12. The Chemical Architecture of Fatty Acids: A Biochemical Overview
Have you ever wondered how your body manages to store months’ worth of metabolic energy in remarkably compact spaces, or how the very boundaries of your cellular organelles are forged to protect sensitive genetic material? The core purpose of this entire slide deck is to thoroughly demystify the structural and chemical architecture of Fatty Acids. We will take you on a journey from their most basic atomic structures all the way to their highly complex esterification into massive energy-storage networks.
Slide 1: Introduction to the Biochemistry of Fatty Acids: The Fundamental Blueprint
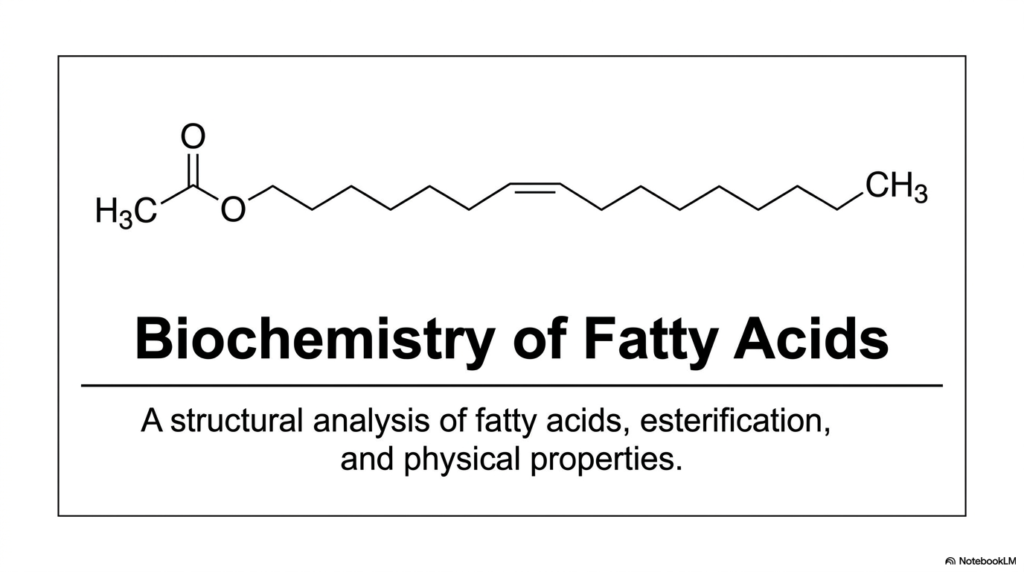
As you look at our first slide, you are introduced to the overarching theme of our biochemistry lecture series: a rigorous structural analysis. At their fundamental core, these molecules are simply composed of long, repeating hydrocarbon chains that are ultimately capped by a single, highly reactive chemical group. But do not let this straightforward visual simplicity fool you. It is the microscopic, seemingly subtle variations within this long carbon backbone that strictly dictate whether a lipid will exist as a harmless, flowing liquid oil at room temperature, or as a dense, solid plaque capable of building up inside a human artery.
Throughout our comprehensive breakdown, we will continuously refer back to this foundational architecture. We will explore exactly how free Fatty Acids behave physically under varying physiological conditions, how they are enzymatically modified by cellular machinery, and how they ultimately bond to other distinct structures. The visual on this slide offers a glimpse of a fully esterified lipid, hinting at the ultimate biochemical fate of most free-circulating lipids. Truly understanding the biochemistry of Fatty Acids is not just an exercise in memorizing static molecular shapes; it is deeply about comprehending the primary biological fuel that relentlessly drives human metabolism and sustains cellular life. Let’s begin our deep dive into the lipid blueprint.
Slide 2: The Dual Anatomy of Fatty Acids: Amphipathic Heads and Hydrophobic Tails
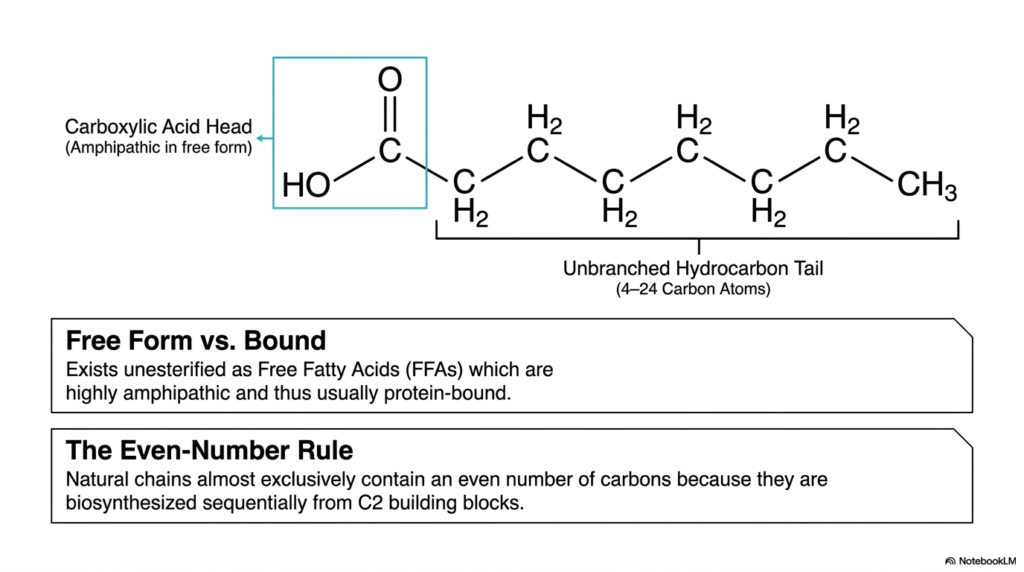
Imagine a unique molecule burdened with a permanent split personality—one side desperately seeking out an aqueous environment, while the opposing side fiercely avoids water at all costs. This distinct dual nature is the defining physical characteristic of free Fatty Acids. Grasping this specific concept is absolutely critical for making logical sense of lipid biochemistry within the human body. The core purpose of this specific slide is to meticulously break down the primary structural domains of a lipid and explicitly explain why they almost universally feature an even number of carbon atoms.
As clearly illustrated in this structural breakdown, Fatty Acids consist of two opposing domains. First, carefully observe the carboxylic acid head highlighted within the blue box. In its free, unesterified biological form, this highly reactive head group is entirely polar and hydrophilic, endowing the overall molecule with its famous amphipathic nature. Because these free Fatty Acids physically act remarkably like biological detergents—which could easily rupture delicate cellular membranes—they are rarely found traveling completely alone in the watery environment of the human bloodstream. Instead, they are almost always transported safely while securely bound to specialized carrier proteins.
Attached directly to this reactive polar head is the defining unbranched hydrocarbon tail, which typically ranges from 4 to 24 carbon atoms in length. This exceptionally long, nonpolar tail region is strictly hydrophobic. When we closely analyze naturally occurring biological lipids, we immediately observe a highly striking structural pattern boldly highlighted at the bottom of the slide: the Even-Number Rule. Almost all naturally synthesized chains contain an even number of carbon atoms. Why does nature insist on this? It ultimately comes down to the body’s internal manufacturing process. During biosynthesis, our cellular machinery sequentially constructs these hydrocarbon chains by exclusively adding distinct two-carbon building blocks.
Slide 3: Decoding the Nomenclature: How We Map Fatty Acids with Delta and Omega Systems
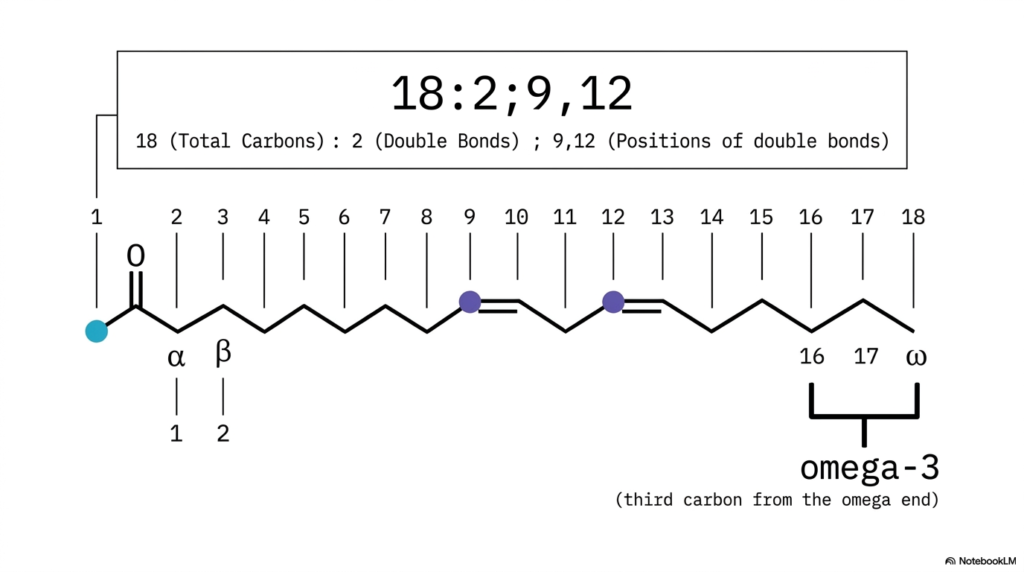
Have you ever stared blankly at a complex biochemical formula and felt as though you were attempting to decipher an encrypted secret code? In the exact science of biochemistry, numerical designations are never arbitrary—they narrate a highly precise structural story. The core purpose of this slide is to effectively teach you how to translate the standard numerical nomenclature of Fatty Acids into a vivid, three-dimensional mental image, while also introducing the highly clinically relevant omega numbering framework used in modern nutrition.
Let us thoroughly break down the standard molecular notation “18:2;9,12” displayed prominently across the slide. This compact numerical formula contains every architectural blueprint you need. The initial number, 18, signifies the total number of carbon atoms in the chain. The specific number immediately following the colon, 2, indicates exactly how many double bonds are present in the structure. Finally, the specific numbers 9 and 12 indicate the exact positions of those double bonds, systematically counting from the highly reactive carboxylic acid head. This universally accepted delta numbering system allows biochemists to quickly communicate the precise chemical nature of diverse Fatty Acids without having to draw them.
However, there is another absolutely critical method we utilize to map these dynamic molecules, particularly in clinical and nutritional contexts: the Omega system. Notice the distinct labels alpha and beta positioned on the second and third carbons. Now, shift your focus to the very end of the tail—the terminal methyl group is designated as the omega carbon.
If you actively count backward from this omega end, the very first double bond you encounter defines the lipid’s nutritional class. In our specific slide example, the first double bond appears precisely on the third carbon from the terminal end, successfully classifying this as a famous omega-3 lipid. Understanding how Fatty Acids fit into both systems is an absolute prerequisite for clinical practice.
Slide 4: The Biological Standards: Why Certain Fatty Acids Dominate in Nature
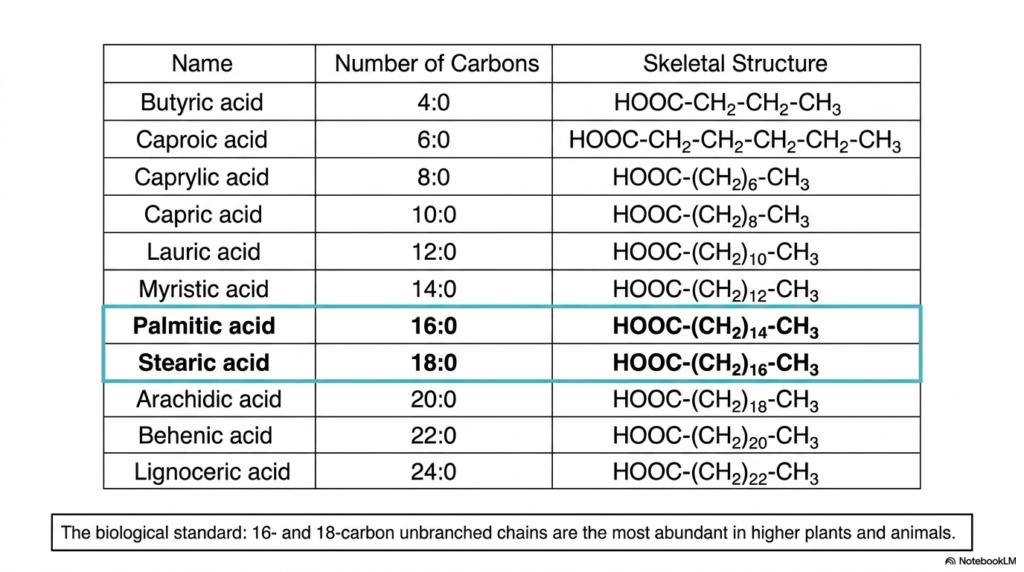
If you were to boldly undertake a comprehensive inventory of every lipid molecule inside the human body, you certainly would not uncover a random, chaotic assortment of carbon chains; rather, you would discover highly specific molecules dominating the biological landscape. Why does biological nature overwhelmingly prefer specific building blocks over others? This slide provides an incredibly detailed catalog of the most prevalent naturally occurring Fatty Acids, with a distinct emphasis on the standard functional molecules that primarily drive higher plant and animal biology.
Carefully looking at the extensive table provided, we can clearly see an array of lipids arranged strictly by their total carbon count. This ranges sequentially from the extremely short 4-carbon butyric acid to the massive 24-carbon lignoceric acid. As we definitively established in our previous slide, every single one of these examples strictly adheres to the biological even-number rule of biosynthesis. However, not all individual Fatty Acids are created functionally equal in terms of their overall biological abundance and physiological utility. The highlighted blue rows explicitly draw our collective attention directly to the true heavyweights of the lipid world: Palmitic acid (16:0) and Stearic acid (18:0).
These specific 16-carbon and 18-carbon unbranched hydrocarbon chains represent the absolute biological standards across countless species. They are, by far, the most overwhelmingly abundant saturated lipids naturally found in higher plants and mammals, including humans. When your highly efficient body actively stores excess dietary energy safely in adipose tissue, or when it rapidly constructs the dense, impermeable sections of your delicate cellular membranes, it heavily relies on these specific palmitic and stearic molecules. Their perfectly straight, fully saturated skeletons actively allow them to pack incredibly tightly together. By intimately familiarizing yourself with these specific Fatty Acids, you are effectively learning the primary metabolic vocabulary of human biology.
Slide 5: Structural Kinks: How Double Bonds Reshape the Geometry of Fatty Acids
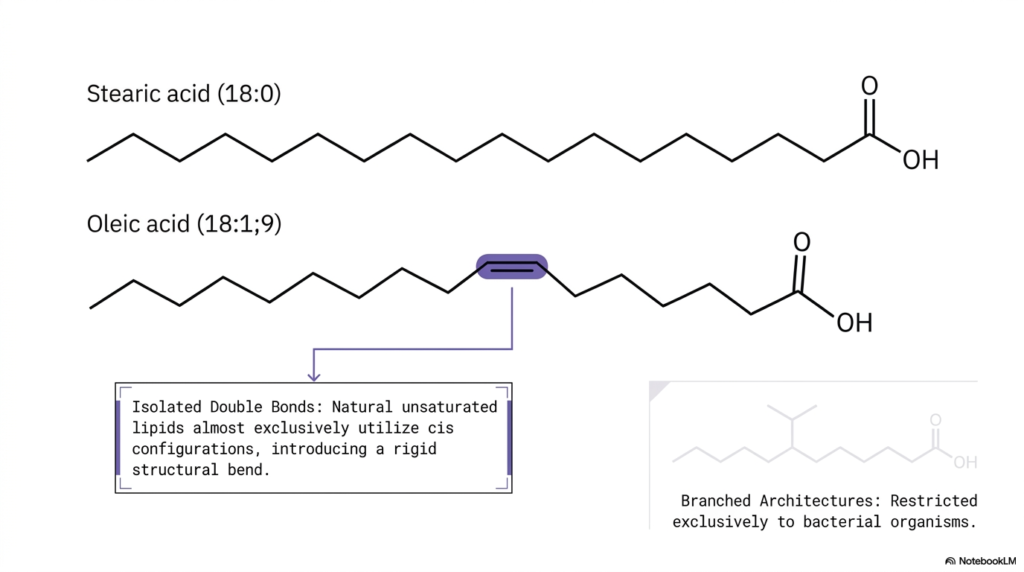
Welcome back to our ongoing structural exploration. In the fascinating realm of advanced cellular biochemistry, even a tiny microscopic modification in a single molecule’s architecture can drastically and permanently alter its entire physiological function. Have you ever pondered why some lipids physically clump together into dense, hard solids while others consistently remain highly fluid, flexible, and slippery? The final answer lies entirely within the physical presence and specific spatial geometry of molecular double bonds. The core purpose of this slide is to visually contrast the dramatically different physical shapes of saturated versus unsaturated Fatty Acids.
Let us carefully examine the top reference molecule featured prominently on the slide: stearic acid (18:0). As we previously learned, this molecule is fully saturated, meaning its long carbon chain contains the maximum possible number of hydrogen atoms and contains only single bonds. As a direct structural result, stearic acid forms a completely straight, highly linear chain. Now, shift your focus immediately below it to the illustrated oleic acid (18:1;9). This unique molecule contains a single, isolated double bond strategically located precisely at the ninth carbon position. In natural biological systems, unsaturated lipids almost exclusively adopt the specific cis double-bond configuration.
What is the ultimate physical consequence of this specific cis configuration? As the accompanying diagram vividly illustrates, it actively introduces a permanent, rigid 30-degree structural bend or kink directly into the middle of the hydrocarbon tail. The perfectly straight geometry of saturated Fatty Acids enables them to stack tightly together. Conversely, the severely bent shape of unsaturated Fatty Acids forcefully pushes adjacent molecules apart, effectively creating vital molecular space. Additionally, the slide briefly draws our active attention to an intriguing evolutionary divergence: highly branched architectures. While mammalian cells rely heavily on straight or kinked chains, highly branched Fatty Acids are found almost exclusively in specialized bacterial organisms.
Slide 6: The Essentiality Rule: Metabolic Limitations in Synthesizing Fatty Acids
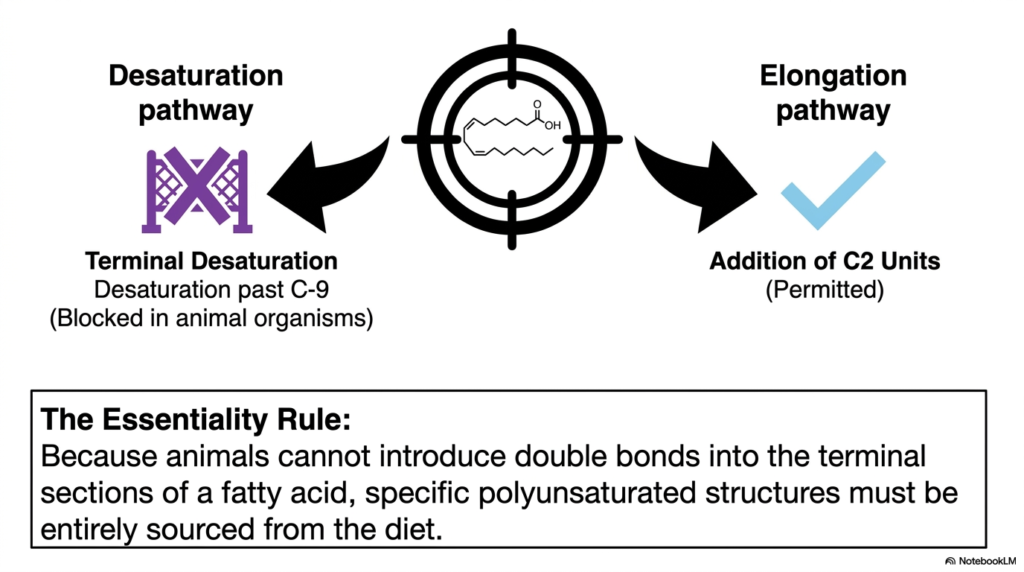
As future medical clinicians and elite biochemistry experts, you will frequently encounter the critical term “essential nutrients” throughout your advanced training, but what exactly renders a molecule biologically essential at a strict structural level? We frequently take highly advanced human metabolism for granted, naturally assuming our cells can effortlessly build any required molecule entirely from basic carbon skeletons. However, our internal cellular machinery actually possesses highly distinct, hardwired enzymatic limitations. The core purpose of this slide is to meticulously define “The Essentiality Rule” by sharply contrasting the permitted metabolic pathways with those strictly blocked, specifically concerning the synthesis of Fatty Acids.
Direct your immediate attention to the left side of the diagram, which visually highlights the crucial desaturation pathway. Desaturation is the critical biochemical process of enzymatically introducing brand new double bonds into a pre-existing carbon chain. While complex human and animal organisms possess the essential enzymes to seamlessly insert double bonds near the reactive carboxylic acid head, we suffer from a severe enzymatic deficiency: terminal desaturation is entirely blocked. Specifically, animal organisms strictly cannot introduce double bonds deep into the distant terminal sections of any molecule past the Carbon-9 (C-9) position. This strict, unyielding enzymatic roadblock powerfully acts as a fundamental functional limit on our endogenous biological synthesis.
Conversely, look closely at the right side of the slide, which perfectly represents the complementary elongation pathway. Unlike restricted terminal desaturation, the systematic addition of newly formed two-carbon units to extend the overall length of the lipid chain is freely permitted and highly active. Because we possess the innate ability to continuously elongate but fundamentally lack the vital ability to desaturate past C-9, we inevitably arrive at the Essentiality Rule. This unbreakable rule dictates that specific polyunsaturated structures—those that absolutely require double bonds deep within their terminal tails—cannot be manufactured internally. Therefore, these precise, structurally complex Fatty Acids must be entirely sourced from the human diet.
Slide 7: From Essential Fatty Acids to Critical Precursors: Synthesizing Arachidonic Acid
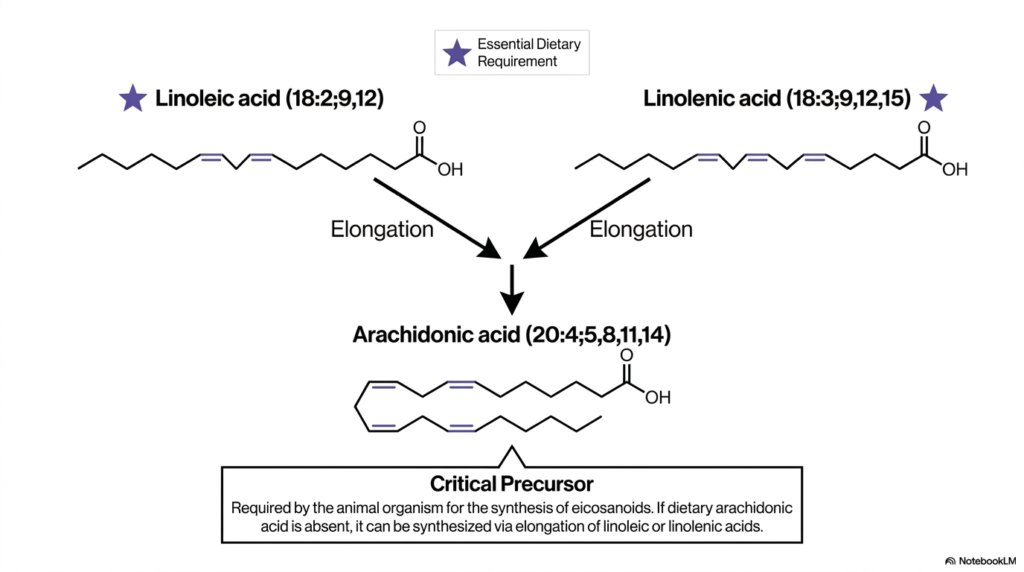
Now that we so deeply understand our inherent biochemical limitations, we must logically ask: how do we actively use the essential nutrients we consume to build the incredibly complex functional signaling molecules our bodies desperately require? Vital cellular signaling, robust immune responses, and highly regulated inflammation pathways are absolutely dependent on highly specific, modified lipid derivatives. The core purpose of this slide is to beautifully trace the highly precise biochemical assembly line that transforms ingested essential dietary lipids into critical functional precursors, using the permitted elongation pathways we previously examined.
At the very top of our intricately detailed diagram, we clearly see two prominently star-marked molecules functionally representing our mandatory essential dietary requirements: linoleic acid (18:2;9,12) and linolenic acid (18:3;9,12,15). As we comprehensively learned from the Essentiality Rule, both of these highly unsaturated Fatty Acids natively contain critical terminal double bonds extending well past the restricted C-9 position, meaning they absolutely must be absorbed directly from our daily food. However, these specific molecules are not merely burned for raw metabolic energy; they act as highly critical biological starting materials. The diagram explicitly features bold downward arrows strongly indicating the highly active elongation pathway.
Through an incredibly complex, highly regulated series of biological chain-lengthening and specific modification steps, these essential dietary lipids are sequentially enzymatically converted into a significantly larger, highly specialized molecule: arachidonic acid (20:4;5,8,11,14). Notice the deeply impressive, heavily kinked molecular structure of arachidonic acid, prominently featuring four distinct cis double bonds. This specific molecule is universally categorized across medical literature as an absolutely critical precursor. If pre-formed dietary arachidonic acid is functionally absent, the biological organism must actively synthesize it internally via the systematic elongation of ingested linoleic or linolenic Fatty Acids.
Slide 8: The Glycerol Scaffolding: The Foundational Anchor Point for Fatty Acids
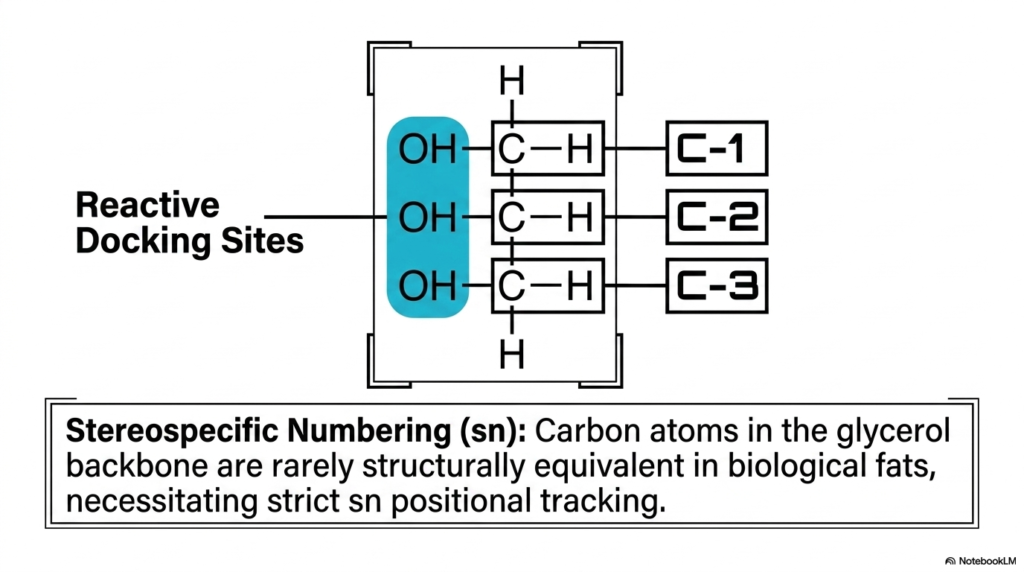
If we consider individual lipids to be the basic structural, high-energy bricks of cellular energy and complex biological membranes, then we must logically ask: what exactly serves as the vital mortar that securely holds them together? Free lipids, aggressively floating unattached around the delicate cell, can be highly volatile and physically act as destructive detergents on cellular membranes. To safely and effectively harness their immense latent energy, biology desperately requires a highly stable, nonreactive framework. The core purpose of this exact slide is to formally introduce the fundamental glycerol backbone, the highly stable biochemical scaffolding that permanently docks multiple Fatty Acids together.
Let us meticulously examine the primary molecular diagram, perfectly centered on the slide. Here, we clearly observe glycerol, a structurally relatively simple three-carbon biological molecule. Securely attached to each of these three central carbons (prominently labeled C-1, C-2, and C-3) is a distinct oxygen-hydrogen pairing. The slide specifically and boldly highlights these three crucial hydroxyl groups in bright blue, labeling them perfectly as the absolutely critical Reactive Docking Sites. These exact microscopic chemical zones represent the precise physical locations where highly reactive incoming free acyl residues will ultimately bind during biological assembly.
However, the complex structural biology of this seemingly simple backbone goes far beyond merely providing a basic attachment point. The descriptive text located below boldly introduces a vital advanced biochemical concept universally known as Stereospecific Numbering (sn). In the highly dynamic, constantly shifting three-dimensional environment of a living biological cell, geometric orientations are never simply arbitrary. Because the various carbon atoms within the core glycerol backbone are rarely structurally functionally equivalent once diverse Fatty Acids begin permanently binding to them, precise biochemists absolutely must accurately track their respective positions.
Slide 9: Assembly Step 1: The Initial Esterification of Fatty Acids
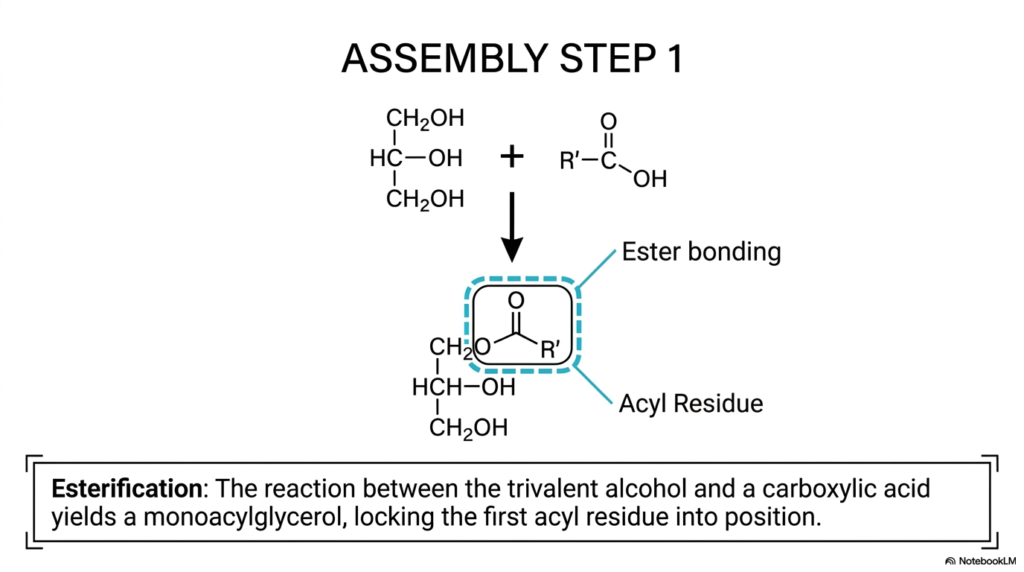
We have now thoroughly and systematically explored both the freely floating individual lipid chains and the completely bare structural glycerol scaffolding; it is finally time to witness the biochemical construction process in full action. How exactly does a living biological cell actively lock a highly reactive, completely free lipid permanently onto a safe storage backbone? Complex biochemical assembly inherently requires highly precise, energetically highly favorable, distinct chemical reactions. The core purpose of this specific slide is to clearly and deliberately outline Assembly Step 1, deeply detailing the fundamental biological condensation reaction—formally termed esterification—that irreversibly binds free Fatty Acids directly to the waiting glycerol framework.
Looking very closely at the balanced chemical equation proudly presented on the upper portion of the slide, we can easily and correctly identify the specific raw materials precisely positioned on the left side: a reactive trivalent alcohol (representing the core glycerol molecule featuring its three highly reactive hydroxyl groups) directly reacting with a generic carboxylic acid (effectively representing our incoming free lipid chain, which is denoted universally by the generic R’ chemical group). The bold, prominent downward arrow distinctly indicates the active biochemical transformation systematically mediated by powerful cellular enzymes. Through the highly precise biological process of esterification, a strong, permanent chemical bond is seamlessly forged.
The newly formed molecular structure, securely enclosed within the highly visible blue dashed box, clearly highlights two major functional features. First, it boldly points out the newly formed ester-bonding structural site. This highly stable covalent bond securely anchors the incoming molecule in place. Second, the lipid chain itself is now formally and permanently referred to as an Acyl Residue, clearly indicating that it is no longer a free, highly reactive acid. This exact initial condensation reaction successfully yields a stable monoacylglycerol, securely locking the very first of the complex Fatty Acids into position.
Slide 10: The Emergence of Molecular Asymmetry: Chiral Centers in Complex Fatty Acids
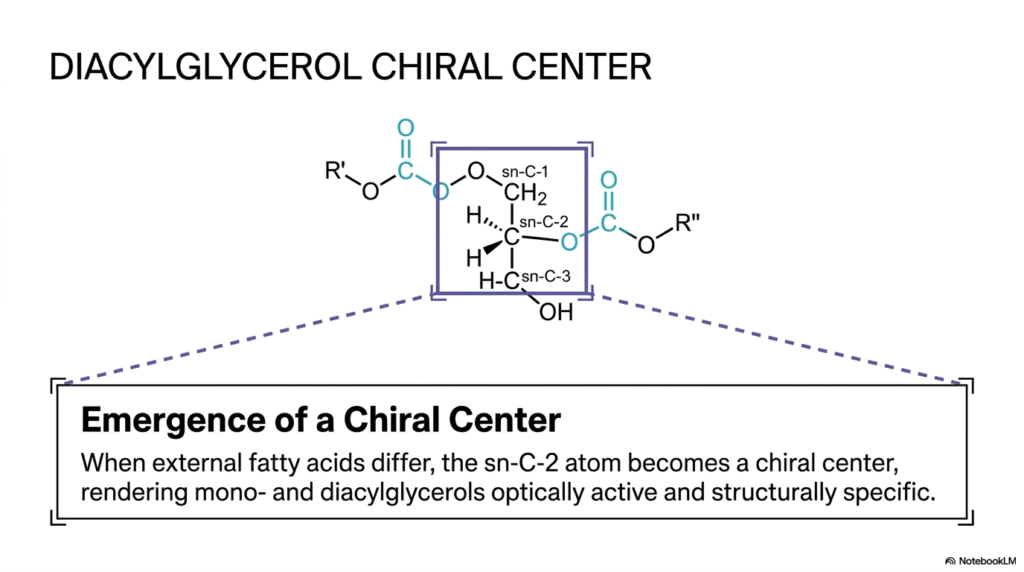
In the invisible, microscopic, flawlessly regulated inner world of advanced cellular biochemistry, precise molecular symmetry—or the distinct lack thereof—fundamentally dictates exactly how biological enzymes interact with their chemical substrates. A specific molecule’s literal three-dimensional “handedness” can entirely change its biological destiny and its subsequent metabolic processing. As we actively continue to build our increasingly complex biological lipids, a highly intriguing, unavoidable spatial phenomenon inevitably arises. The core purpose of this particular slide is to deeply explain exactly how the physical addition of highly diverse Fatty Acids to a perfectly simple glycerol backbone spontaneously generates permanent molecular asymmetry.
Take an incredibly close, highly analytical look at the detailed, specific molecular structure of the intermediate diacylglycerol, prominently shown directly at the center of the diagram. We can clearly and definitively identify the central glycerol backbone, with precise carbon positions at sn-C-1, sn-C-2, and sn-C-3. At this highly specific intermediate stage of complex biochemical assembly, two completely distinct acyl residues (formally labeled R’ and R”) have already been successfully esterified to the primary scaffolding. However, pay highly specific, incredibly careful functional attention to the exact central carbon atom, boldly labeled sn-C-2.
The informative slide carefully outlines an absolutely critical, unbreakable rule of advanced lipid stereochemistry: precisely when the external functional acyl chains, actively bound to the structural backbone, physically differ from one another, the specific central sn-C-2 atom instantly and permanently transforms into a true biological chiral center. This newly formed chiral center renders the previously symmetrical molecule completely optically active and highly stereospecific in three-dimensional space. By systematically arranging functionally distinct Fatty Acids around this brand-new chiral center, the intelligent biological cell ensures that highly specific downstream metabolic enzymes can flawlessly recognize and precisely interact with the biologically correct structural isomer.
Slide 11: The Ultimate Energy Reservoir: Storing Fatty Acids as Neutral Fats
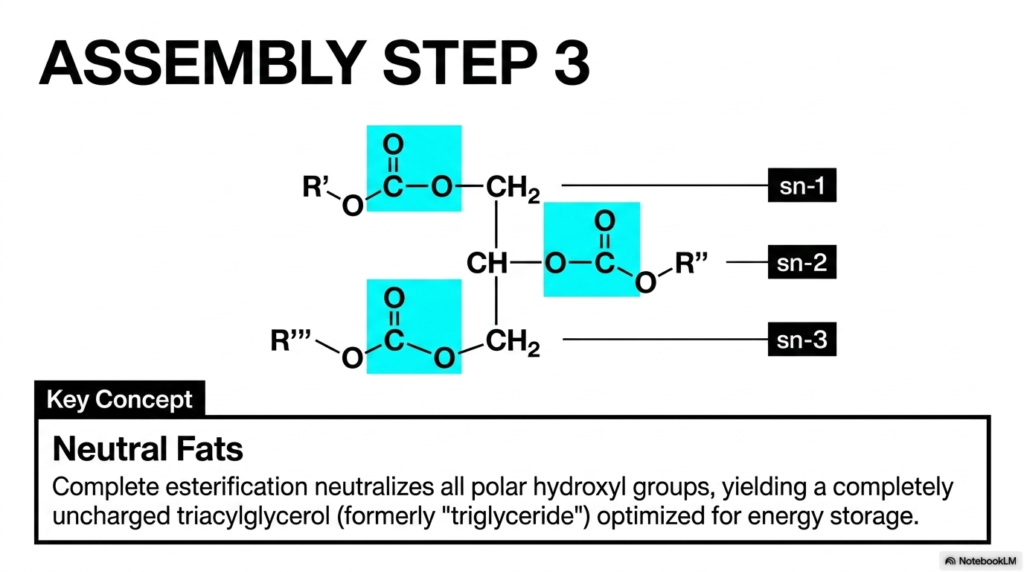
Why exactly doesn’t the unbelievably massive amount of pure fat safely stored continuously within our human bodies simply instantly dissolve outward into the highly watery, fluid environment of our circulating blood and cellular cytoplasm? To successfully and safely store truly vast internal reserves of metabolic energy over extensively long temporal periods, highly evolved biological systems absolutely must completely chemically neutralize the innate, destructive reactivity of their stored energy molecules. The primary purpose of this beautifully illustrated slide is to visually demonstrate Assembly Step 3, the final stage of comprehensive lipid construction that yields the highly stable triacylglycerol.
Carefully observe the completely assembled, highly massive molecular structure heavily dominating the precise center of the informational slide. All three potentially available chemical docking sites located permanently on the primary glycerol backbone (labeled highly clearly as sn-1, sn-2, and sn-3) have now been successfully and completely fully occupied by three entirely distinct functional acyl residues, boldly marked conceptually as R’, R”, and R”’. The extremely bright blue-highlighted boxes purposefully draw our immediate, functional attention to the three incredibly stable covalent ester bonds that have now permanently locked these chains perfectly into place.
The highly critical overarching biochemical core concept explained thoroughly here is the biological formation of what are termed Neutral Fats. Before undergoing this complete, systematic esterification process, the freely floating individual Fatty Acids natively possessed highly polar, biologically reactive carboxylic acid heads. However, the precise biological process of complete, three-part esterification fundamentally permanently alters their underlying chemical reactivity. By successfully and permanently locking a unique lipid chain into every available slot, the biological condensation reaction effectively neutralizes all polar hydroxyl groups. The triumphant end biological result is a completely uncharged, totally nonpolar, massive structural molecule, fundamentally optimized exclusively for the dense energy storage of Fatty Acids.
Slide 12: Combinatorial Diversity and the Positional Preference of Fatty Acids
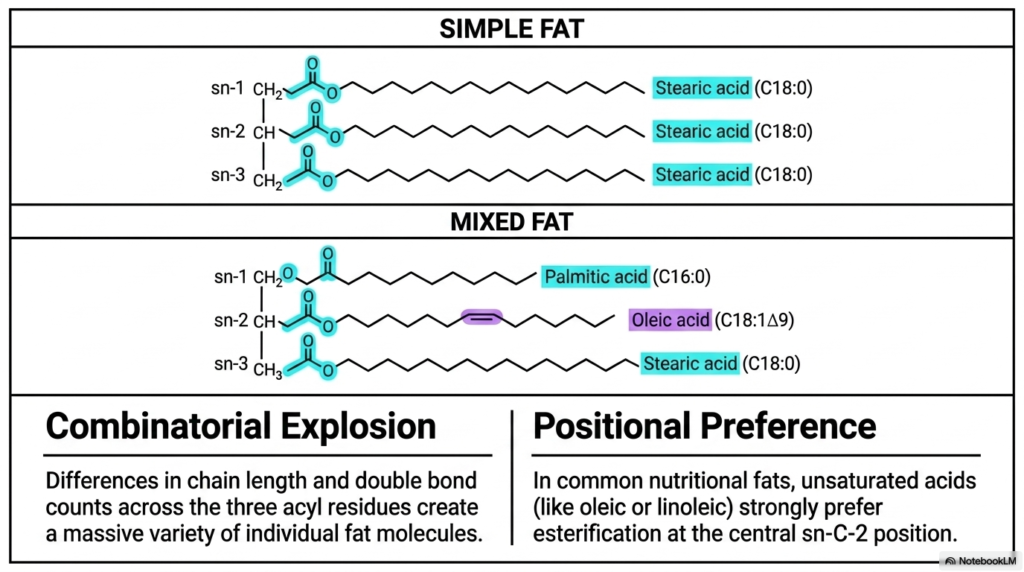
If you were to microscopically look deeply into the dense fat tissue of a complex human, a grazing cow, or even an actively growing plant, you definitively would not easily find just one highly uniform, completely perfectly replicated type of simple triglyceride repeated endlessly; rather, you would uncover a dizzying, incredibly complex array of potentially millions of highly unique, distinct molecular structural variations. How exactly does intelligent biological nature physically generate such truly massive structural diversity entirely from just a very small, limited handful of basic fundamental building blocks? The core purpose of this slide is to clearly illustrate both the staggering mathematical structural diversity of biological lipid structures and the strict positional rules governing Fatty Acids.
The highly detailed graphic cleanly contrasts two primary overarching structural categories of diverse biological fats. First, perfectly located in the top visual panel, we definitively have the generic Simple Fat. In this highly specific, visually uniform example, physically identical saturated stearic acid chains fully and uniformly occupy all three available sn positions. However, these perfectly uniform simple fats are exceedingly rare in biology. The dynamic visual panel, physically positioned immediately below it, strongly highlights the vastly more functionally common Mixed Fat, precisely where three entirely structurally different acyl residues uniformly occupy the three central docking sites. Because biology features mathematically endless physical variations, this instantly and forcefully creates a truly massive Combinatorial Explosion.
Yet, despite this truly vast underlying mathematical-statistical potential for highly random, chaotic biological assembly, our highly evolved cellular enzymes do not blindly attach individual lipids chaotically. The informative right visual panel powerfully highlights the unbreakable, strict biological rule known universally as Positional Preference. In our highly common daily nutritional fats and complex cellular lipid membranes, heavily unsaturated chains consistently exhibit a remarkably strong, enzymatically intensely driven distinct preference for secure esterification precisely at the absolute central sn-C-2 position. By carefully and strictly regulating exactly where specific Fatty Acids go, the body precisely tunes its lipids.
Slide 13: Micro-Structure Dictates Macro-State: The Physical State of Fatty Acids
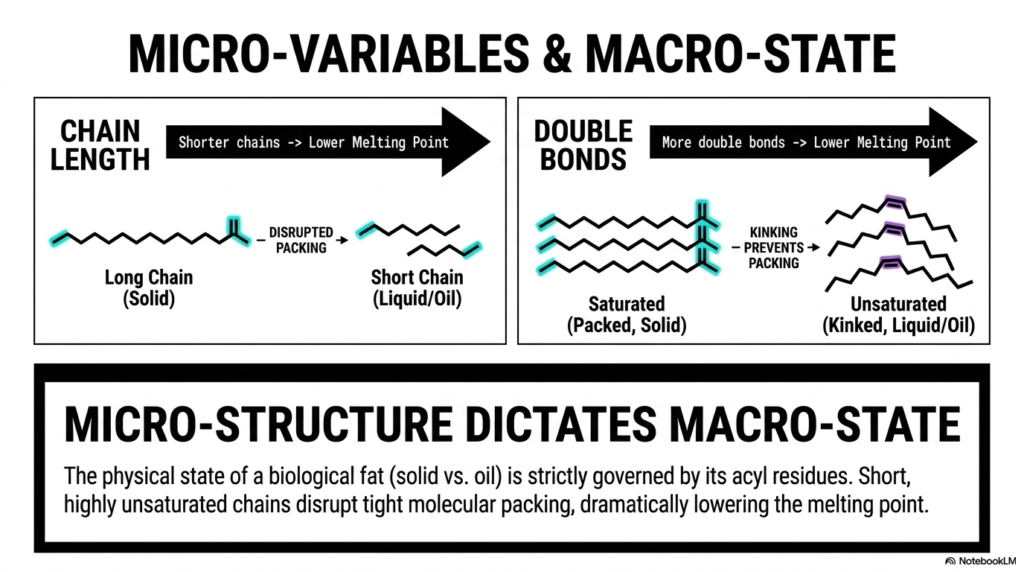
Have you ever stared at a solid stick of butter and a bottle of flowing liquid olive oil and wondered what truly separates them at a microscopic level? They are both composed entirely of dietary lipids, yet their visible macro-states are completely opposed. The core purpose of this powerful slide is to finally bridge the massive conceptual gap between invisible molecular geometry and highly visible physical properties, proving that the melting point of a biological fat is strictly governed by the specific architecture of its constituent Fatty Acids.
The slide beautifully breaks this complex concept down into two highly influential structural micro-variables. First, let us closely examine “Chain Length” illustrated on the left side of the diagram. The graphic sharply contrasts a long-chain lipid with a much shorter-chain lipid. The governing physical rule here is elegant yet profoundly impactful: shorter carbon chains intrinsically possess a significantly lower melting point. A long, perfectly uninterrupted hydrocarbon chain has more surface area, allowing it to pack tightly and form a rigid solid via strong intermolecular forces. Conversely, shorter chains structurally disrupt this highly orderly packing, pushing the macroscopic substance toward a fluid, liquid state at room temperature.
The second micro-variable is the undeniable presence of “Double Bonds,” clearly shown on the right side. Here, we see a highly ordered, tightly packed stack of saturated Fatty Acids, boldly labeled “Solid.” Because these fully saturated molecules are perfectly straight, they align flawlessly. Directly next to them, we observe a chaotic, loosely arranged stack of highly unsaturated lipids. As we established earlier, cis double bonds introduce rigid, permanent structural kinks. These distinct kinks actively and forcefully prevent tight molecular packing. Therefore, the unbreakable rule states that more double bonds heavily disrupt packing, yielding a fluid liquid oil. Ultimately, the physical state of any fat is utterly dictated by its exact blend of Fatty Acids.
Slide 14: Conformational Packing: Maximizing the Energy Density of Fatty Acids
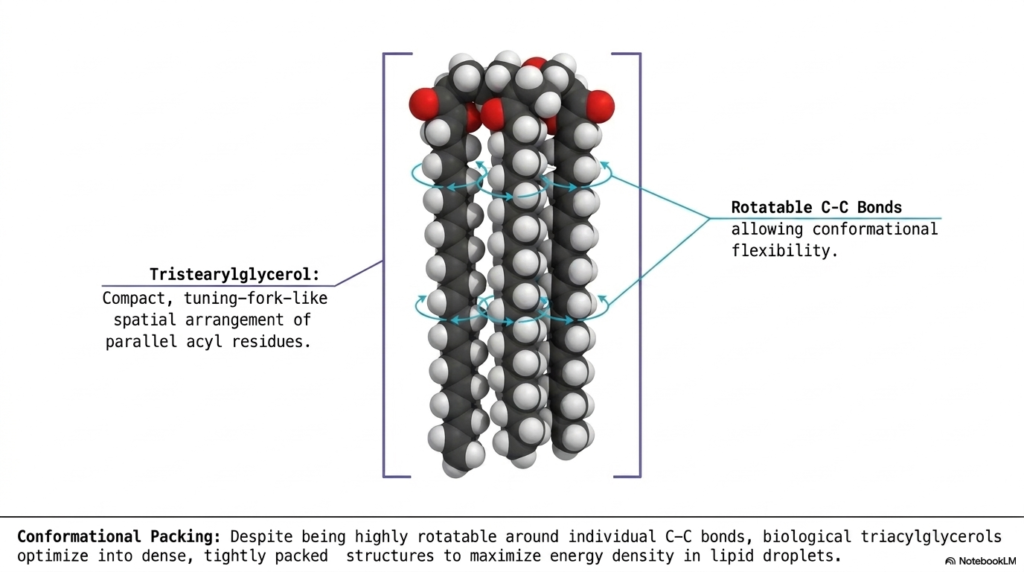
For our final slide in this comprehensive biochemical journey, we must ask the ultimate biological architectural question: how does a living cell actually cram the absolute maximum amount of energetic material into the incredibly microscopic space of a lipid droplet? As we have seen throughout this deck, complex lipids can be massively long, highly unruly, and constantly shifting. To successfully achieve true maximum energy density without rupturing the cell, these dynamic molecules must fold into highly specific, ultra-compact geometric shapes. The core purpose of this concluding slide is to clearly reveal the exact three-dimensional “Conformational Packing” strategies that fully assembled triacylglycerols use to compactly store Fatty Acids.
Observe the striking, highly detailed 3D space-filling molecular model presented squarely in the center of the diagram. This represents tristearylglycerol, a simple fat composed entirely of three straight, fully saturated stearic acid chains. Notice its highly distinct, unbelievably compact shape. Rather than flailing wildly in all possible random directions, the three incredibly long acyl residues align perfectly parallel to one another. This specific geometric folding creates a highly organized structure that biochemists frequently describe as a tuning-fork-like spatial arrangement. It is this exact shape that allows lipid droplets to become incredibly dense.
How do these stiff carbon chains manage to achieve this tight parallel alignment? The diagram astutely points out the crucial role of Rotatable C-C Bonds. The single covalent bonds located between the carbon atoms in saturated Fatty Acids are remarkably flexible, allowing for massive conformational rotation entirely along the hydrocarbon tail. Through the constant rotation of these bonds, the triacylglycerols literally fold themselves, optimizing into extremely dense, tightly packed parallel structures. By actively locking their attached Fatty Acids into this highly organized tuning-fork conformation, organisms can dramatically maximize the energy density stored within their tissues, creating the perfect biological battery.
Please read our Content Disclaimer Statement.
Check out our social media channels: