8. Carbohydrate Biochemistry: An Academic Overview
When most people out in the real world hear the word “carbohydrate,” they instantly think of a plate of pasta, a loaf of bread, or the impending exhaustion of a sugar crash. But as you transition into the rigorous, high-stakes fields of medicine and advanced cellular biology, you need to completely rewire how you view these specific molecules. Carbohydrates are not just simple dietary fuel; they are highly complex, structurally dynamic, and incredibly versatile building blocks that make human life fundamentally possible.
The core purpose of this slide deck is to guide your understanding from basic high school biology to advanced molecular architecture. We are going to thoroughly explore the precise chemical mechanisms, the dynamic three-dimensional structures, and the critical metabolic reactions of monosaccharides. By the end of this journey, you will understand exactly how the unique shape and reactivity of a single microscopic glucose molecule dictate human health, disease, and cellular function. Let’s dive into the endlessly fascinating microscopic world of sugar!
Slide 1: The Architectural Foundation of Carbohydrate Biochemistry
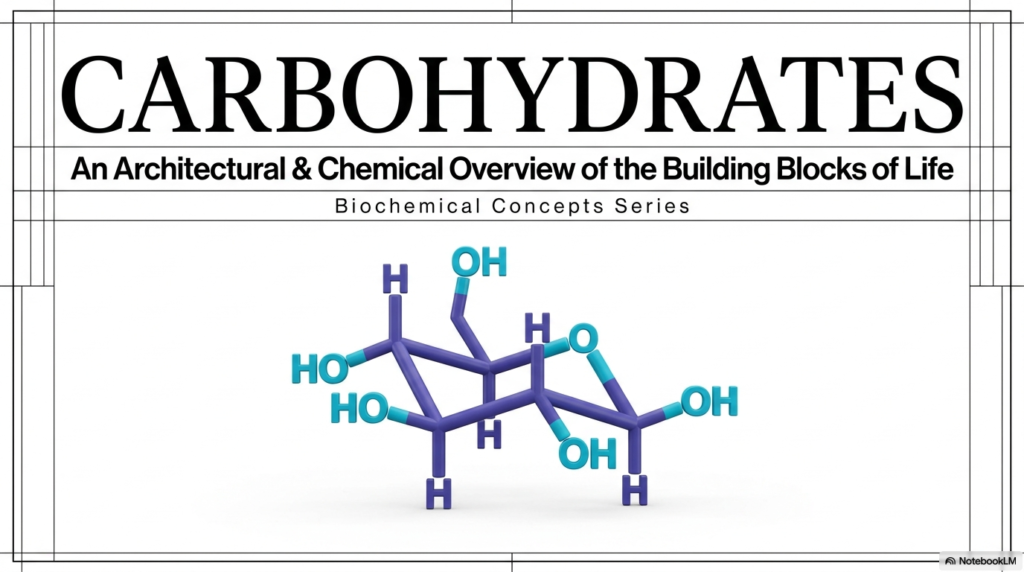
Welcome to the very beginning of our journey into the microscopic world of cellular building blocks! When you look at the first slide of our presentation, you are greeted by a beautiful, three-dimensional ball-and-stick model of a monosaccharide. This image serves as the perfect visual anchor for the entire field of Carbohydrate Biochemistry. To the untrained eye, it might just look like a random assortment of blue and teal spheres connected by sticks. However, as future medical professionals and research scientists, you must view this structure as a highly specific, finely tuned architectural marvel.
In the study of Carbohydrate Biochemistry, we quickly learn that structure is destiny. The precise orientation of those hydroxyl groups—the teal spheres—determines exactly how this molecule will interact with water, how it will bind to complex cellular receptors, and whether it can be efficiently broken down for rapid energy. Historically, sugars were thought of simply as hydrated carbons, a basic chemical concept. But modern analysis reveals a landscape of extreme complexity. The spatial arrangement shown in this cyclic model is what allows sugars to form massive, complex functional polymers.
Think of this single molecule as a highly versatile molecular Lego brick. Depending on how you biochemically snap it together with other structural bricks, you can build a flexible, temporary energy reserve or an impenetrable bacterial cell wall. Understanding this foundational molecular architecture is absolutely critical. It is the core basis for everything from how your brain functions during an exhausting exam to how your immune system identifies foreign pathogens. As we progress through this slide deck, keep this 3D image in your mind.
We are going to unpack how this simple structural blueprint dynamically adapts and reacts to keep the human body alive. Mastering these concepts in Carbohydrate Biochemistry is your first crucial step toward succeeding in metabolic science.
Slide 2: The Four Major Biological Roles in Carbohydrate Biochemistry
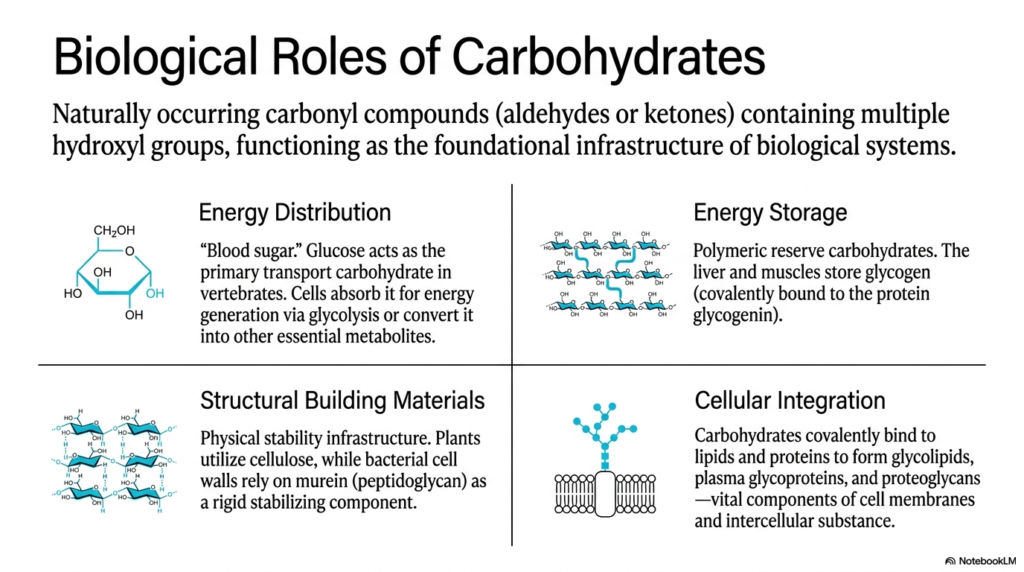
If you want to understand exactly why we dedicate so much precious time to Carbohydrate Biochemistry in medical school, you only need to look at the immense biological utility outlined on this second slide. Carbohydrates are naturally occurring carbonyl compounds with multiple hydroxyl groups, but their function extends infinitely beyond simply providing “blood sugar.” This slide beautifully categorizes their biological roles into four major quadrants: Energy Distribution, Energy Storage, Structural Building Materials, and Cellular Integration. Each of these unique domains represents a massive, non-negotiable pillar of human physiology and cellular survival.
First, let’s talk about Energy Distribution. Glucose is the primary transport carbohydrate in vertebrates. It aggressively flows through your bloodstream, ready to be instantly absorbed by cells and converted into ATP through the rapid process of glycolysis. Without this fundamental aspect of Carbohydrate Biochemistry, the high-energy demands of the human brain and muscular system could never be met. But the body is incredibly efficient, which brings us to Energy Storage. When you consume excess glucose, your liver and muscles don’t just throw it away. They cleverly link these monosaccharides together to form glycogen, a massive, densely branched polymeric structure covalently anchored to a core protein called glycogenin.
Beyond energy, carbohydrate biochemistry strongly dictates structural integrity and complex cellular communication. While humans don’t rely on structural cellulose as plants do, we constantly interact with bacterial cell walls composed of murein (peptidoglycan). Understanding this structural carbohydrate is essential because life-saving antibiotics like penicillin work exclusively by destroying this very infrastructure! Finally, Cellular Integration illustrates how carbohydrates are covalently bound to lipids and proteins. These complex glycoproteins and glycolipids heavily coat the surface of every cell in your body. Your immune system relies entirely on these specific sugar tags to recognize which cells belong to you and which are highly dangerous foreign invaders.
Slide 3: Mastering Open Chain Architecture in Carbohydrate Biochemistry
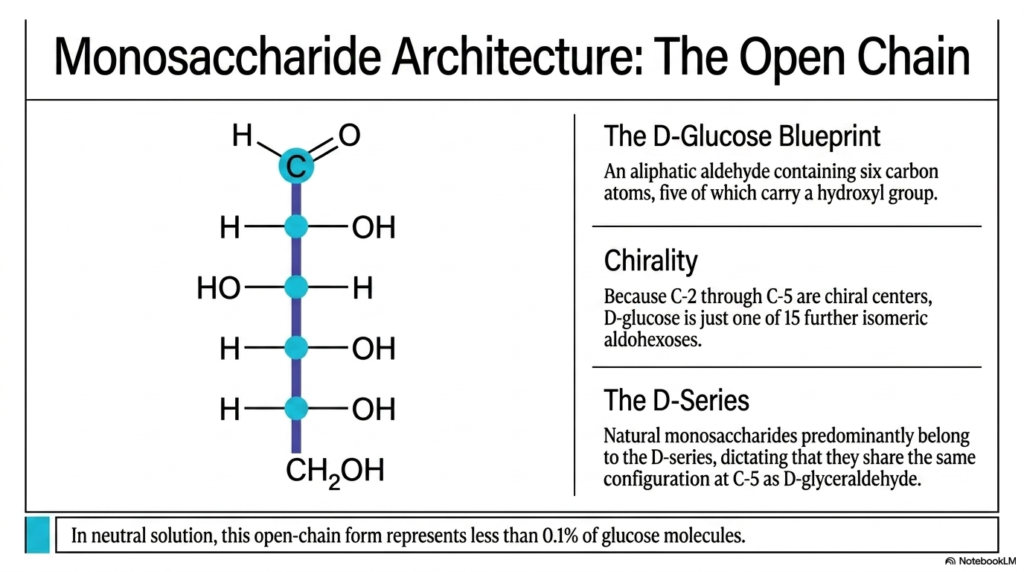
To truly grasp the mechanisms driving Carbohydrate Biochemistry, we must strip the massive sugar molecule down to its most basic, two-dimensional blueprint: the open-chain form. Slide 3 introduces us to the linear architecture of D-glucose, an aliphatic aldehyde containing exactly six carbon atoms, five of which carry a highly reactive hydroxyl group. While it might look like a simple, unassuming straight line of carbon atoms, this diagram is densely packed with critical stereochemical information.
The real magic—and the real challenge for biochemistry students—lies in the concept of molecular chirality. Because carbons 2, 3, 4, and 5 are chiral centers, meaning they are bonded to four entirely different chemical groups, D-glucose is just one of 16 possible isomeric aldohexoses!
This staggering level of structural variation is a defining hallmark of Carbohydrate Biochemistry. A microscopic physical change, such as flipping a single hydroxyl group from the right side to the left side of the carbon chain, generates an entirely different sugar with wildly different biological properties and enzyme affinities. Our bodies are incredibly specific, primarily recognizing and actively utilizing sugars from the “D-Series.” As the slide explicitly notes, a sugar belongs to the D-series if the hydroxyl group on its highest-numbered chiral center (Carbon-5) points to the right, perfectly matching the orientation of standard D-glyceraldehyde.
However, there is a massive biological caveat that you must permanently burn into your memory when studying Carbohydrate Biochemistry. We explicitly draw glucose in this straight, open-chain schematic primarily for educational convenience—to easily count the constituent carbons and identify the stereoisomer. But in the neutral, aqueous environment of a living human cell, this open-chain form practically does not exist! It actually represents less than 0.1% of the glucose molecules present. Why? Because the open chain is thermodynamically restless. The aldehyde group at the top is highly reactive, driving the entire molecule to aggressively fold in on itself, setting the perfect stage for the crucial cyclization reaction.
Slide 4: Hemiacetal Formation and Ring Equilibrium in Carbohydrate Biochemistry
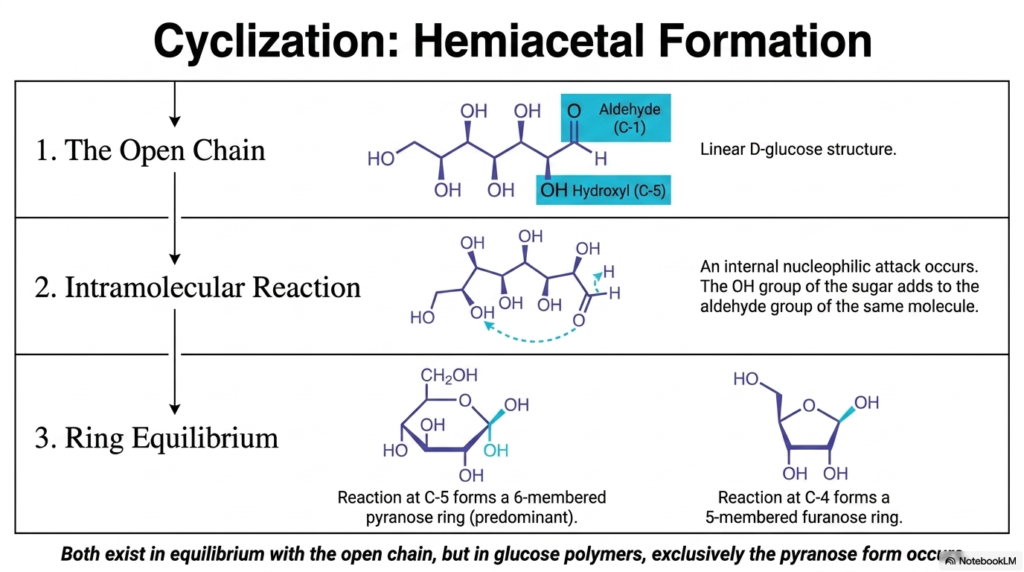
Slide 4 captures what is arguably the most elegant and crucial chemical transformation in all of Carbohydrate Biochemistry: intramolecular cyclization. As we just established, the open linear chain of D-glucose is highly unstable in the warm, water-based environment of our cellular cytoplasm. Because the long carbon backbone is highly flexible, it physically bends and twists, bringing the highly reactive aldehyde group at Carbon-1 into dangerously close proximity with the nucleophilic hydroxyl group at Carbon-5. What happens next is a fundamental, spontaneous reaction that every future physician must deeply understand.
An internal nucleophilic attack naturally occurs. The oxygen atom of the C-5 hydroxyl group aggressively strikes the electrophilic carbonyl carbon at C-1. This rapid reaction forms a tightly closed molecular loop, specifically known in Carbohydrate Biochemistry as a hemiacetal. Understanding this specific, vital ring formation is absolutely paramount because it permanently defines the highly stable shape the sugar will take before it can be used for vital biological functions. Once the stable ring forms, the molecule exists in a constant dynamic equilibrium, but the balance heavily, almost exclusively, favors the closed cyclic structure over the fragile open chain.
Fascinatingly, the flexible nature of the carbon chain means cyclization could theoretically occur at distinct carbon sites, directly leading to rings of different sizes. As the slide explicitly illustrates, if the C-5 hydroxyl group attacks, we generate a remarkably stable 6-membered ring known as a pyranose. If the C-4 hydroxyl group were to attack instead, we would end up with a much tighter 5-membered furanose ring. A core, non-negotiable rule of Carbohydrate Biochemistry is understanding thermodynamic stability. While both ring sizes exist in a watery equilibrium alongside the tiny fraction of open-chain molecules, nature strongly prefers the 6-membered pyranose form. In natural glucose polymers, glucose is found exclusively in this highly stable pyranose configuration!
Slide 5: Visualizing 3D Sugar Projections in Carbohydrate Biochemistry
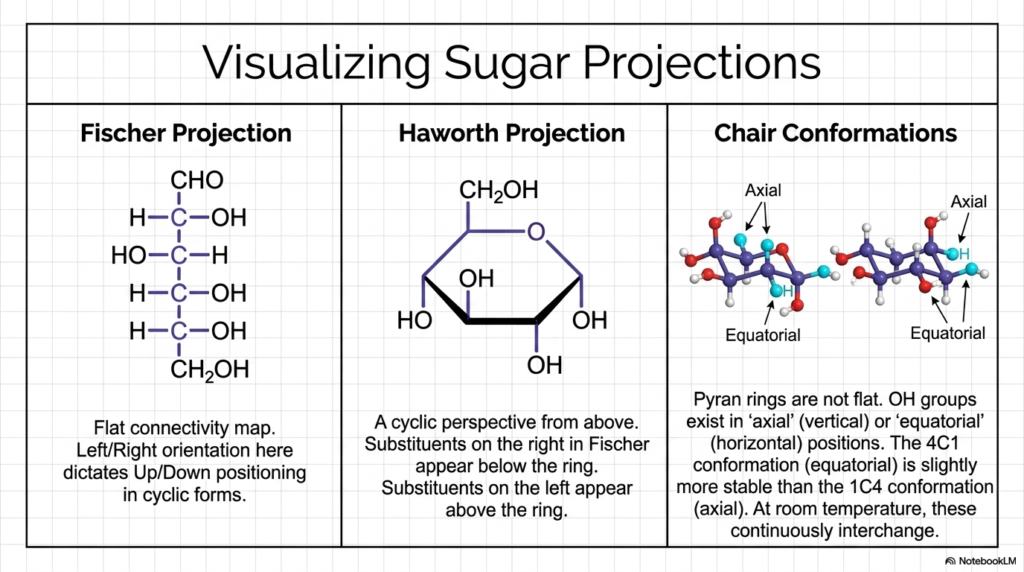
Welcome to the specific visual language of Carbohydrate Biochemistry! Because sugar molecules are complex, three-dimensional physical objects, structural biochemists need standardized methods to accurately represent them on a flat, two-dimensional surface. Slide 5 successfully breaks down the three primary projection models you will continuously encounter in your advanced studies: the Fischer Projection, the Haworth Projection, and the highly accurate Chair Conformation. First, we have the Fischer Projection. Think of this as the perfectly flat, architectural floor plan. It is highly artificial, but it brilliantly allows us to see exactly which hydroxyl groups point left and which point right.
Next is the famous Haworth Projection, which is universally loved by students of Carbohydrate Biochemistry because it is relatively easy to draw and interpret quickly. It gives us a cyclic perspective from slightly above, depicting the newly formed pyranose ring as a perfectly flat hexagon. To easily translate from Fischer to Haworth, we use a very simple rule: substituents on the right side of the Fischer projection point “down” below the ring, and those on the left point “up.” However, you must realize the Haworth projection is biologically inaccurate. The strict reality of Carbohydrate Biochemistry dictates that six-membered carbon rings are never truly flat due to the natural, inflexible tetrahedral bond angles of the carbon atoms.
This brings us directly to the Chair Conformation, the absolute closest visual representation of physical reality. The ring actually puckers into a distinct zigzag shape resembling a reclining lawn chair. In this 3D reality, hydroxyl groups must exist in one of two distinct positions: “axial” (sticking straight up or down like a rigid flagpole) or “equatorial” (pointing outward horizontally along the molecule’s equator). This is exactly where steric hindrance—the physical clashing of bulky atoms—becomes crucially important. The 4C1 chair conformation, where the bulky OH groups are perfectly spaced out in the roomy equatorial positions, is significantly more physically stable than the crowded 1C4 axial conformation.
Slide 6: Stereochemistry, Anomers, and Mutarotation in Carbohydrate Biochemistry
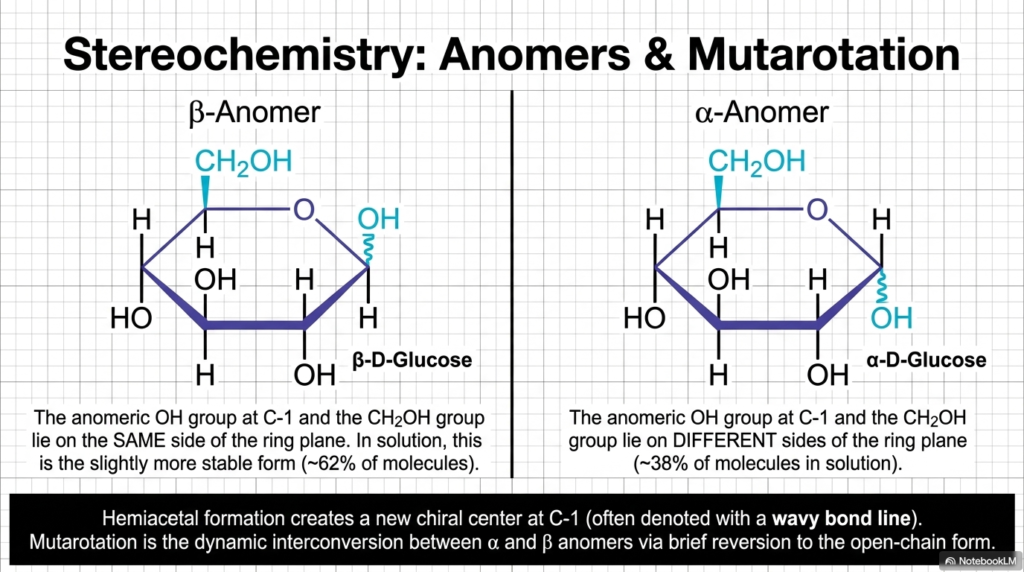
When the highly flexible open-chain glucose molecule suddenly cyclizes into a closed hemiacetal ring, a profoundly important chemical event occurs in Carbohydrate Biochemistry: a brand-new chiral center is magically born out of thin air. The original aldehyde carbon (Carbon-1), which previously had only three chemical attachments, suddenly gains a fourth entirely unique bond. This newly formed chiral center is referred to as the “anomeric carbon” and is often denoted in technical diagrams with a wavy bond line. Because this new OH group can structurally lock into place, pointing either up or down, it directly gives rise to two distinct stereoisomers called anomers: the alpha (α) anomer and the beta (β) anomer.
In the precise, strict language of Carbohydrate Biochemistry, we chemically differentiate these anomers by analyzing the spatial relationship between the anomeric OH group at C-1 and the terminal CH2OH group projecting from the top of the ring. In the beta-anomer of D-glucose, the OH group at C-1 and the CH2OH group structurally project onto the exact same side of the ring plane. Because this specific physical configuration actively minimizes atomic crowding (placing both groups in the roomy equatorial position in 3D space), the beta-anomer is thermodynamically more stable, accounting for roughly 62% of free glucose molecules in a given solution. In sharp contrast, the alpha-anomer features these groups on opposite sides, firmly accounting for the remaining 38%.
But here is the most absolutely fascinating part of Carbohydrate Biochemistry: these rings do not just pick a shape and stay permanently frozen. They undergo a highly dynamic, incredibly continuous process called mutarotation. Because the internal hemiacetal bond is readily reversible in water, the sugar ring is constantly popping open into the linear chain for a fraction of a millisecond and quickly snapping back shut. Every time it closes, it has a distinct chance of flipping its configuration from alpha to beta, or from beta to alpha. This beautifully proves that simple sugars are energetic, breathing molecules!
Slide 7: Polarimetry and the Analytical Measurement of Carbohydrate Biochemistry
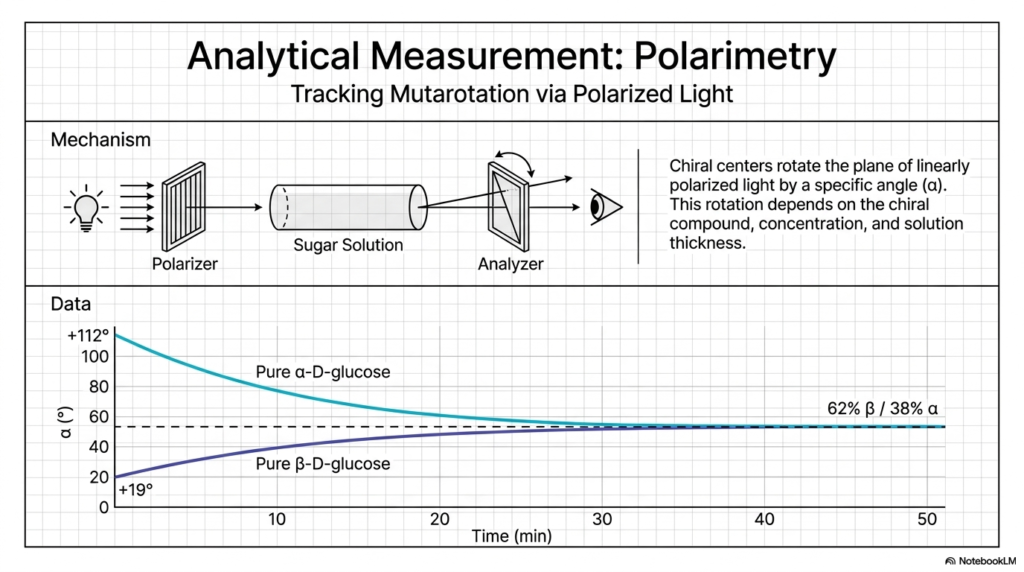
You might logically be wondering: if this rapid mutarotation is happening at an invisible microscopic level, how do clinical biochemists actually know it exists? We cannot simply put a single cell under a standard light microscope and watch the tiny molecules pop open and closed. In Carbohydrate Biochemistry, we rely heavily on a brilliant, historically significant analytical technique known as polarimetry, which is beautifully illustrated here on Slide 7.
Because sugar molecules are strictly chiral—meaning they fundamentally lack internal planes of symmetry—they have a highly unique physical ability to interact with beams of light. When we carefully pass a beam of linearly polarized light through a glass tube containing a solution of a sugar, the chiral molecules physically rotate the light by a very specific, accurately measurable angle.
This exact rotation angle (denoted universally as alpha, α) depends entirely on the specific chiral compound, the precise concentration of the solution, and the physical length of the glass tube. This slide wonderfully showcases one of the most famous, foundational experiments in all of Carbohydrate Biochemistry. If a scientist isolates completely pure, crystallized alpha-D-glucose and completely dissolves it in water, the initial burst of light rotation will read exactly +112 degrees. Conversely, if they strictly isolate pure beta-D-glucose, the initial rotation predictably reads a much lower +19 degrees. But as you watch the graphed data over time, something deeply remarkable logically occurs right before your eyes!
The high optical angle of the pure alpha solution slowly and predictably drops, while the low angle of the pure beta solution steadily rises. Eventually, as the minutes continually tick by, both solutions perfectly plateau at the exact same middle value: +52.7 degrees. This converging graph visually represents the abstract concept of mutarotation in real time! The molecules in both separate solutions are spontaneously opening and reforming, constantly flipping their stereochemistry until they perfectly reach their thermodynamic equilibrium mixture of 62% beta and 38% alpha. This precise analytical measurement is the bedrock proof driving the entirety of Carbohydrate Biochemistry.
Slide 8: Polymerization and Glycosidic Bonds in Carbohydrate Biochemistry
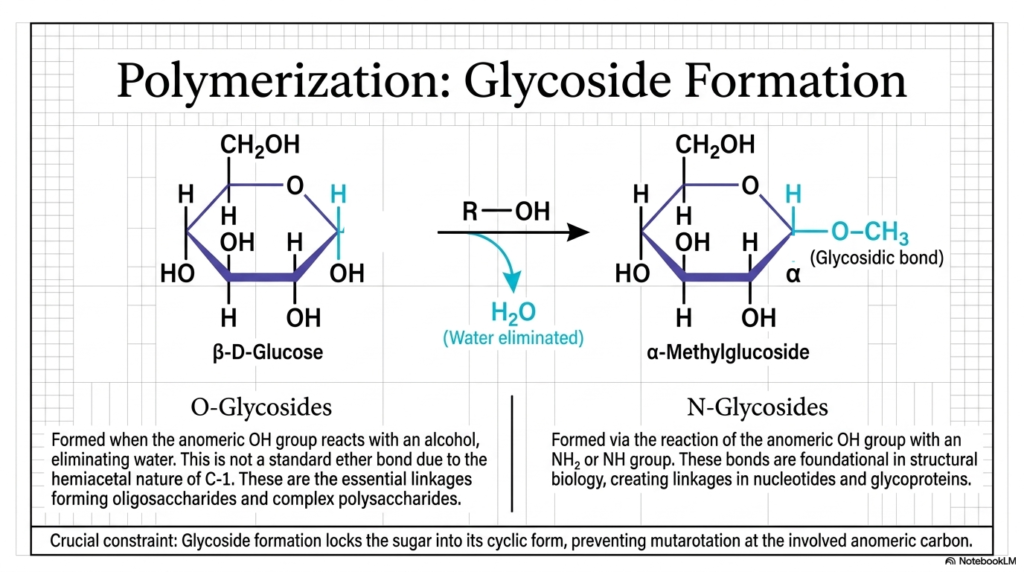
We are now officially graduating from understanding single, highly isolated molecular units to constructing massive, functional biological polymers. Slide 8 properly introduces the exact chemical mechanism of glycoside formation, which is undeniably the most important, critical linking reaction in all of Carbohydrate Biochemistry. When the highly reactive anomeric hydroxyl group of a closed cyclic sugar (like beta-D-glucose) successfully encounters an alcohol group (R-OH) from another molecule, a classic, textbook dehydration synthesis reaction rapidly occurs. A single, distinct molecule of water is completely eliminated, and a brand-new, incredibly strong covalent bond is permanently forged between the two structures.
The resulting unified, massive structure is explicitly called a glycoside, and the highly vital new chemical connection is the glycosidic bond. It is absolutely critical to note in Carbohydrate Biochemistry that this is not just a standard, easily broken ether bond. Because it involves the unique, complex hemiacetal nature of the C-1 carbon, it requires highly specific cellular enzymes to build it carefully and break it safely. If the sugar binds to another typical hydroxyl group, we form O-Glycosides. These are the fundamental, structural linkages that aggressively string single sugars together to physically create everything from the sweet lactose in the milk you drink to the massive, heavily branched glycogen stores packed deeply into your muscle tissue.
Alternatively, if the reactive anomeric carbon reacts with a nitrogen-containing amine group, we specifically form N-Glycosides. This highly specific, fascinating subset of Carbohydrate Biochemistry is profoundly important to advanced cellular genetics; it is the exact, undeniable chemical mechanism that functionally links sugar rings to specialized nitrogenous bases, actively building the structural backbone of your DNA and RNA! Finally, there is one absolutely crucial biochemical constraint you must memorize: forming a glycosidic bond permanently locks the sugar ring into its closed cyclic form, preventing any further mutarotation at that site. This ensures our complex physiological polymers remain highly structurally sound!
Slide 9: Metabolic Derivatives and Redox Reactions in Carbohydrate Biochemistry
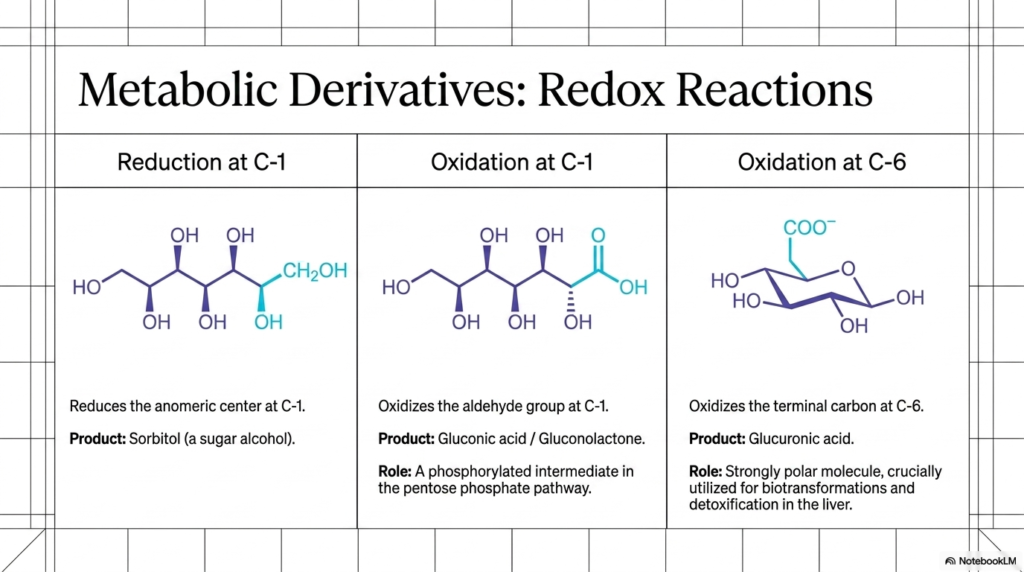
Monosaccharides do not merely link together to form large static polymers; they also eagerly undergo profound, highly targeted chemical modifications to serve wildly specialized metabolic needs. Slide 9 effectively highlights three extremely vital oxidation-reduction (redox) reactions that directly form the functional core of metabolic Carbohydrate Biochemistry. First, let’s carefully look at reduction. If highly specific cellular enzymes successfully reduce the anomeric center at C-1, thereby converting the aldehyde group back into a standard alcohol, we obtain a sugar alcohol, such as Sorbitol.
In medical Carbohydrate Biochemistry, understanding this dangerous pathway is crucial. In chronically ill patients with poorly managed diabetes, excess glucose is continuously reduced to sorbitol in the fragile lens of the eye. Because sorbitol cannot easily cross membranes, it aggressively draws in water osmotically, directly causing devastating diabetic cataracts!
Next, we shift our critical focus to oxidation at the top of the sugar molecule. If we carefully and enzymatically oxidize the reactive aldehyde group at C-1, we generate Gluconic acid (which often rapidly cyclizes to gluconolactone). This is definitely not just random, unimportant chemical trivia. This highly specific oxidized derivative, once successfully phosphorylated, is a mandatory, completely non-negotiable intermediate in the vital pentose phosphate pathway. This protective cellular pathway is absolutely essential for actively protecting your red blood cells from lethal oxidative stress by continuously generating highly protective NADPH.
Finally, we comprehensively examine a massive modification at the bottom of the molecule. If we leave the C-1 aldehyde completely alone but selectively oxidize the terminal carbon at C-6, we generate Glucuronic acid. This entirely unique molecule is an absolute clinical superstar in Carbohydrate Biochemistry. Because the newly formed carboxylic acid group makes the entire molecule exceptionally polar and highly water-soluble, your healthy liver aggressively attaches glucuronic acid to highly toxic drugs, spent steroid hormones, and bilirubin. This critical detoxification process essentially ensures that these dangerous, hydrophobic waste products can be safely and easily flushed out in your urine!
Slide 10: Structural Shifts and Epimerization in Carbohydrate Biochemistry
Slide 10 deeply delves into the remarkable, practically magical flexibility of the simple monosaccharide carbon skeleton, perfectly highlighting a fascinating structural shift universally known as epimerization. In the strict, highly specific terminology of Carbohydrate Biochemistry, epimers are a specialized type of stereoisomer. They are distinct pairs of sugars whose chemical structures are identical in every way except for the exact spatial configuration at one highly specific chiral center. If you look carefully at the slide’s detailed visual, you can clearly see that D-glucose and D-mannose are perfect examples. They are strictly identical molecules, save for the fact that the hydroxyl group at Carbon-2 points in the completely opposite direction!
What is truly, wonderfully fascinating about Carbohydrate Biochemistry is that these differing, highly distinct sugars can actually interconvert under the right specific chemical conditions. In a simple weakly alkaline solution in a standard chemistry lab, D-glucose exists in a highly dynamic equilibrium with D-mannose and D-fructose. This truly remarkable, complex structural transformation occurs by temporarily and energetically shuffling double bonds and protons to form an “enediol intermediate.” This incredibly transient, highly unstable molecular state briefly features a double bond between two carbons that simultaneously bear hydroxyl groups, allowing the molecule to rapidly pivot its stereochemistry before settling back into a stable ring.
While this slide beautifully and perfectly illustrates the shifting chemical equilibrium in a simple glass beaker, this exact Carbohydrate Biochemistry concept is tightly and aggressively controlled in the living human body by highly specific, complex enzymes perfectly called epimerases and isomerases. These incredibly fast enzymes enable your living cells to rapidly and efficiently convert the dietary fructose you easily get from an apple into usable glucose-based metabolic intermediates. Understanding this highly efficient enzymatic shuffling is the exact key to truly mastering how the body universally manages diverse dietary sugar inputs!
Slide 11: Esterification and Metabolic Activation in Carbohydrate Biochemistry
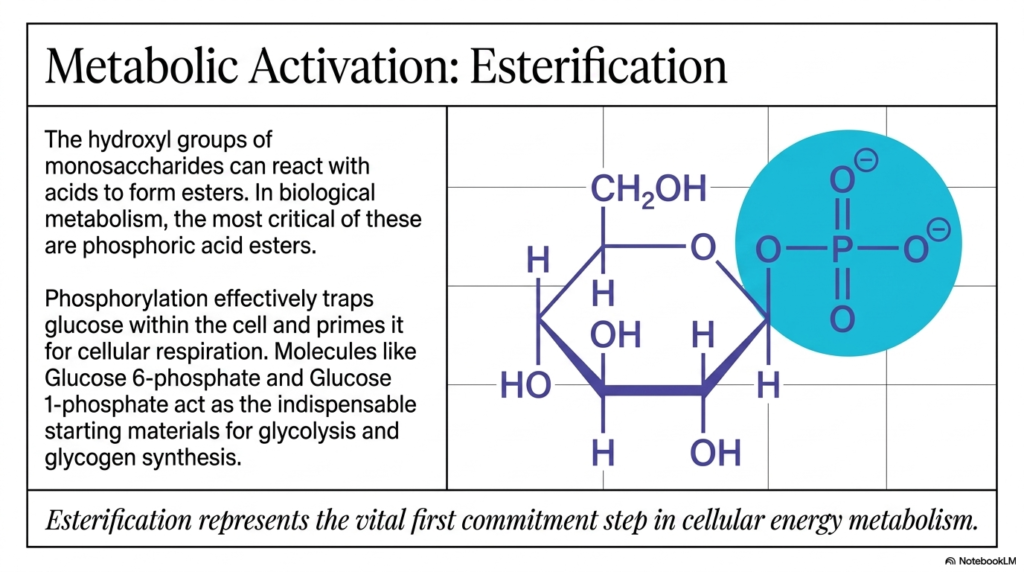
If there is literally one single, monumental slide in this entire lengthy presentation that essentially dictates whether a fragile human cell ultimately lives or dies, it is absolutely Slide 11. Here, we delve deeply into the critical concept of metabolic activation via rapid esterification. The numerous, highly exposed hydroxyl groups densely decorating a monosaccharide ring are chemically reactive and prime, easy targets for rapid reaction with cellular acids to quickly form complex esters. In the grand physiological, life-sustaining scheme of Carbohydrate Biochemistry, the most critically important esters by far are those successfully formed by rapid reactions with bulky phosphoric acid.
When a fresh glucose molecule first eagerly enters a living human cell directly from the bloodstream, the first crucial thing that violently happens is a massive enzymatic ambush. A specialized enzyme, perfectly called hexokinase, violently slaps a massive, highly negatively charged phosphate group directly onto the 6th carbon of the fragile sugar, instantly and aggressively creating Glucose 6-phosphate. Why is this specific, rapid aspect of Carbohydrate Biochemistry so incredibly, non-negotiably vital to human survival? Because the protective plasma membrane of a living cell is highly hydrophobic and tightly regulates what can easily diffuse in and out. By aggressively attaching a highly polar, heavily charged phosphate group, the cell effectively and permanently “traps” the energetic glucose completely inside its walls!
Furthermore, this rapid phosphorylation step perfectly primes the sugar molecule with high, unstable chemical energy, physically destabilizing it just enough to perfectly prepare it for the brutal, demanding rigors of deep cellular respiration. Molecules identical to Glucose 6-phosphate serve as indispensable, highly energized cellular starting materials for both rapid glycolysis and massive glycogen synthesis. In Carbohydrate Biochemistry, rapid esterification isn’t just a random structural modification; it is the absolutely vital first step in all of human cellular energy metabolism. Without this exact trapping mechanism, our cells would instantly and catastrophically starve!
Slide 12: Synthesizing The Glucose Reaction Network in Carbohydrate Biochemistry

We have finally and successfully arrived at the grand, sweeping synthesis of absolutely everything we have comprehensively learned so far. This final, beautifully organized slide serves as your ultimate, highly valuable, high-yield cheat sheet for Carbohydrate Biochemistry, perfectly mapping out the incredibly vital “Glucose Reaction Network.” It beautifully and succinctly summarizes exactly how the seemingly simple, highly flexible six-carbon D-glucose blueprint chemically adapts, violently shape-shifts, and specifically reacts to serve a wildly diverse array of complex biological human functions.
Let’s quickly and effectively review the incredible, complex journey we just comprehensively took through Carbohydrate Biochemistry. We clearly saw how rapid Hemiacetal Formation practically naturally leads directly to solid Pyranose Ring Formation, perfectly satisfying the strict thermodynamic need for a highly stable, physiological structural state. We deeply explored how specialized Glycoside Formation beautifully utilizes both O- and N-linkages to rapidly string those tiny rings together, essentially single-handedly building massive energy-storing polysaccharides and constructing the literal chemical backbone of our highly complex genetic code. We learned that rapid Esterification produces the critical Glucose 6-Phosphate, which serves as the ultimate, inescapable cellular trap and the mandatory, vital starting gun for life-sustaining glycolysis.
We also critically reviewed how highly specific C-6 Oxidation rapidly generates highly polar Glucuronic Acid, which efficiently serves as the liver’s primary, utterly indispensable chemical tool for detoxifying the blood and safely clearing highly dangerous waste. Finally, we saw firsthand how rapid Epimerization brilliantly generates vital isomers, such as Mannose and Fructose, drastically and brilliantly diversifying the highly available metabolic pathways our constantly hungry human cells can easily utilize. Master this single, brilliantly organized summary table, and you essentially master the core, foundational underlying logic of Carbohydrate Biochemistry. You now deeply understand, with absolute clarity, that a sugar molecule is a dynamic, highly engineered chemical machine driving human life!
Please read our Content Disclaimer Statement.
Check out our social media channels: