7. Biological Redox Reactions: The Molecular Ledger
Have you ever wondered what actually happens at the microscopic level when you drink a cup of coffee or eat a carbohydrate-rich meal to “get some energy”? As future doctors and scientists, you know it is not just magic—it is a highly orchestrated series of molecular events. The core engine driving all of this is an elegant, invisible accounting system inside your cells. The purpose of this slide deck is to demystify that system by exploring the fundamental mechanics of electron transfer. We are going to break down how cells safely manage unstable energy without spontaneously combusting, turning complex biochemistry into a clear, understandable narrative. Grab your notebooks, and let’s dive in!
Slide 1: An Introduction to Biological Redox Reactions and the Molecular Ledger

Welcome to the foundation of cellular bioenergetics! When we talk about keeping a living organism alive, we are fundamentally talking about Biological Redox Reactions. Think of the cell as a massive, bustling metropolis, and in this city, energy is not exchanged in dollar bills; it is exchanged in the form of electrons. This opening slide introduces the concept of the “Molecular Ledger,” a perfect analogy for how these processes operate. Just as an accountant tracks every penny moving in and out of a bank account to ensure the books balance, cellular biochemistry tracks every single electron.
To truly understand Biological Redox Reactions, you must first realize that nature is an incredibly strict bookkeeper. We are looking at a structural analysis of electron transfers, which means we care deeply about the specific physical pathways these subatomic particles take. The slide highlights three major pillars we will explore: electron transfers, reduction equivalents, and coenzyme mechanics. When you see the term “reduction equivalents,” think of it as the specific currency denomination being handed from one molecule to another. Sometimes it is a single coin, and sometimes it is a bundle of cash.
Furthermore, Biological Redox Reactions do not happen in a vacuum. They are facilitated by complex molecular machines known as coenzymes. These coenzymes are essentially the bank tellers of the cellular world, safely handling the dangerous currency of electrons. As medical and biology students, mastering this concept of the ledger early on will make everything from cellular respiration to pharmacology infinitely easier to digest. We will see how every biochemical pathway relies on the fact that electrons are never simply lost into the ether; they are intentionally, precisely relocated to drive the machinery of life.
Slide 2: The Paired Nature of Biological Redox Reactions
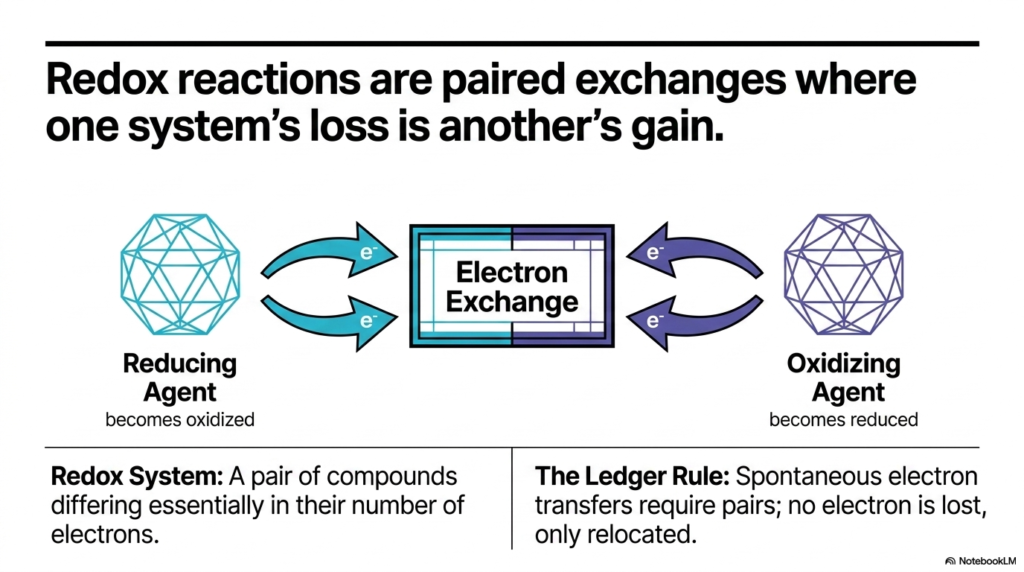
Now that we understand the cell acts as a strict accountant, we need to look at the golden rule of that accounting system: the paired exchange. In the world of Biological Redox Reactions, one molecule’s loss is absolutely always another molecule’s gain. You cannot have an oxidation event without a simultaneous reduction event. This slide brilliantly illustrates this core concept by showing the continuous, complementary flow of electrons between two distinct entities.
Let us break down the terminology that often trips up students. The “Reducing Agent” is the generous donor. It gives away its electrons and, in doing so, becomes oxidized. Conversely, the “Oxidizing Agent” is the eager recipient; it accepts those electrons and becomes reduced. This interplay creates what we call a redox system—a pair of compounds that differ only in the number of electrons they contain. It is crucial to view Biological Redox Reactions not as isolated incidents, but as an intimate molecular dance between these pairs.
This brings us to “The Ledger Rule” highlighted on the slide. Because spontaneous electron transfers require these pairs, no electron is ever truly lost; it is only relocated. This rule is the absolute bedrock of Biological Redox Reactions. If an electron were to simply float away uncoupled, it would wreak havoc on the cellular environment. Therefore, the cell has evolved to ensure that every time an electron leaves a donor, an acceptor molecule is perfectly positioned to catch it. Understanding this paired dependency allows you to predict how metabolic pathways chain together, passing electrons down the line like a molecular bucket brigade to ultimately generate the ATP that keeps us alive.
Slide 3: Redox Potential Dictates the Flow of Biological Redox Reactions
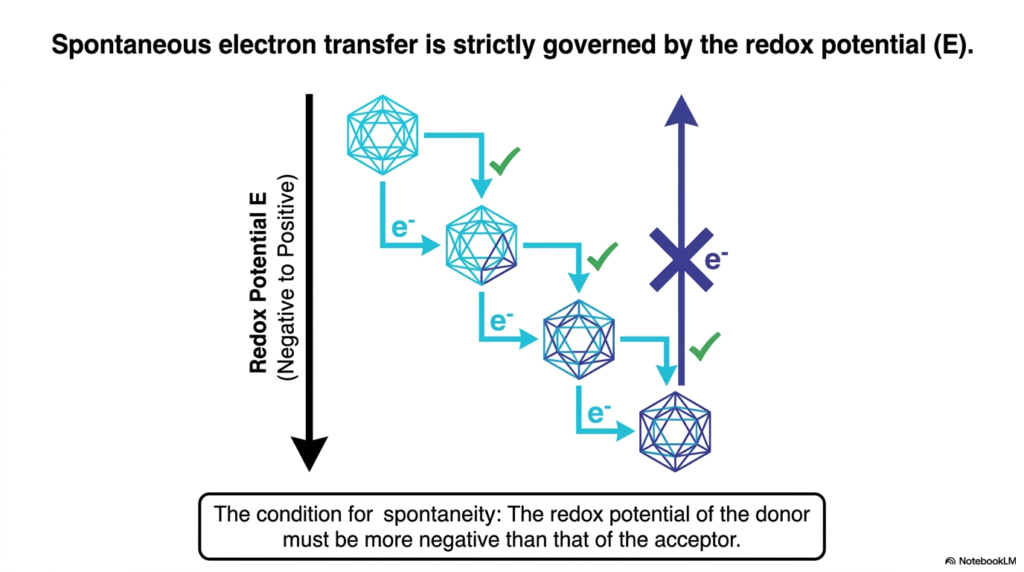
If electrons are the water flowing through the cellular landscape, what determines the direction of the river? The answer is Redox Potential (E). This slide is critical because it explains the directional rules governing all Biological Redox Reactions. Electrons do not just wander randomly; they fall down an energetic gradient. The redox potential is effectively a measure of a molecule’s affinity for electrons.
To make sense of this, picture a staircase. At the top of the staircase are molecules with a highly negative redox potential. These molecules are desperate to give away their electrons; they are excellent electron donors. At the bottom of the staircase are molecules with a positive redox potential. These are the ultimate electron acceptors, pulling electrons toward themselves with immense chemical force. In Biological Redox Reactions, spontaneous electron transfer only happens in one direction: from a more negative redox potential to a more positive one. The slide illustrates this beautifully with a downward arrow, showing successful electron transfers (marked with green checks) cascading down the energetic staircase.
Why is this important for you to know? Because the entire electron transport chain in our mitochondria is based on this exact principle. Biological Redox Reactions are strung together in a specific sequence so that electrons fall step-by-step from NADH all the way down to Oxygen. Notice the blue arrow on the slide marked with an “X”—this shows that an electron cannot spontaneously jump back up the energy gradient to a more negative potential. Without an input of energy, the flow is strictly unidirectional. Grasping this thermodynamic reality helps you understand why cells extract energy in careful, controlled stages rather than one explosive, unusable burst.
Slide 4: Tracking Reduction Equivalents in Biological Redox Reactions
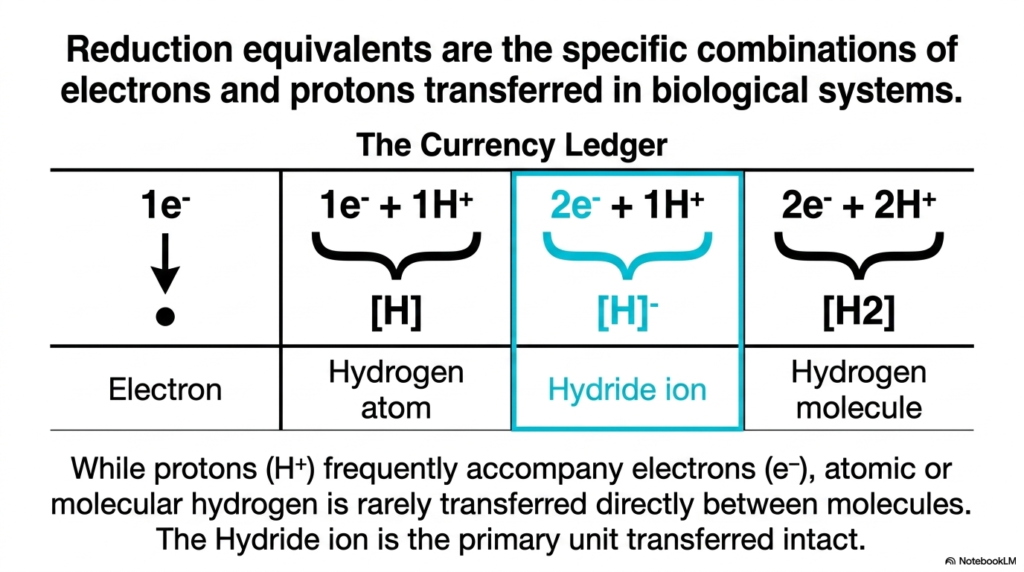
We have established that electrons move down an energy gradient, but how exactly are they packaged in transit? This slide introduces “The Currency Ledger,” breaking down the specific physical forms that electron transfers take. In the study of Biological Redox Reactions, we refer to these packages as “reduction equivalents.” It is rarely just a naked electron flying through the crowded cellular space; the packaging matters immensely for cellular safety and enzyme specificity.
Let us examine the options. The simplest form is a lone electron (e−). Next, we have the hydrogen atom, which is one electron paired with one proton (1e−+1H+). Then, highlighted in blue, we have the Hydride ion (2e−+1H+), and finally, the hydrogen molecule (2e−+2H+). While it is common for protons to accompany electrons in biological systems to maintain charge neutrality, atomic or molecular hydrogen is rarely passed directly between molecules. Instead, as the slide emphasizes, the Hydride ion is the undisputed star of the show.
In many of the most crucial Biological Redox Reactions, the Hydride ion is the primary unit transferred intact. Why is this a game-changer? Because transferring two electrons simultaneously alongside a proton is a highly efficient, chemically safe way to move massive amounts of reducing power. It avoids leaving single, unpaired electrons hanging around in intermediate states. Whenever you look at major metabolic pathways—like the Krebs cycle—and see NADH being produced, you are witnessing the direct transfer of a Hydride ion. Understanding the exact nature of this molecular currency is essential for mastering the mechanics of Biological-Redox Reactions and appreciating how nature elegantly solves the problem of safe energy transport.
Slide 5: Managing the Free Radical Threat in Biological Redox Reactions
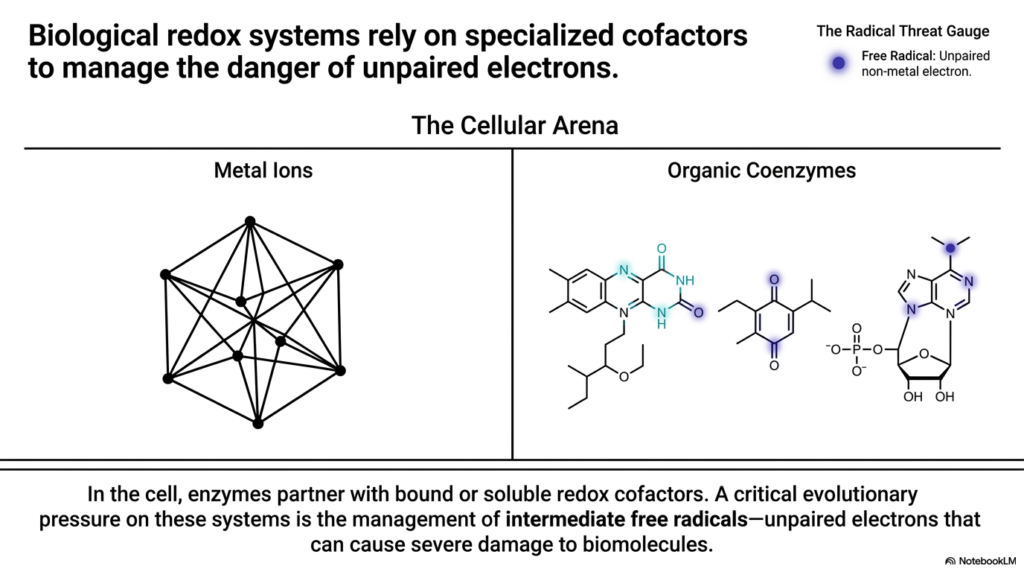
Now we must address the dark side of cellular metabolism: the danger of free radicals. This slide takes us into “The Cellular Arena,” showing how Biological Redox Reactions manage the inherent risks of moving electrons. An electron is essentially a packet of highly reactive energy. If it is not carefully controlled, an unpaired electron (a free radical) can wreak havoc throughout the cell, violently reacting with DNA, proteins, and lipid membranes, causing catastrophic structural damage.
To safely facilitate Biological Redox Reactions, the cell relies on specialized cofactors. You can think of these cofactors as heavily armored transport vehicles. The slide divides these into two main categories: Metal Ions and Organic Coenzymes. Enzymes partner with these bound or soluble cofactors to create safe environments for electron transfer. The “Radical Threat Gauge” on the slide points out the ultimate danger: the unpaired non-metal electron.
A massive evolutionary pressure shaping Biological Redox Reactions is the management of these intermediate free radicals. As electrons are passed from donor to acceptor, there are often intermediate steps where an electron briefly sits alone. If an organic molecule holds this unpaired electron, it becomes highly unstable. The brilliant evolutionary design of our cellular machinery is specifically tailored to hide, stabilize, or instantly pass along these unpaired electrons before they can interact with vulnerable biomolecules. As future clinicians, understanding this slide is your foundation for understanding aging, radiation damage, and the pathology of oxidative stress in human diseases.
Slide 6: How Metal Complexes Safely Navigate Biological Redox Reactions
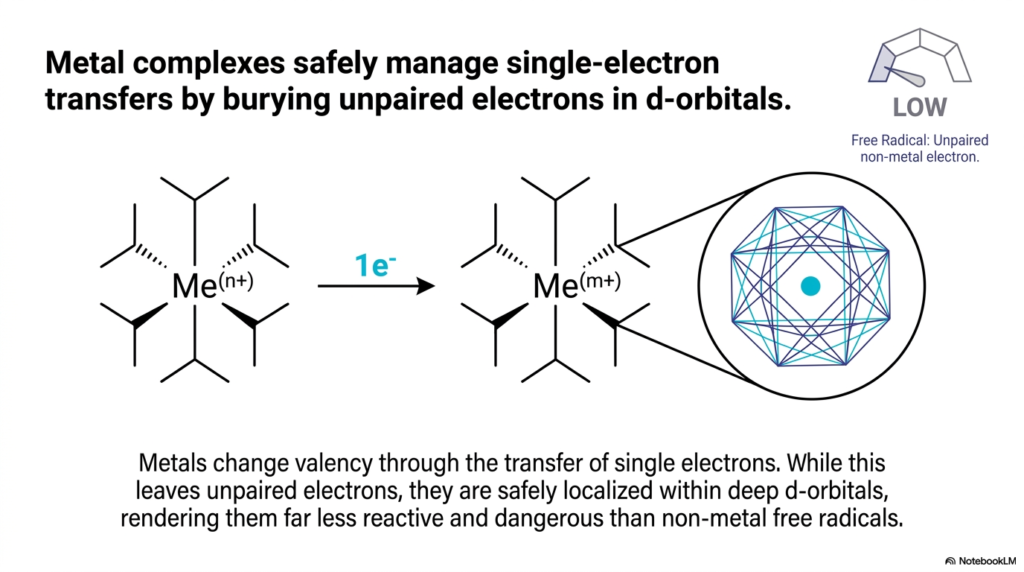
Following our discussion on cellular danger, this slide focuses on one of the cell’s most reliable defense mechanisms: Metal Complexes. In the vast landscape of Biological Redox Reactions, transition metals like iron and copper act as highly stable electron handlers. The slide explains exactly why metals are well-suited for single-electron transfer, earning them a “LOW” rating on the Radical Threat Gauge.
The secret lies in atomic structure. Metals change their valency (their charge) by accepting or donating single electrons. Normally, as we learned, an unpaired electron is a massive liability. However, metal ions possess deep, complex atomic orbitals—specifically, the d-orbitals. When a metal complex participates in Biological Redox Reactions and gains a single, unpaired electron, it buries that electron deep within its d-orbital structure.
This architectural feature is a lifesaver. Because the unpaired electron is safely localized and shielded by the rest of the metal complex’s structure, it becomes far less reactive. It is essentially locked in a molecular vault. This allows metalloproteins (such as the cytochromes in your mitochondria) to safely pass electrons one by one along the electron transport chain. In the context of Biological Redox Reactions, metal ions are the ultimate shock absorbers, accepting volatile single electrons and holding them securely until the next molecule in the chain is ready to receive them, thereby drastically reducing the risk of accidental cellular damage.
Slide 7: The Flavin Two-Step and Protein Binding in Biological Redox Reactions
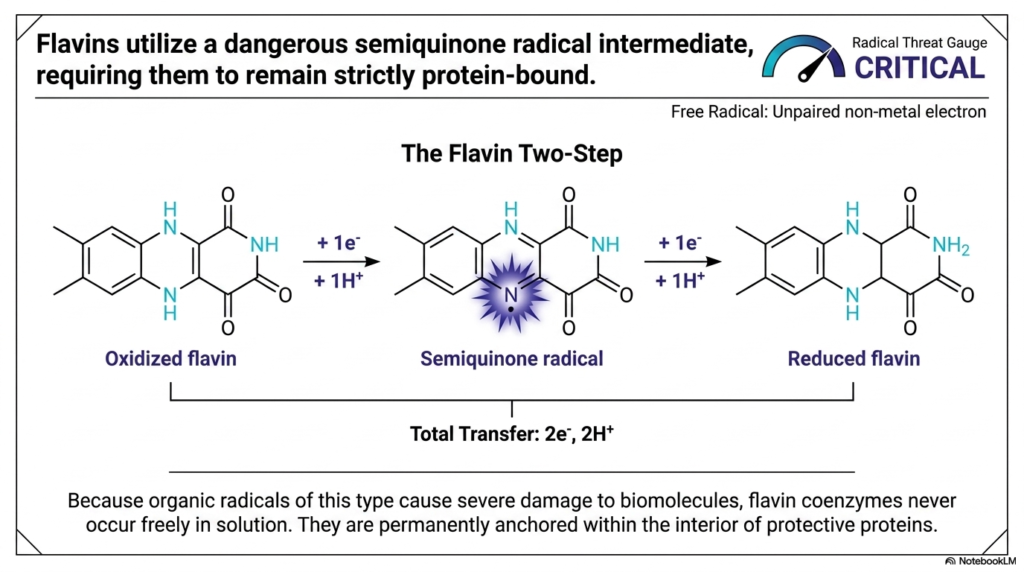
While metal complexes are relatively safe, organic coenzymes have a much higher risk profile, and Flavins are a prime example. This slide introduces “The Flavin Two-Step,” a fascinating but perilous mechanism within Biological Redox Reactions. Flavins, such as FAD and FMN, are essential carriers that can transfer a total of two electrons and two protons. However, unlike other carriers, they do this sequentially—one electron at a time.
This sequential transfer creates a massive problem. After the first electron and proton attach to the oxidized flavin, a “Semiquinone radical” intermediate is formed. Look at the Radical Threat Gauge on this slide: it is firmly in the “CRITICAL” zone. In Biological Redox Reactions, an organic radical such as a semiquinone is highly reactive. It features an unpaired non-metal electron right on the nitrogen ring, desperately seeking to react with anything nearby.
To survive the use of such a hazardous molecule, nature engineered a strict structural failsafe. Because these radical intermediates would cause severe biomolecular destruction if left unchecked, flavin coenzymes never occur freely floating in the cellular solution. As the slide notes, they are permanently anchored within the deep interior of protective proteins. By trapping the flavin deep inside a protein pocket, the cell ensures that Biological Redox Reactions involving this explosive semiquinone intermediate happen in a strictly quarantined environment, preventing the radical from tearing apart the rest of the cell.
Slide 8: Quinones and Cellular Shields in Biological Redox Reactions
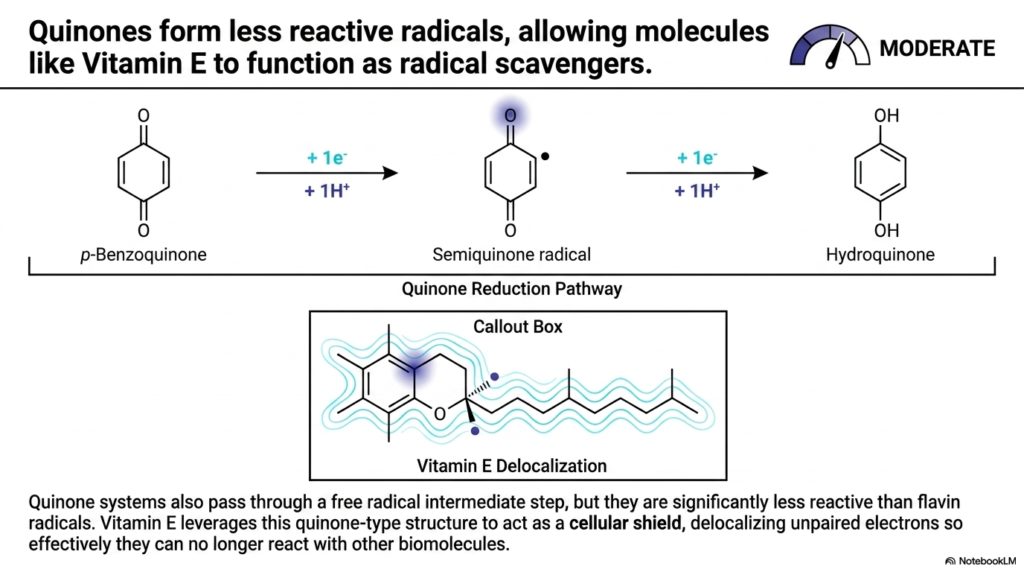
Moving slightly down the danger scale, we arrive at the Quinones. This slide provides an excellent comparison to the volatile flavins we just discussed. In the realm of Biological Redox Reactions, quinones also operate by passing through single-electron steps, forming a semiquinone radical intermediate. However, their chemical structure makes them significantly less dangerous, earning them a “MODERATE” threat rating.
The difference lies in electron delocalization. When p-Benzoquinone accepts an electron and becomes a semiquinone radical, the unpaired electron is not isolated to a single, highly reactive point. Instead, the aromatic ring structure allows the electron’s charge to be spread out, or delocalized, across the molecule. This makes the radical far less aggressive. Because of this inherent stability, quinones do not need to be locked deep inside protein cages like flavins do; they can exist in lipid membranes.
This principle is exactly how Vitamin E functions as a protective antioxidant. In Biological Redox Reactions, Vitamin E leverages its quinone-type structure to act as a “cellular shield.” When a dangerous free radical threatens a cell membrane, Vitamin E intercepts it. It accepts the unpaired electron, but thanks to its extensive delocalization network (highlighted in the Callout Box), it safely absorbs the reactive energy. This effectively neutralizes the threat, proving that manipulating molecular structure is how cells master Biological Redox Reactions to preserve health and prevent tissue damage.
Slide 9: The Cascade of Reactive Oxygen Species in Biological Redox Reactions
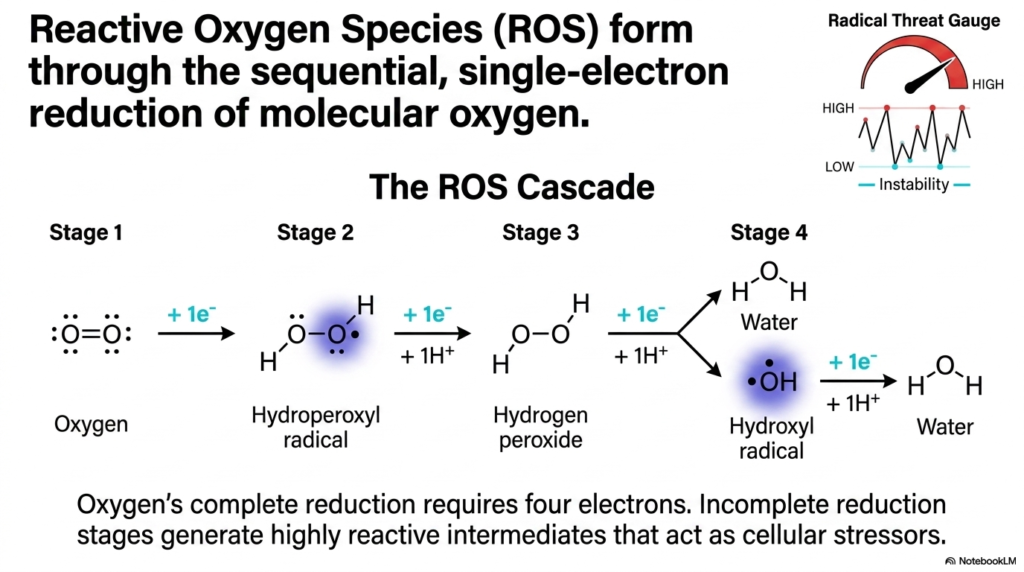
This slide brings us to the ultimate cellular double-edged sword: Oxygen. We literally breathe to supply oxygen to our electron transport chains, but incomplete oxygen processing generates catastrophic cellular stress. This slide diagrams the “ROS Cascade,” showing exactly how Reactive Oxygen Species form through single-electron steps during Biological Redox Reactions.
To fully and safely reduce a molecule of oxygen into water, the cell must deliver exactly four electrons. The danger arises when oxygen is reduced incompletely. If molecular oxygen accidentally picks up just one electron, it becomes a Hydroperoxyl radical (also known as superoxide). A second electron turns it into Hydrogen peroxide, and a third generates the exceptionally destructive Hydroxyl radical. In the study of Biological Redox Reactions, this sequential, single-electron dripping is the primary source of endogenous cellular damage.
The Radical Threat Gauge on this slide goes off the charts as instability increases. These highly reactive intermediates act as severe cellular stressors, aggressively stealing electrons from nearby lipids and DNA to stabilize themselves. This cascade perfectly illustrates why the cell works so incredibly hard to manage its Biological Redox Reactions. The precise molecular machinery we have discussed—the metal complexes, the buried flavins, the quinone shields—are all evolutionary defenses designed specifically to prevent this exact ROS cascade from ripping the cell apart from the inside out.
Slide 10: The Ultimate Safety of Pyridine Nucleotides in Biological Redox Reactions
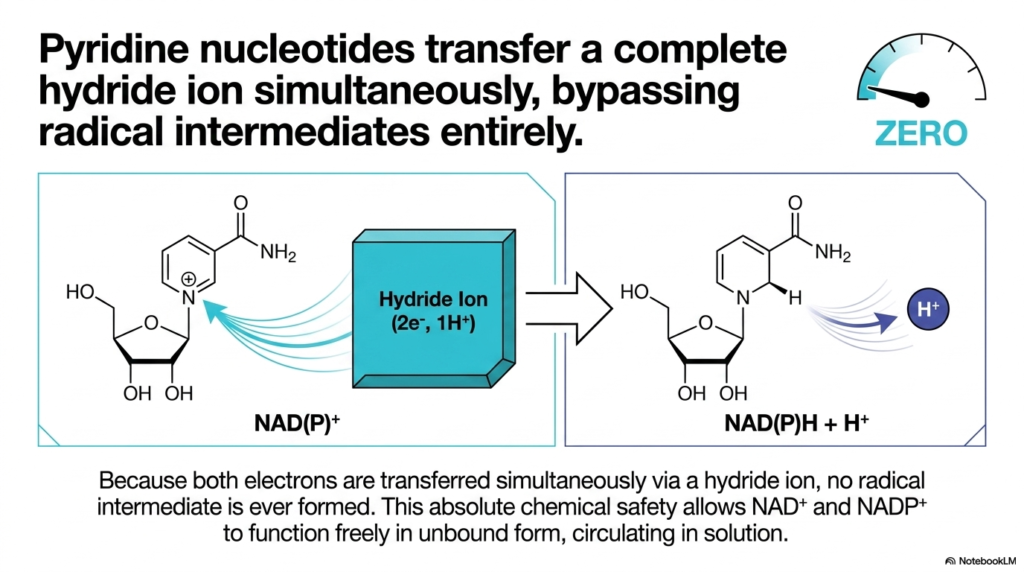
After looking at all the dangers of radical intermediates, we finally arrive at the gold standard of safety: the Pyridine Nucleotides. This slide introduces NAD+ and NADP+, which are arguably the most important organic carriers in all of Biological Redox Reactions. Notice the Radical Threat Gauge is firmly set to “ZERO.” How do these molecules achieve such absolute chemical safety?
The secret is that pyridine nucleotides entirely bypass the single-electron step. Instead of taking electrons one at a time, they transfer a complete Hydride ion, which, as we learned on Slide 4, is a neat package of two electrons and one proton (2e−, 1H+). Because both electrons are transferred simultaneously onto the molecule, a radical intermediate is never formed. Not even for a microsecond.
This absolute safety profile dramatically changes how the cell utilizes these molecules. In the world of Biological Redox Reactions, risk dictates location. Because flavins are dangerous, they are permanently bound to proteins. But because NAD+ and NADP+ form no radical intermediates, they have no need for a protein cage. They are freely soluble and unbound, circulating through the cytoplasm and mitochondrial matrix like safe, reliable delivery trucks. Understanding this simultaneous hydride transfer is key to understanding why NAD+ is the universal, free-floating currency of Biological Redox Reactions across nearly all living organisms.
Slide 11: Resonance Structures Guiding Biological Redox Reactions
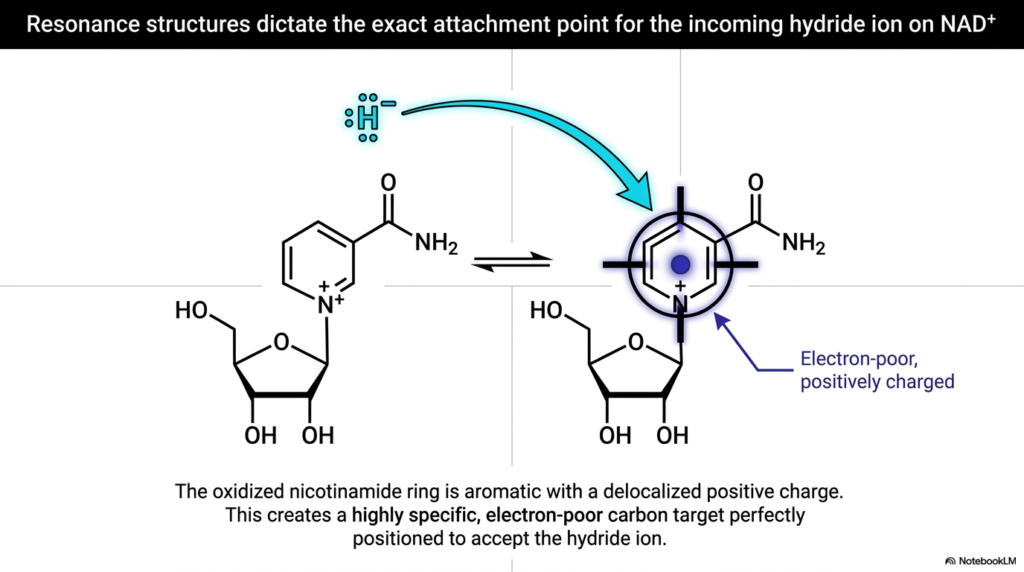
While we know that NAD+ safely accepts a whole hydride ion, we must ask: how does the hydride ion know exactly where to attach? Biology is not random; it is highly specific. This slide delves into the physical chemistry of Biological Redox Reactions, revealing how resonance structures dictate molecular targeting. It provides a beautiful visual of the oxidized nicotinamide ring of NAD+.
The nicotinamide ring is aromatic, meaning its electrons are shared across the ring. Because of the positively charged nitrogen atom within the ring structure, there is a delocalized positive charge spread out over the molecule. This creates a very specific, localized region of the ring that is incredibly electron-poor. In the context of Biological Redox Reactions, electrons—like the heavily negative hydride ion—are inherently attracted to positive, electron-poor environments.
The slide highlights this specific carbon atom with a target aura. The ring’s structure essentially forms a biological landing pad, perfectly positioning the incoming hydride ion. It cannot attach just anywhere; it is magnetically drawn to the exact carbon atom necessary for the reaction to proceed. This level of molecular precision ensures that Biological Redox Reactions occur swiftly, efficiently, and without harmful side reactions. It is a stunning example of how molecular architecture directly dictates biological function down to the subatomic level.
Slide 12: The Master Matrix Summarizing Biological Redox Reactions
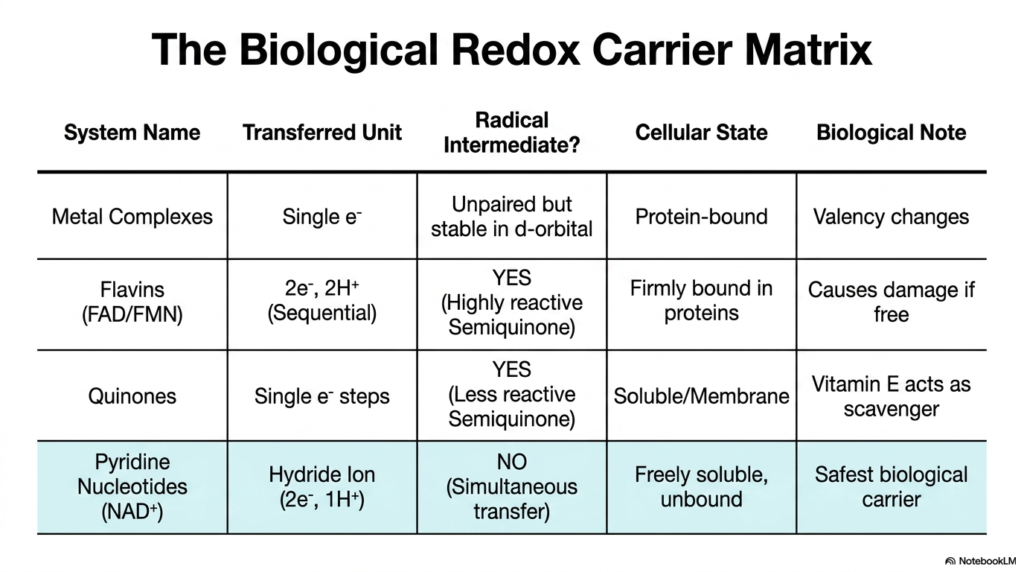
We have covered an immense amount of biochemical ground, and this final slide brings it all together into a beautiful, cohesive summary: “The Biological Redox Carrier Matrix.” This table is your ultimate cheat sheet for understanding how cells manage the flow of energy. By comparing the four major systems we have discussed, you can clearly see the grand evolutionary logic underlying all Biological Redox Reactions.
Let us review the patterns. Metal Complexes manage single electrons safely due to their deep d-orbitals, mostly functioning while protein-bound to change valency safely. Flavins (FAD/FMN) are forced to take sequential steps that create highly reactive semiquinone radicals; therefore, nature mandates they remain firmly bound within protective protein cages. Quinones also form radicals, but their delocalized structures make them less reactive, allowing them to exist as soluble membrane scavengers. Finally, Pyridine Nucleotides (NAD+) represent the safest system in Biological Redox Reactions, transferring simultaneous hydride ions with zero radical intermediates, allowing them to float freely as the cell’s primary energy currency.
This matrix perfectly illustrates the core theme of our lecture. The cell is a master accountant, balancing the energetic need to move electrons against the lethal danger of free radicals. By deeply understanding the rules laid out in this matrix, you are no longer just memorizing pathways; you understand the why behind cellular metabolism. Keep these principles of Biological Redox Reactions in mind as you move forward in your medical and scientific careers—they are the spark that sustains all living things.
Check out our social media channels:
Please read our Content Disclaimer Statement.