1. Structural Biochemistry and The Architecture of Life
Welcome, future doctors, biomedical researchers, and science enthusiasts! Have you ever looked at a jar of simple chemicals and wondered what physically separates those lifeless powders from a living, breathing human being? The breathtaking answer lies entirely in the microscopic realm of structural biochemistry. Our goal in this comprehensive guide is to bridge the gap between foundational chemistry and the incredibly complex mechanisms of biological systems. We are going to explore how mere handfuls of elements come together, following strict rules of physics and thermodynamics, to form the exact molecular structures that power every single heartbeat, thought, and metabolic pathway in your body. Let’s dive into the structural biochemistry behind our cellular architecture and decode the absolute foundations of life!
Check out our TikTok Slideshow for this lecture.
Slide 1: Introduction to Biochemistry’s Structural Foundations
Welcome to the fascinating intersection where foundational chemistry becomes biology. Biochemistry isn’t just about memorizing endless metabolic pathways for your exams; it is the essential science that explains the literal molecular underpinnings of life itself. When you strip away the incredible complexity of tissues, organs, and cells, every living organism is ultimately governed by atomic interactions. In this opening sequence, we set the stage by recognizing that biology is simply chemistry in action.
Our structural foundations begin with a visual analysis of elements, bonds, and molecular geometry. Think about the sheer scale of this: the physical and chemical behavior of any biomolecule is absolutely dictated by its constitution—the specific types of atoms it contains and precisely how they are bonded together in three-dimensional space. This slide deck is carefully structured to guide you through these fundamental chemical concepts, which are essential for understanding complex cellular and metabolic processes later on. We will explore everything from orbital hybridization and chemical bonding to the complexities of molecular structure and electron resonance. We will also cover vital noncovalent interactions, such as hydrogen bonds, and examine the diverse forms of isomerism that govern biomolecular function. Whether you are studying medicine, pharmacy, or the biological sciences, grasping these core concepts is an essential first step toward mastering the physical and chemical behavior of life’s molecules, setting the stage for advanced pharmacology and pathology.
Slide 2: The Elements of Life and Biological Abundance

Take a quick look at the periodic table of elements. While there are dozens of stable elements found in nature, it is incredibly surprising to learn that over 99% of animal body mass is constructed from just four primary macroelements: Hydrogen, Oxygen, Carbon, and Nitrogen. Hydrogen and oxygen alone combine to form water, which constitutes a massive 60 to 70% of a cell’s total mass. Together with carbon and nitrogen, they form the reactive core of all organic biomolecules—like the proteins, lipids, and nucleic acids necessary for life.
But human biology isn’t just made of these four! Biological systems rely heavily on a second critical group of elements, primarily inorganic ions, which make up about 0.5% of body mass. These include alkali metals such as Sodium (Na) and Potassium (K), alkaline earth metals such as Magnesium (Mg) and Calcium (Ca), and halogens such as Chlorine (Cl). These are universally ionized in cells and absolutely vital for nerve action potentials, muscle contraction, and fluid balance. Finally, living things require trace elements at the microscopic tip of the biological pyramid. Transition metals such as Iron (Fe), Zinc (Zn), Copper (Cu), Cobalt (Co), and Manganese (Mn), along with nonmetals like Iodine (I) and Selenium (Se), are fundamentally important. For instance, iron is the essential core of the oxygen-transport molecule hemoglobin. Deficiencies in these specific trace elements lead to profound, systemic human diseases such as anemia and metabolic dysfunction.
Slide 3: Electron Configurations and the Octet Rule
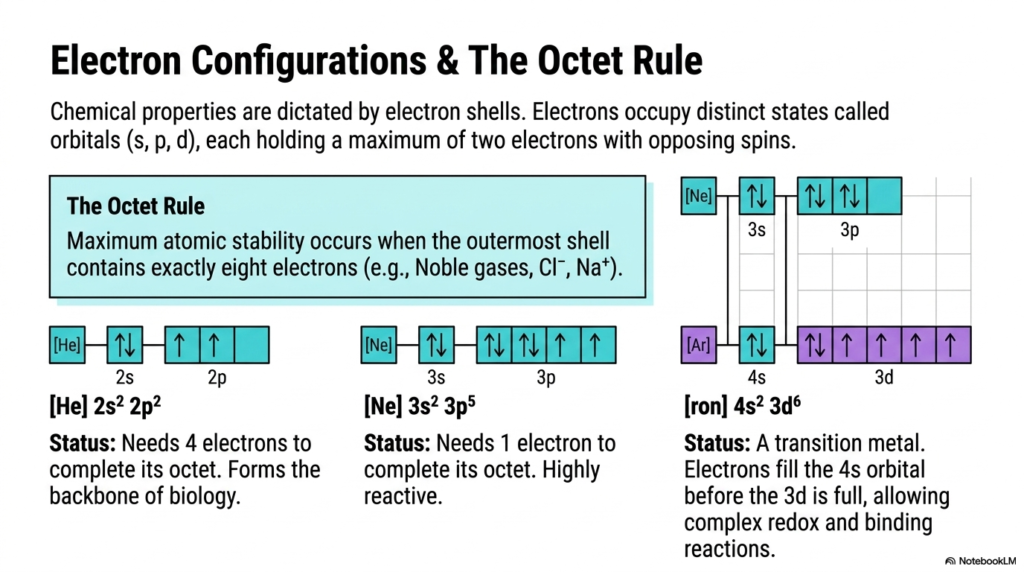
To truly understand how complex biomolecules form, we have to zoom way in on the electrons. The chemical properties of atoms—and the specific types of bonds they form—are strictly dictated by their electron shells. Electrons don’t just orbit the atomic nucleus randomly; they occupy distinct, mathematically defined spatial states known as orbitals (labeled s, p, and d). As atoms grow larger, these orbitals fill sequentially, with each individual orbital holding a maximum of two electrons that must have opposing spins.
The golden rule of chemical bonding that drives electron-seeking behavior is the “Octet Rule.” Maximum atomic stability occurs when an atom’s outermost shell contains exactly eight electrons, perfectly mimicking the highly stable, unreactive configuration of noble gases like Neon or Argon, or ions like Cl⁻ and Na⁺. Take Carbon (C), for example: its electron configuration is [He] 2s² 2p². It needs exactly 4 more electrons to complete its octet, driving it to form four covalent bonds and making it the ultimate backbone of biology. Highly reactive Chlorine (Cl) only needs 1 electron to complete its octet. Meanwhile, transition metals like Iron (Fe) break standard intuitive rules: their electrons fill the 4s orbital before the 3d orbitals are full. This unique electron configuration allows transition metals to facilitate highly complex redox and binding reactions, making them indispensable in medical biotechnology and the catalytic centers of our metabolic enzymes.
Slide 4: Orbital Hybridization and 3D Molecular Geometry
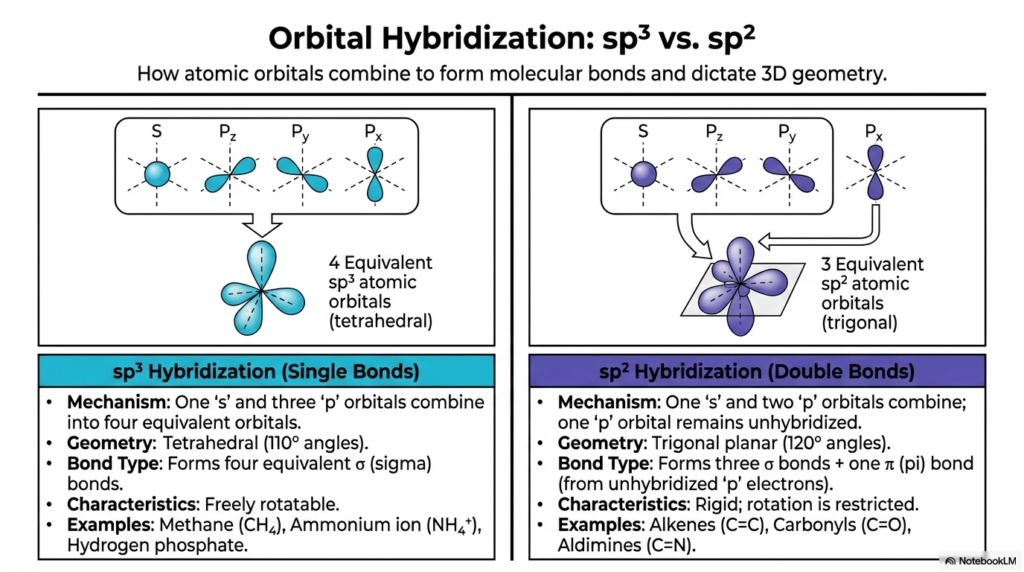
Now, let’s talk about how these atoms actually connect to build cellular structures. Stable covalent bonds form when atomic orbitals combine. A pivotal concept in organic biochemistry is orbital hybridization. Taking Carbon as a prime example, its spherical ‘s’ orbital and three ‘p’ orbitals can mix together to form four mathematically equivalent “sp³” hybrid orbitals. These sp³ orbitals physically arrange themselves in a 3D tetrahedral geometry with 110-degree angles, creating four equivalent sigma (σ) bonds. This exact sp³ hybridization allows for free rotation and defines the molecular geometry of methane, the ammonium ion, and hydrogen phosphate.
Not all molecules form purely tetrahedral structures. A second vital type of hybridization, sp², occurs when an ‘s’ orbital mixes with only two ‘p’ orbitals, leaving one ‘p’ orbital completely unhybridized. The three resulting sp² orbitals lie perfectly flat in a trigonal planar geometry at 120-degree angles, forming standard sigma bonds. The remaining unhybridized ‘p’ orbital forms a pi (π) bond. A sigma bond combined with a pi bond forms a double bond! These double bonds—such as the alkenes (C=C), carbonyls (C=O), and aldimines (C=N)—are fundamentally rigid, and rotation is severely restricted. Without these two distinct types of hybridization dictating bond angles and rigidity, the complex, three-dimensional shapes of your cellular proteins and genetic DNA simply couldn’t exist, making these atomic properties the foundational blueprint for all pharmacological drug design.
Slide 5: Resonance and Delocalized Electron Networks
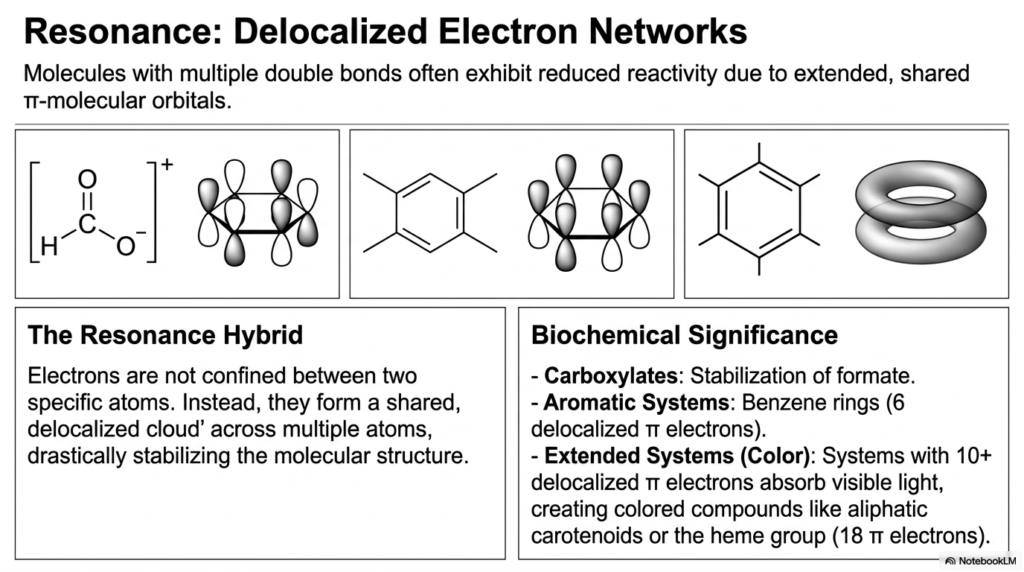
In many critical biomolecules that contain multiple double bonds, the behavior of electrons is far more fluid and dynamic than our simple structural formulas might suggest. Rather than being strictly confined to a single pi bond between two specific, adjacent atoms, pi electrons can actually be shared across an extended, overlapping electron cloud that encompasses multiple atoms at once. This incredible quantum phenomenon is known as resonance, and it drastically stabilizes the molecular structure.
Molecules exhibiting this widespread electron delocalization are known as resonance hybrids. Because they have extended, shared pi-molecular orbitals, they often exhibit massively reduced reactivity. Excellent biochemical examples of resonance stabilization include carboxylate groups (e.g., formate) and aromatic ring systems (e.g., benzene rings, which utilize 6 delocalized pi electrons), found in essential amino acids. Interestingly, when a resonance system is highly extended—typically containing 10 or more delocalized pi electrons—it can actually absorb light within the visible spectrum! This fascinating physical property gives rise to highly colored biological compounds, such as the vibrant aliphatic carotenoids found in plants and the heme group (featuring 18 pi electrons) that carries oxygen in your blood. These delocalized networks are critically important for human survival, safely managing high-energy electrons during cellular respiration and protecting our tissues from oxidative damage.
Slide 6: Modeling Molecular Structure for Reactivity and Shape
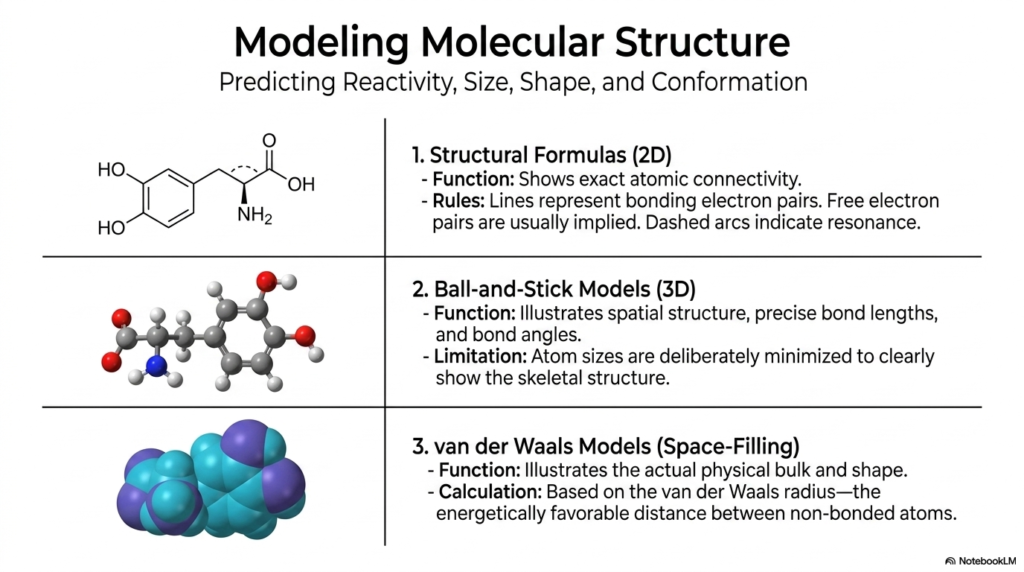
To fully grasp biochemical mechanisms and pharmacology, you have to be able to accurately predict reactivity, size, shape, and conformation. Biochemists utilize three distinct types of molecular models to understand these microscopic structures. First, traditional two-dimensional Structural Formulas are highly useful for mapping exact atomic connectivity. In these drawings, lines represent bonding electron pairs, free electron pairs are usually implied, and dashed arcs indicate the resonance we just discussed.
However, biology operates exclusively in 3D. To understand spatial orientation, Ball-and-Stick Models are heavily employed. These models illustrate exact spatial structures, providing precise bond lengths and bond angles. While highly educational, the atom sizes are deliberately minimized to clearly show the skeletal structure. Finally, to appreciate the true physical volume and shape of a molecule, we rely on van der Waals Space-Filling Models. These models represent atoms as intersecting spheres based on their van der Waals radius—the energetically most favorable distance between non-bonded atoms. In modern medicine and biotechnology, utilizing these sophisticated 3D space-filling models allows researchers to visualize exactly how a therapeutic compound might physically wedge itself into the active site of a viral enzyme, which is the exact process used to develop life-saving antiviral medications!
Slide 7: Electronegativity and the Power of Bond Polarity
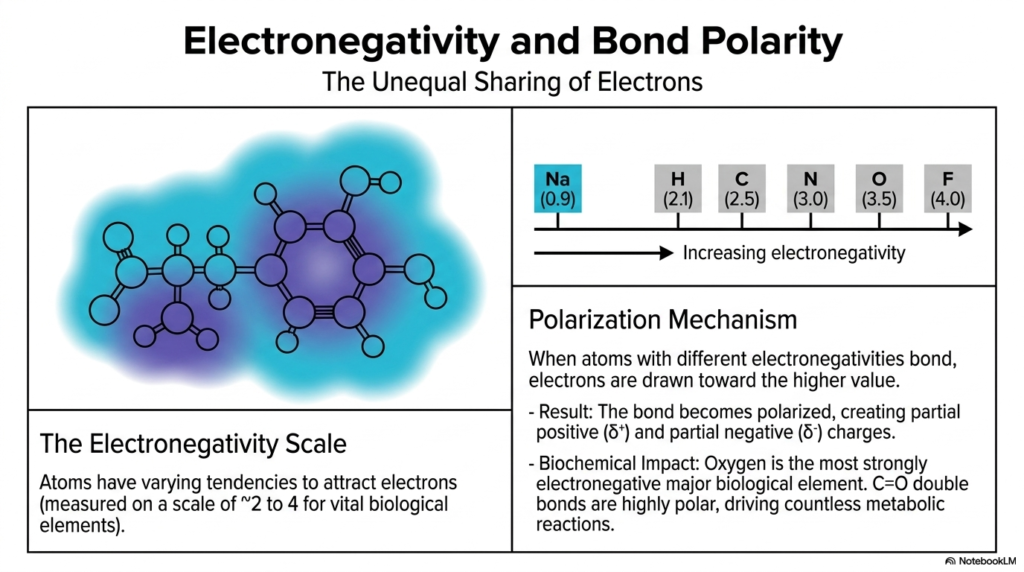
Let’s talk about molecular magnetism and why certain molecules dissolve perfectly in your bloodstream. A critical factor in molecular chemistry is electronegativity, the intrinsic tendency of an atom to attract electrons. Different elements possess varying tendencies to attract electrons, measured on a scale of roughly 2 to 4 for vital biological elements. For example, Sodium (Na) is very low at 0.9, Carbon (C) is 2.5, Nitrogen (N) is 3.0, and Oxygen (O) sits very high at 3.5.
When two atoms with different electronegativities form a bond, the polarization mechanism kicks in. The shared electrons are fiercely drawn toward the higher value. This unequal sharing means the bond becomes polarized, creating a partial positive (δ⁺) and a partial negative (δ⁻) charge across the molecule. The biochemical impact of this is absolutely staggering. Oxygen is the most strongly electronegative major biological element. When it forms a carbonyl group (C=O) with carbon, the bond is highly polar. These partial charges strictly determine how specific regions of a molecule will attract or repel other molecules. This polarity drives countless metabolic reactions, dictates water solubility, and precisely controls the binding mechanisms between pharmaceutical drugs and human cellular receptors.
Slide 8: Hydrogen Bonds: The Non-Covalent Glue of Biology
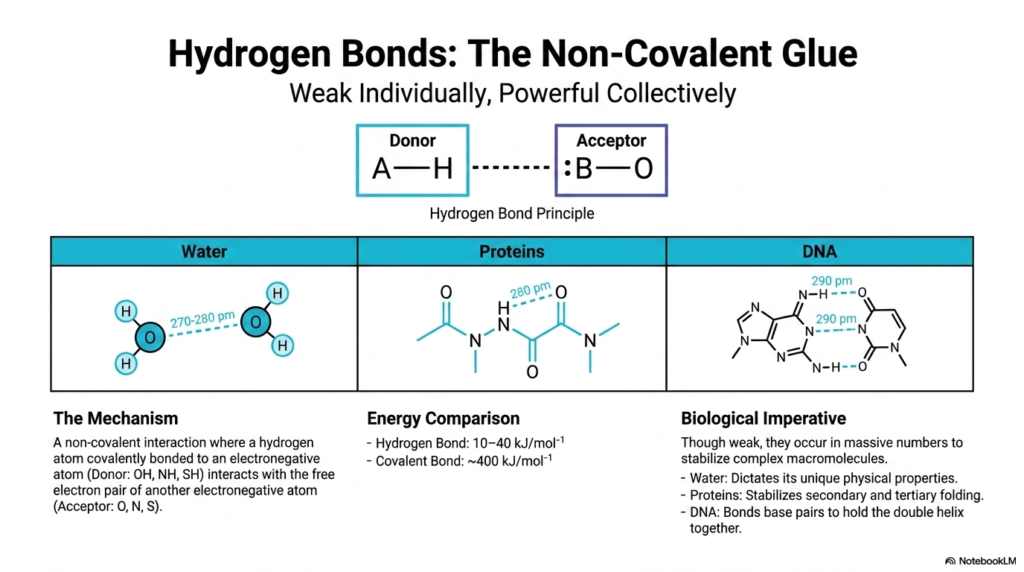
Among all the non-covalent interactions operating in biology, the hydrogen bond is undoubtedly the undisputed heavyweight champion. The hydrogen bond principle is incredibly elegant: it is a non-covalent interaction where a hydrogen atom covalently bonded to a highly electronegative Donor atom (like an OH, NH, or SH group) interacts with the free electron pair of another electronegative Acceptor atom (like Oxygen, Nitrogen, or Sulfur).
Individually, these hydrogen bonds are relatively weak. A quick energy comparison shows that hydrogen bonds have only about 10 to 40 kilojoules per mole of bonding energy, compared to the massive ~400 kilojoules per mole of a strong covalent bond. However, their biological imperative is that they occur in massive, interconnected numbers to collectively stabilize complex macromolecules. The distance between these bonds is tiny, sitting around 270-290 picometers. They dictate the unique physical properties of Water, which makes up most of your body. In Proteins, they form the vital cross-links that stabilize secondary and tertiary folding. In DNA, hydrogen bonds are the exact forces holding the base pairs of the double helix together! Every time a cell in your body divides, it is the precise, manageable strength of these hydrogen bonds that allows your DNA to temporarily unzip, flawlessly replicate, and securely zip back together to prevent genetic diseases.
Slide 9: Geometric Isomerism and the cis-trans Divide
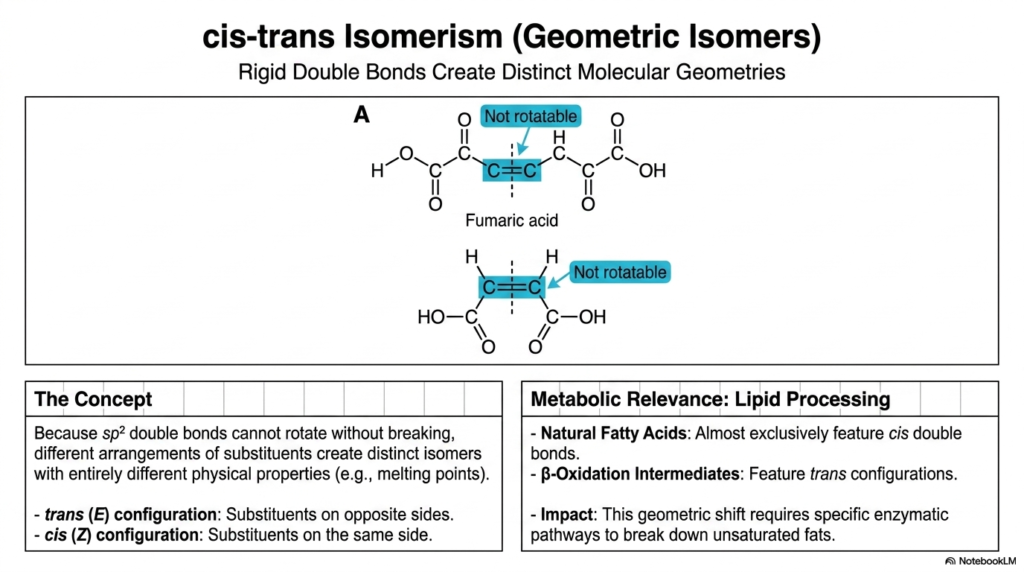
In biochemistry, rigid structures strictly determine metabolic function, leading to cis-trans isomerism, also known as geometric isomerism. This concept arises exclusively around the sp² double bonds we explored earlier. Because these double bonds cannot rotate without completely breaking their pi orbitals, different arrangements of substituents around the bond create distinct isomers with entirely different physical properties, such as melting points.
If the chemical substituents are locked on the opposite sides of the double bond, it is a trans (E) configuration. If they are locked on the same side, it is a cis (Z) configuration. A perfect textbook example is fumaric acid. This spatial distinction has massive metabolic relevance, particularly in lipid processing. Naturally occurring unsaturated fatty acids almost exclusively feature cis double bonds, which introduce a “kink” into the chain and keep cellular membranes fluid and healthy. Conversely, β-oxidation intermediates—the molecules formed when your body breaks down fats for energy—feature trans configurations. This immense geometric shift requires entirely specific enzymatic pathways to process. This is exactly why artificial trans fats in processed foods are so dangerous; they hijack normal lipid metabolism pathways, leading directly to cardiovascular disease.
Slide 10: Conformational Isomerism and Molecular Flexibility
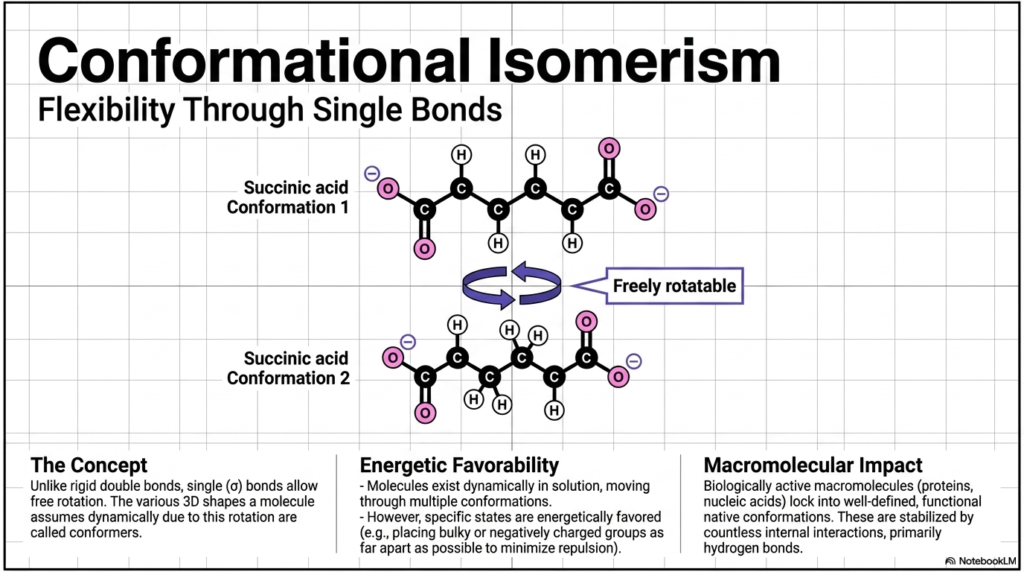
Unlike the highly rigid double bonds of geometric isomers, standard single (σ) covalent bonds allow for continuous, free rotation. This vital flexibility means that a single molecule can dynamically assume various 3D shapes in a cellular solution. These different, temporary spatial forms are known as conformers, and the overall phenomenon is conformational isomerism.
Take a metabolic intermediate, such as succinic acid. It can rapidly twist and turn, adopting multiple conformations (Conformation 1 vs. Conformation 2). However, this molecular gymnastics is governed by energetic favorability. Molecules will naturally spend the vast majority of their time in specific states that are energetically favored—usually by twisting to place bulky or negatively charged groups as far apart as possible to drastically minimize atomic repulsion. This has a profound macromolecular impact. While small molecules flit between shapes, biologically active macromolecules like massive proteins and nucleic acids physically lock into well-defined, functional “native conformations.” These highly specific shapes are stabilized by countless internal interactions, primarily our old friend, the hydrogen bond. If a protein fails to achieve this specific conformation, it misfolds. Protein misfolding is the exact pathological mechanism behind devastating, incurable human conditions like Alzheimer’s and Parkinson’s disease.
Slide 11: Optical Isomerism, Chirality, and Molecular Mirrors
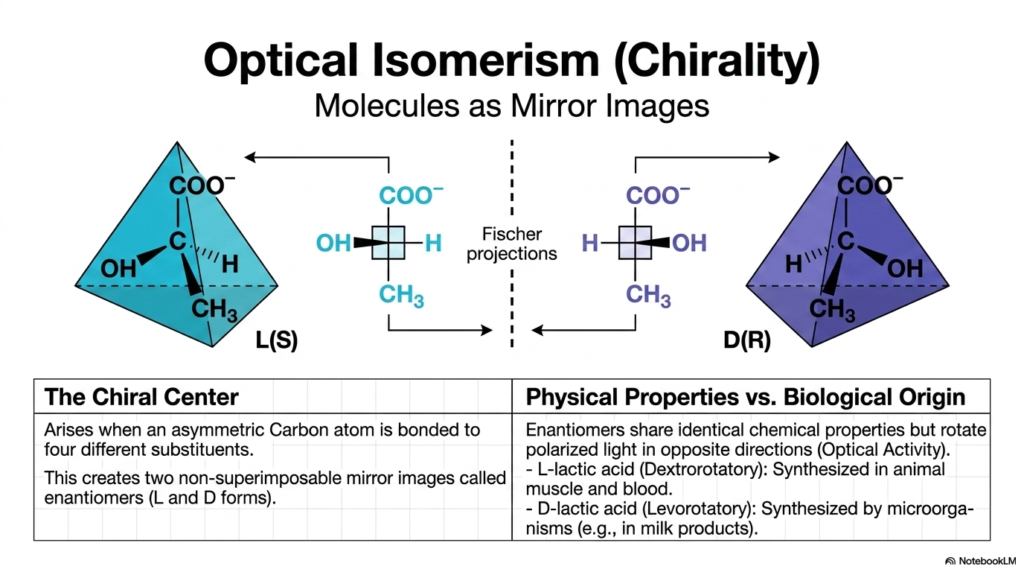
Our exploration of 3D geometry takes a fascinating turn with optical isomerism, deeply rooted in the concept of chirality. The concept of the “chiral center” arises specifically when an asymmetric carbon atom is bonded to four entirely different substituents. This unique asymmetry magically creates two distinct molecular forms called enantiomers, which are completely non-superimposable mirror images of one another. Think of them exactly like your left and right hands!
These enantiomers are typically designated as the L(S) and D(R) forms, and they can be represented using visual tools such as Fischer projections. When considering physical properties versus biological origin, enantiomers exhibit identical standard chemical properties. However, they rotate polarized light in opposite directions (a property called Optical Activity). In biology, chirality is literally a matter of life and death. For example, L-lactic acid is naturally synthesized in human and animal muscle and blood, whereas D-lactic acid is synthesized primarily by microorganisms, including those found in milk products. Human cells recognize and utilize only specific enantiomers, thereby preventing chaotic biochemical cross-reactions. When pharmaceutical companies synthesize drugs, they must rigorously separate the L and D enantiomers, as one mirror image might cure a disease, while its exact mirror counterpart could be highly toxic.
Slide 12: Enzymatic Stereospecificity in Metabolism
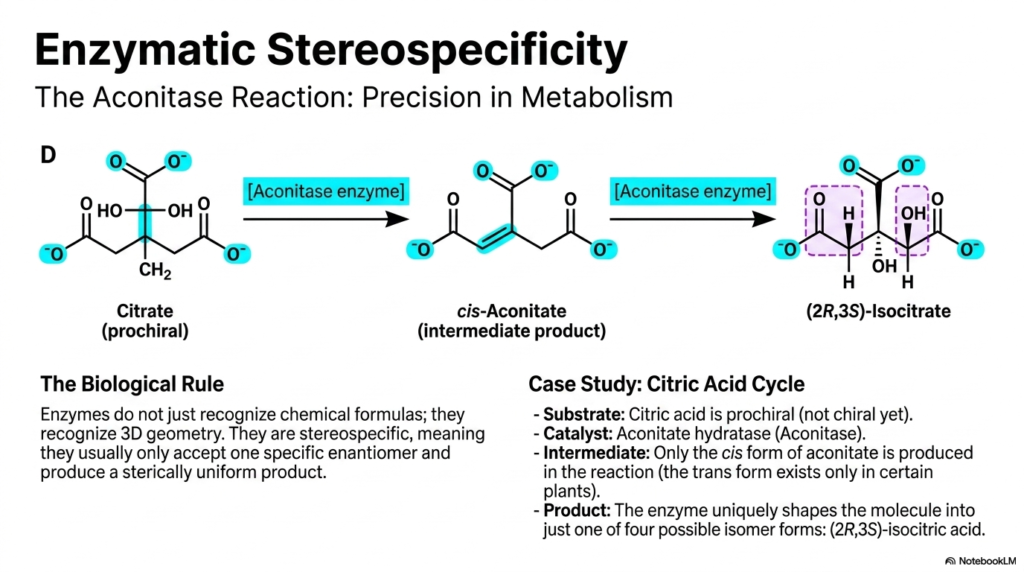
To understand why chirality matters so much, we must examine enzymatic stereospecificity. The biological rule is simple but absolute: enzymes do not just recognize basic chemical formulas; they recognize precise 3D geometry. Enzymes are inherently stereospecific, meaning they usually only accept one specific enantiomer as a substrate and produce a sterically uniform product.
A phenomenal case study is the Aconitase reaction, found deep within the Citric Acid Cycle, the engine of cellular energy. The cycle’s starting substrate, Citrate, is technically prochiral—meaning it is not yet chiral but has the potential to become chiral. The enzyme Aconitase (or aconitate hydratase) binds citrate, forming an intermediate product called cis-aconitate. Of all the potential geometric shapes, only the cis form is produced in human metabolism. From there, the enzyme tightly manipulates the intermediate to uniquely shape the final molecule into just one of four possible isomer forms: exactly (2R,3S)-isocitrate. This jaw-dropping level of metabolic precision ensures that energy production operates flawlessly, without producing wasteful byproducts. In biotechnology, leveraging this strict stereospecificity allows scientists to bioengineer bacteria to produce highly pure, complex medicines that synthetic chemistry simply cannot match.
Slide 13: The Chemical Architecture of Life Summarized
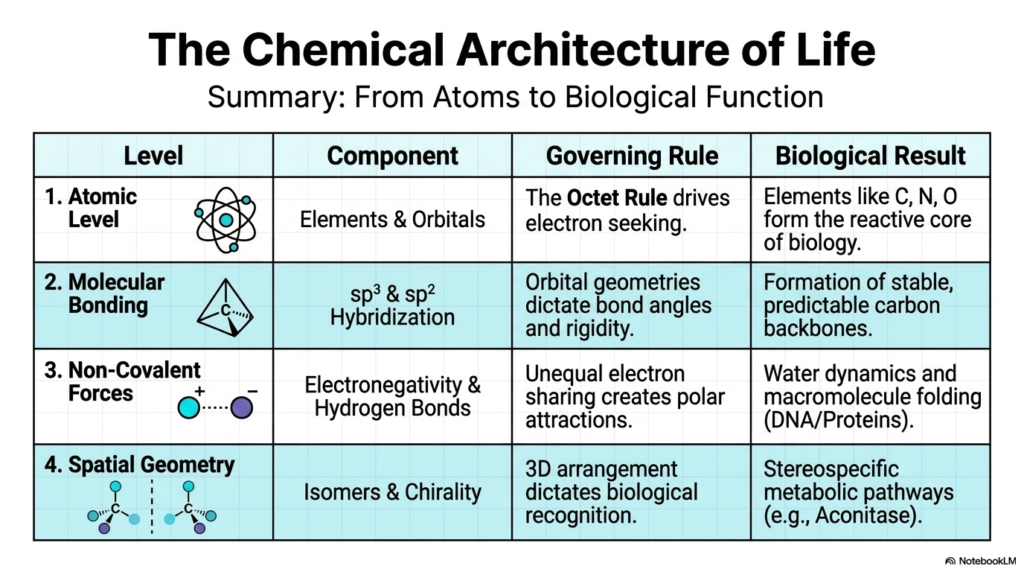
We have reached the grand summary of life’s chemical architecture, mapping the magnificent journey from individual atoms to complex biological functions! Let’s review this incredible structural blueprint.
At the Atomic Level, elements like Carbon, Nitrogen, and Oxygen form the reactive core of biology, with their behavior entirely governed by the electron-seeking drive of the Octet Rule. Moving up to Molecular Bonding, we see how the intricate mixing of sp³ and sp² hybridization dictates bond angles and rigidity, resulting in highly stable, predictable carbon backbones. Once these molecules are formed, Non-Covalent Forces take over. The unequal sharing of electrons due to differences in electronegativity creates polar attractions, particularly Hydrogen Bonds, which govern water dynamics and drive the crucial folding of macromolecules such as DNA and proteins. Finally, Spatial Geometry layers provide the ultimate level of control. The 3D arrangement of atoms via geometric and optical isomers (chirality) dictates biological recognition, driving stereospecific metabolic pathways such as the Aconitase reaction.
Understanding these fundamental structural biochemistry concepts is not just an academic exercise—it is the direct key to modern medicine. By mastering this blueprint, we empower ourselves to design better pharmaceutical drugs, cure genetic diseases, and truly comprehend the absolute beauty of biological design!
Check out our social media channels:
Please read our Content Disclaimer Statement.