19. Understanding Protein Structure: An Academic Guide
Imagine trying to build a fully autonomous, microscopic machine using only a string of twenty different types of snap-together beads. Nature accomplishes this exact engineering feat every single second within living cells. The secret lies not just in the beads themselves, but in how they fold, twist, and chemically lock together to form complex three-dimensional engines. This comprehensive visual guide breaks down the fundamental principles of biochemistry to show exactly how linear chains transform into dynamic cellular tools. The core purpose of this slide deck is to demystify the precise biochemical rules and physical forces that govern the intricate folding and ultimate biological function of these vital macromolecules.
Slide 1: Introduction to Protein Structure and Biochemical Architecture
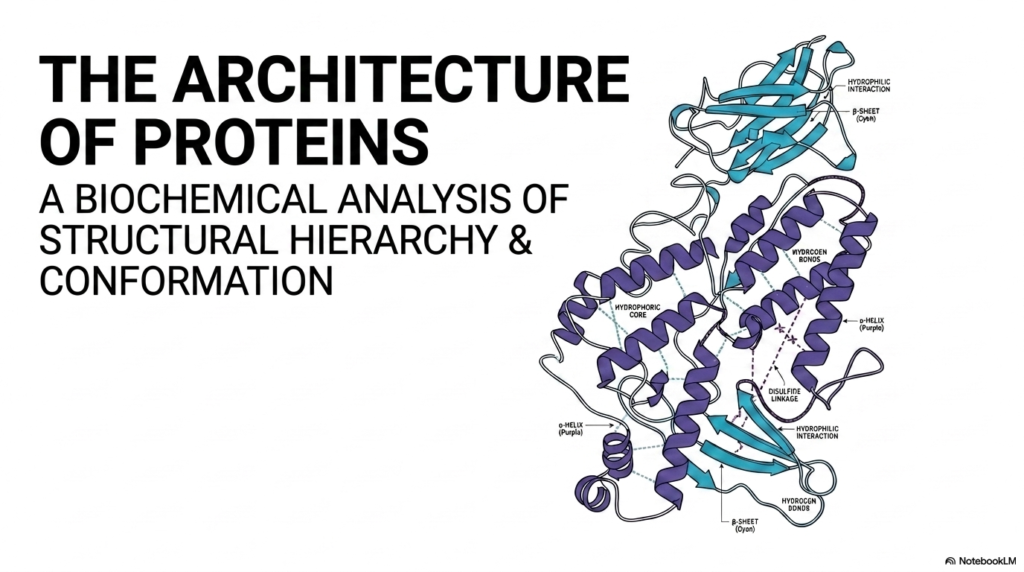
The opening visual provides a beautiful overview of the intricate nature of Protein Structure. It displays a classic ribbon diagram, showcasing the complex folding patterns that characterize a highly functional macromolecule. The fundamental concept highlighted here is that biological utility is entirely dependent on this precise three-dimensional conformation. Without this exact spatial arrangement, biological life as it is currently understood could not exist. Mastering the basics of Protein Structure is the first and most crucial step in unlocking the broader mysteries of molecular biology and cellular mechanics for any serious student.
The diagram illustrates various distinct geometric regions within the folded molecule. Notice the purple helical coils and the cyan arrow-like sheets winding throughout the core. These specific shapes do not form randomly; they are dictated by strict, unchanging biochemical rules. The image also highlights the hydrophobic core, a critical feature of protein structure where water-repelling amino acids naturally cluster in the center. This specific clustering away from the watery cellular environment serves as a major thermodynamic driving force for the entire folding process.
Furthermore, the visual highlights specific stabilizing interactions, such as hydrogen bonds and covalent disulfide linkages. These chemical bonds act exactly like the internal architectural cross-beams in a skyscraper, securely locking the intricate Protein Structure into its final, energetically stable form. Meanwhile, the hydrophilic interactions on the exterior allow the molecule to remain perfectly soluble in the aqueous cellular fluid. Together, these coordinated forces ensure the biological machine remains intact and ready to perform its highly specific cellular tasks.
To fully grasp this overarching concept, one must examine the dynamic interplay between the purple alpha-helices and the cyan beta-sheets shown in the diagram. These specific secondary elements always pack tightly together, minimizing empty internal space while maximizing overall structural integrity. The precise coordination of these moving parts demonstrates that a fully realized Protein Structure is a true marvel of microscopic engineering. This opening slide sets the necessary stage for a much deeper dive into the exact thermodynamic rules governing this incredible biological architecture.
Slide 2: The Four-Tiered Hierarchy of Protein Structure
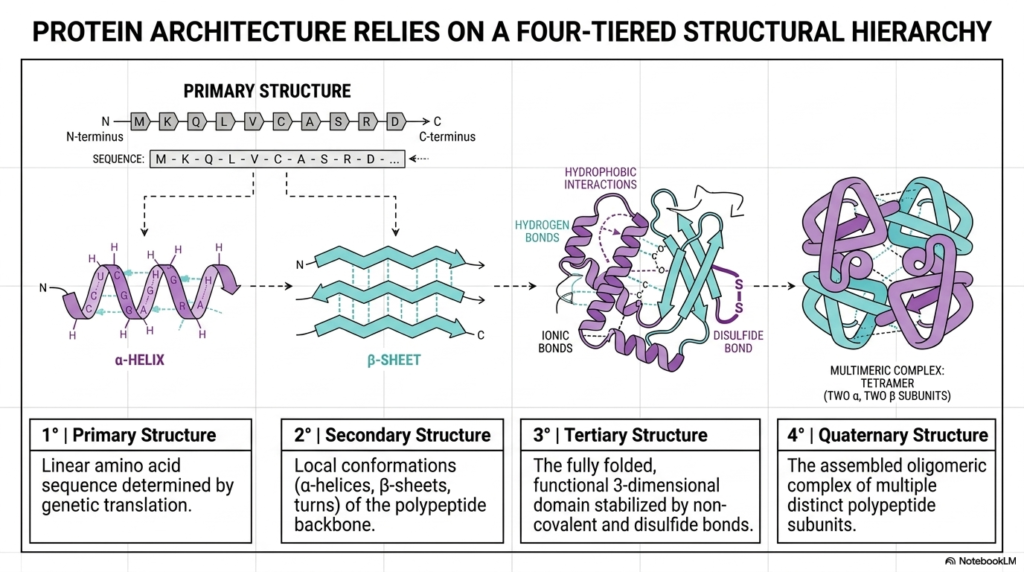
Biological macromolecules do not simply snap into their final shapes instantly. Instead, a functional Protein Structure relies on a highly rigorous, four-tiered structural hierarchy. This slide beautifully illustrates this exact stepwise progression, beginning with the linear amino acid chain at the far left and culminating in a complex multi-subunit assembly on the right. Students must understand that each level of this hierarchy serves as the direct physical foundation for the next. This logical progression is the cornerstone of all biochemical architecture.
The first two tiers form the absolute baseline of molecular design. The primary structure is simply the linear sequence of amino acids, which is directly dictated by genetic translation. This simple string then spontaneously folds into a secondary structure, which consists of local, stable conformations such as alpha-helices and beta-sheets shown in purple and cyan. These initial local folds are crucial because they dictate the geometric boundaries for the rest of the developing Protein Structure.
Moving further along the diagram, the tertiary structure represents the fully folded, functional three-dimensional domain. This specific shape is heavily stabilized by a diverse network of non-covalent interactions and strong disulfide bonds. For many biological molecules, this third tier represents the final, active Protein Structure. However, more complex cellular tasks often require an even higher level of organization, which brings us to the final tier of this molecular hierarchy.
The final tier is the quaternary structure, representing the assembled oligomeric complex of multiple, distinct polypeptide subunits. The multimeric complex shown on the far right, a tetramer consisting of two alpha and two beta subunits, perfectly illustrates this concept. By linking multiple folded chains together, cells can construct massive machinery capable of highly sophisticated cooperative regulation. Every single tier of this architectural hierarchy must be assembled perfectly to ensure cellular viability.
Slide 3: Primary Sequences Dictate Final Protein Structure
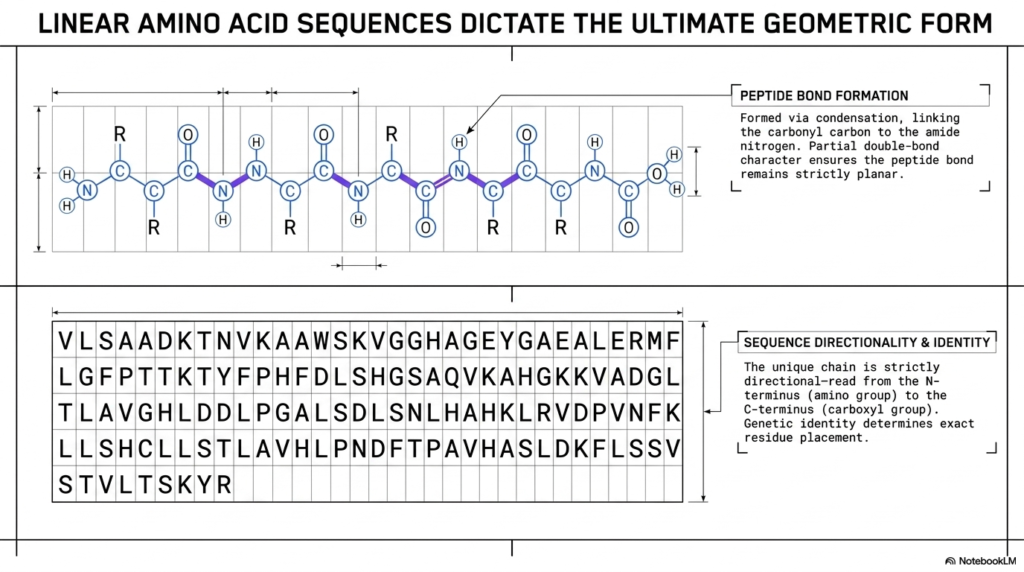
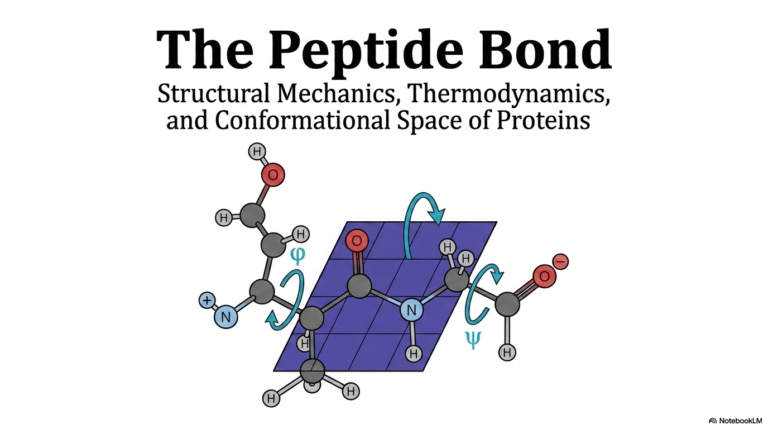
The linear sequence of amino acids is far more than a simple list of molecules; it is the ultimate blueprint. The final, complex Protein Structure is entirely encoded within this specific sequential arrangement. The top panel illustrates exactly how individual amino acids are chemically linked together via condensation reactions. This crucial process links the carbonyl carbon of one residue directly to the amide nitrogen of the next, forming the foundational backbone of the entire biological molecule.
A critical biochemical feature highlighted here is the peptide bond itself. Because this specific linkage possesses partial double-bond character, it remains strictly planar and highly rigid. This inherent rigidity severely restricts the main chain’s flexibility, a key physical constraint on the overall Protein Structure. The molecule cannot simply bend in any arbitrary direction; it must strictly obey the geometric limitations imposed by these planar peptide bonds along the structural backbone.
The bottom panel emphasizes the vital importance of sequence directionality and unique identity. The sequence of letters represents specific amino acid residues, and this unique chain is always read directionally from the N-terminus to the C-terminus. The exact placement of each specific residue, directly determined by genetic identity, dictates every subsequent folding event. Any mutation altering this unique sequence can catastrophically disrupt the entire final Protein Structure.
Understanding this linear blueprint is absolutely essential for medical students studying genetic diseases. When the genetic code introduces a single incorrect amino acid into this specific linear sequence, the entire downstream structural hierarchy can completely unravel. The highly specific sequence of residues shown in the grid demonstrates exactly how much chemical information is securely stored within the most basic primary tier of molecular design.
Slide 4: Ramachandran Plots and the Limits of Protein Structure
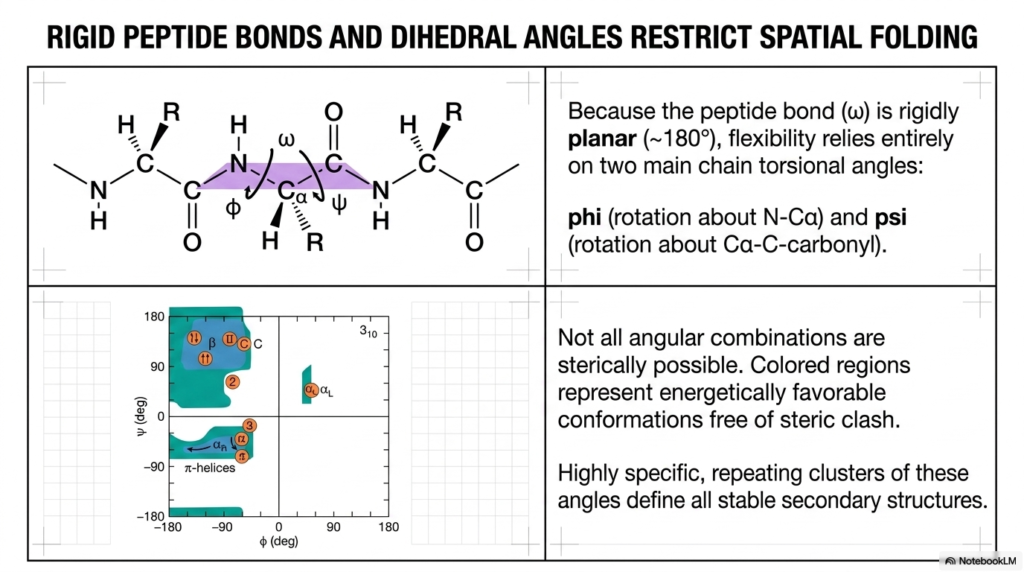
Because the central peptide bond is rigidly planar, the molecule’s flexibility relies entirely on two specific main-chain torsional angles. This slide introduces phi and psi, the angles of rotation around the alpha-carbon that dictate spatial folding. The fundamental concept here is that not all angular combinations are sterically possible. Understanding these specific rotational limits is vital for comprehending how a functional Protein Structure physically navigates the complex transition from a linear string into a compact three-dimensional shape.
The Ramachandran plot shown on the bottom left is a foundational tool in structural biochemistry. It graphically maps all energetically favorable spatial conformations that are entirely free of molecular steric clashes. The colored regions represent the only allowable angular combinations in which atoms do not physically collide. This geometric mapping proves that a stable Protein Structure is heavily constrained by fundamental physical laws and chemical thermodynamics.
Notice how the specific repeating clusters of these allowable angles perfectly define all stable secondary structures. The areas marked for alpha-helices and beta-sheets align exactly with the most energetically favorable regions on the entire plot. If the torsional angles stray outside these highly specific colored zones, the resulting steric hindrance would instantly destabilize the overall Protein Structure. Nature fundamentally prefers energy minimization above all else.
By using these energetically favorable angles, polypeptides reliably fold into predictable, highly stable local conformations. The Ramachandran plot elegantly visually proves that protein folding is not a random, chaotic search through endless spatial possibilities. Instead, it is a highly directed, thermodynamically driven journey toward the lowest possible energy state, ensuring the reliable and consistent construction of essential cellular machinery.
Slide 5: The Alpha-Helix in Protein Structure
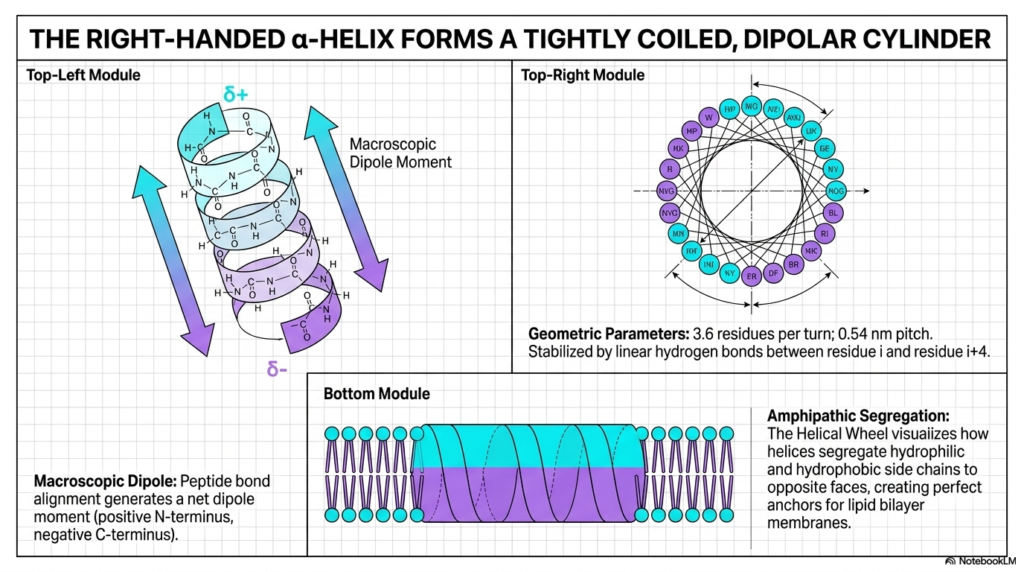
The right-handed alpha-helix is one of the most recognizable and ubiquitous features of cellular architecture. As shown in the top-left module, it forms a tightly coiled, dipolar cylinder that securely stabilizes the local backbone. This crucial structural element is a major pillar of Protein Structure, widely used by nature to build robust, highly functional molecular domains. The specific geometry of this coil is mathematically precise, featuring exactly 3.6 residues per turn.
A key chemical feature of the alpha-helix is its macroscopic dipole moment. Because all the individual peptide bonds perfectly align in the same vertical direction, they generate a net positive charge at the N-terminus and a net negative charge at the C-terminus. This distinct polarity allows the localized Protein Structure to interact very specifically with other charged molecules and critical functional domains. It also heavily relies on linear hydrogen bonds between residue i and i+4 for profound structural stabilization.
The helical wheel diagram in the top-right module brilliantly visualizes the concept of amphipathic segregation. By looking straight down the central barrel of the helix, students can clearly see how hydrophilic and hydrophobic side chains deliberately segregate to completely opposite faces. This specific chemical segregation allows the overall Protein Structure to exhibit two distinct chemical environments simultaneously. This dual nature is absolutely essential for complex cellular functions.
The bottom module perfectly illustrates the practical biological utility of this amphipathic segregation. Because one distinct face is entirely hydrophobic, it acts as a perfect molecular anchor, seamlessly integrating into the oily interior of a lipid bilayer membrane. Meanwhile, the opposite hydrophilic face safely interacts with the watery exterior environment. This elegant biochemical design allows these structural cylinders to safely bridge the crucial gap between cellular compartments.
Slide 6: Beta-Sheets and Their Role in Protein Structure
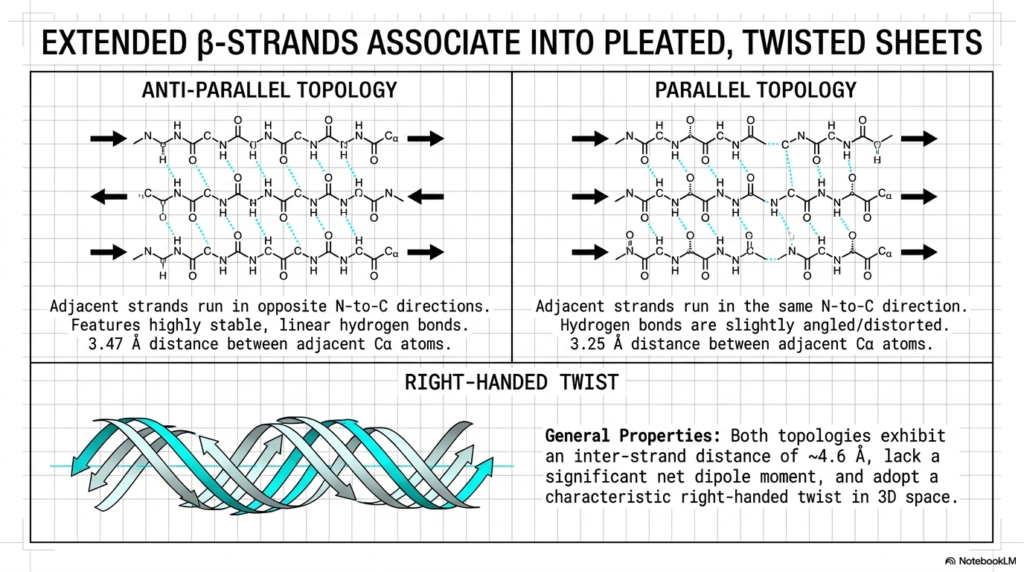
Alongside the helical coil, extended beta-strands represent the second major foundational pillar of molecular architecture. These individual strands reliably associate to form broad, pleated, and twisted sheets, as elegantly illustrated in this slide. These extended sheets play an absolutely vital role in establishing the core stability of a complex Protein Structure. Unlike the tightly wound helix, these structures stretch the polypeptide backbone into a much more extended, linear conformation.
The diagram brilliantly details the two primary topological arrangements: anti-parallel and parallel. In the anti-parallel topology on the left, the adjacent strands run in strictly opposite N-to-C directions. This specific opposing arrangement enables highly stable, perfectly linear hydrogen bonds to form rapidly between adjacent backbones. This strong, linear bonding pattern makes the antiparallel protein structure incredibly rigid and mechanically resilient under applied cellular stress.
Conversely, the parallel topology on the right features adjacent strands running in the exact same direction. Because of this parallel alignment, the bridging hydrogen bonds must become slightly angled or heavily distorted to connect properly. While slightly less thermodynamically stable than their anti-parallel counterparts, these parallel sheets still form vital structural components within a broader Protein Structure. Both specific topologies exhibit a distinct inter-strand distance that maximizes hydrogen bonding efficiency.
Finally, the bottom illustration highlights the characteristic right-handed twist adopted by these structures in three-dimensional space. These sheets rarely remain perfectly flat in biological environments; they naturally twist to relieve inherent steric strain and optimize their thermodynamic packing. This specific twisting motion is a ubiquitous feature of molecular architecture, allowing these broad, pleated sheets to smoothly form closed barrels and highly complex active sites within the folded macromolecule.
Slide 7: Turns and Loops Shaping Protein Structure
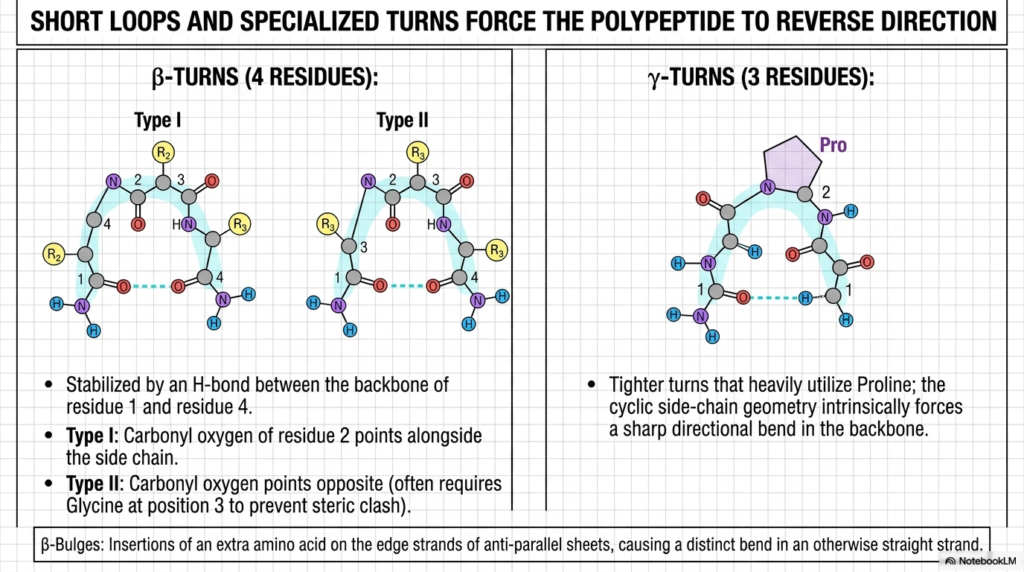
While helices and sheets form the rigid structural framework, short loops and specialized turns are equally important for overall compaction. These specific turning elements forcefully compel the linear polypeptide backbone to rapidly reverse its spatial direction. Without these essential directional changes, a functional Protein Structure would simply be an endlessly long, completely useless linear string. These tight turns are the exact mechanisms that allow macromolecules to fold into dense, highly compact spherical shapes.
The slide elegantly details two major categories of directional reversal: the beta-turn and the tighter gamma-turn. Beta-turns explicitly require exactly four distinct residues to complete the required geometric U-turn. As shown in the diagram, these specific turns are heavily stabilized by a crucial internal hydrogen bond between the backbone of residue one and residue four. This localized bonding firmly locks the bend into the broader Protein Structure, preventing it from unraveling in the cellular environment.
The differences between Type I and Type II beta-turns highlight the extreme sensitivity of molecular folding. In Type II turns, the carbonyl oxygen points in the exact opposite direction, which almost always requires a tiny Glycine residue at the third position to successfully prevent a massive steric clash. This incredibly specific spatial requirement proves that even the smallest, tightest corners of a Protein Structure are entirely dependent on highly exact amino acid sequences.
The gamma-turn on the right showcases an even tighter directional reversal, utilizing only three specific residues. This incredibly sharp bend heavily utilizes Proline, whose unique cyclic side-chain geometry intrinsically forces a harsh, unavoidable directional kink directly into the main backbone. By strategically placing specific residues like Proline and Glycine, nature can expertly and precisely control the spatial trajectory of the entire molecular chain.
Slide 8: Amino Acid Preferences Guiding Protein Structure
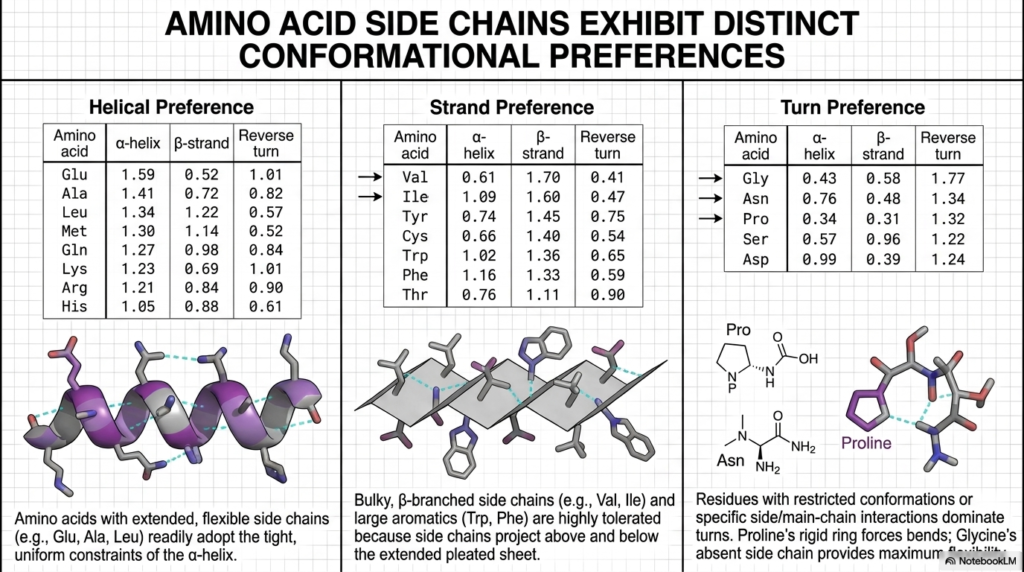
Not all amino acids are structurally equal; each exhibits distinct and measurable conformational preferences. This particular slide explores how the unique chemical side chains of individual residues directly influence local folding patterns. Understanding these specific spatial biases is absolutely crucial for predicting how a primary sequence will eventually transition into a functional Protein Structure. These statistical preferences are not random; they are entirely rooted in fundamental thermodynamics and steric physics.
The table on the far left reveals that amino acids with extended, highly flexible side chains readily adopt the tight constraints of the alpha-helix. Residues like Glutamate, Alanine, and Leucine score very highly for helical preference because they can easily project outward without physically interfering with the tightly coiled backbone. This natural chemical bias ensures that specific regions of the growing Protein Structure spontaneously form essential helical cylinders during the early stages of thermodynamic folding.
Conversely, the middle panel demonstrates that bulky, highly branched side chains show a massive statistical preference for the extended beta-strand. Large aromatic residues like Tryptophan and Phenylalanine are highly tolerated here because their massive side chains can project freely above and below the extended, pleated sheet. This specific geometric freedom prevents the disastrous steric clashing that would instantly destabilize a more tightly packed Protein Structure, favoring the formation of broad, flat molecular surfaces.
The panel on the right highlights the absolute dominance of Glycine and Proline in turn preferences. Proline’s incredibly rigid, cyclic ring forces an unavoidable bend, while Glycine’s total lack of a bulky side chain provides maximum spatial flexibility. These two specific residues act as the crucial molecular hinges of the biological machine. Their highly strategic placement reliably forces the required sharp directional reversals necessary to achieve a fully compact, native state.
Slide 9: Alpha-Keratin: A Fibrous Protein Structure
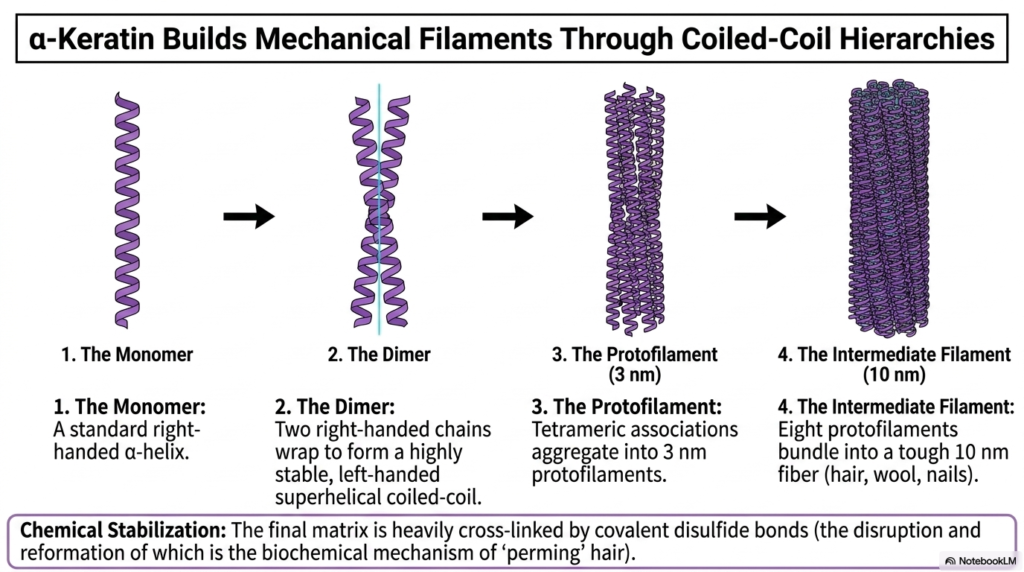
Alpha-keratin perfectly illustrates how nature utilizes repeating secondary elements to construct massive, highly durable mechanical filaments. This slide beautifully breaks down the precise structural evolution from a single monomer entirely up to a tough biological fiber. For structures like hair, wool, and nails, the primary biological objective is sheer mechanical strength rather than complex enzymatic activity. Therefore, the resulting Protein Structure must be rigorously optimized for extreme physical durability and high tensile resistance.
The assembly process begins with a standard, right-handed alpha-helical monomer. However, step two reveals the brilliant biochemical trick: two of these right-handed chains tightly wrap around each other to successfully form a left-handed superhelical coiled-coil. This opposing directional twisting acts exactly like the twisting of individual fibers in a heavy structural rope, massively amplifying the physical strength of the localized Protein Structure and actively preventing the distinct helices from simply unspooling under stress.
As the structural hierarchy rapidly advances, these stable dimeric coiled-coils aggregate into larger tetrameric associations known as three-nanometer protofilaments. These intermediate structures then heavily bundle together, perfectly aligning to form the ultimate ten-nanometer intermediate filament. This highly complex, multi-tiered assembly process demonstrates how a relatively simple Protein Structure can be extensively multiplied and structurally bundled to achieve remarkable macroscale mechanical toughness in the host organism.
Finally, the slide importantly notes the crucial role of chemical stabilization via strong covalent disulfide bonds. The fully assembled biological matrix is heavily cross-linked by these incredibly strong bonds, which permanently lock the thick fibers tightly together. The disruption and subsequent artificial reformation of these specific chemical cross-links is the exact biochemical mechanism behind ‘perming’ human hair, providing a perfectly relatable, real-world application of advanced structural biochemistry.
Slide 10: The Triple Helix of Collagen’s Protein Structure
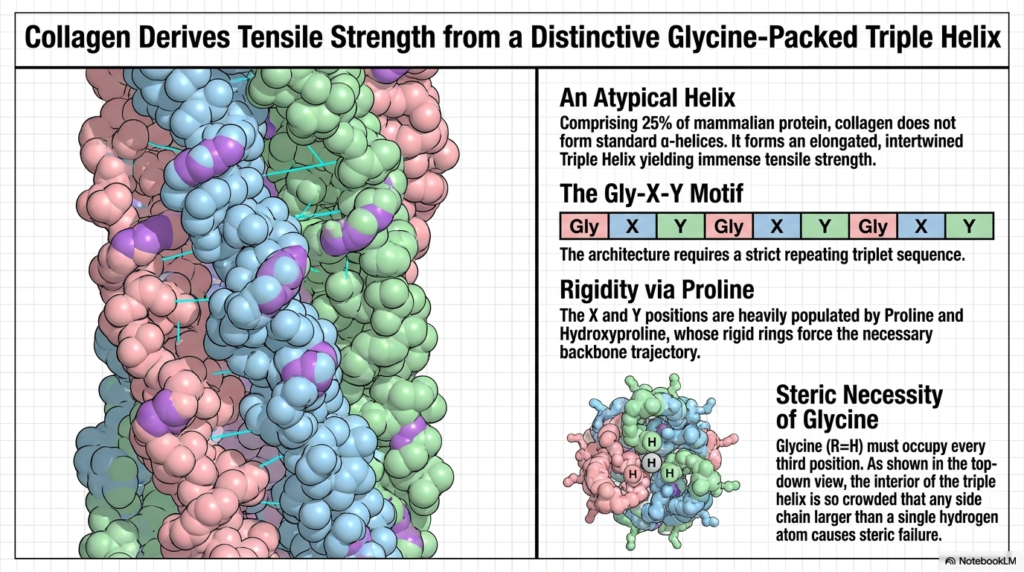
Collagen, representing a massive 25% of all mammalian protein, requires a completely unique structural approach to achieve its required immense tensile strength. Unlike the standard cylinders discussed previously, collagen does not utilize traditional alpha-helices. Instead, its specific Protein Structure is defined by a highly atypical, elongated, and extensively intertwined Triple Helix. This unique molecular architecture is absolutely essential for physically supporting the vast connective tissues, tendons, and bones throughout the entire human body.
The fundamental architectural requirement for this unique structure is the strict, relentlessly repeating Gly-X-Y triplet motif. This specific primary sequence is completely non-negotiable; without this exact repeating pattern, the robust Triple Helix simply cannot form. This proves yet again how uniquely tied the final, functional Protein Structure is to its initial genetic blueprint. Every single geometric feature of this massive cable is entirely dependent on this distinct, highly repetitive genetic code.
The rigidity of this massive cable is primarily conferred by the high content of Proline and Hydroxyproline at the X and Y positions. Their uniquely rigid cyclic rings forcefully dictate the necessary, highly extended spatial trajectory of the main backbone. Furthermore, the far-right panel highlights the absolute steric necessity of Glycine. Because Glycine has only a single, tiny hydrogen atom as a side chain, it is the only residue small enough to physically fit.
As shown in the detailed top-down view, the internal core of this specific Protein Structure is incredibly crowded. If any side chain larger than Glycine attempts to occupy every third central position, it immediately causes catastrophic steric failure, actively unraveling the massive triple helix. This incredible evolutionary design perfectly demonstrates how sheer tensile strength is ultimately achieved through extremely tight, highly optimized internal spatial packing and rigorous chemical precision.
Slide 11: Silk Fibroin and Beta-Sheet Protein Structure
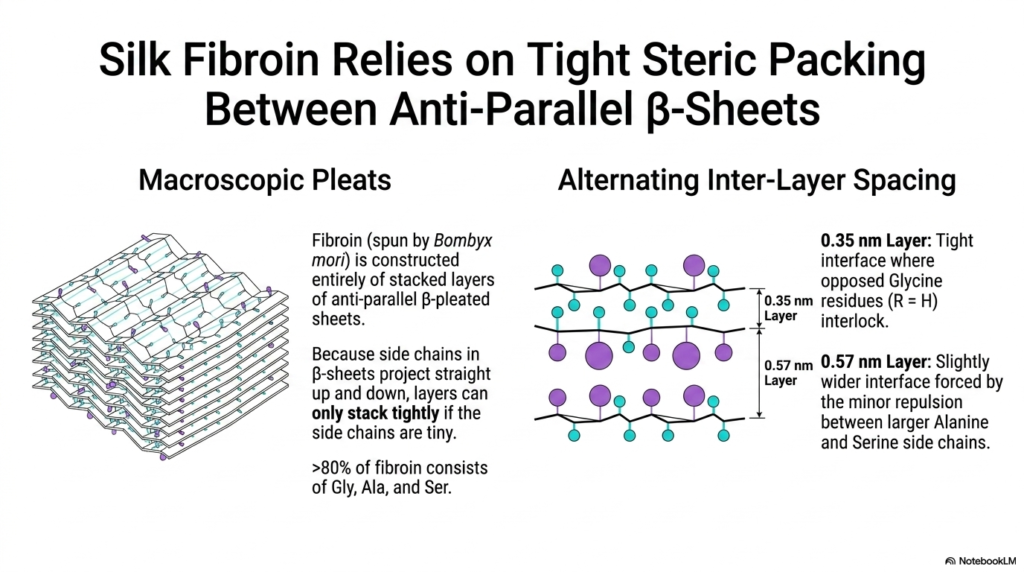
Silk fibroin, the highly resilient material spun by the Bombyx mori silkworm, offers a fascinating look into the macroscopic capabilities of extended molecular sheets. This slide explicitly demonstrates how a biological material can be constructed entirely from massive, extensively stacked layers of antiparallel beta-pleated sheets. This highly specialized Protein Structure eschews bulky helices entirely, relying completely on rigorous, dense geometric packing to achieve its famous combination of extreme tensile strength and smooth flexibility.
The macroscopic pleats shown on the left rely heavily on an incredibly tight molecular interface. Because the chemical side chains in these extended sheets always project straight up and down, the distinct broad layers can stack tightly together only if those side chains are incredibly tiny. Therefore, the entire Protein Structure of this material is deliberately optimized by evolution to utilize only the smallest possible amino acid residues.
Over 80% of the entire silk fibroin sequence consists entirely of Glycine, Alanine, and Serine. The diagram on the right beautifully illustrates how this specific composition perfectly creates alternating inter-layer spacing. The incredibly tight 0.35-nanometer layer features opposing Glycine residues that perfectly interlock. This microscopic interlocking mechanism securely binds the extended Protein Structure together, preventing the broad sheets from sliding completely apart under heavy mechanical pulling force.
The slightly wider 0.57-nanometer layer accommodates the minor spatial repulsion imposed by the slightly larger Alanine and Serine side chains. This brilliant alternating architectural design allows the massive molecular sheets to pack incredibly densely while still retaining a slight degree of crucial microscopic flexibility. Understanding this highly specific geometric arrangement is essential for advanced students studying the remarkable mechanical properties of specialized biological fibers and biomaterials.
Slide 12: Super-Secondary Motifs in Protein Structure
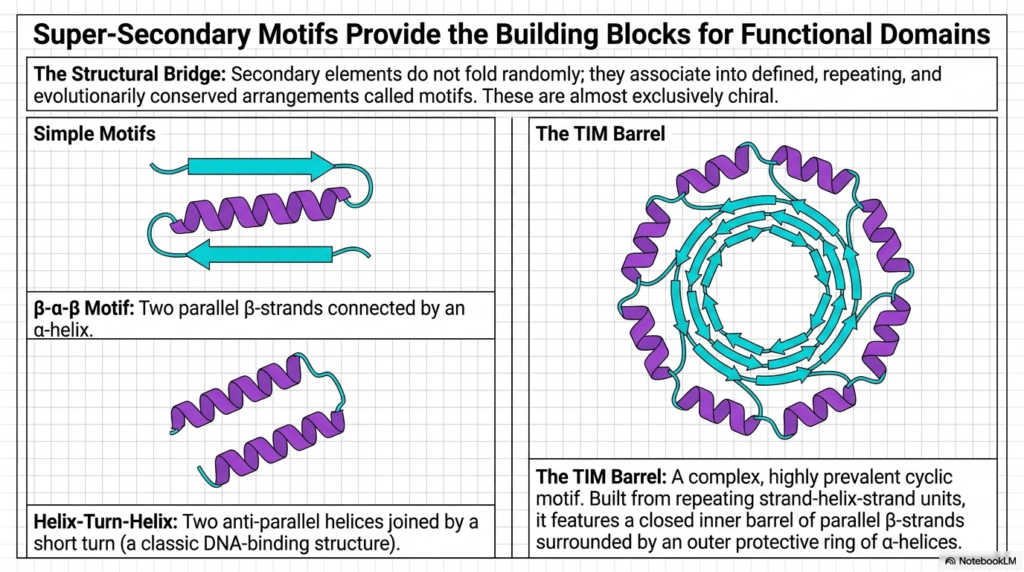
Secondary structural elements do not just fold randomly within the cellular environment; they deliberately associate into highly defined, repeating, and evolutionarily conserved spatial arrangements. These incredibly specific arrangements are known as super-secondary motifs. This slide highlights how these simple structural bridges actively serve as the foundational building blocks for much larger, highly functional biological domains. Recognizing these distinct repeating geometric patterns is incredibly crucial for properly decoding any complex, newly discovered Protein Structure.
The panel on the left illustrates two incredibly common, simple motifs. The beta-alpha-beta motif elegantly connects two parallel beta-strands using an intervening structural alpha-helix. Similarly, the helix-turn-helix motif securely joins two anti-parallel helical cylinders together using a very short, highly specific directional turn. These seemingly simple geometric combinations are ubiquitous in nature and are widely used to reliably construct the highly stable internal framework of a much larger functional Protein Structure.
The right panel introduces the incredibly complex and highly prevalent TIM Barrel. Built entirely from seamlessly repeating strand-helix-strand units, it perfectly features a completely closed inner barrel of robust parallel beta-strands, safely surrounded by an outer protective ring of sturdy alpha-helices. This beautiful cyclic architecture represents a pinnacle of structural evolution, providing a highly stable, completely enclosed internal chemical pocket perfectly suited for highly specific enzymatic catalysis.
These super-secondary motifs physically demonstrate that nature heavily relies on a highly modular design strategy. By repeatedly utilizing these proven, highly stable geometric building blocks, cells can efficiently construct an incredibly vast array of diverse molecular machinery. Understanding how these distinct motifs seamlessly assemble together provides deep, critical insights into the incredible evolutionary history and functional biochemical mechanisms hidden within any large-scale Protein Structure.
Slide 13: Non-Covalent Forces Driving Tertiary Protein Structure
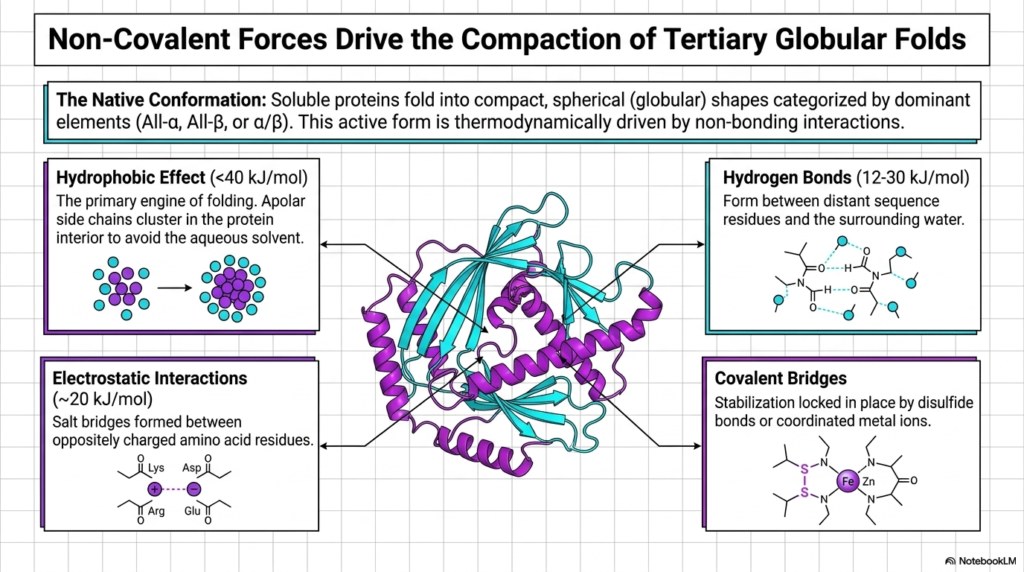
Once the local motifs are formed, a highly diverse array of chemical forces drives the final compaction into the fully functional, tertiary globular fold. Soluble macromolecules must reliably fold into these highly compact, spherical shapes to function properly within the active cellular environment. This specific active form is entirely thermodynamically driven by crucial non-bonding interactions. This complex slide perfectly details the exact specific physical engines that reliably power the final, critical stages of Protein Structure assembly.
The absolute primary engine of this massive folding process is the hydrophobic effect. As shown in the top-left panel, apolar, water-repelling side chains rapidly cluster deeply within the internal protein interior to violently avoid the surrounding aqueous solvent. This massive, thermodynamically favorable clustering firmly establishes the dense core of the entire Protein Structure. It provides the massive bulk stabilization energy absolutely required to physically overcome the vast conformational entropy of the highly flexible linear backbone.
Simultaneously, crucial hydrogen bonds and highly specific electrostatic interactions actively stabilize the folding exterior. Hydrogen bonds form rapidly between distant residues in the sequence, while electrostatic salt bridges firmly lock oppositely charged amino acids together. Although individually incredibly weak, the massive, combined sum of these numerous non-covalent interactions provides immense thermodynamic stability to the overall active Protein Structure, ensuring it safely maintains its specific required geometry under shifting cellular conditions.
Finally, the slide highlights the vital role of specific covalent bridges, such as strong disulfide bonds and heavily coordinated metal ions like Zinc and Iron. These extremely strong, permanent chemical linkages act as rigid molecular staples, permanently locking the highly flexible final conformation safely into its required native state. Without these crucial stabilizing forces acting in perfect chemical unison, the entire functional Protein Structure would rapidly denature and catastrophically fail entirely.
Slide 14: Quaternary Assembly and Complex Protein Structure
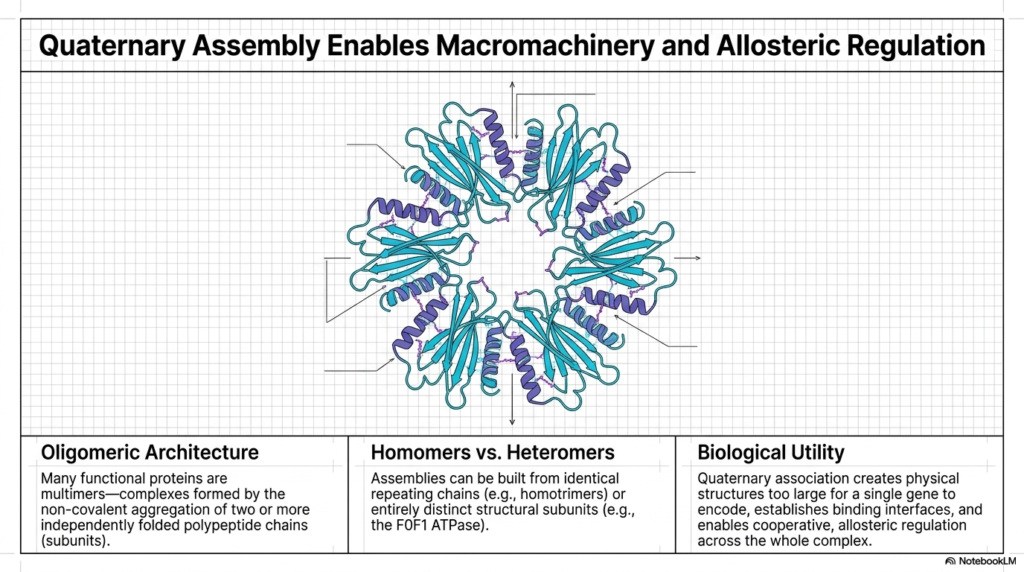
While many molecules function perfectly as single, isolated chains, nature often demands significantly more complex macromachinery. This slide elegantly introduces quaternary assembly, the final crucial tier of structural organization. Many incredibly essential functional proteins are actually massive multimers—complexes formed by the highly specific, non-covalent aggregation of two or more completely independently folded polypeptide subunits. This advanced assembly process creates a truly massive Protein Structure capable of highly sophisticated cooperative regulation.
The beautiful central diagram perfectly illustrates a highly complex oligomeric architecture. The distinct individual colored subunits safely assemble together to create a massive physical structure, which is heavily required for advanced biological tasks. These distinct assemblies can be readily built from identical, endlessly repeating homomeric chains or entirely distinct, highly specialized heteromeric subunits. This incredible modular approach enables the highly efficient biological construction of a massive Protein Structure using relatively small genetic blueprints.
The biological utility of this high-level quaternary assembly is absolutely immense. First, it reliably creates massive physical structures that are simply far too large for any single gene to encode safely and efficiently. By linking smaller, pre-folded subunits tightly together, the cell vastly reduces the risk of disastrous translational errors while simultaneously constructing an incredibly large, highly complex Protein Structure capable of spanning vast cellular distances or entirely breaching robust, thick lipid membranes.
Furthermore, this multi-subunit architecture directly enables advanced allosteric regulation across the entire massive complex. When one distinct subunit physically binds a target ligand, it can instantly mechanically transmit that specific conformational change directly across the binding interfaces to safely activate or cleanly inhibit all the other connected subunits simultaneously. This incredibly sophisticated structural communication is exactly what makes advanced, multi-part Protein Structure the ultimate highly responsive machinery of biological life.
Slide 15: Insulin Maturation and Hierarchical Protein Structure
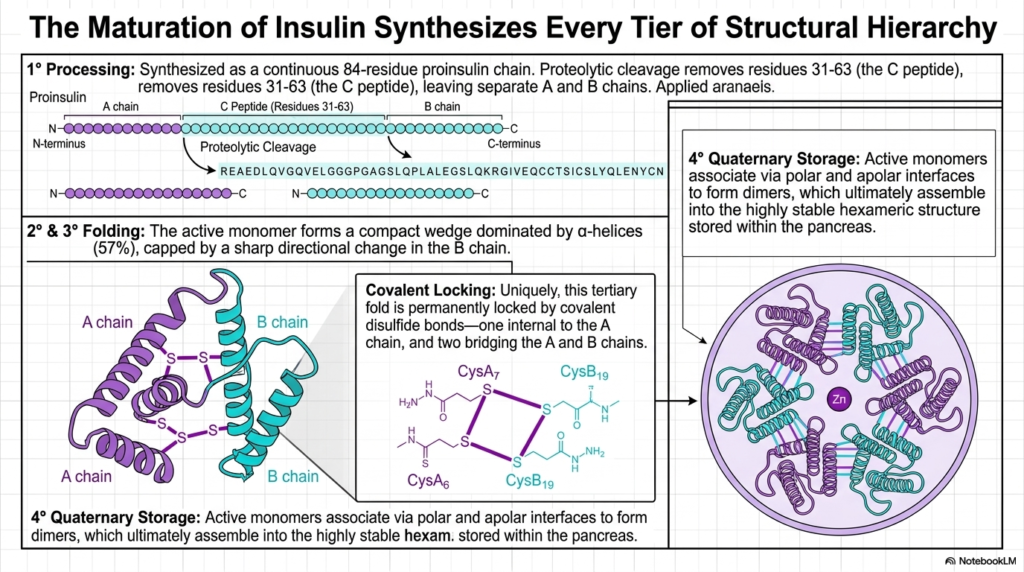
The complex biochemical maturation of the hormone insulin serves as a perfect, real-world summary, seamlessly synthesizing every single tier of the structural hierarchy discussed throughout this visual guide. The process absolutely begins with primary processing. Insulin is initially synthesized as a continuous 84-residue linear proinsulin chain. This highly specific primary sequence is the starting point for the final Protein Structure, proving once again that the linear blueprint contains all necessary downstream folding instructions.
Next, a crucial proteolytic cleavage site surgically removes residues 31 through 63, thereby completely isolating the distinct A and B chains. Following this, the active monomers rapidly fold into a highly compact tertiary wedge dominated by alpha-helical structure. This specific compact Protein Structure is permanently locked into its required active configuration by three extremely strong covalent disulfide bonds. One bond is entirely internal to the A chain, while two others securely bridge the A and B chains.
The final, fully functional tertiary fold is a direct result of these tightly coordinated helical motifs and rigid covalent locks. However, the true genius of insulin’s design lies in its incredible quaternary storage capability. As shown in the final panel, the highly active monomers reliably associate via highly specific polar and apolar chemical interfaces to successfully form secure dimeric pairs. This critical multimeric assembly is a hallmark of advanced, highly regulated Protein Structure.
Ultimately, these secure dimers reliably assemble into a massive, highly stable hexameric ring, tightly coordinated around central Zinc ions. This beautiful quaternary hexamer is the exact structural form safely stored deeply within the human pancreas. When the body urgently requires the active hormone, this massive hexameric Protein Structure rapidly and safely dissociates directly back into highly active monomers, perfectly demonstrating the incredible dynamic functional power of multi-tiered structural biological architecture.
Please read our Content Disclaimer Statement.
Check out our social media channels: