13. Phospholipids: Molecular Structures and Cell Membrane Functions
If you have ever wondered how a cell prevents itself from turning into a disorganized, watery soup, you are in the right place. Our cells are essentially complex bags of water, and maintaining the strict boundaries between the “inside” and the “outside” is the defining feature of biological life. The core purpose of this slide deck, titled “Structural and Functional Architecture of Compound Lipids,” is to explore exactly how nature constructs these smart barriers. We will dive deep into the molecular architecture, exploring the diverse backbones, headgroups, and functional domains that make cellular membranes the ultimate dynamic interface. Let’s get started!
Slide 1: Introduction to Structural Phospholipids and the Membrane Architecture
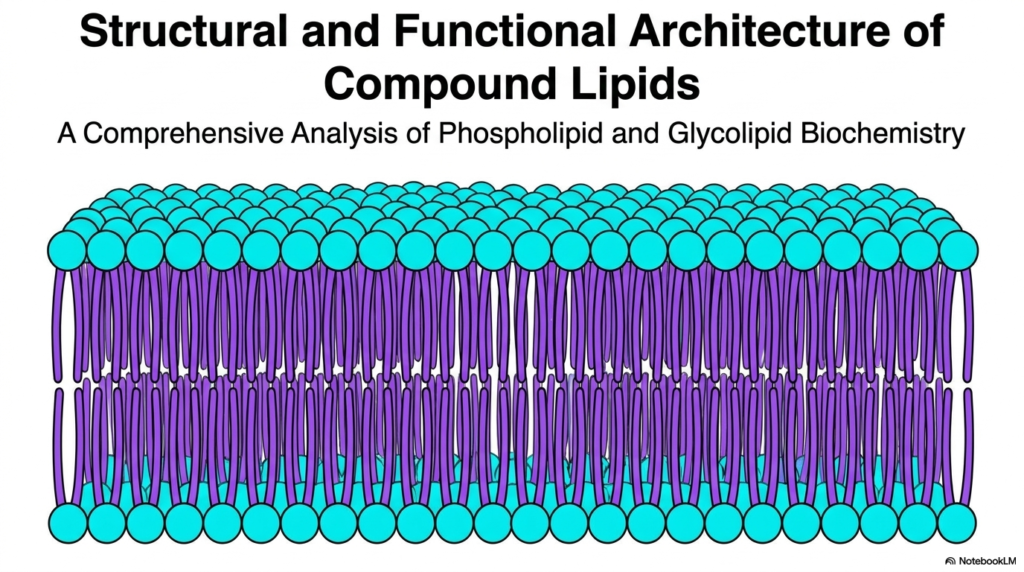
When you look at this first slide, you are looking at the fundamental canvas of all biological life: the lipid bilayer. For decades, biology textbooks simplified the cellular membrane as just a passive, fatty wrapping paper. However, as modern biochemistry has revealed, this structure is a highly dynamic, intelligent barrier primarily constructed by Phospholipids. Unlike the triacylglycerols or storage fats that your body uses for long-term energy reserves, the molecules depicted here are structural powerhouses. They are explicitly designed to form sheets rather than oil droplets.
What you see here is a vast, continuous ocean of Phospholipids arranging themselves in a dual-layered formation. The little blue spheres represent the water-loving polar headgroups, while the purple squiggly lines represent the water-fearing lipid tails. Because biological systems are aqueous—meaning both the inside of the cell (cytoplasm) and the outside environment are water-based—these Phospholipids spontaneously self-assemble to protect their tails from the water. This spontaneous assembly requires absolutely no enzymatic energy; it is driven entirely by thermodynamics and the hydrophobic effect.
Understanding this basic architecture is the first step in mastering cell biology. Every single drug you will ever prescribe, every nutrient your cells absorb, and every nerve impulse your brain fires must negotiate with this tightly packed barrier of Phospholipids. This architectural foundation dictates membrane fluidity, permeability, and even the ability of the cell to divide. As we move through this deck, we will unpack exactly how varying the chemical structure of these molecules completely changes the functional destiny of the membrane.
Slide 2: The Amphipathic Architecture of Phospholipids
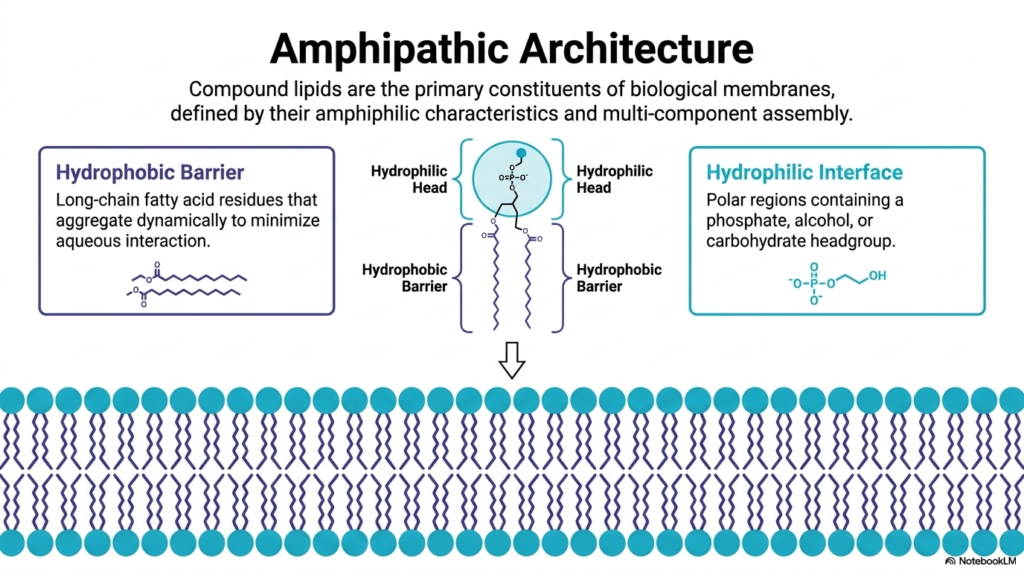
Building upon our introduction, this slide takes a closer look at the dual personality of membrane lipids. The defining biochemical characteristic of these structural molecules, particularly the Phospholipids, is their “amphipathic” or amphiphilic nature. This means they possess two distinct, contradictory chemical properties within the exact same molecule.
On one side, we have the “Hydrophobic Barrier.” This consists of long-chain fatty acid residues—hydrocarbon chains that are completely nonpolar. Because they cannot form hydrogen bonds with water, they aggregate dynamically to minimize aqueous interaction. In the diagram, these are the zigzagging tails pointing inward. On the opposite side, we have the “Hydrophilic Interface.” This is the polar region containing a negatively charged phosphate group, along with an attached alcohol or carbohydrate headgroup.
Why is this amphipathic split so critical? Because it allows Phospholipids to act as the ultimate biological mediators. The polar heads happily interface with the watery environments of the extracellular fluid and the cytosol, engaging in hydrogen bonding and electrostatic interactions. Meanwhile, the hydrophobic tails associate via Van der Waals forces, creating an oily, impermeable core. This core prevents the free diffusion of ions, polar molecules, and large proteins. Without the specific amphipathic architecture of these Phospholipids, your cells would constantly leak their internal contents, making life impossible. This structural duality is the exact reason why cells can maintain complex internal environments perfectly distinct from the chaos of the outside world.
Slide 3: The Biochemical Backbone Framework of Phospholipids and Sphingolipids
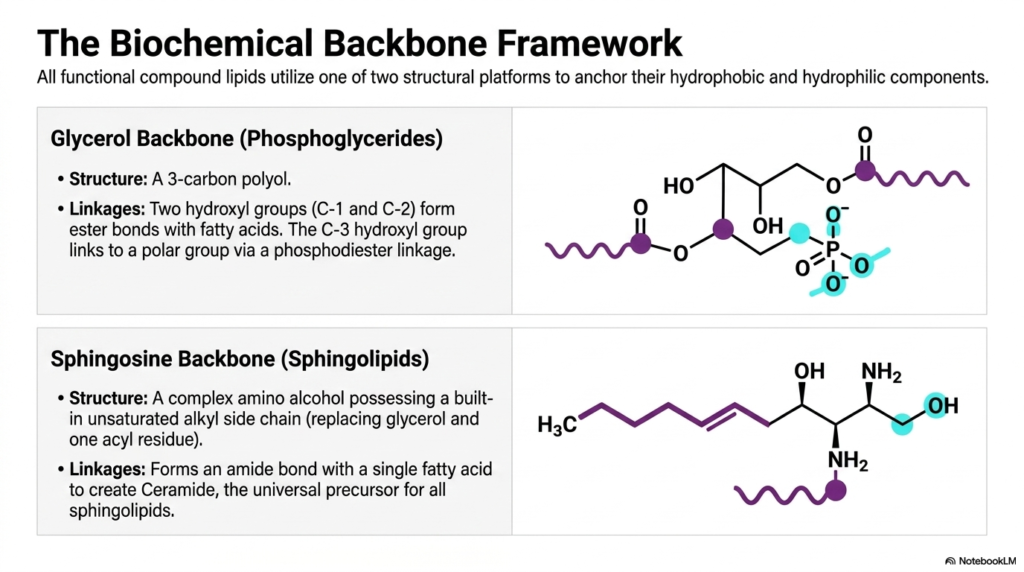
If you want to understand how a skyscraper is built, you first look at its steel frame. The same logic applies to compound lipids. Every functional lipid in your cellular membranes requires a molecular platform—a biochemical backbone—to anchor the hydrophobic tails and the hydrophilic heads together. This slide introduces the two primary platforms used by nature: the glycerol backbone and the sphingosine backbone.
The top panel displays the glycerol backbone, the foundational structure for most Phospholipids in the human body. Glycerol is a simple 3-carbon polyol. In phosphoglycerides, the first two hydroxyl groups (at C-1 and C-2) form ester bonds with long-chain fatty acids. This forms the hydrophobic dual-tail. The crucial third hydroxyl group at C-3 does not attach to a fat; instead, it forms a phosphodiester linkage to a polar group. This specific linkage is what grants these Phospholipids their structural versatility and amphipathic character.
The bottom panel introduces a completely different architectural strategy: the sphingosine backbone. Instead of a simple 3-carbon alcohol, sphingosine is a complex, long-chain amino alcohol that already possesses a built-in unsaturated hydrocarbon chain. Because of this, it only needs to attach one additional fatty acid via an amide bond to create “Ceramide,” the universal precursor for all sphingolipids. While many sphingolipids also incorporate a phosphate group—making them a specialized subtype of Phospholipids—their unique amino alcohol backbone provides different chemical stabilities and packing geometries, which are heavily utilized in the protective membranes of your nervous system.
Slide 4: Major Glycerophospholipids and the Electrical Charge of Phospholipids
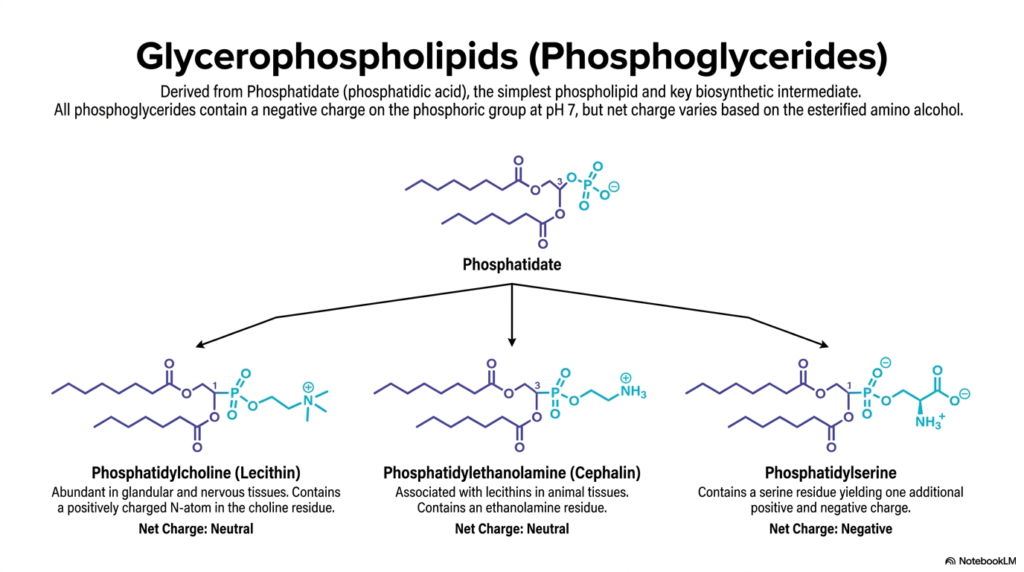
Now that we understand the glycerol backbone, we can explore the specific molecules that make up the bulk of your cellular membranes. This slide details the most common glycerophospholipids, all of which are derived from a parent molecule called Phosphatidate (or phosphatidic acid). Phosphatidate is the absolute simplest of the Phospholipids, serving as the essential biosynthetic intermediate for all the more complex lipids shown here.
The chemical identity—and more importantly, the net electrical charge—of these Phospholipids depends entirely on the specific amino alcohol esterified to the phosphate group. On the left, we have Phosphatidylcholine (Lecithin). It features a bulky choline headgroup containing a positively charged nitrogen atom. Because this positive charge perfectly balances the negative charge of the phosphate, Phosphatidylcholine has a net neutral charge at physiological pH. Due to its cylindrical shape, it is the most abundant lipid forming the flat structural planes of your membranes.
In the middle is Phosphatidylethanolamine (Cephalin), which also carries a net neutral charge but features a smaller ethanolamine headgroup. This smaller head gives the lipid a cone-like shape, making it crucial for areas of the membrane that need to curve or bend. Finally, on the right, we have Phosphatidylserine. By attaching the amino acid serine, this lipid gains an extra carboxyl group, resulting in a distinct net negative charge. In healthy cells, these negatively charged Phospholipids are strictly confined to the inner leaflet of the membrane, acting as a crucial docking site for intracellular signaling proteins.
Slide 5: The Role of Phospholipids in Cellular Signal Transduction
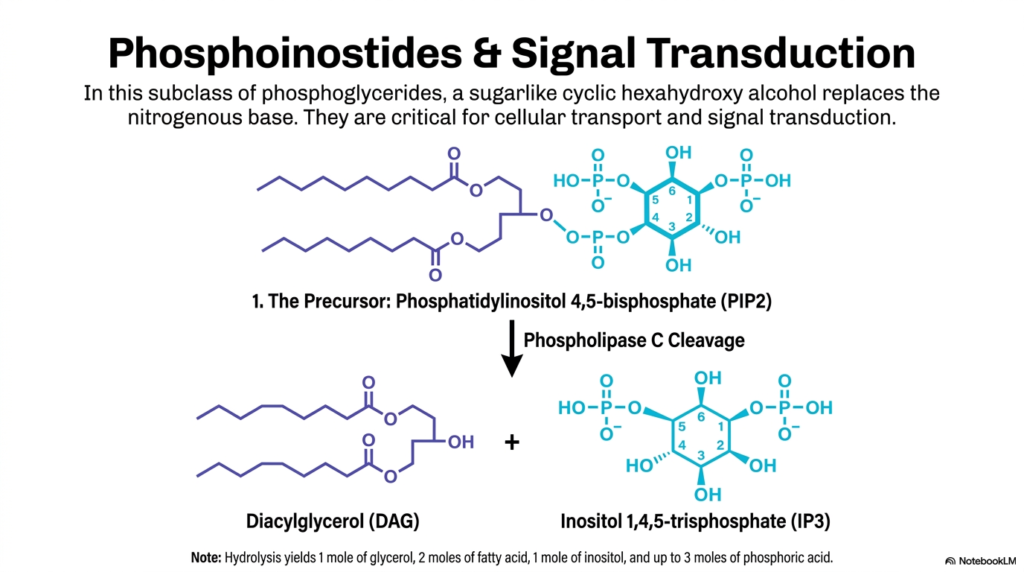
Membrane lipids are not just passive bricks in a wall; they are active, dynamic participants in cellular communication. This slide illustrates one of the most vital signaling pathways you will encounter in medicine, highlighting how specialized Phospholipids known as phosphoinositides are utilized for signal transduction.
In this specific subclass of Phospholipids, the standard nitrogenous base is replaced by a sugar-like cyclic alcohol called myo-inositol. The star player here is Phosphatidylinositol 4,5-bisphosphate, commonly referred to as PIP2. PIP2 sits quietly in the inner leaflet of the cell membrane until a signal—like a hormone or a neurotransmitter—binds to a receptor on the outside of the cell. This binding event activates an enzyme called Phospholipase C.
Phospholipase C acts like a pair of molecular scissors, specifically targeting these PIP2 Phospholipids. It cleaves the molecule right at the phosphodiester bond, instantly generating two highly active intracellular “second messengers”: Diacylglycerol (DAG) and Inositol 1,4,5-trisphosphate (IP3). IP3 is water-soluble and rushes into the cytoplasm to release massive stores of calcium from the endoplasmic reticulum. Meanwhile, DAG remains embedded in the membrane to activate Protein Kinase C. Through this single cleavage event, a structural membrane lipid dictates massive downstream physiological changes, proving that lipids are just as crucial to cellular communication as proteins!
Slide 6: Sphingomyelin and the Neural Phospholipids
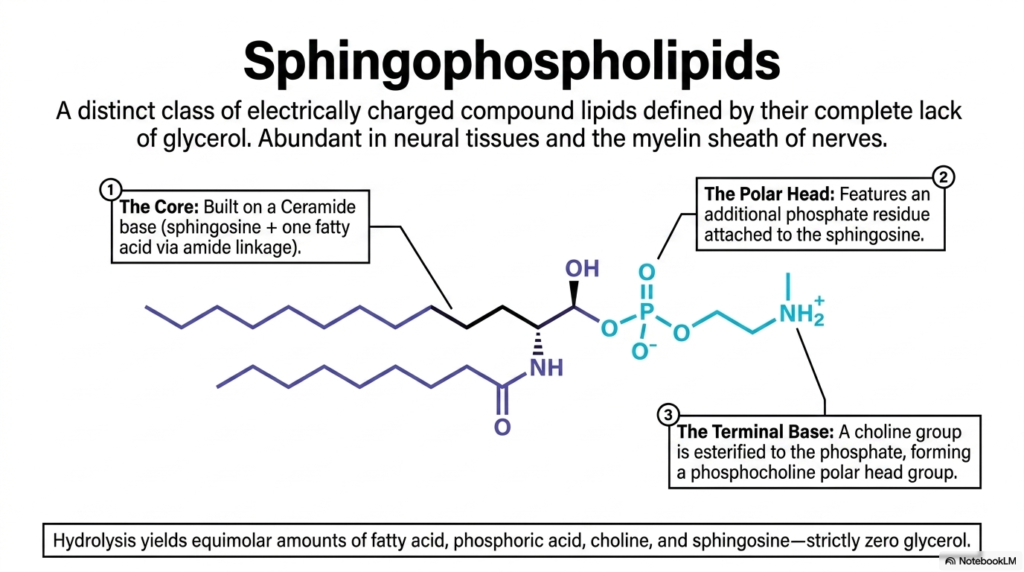
While glycerol-based lipids dominate most of the body, the nervous system requires a much more robust insulation strategy. This slide introduces the sphingophospholipids, a distinct class of electrically charged compound lipids defined by their complete lack of a glycerol backbone. The most famous and biologically significant member of this family is sphingomyelin.
Despite missing glycerol, sphingomyelin is still officially classified among the Phospholipids because it contains a phosphate group in its polar head. The core of this molecule is a ceramide base—composed of a sphingosine backbone attached to a single fatty acid via an incredibly stable amide linkage. To create sphingomyelin, the cell attaches a phosphate residue to the sphingosine, and then esterifies a choline group to that phosphate, forming a phosphocholine polar headgroup.
Why do medical students need to care about this specific variation of Phospholipids? Because sphingomyelin is exceptionally abundant in neural tissues and is the primary structural component of the myelin sheath—the fatty insulation that wraps around the axons of your nerve cells. The unique geometry and hydrogen-bonding capabilities of the ceramide backbone allow these lipids to pack tightly together, creating an incredibly effective electrical insulator. When the body fails to properly synthesize or maintain these specific neural Phospholipids, it leads to devastating demyelinating diseases like Multiple Sclerosis, where nerve impulses literally short-circuit.
Slide 7: Moving Beyond Phospholipids: Simple Glycosphingolipids
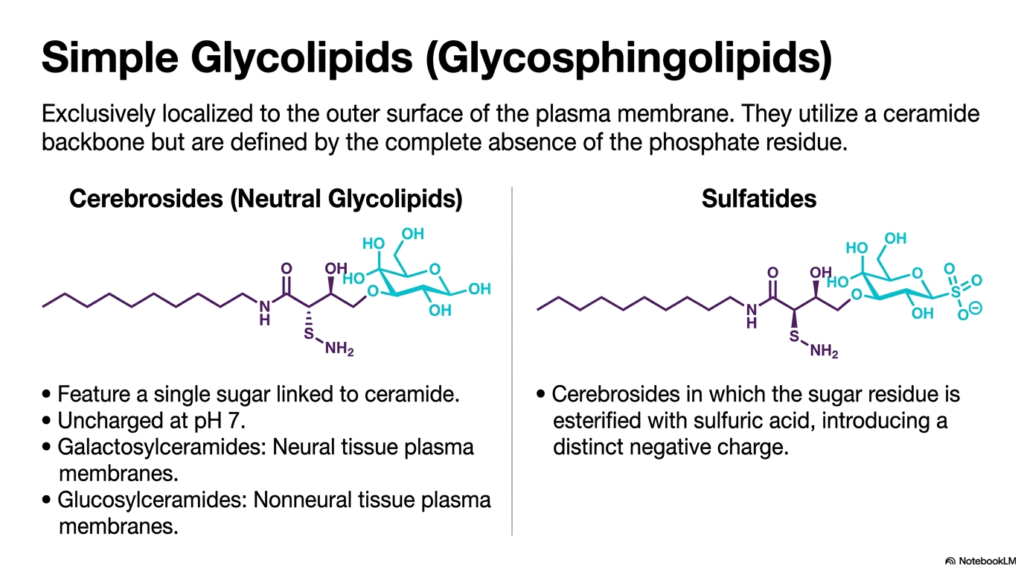
As we journey outward to the very edge of the cell, we encounter a different breed of lipids. This slide introduces the simple glycolipids, specifically the glycosphingolipids. Unlike the Phospholipids we have discussed so far, these molecules are defined by the absolute absence of a phosphate residue. Instead of phosphate, they use complex carbohydrates to interface with the aqueous environment.
These glycolipids share the same tough ceramide backbone (sphingosine plus a fatty acid) as the neural lipids, but their heads are completely different. The simplest variants are the Cerebrosides, which feature just a single sugar unit linked to the ceramide. Because they lack a negatively charged phosphate group, neutral cerebrosides remain uncharged at physiological pH. For example, galactosylceramides are highly enriched in neural tissue membranes, while glucosylceramides are found more commonly in non-neural tissues.
Another crucial variant shown here is the Sulfatide. By esterifying the sugar residue of a cerebroside with sulfuric acid, the cell introduces a distinct negative charge to the molecule. It is absolutely vital to note that unlike Phospholipids, which are found on both sides of the lipid bilayer, these glycolipids are exclusively localized to the outer surface of the plasma membrane. They act as the “face” of the cell, interacting with the extracellular matrix and neighboring cells, rather than providing the core structural barrier.
Slide 8: Complex Glycolipids alongside Phospholipids
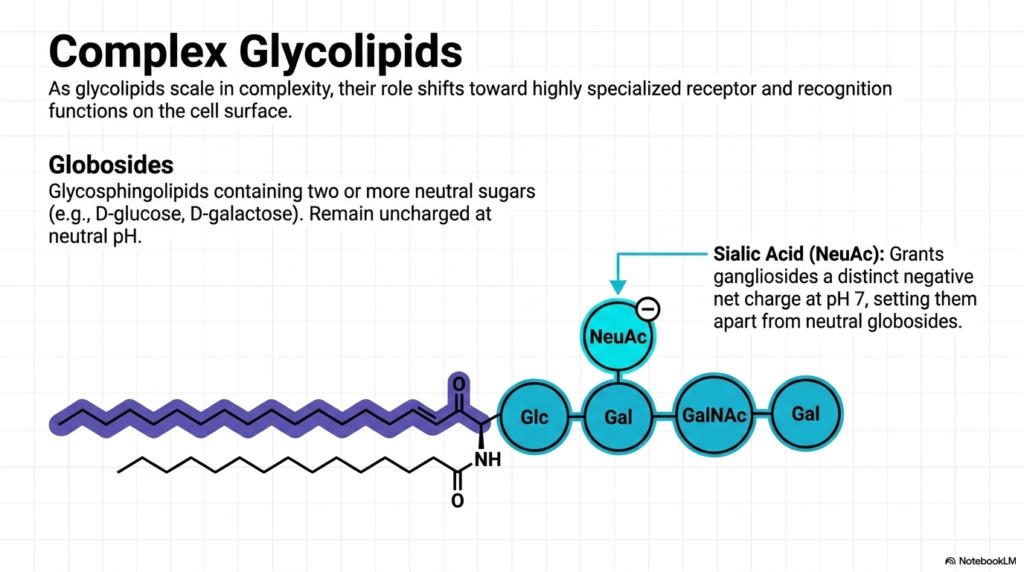
As glycolipids scale in structural complexity, their biological role shifts dramatically from basic barrier support to highly specialized receptor and recognition functions. This slide details the complex glycolipids, which share the outer membrane space with traditional Phospholipids but extend massive, intricate carbohydrate branches far out into the extracellular space.
We categorize these complex molecules into two main groups: Globosides and Gangliosides. Globosides are simply glycosphingolipids containing two or more neutral sugars, such as D-glucose or D-galactose. Because their sugars are neutral, the entire molecule remains uncharged. Gangliosides, however, are the heavyweights of the lipid world. They feature massive oligosaccharide chains that include a very specific, charged sugar molecule called N-acetylneuraminic acid (NeuAc), more commonly known as Sialic Acid.
The presence of this Sialic Acid grants gangliosides a distinct, strong negative net charge at physiological pH. While Phospholipids handle the internal signaling and structural integrity of the bilayer, gangliosides act as sophisticated identification tags and cellular receptors on the surface. They play massive roles in cell-to-cell recognition, embryonic development, and immunity. In fact, many dangerous pathogens and bacterial toxins (like the Cholera toxin) specifically hunt for and bind to these complex gangliosides to gain entry into your cells!
Slide 9: A Taxonomic Summary of Phospholipids and Glycolipids

Biochemistry involves mastering an overwhelming number of molecules, so this slide provides a crucial taxonomic summary of the compound lipid families. This table is an excellent high-yield review tool for distinguishing between the various classes of Phospholipids and glycolipids based on their backbone, fatty acid count, headgroup linkage, and net charge.
The top half of the table is dominated by the phosphoglycerides and phosphoinositides—the classic glycerol-based Phospholipids. You can clearly see that they all utilize a glycerol backbone and contain two esterified fatty acids. Their diversity and net charge (neutral vs. negative) are determined entirely by their specific headgroups, such as choline, serine, or myo-inositol.
The bottom half of the table shifts to the sphingosine-based lipids. Notice that sphingomyelin bridges the two worlds: it is categorized under sphingophospholipids because it contains a phosphate group, making it part of the Phospholipids family, yet it utilizes the single-fatty-acid amide linkage characteristic of sphingosine. Finally, the simple and complex glycolipids (cerebrosides, sulfatides, and gangliosides) lack phosphate entirely, relying on sugars and sulfates to determine their biochemical properties and charges. Memorizing these fundamental differences is essential for understanding how cells construct highly specific, functionally distinct regions within their membranes.
Slide 10: Lipid Rafts and the Spatial Organization of Phospholipids
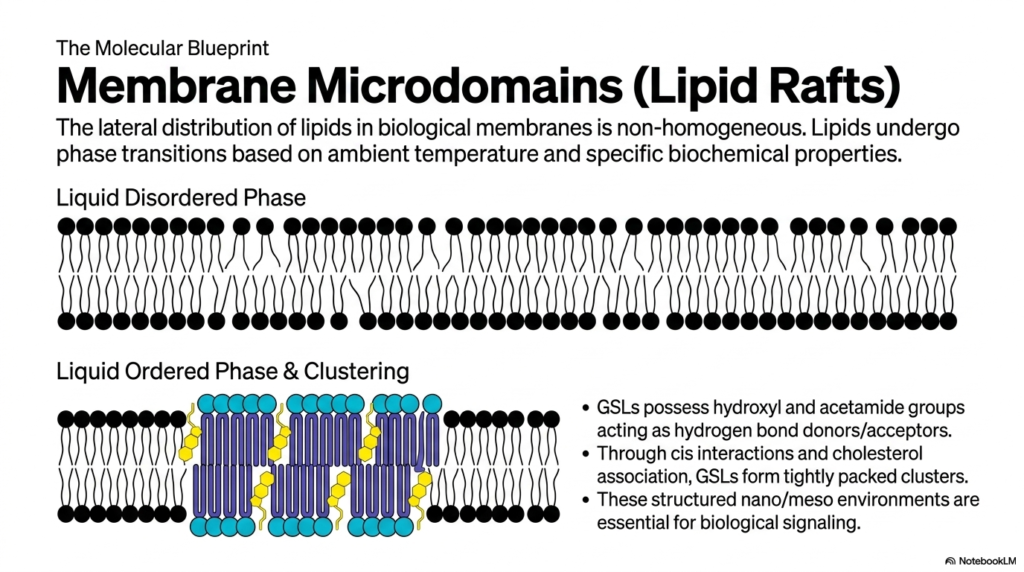
Now that we have cataloged the lipid types, we need to look at how they actually behave together in a living membrane. Biological membranes are not perfectly homogenous, well-mixed oceans. Instead, they organize into highly specific domains. This slide introduces the concept of Membrane Microdomains, more popularly known as “Lipid Rafts,” which float within the broader sea of standard Phospholipids.
In a standard “Liquid Disordered Phase” (the top illustration), the unsaturated tails of normal glycerophospholipids are kinked and loosely packed, creating a highly fluid, flexible membrane. However, when cells need to create a stable platform for signaling proteins, they utilize glycosphingolipids (GSLs) and cholesterol to form a “Liquid Ordered Phase” (the bottom illustration). GSLs possess hydroxyl and acetamide groups that act as powerful hydrogen bond donors and acceptors.
Through these strong cis interactions and tight association with cholesterol, these specialized lipids pull together tightly, forming dense, highly ordered clusters. These lipid rafts are structurally distinct and physically thicker than the surrounding sea of disordered Phospholipids. By segregating specific lipids into these tightly packed micro-environments, the cell creates specialized signaling hubs. These rafts act as docking bays, ensuring that the necessary receptor proteins and enzymes are kept close together so they can interact rapidly when a biological signal arrives.
Slide 11: Trans-bilayer Interdigitation Across the Phospholipid Sea
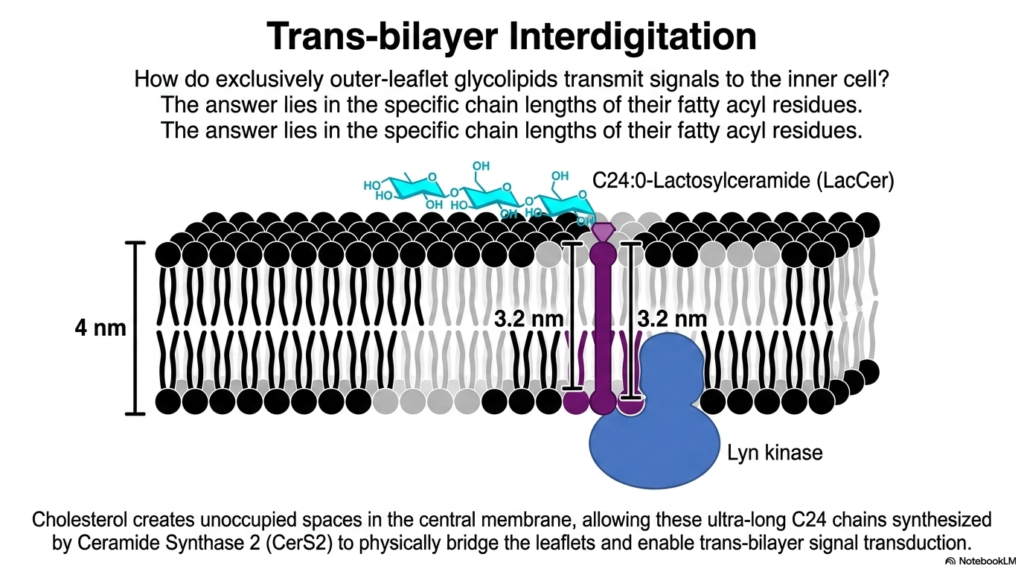
A major structural puzzle in cell biology is how a signal received on the purely extracellular outer leaflet can be transmitted to the intracellular machinery, especially when the lipid bilayer physically separates them. This slide reveals the ingenious biochemical solution: trans-bilayer interdigitation. While standard Phospholipids usually stay strictly within their own leaflet, certain specialized lipids break the rules.
Here we see a specific outer-leaflet glycolipid called Lactosylceramide (LacCer). Look closely at its biochemical structure: it features incredibly long fatty acyl chains, specifically the ultra-long C24 chain synthesized by the enzyme Ceramide Synthase 2 (CerS2). The total width of a cellular membrane is roughly 4 nanometers. A standard phospholipid tail only reaches halfway across. However, the ultra-long 3.2-nanometer tail of LacCer physically pushes past the midplane of the membrane!
To make room for this massive lipid tail, cholesterol creates unoccupied spaces in the central hydrophobic core among the normal Phospholipids. By physically bridging both leaflets, the ultra-long tail of the outer-leaflet LacCer can directly interact with inner-leaflet signaling proteins, such as the Lyn kinase shown in blue. This structural phenomenon, known as interdigitation, proves that the two layers of the membrane are not isolated systems; they are physically and mechanically coupled, allowing for instantaneous signal transduction across the boundary.
Slide 12: Microdomain Contextures Sinking into Phospholipids
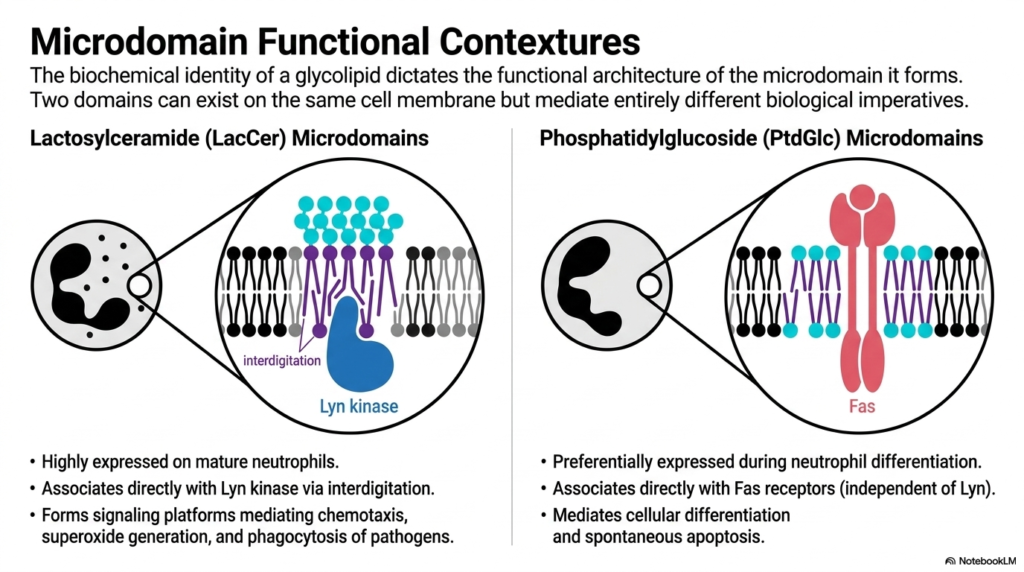
Building on the concept of interdigitation, this slide demonstrates how the biochemical identity of a single lipid dictates massive biological imperatives. Even when two different microdomains exist on the exact same neutrophil cell membrane, floating in the exact same ocean of Phospholipids, they mediate entirely different cellular outcomes based on their core lipid composition.
On the left, we have Lactosylceramide (LacCer) microdomains. As discussed, these domains utilize interdigitation to directly associate with Lyn kinase. These specific platforms are highly expressed on mature neutrophils and serve as the command centers for active immune responses, mediating chemotaxis, superoxide generation, and the phagocytosis of invading pathogens. They are built for action and defense.
On the right, we contrast this with Phosphatidylglucoside (PtdGlc) microdomains. PtdGlc is an unusual lipid that preferentially pairs with Fas receptors, completely independent of Lyn kinase. Instead of triggering an attack, these microdomains mediate cellular differentiation and spontaneous apoptosis (programmed cell death). This is a profound concept for medical students to grasp: the physical clustering of specific membrane lipids alongside standard Phospholipids isn’t just structural trivia; it literally determines whether an immune cell will attack a bacterium or trigger its own cellular suicide.
Slide 13: Pattern Recognition Alongside Membrane Phospholipids
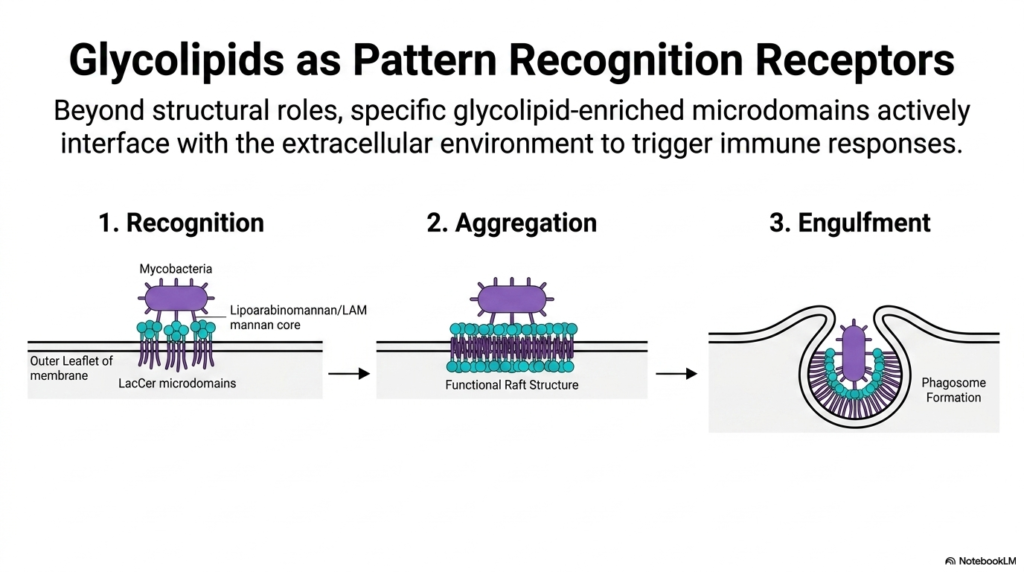
This slide takes the biochemical concepts we’ve discussed and applies them to a real-world immunological event. Beyond their structural roles, glycolipid-enriched microdomains actively interface with the extracellular environment to act as “Pattern Recognition Receptors.” While standard Phospholipids maintain the barrier, these raft structures hunt for danger.
The diagram breaks down the immune response into three phases. First is “Recognition.” Notice how the outer-leaflet LacCer microdomains physically bind to the lipoarabinomannan (LAM) and mannan cores present on the surface of dangerous mycobacteria. The glycolipids themselves are directly identifying the pathogen!
Once recognition occurs, we move to “Aggregation.” The binding event causes multiple, smaller LacCer microdomains to rapidly cluster together into a massive, functional raft structure. This large-scale lipid aggregation displaces the normal fluid Phospholipids, creating a dense signaling platform. Finally, this massive signaling hub triggers “Engulfment.” The intense intracellular signaling (mediated by the interdigitation we discussed earlier) causes the cell’s cytoskeleton to physically remodel the membrane, reaching up and swallowing the bacteria to form a closed vesicle called a phagosome. Here, the architectural lipids act as both the sensor and the physical mechanism of immune defense.
Slide 14: Restructuring Phospholipids and Membranes for Antigen Presentation
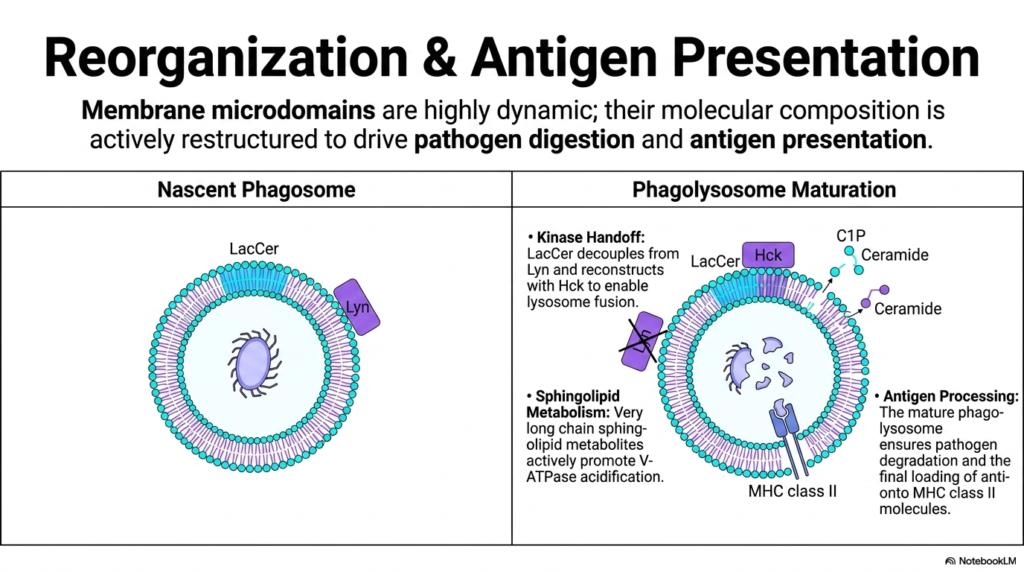
Our final slide explores the climax of the cellular immune response, illustrating how incredibly dynamic these lipid structures truly are. Once the bacteria is engulfed into a nascent phagosome, the lipid membrane cannot remain static. Its molecular composition must be actively restructured—moving away from structural Phospholipids and toward specialized lipid metabolites—to drive pathogen digestion and antigen presentation.
In the initial nascent phagosome, the LacCer microdomains are still coupled with Lyn kinase. However, as the phagolysosome matures, a critical “Kinase Handoff” occurs. LacCer physically decouples from Lyn and reconstructs a new signaling complex with Hck kinase. This specific biochemical switch is required to enable the phagosome to fuse with lysosomes.
Simultaneously, the cell undergoes massive sphingolipid metabolism. Enzymes rapidly break down complex lipids to generate very long-chain sphingolipid metabolites like ceramides. These newly generated lipids actively promote the function of V-ATPases, massive protein pumps that flood the phagolysosome with acid to dissolve the pathogen. Ultimately, this complete reorganization of the Phospholipids and glycolipids ensures total pathogen degradation and allows the cell to execute its final, crucial task: loading the digested bacterial antigens onto MHC class II molecules to alert the rest of the immune system. The membrane is never static; it is a constantly evolving biochemical machine!
Please read our Content Disclaimer Statement.
Check out our social media channels: