33. Understanding Metabolic Regulation: Fundamental Mechanisms
Imagine a bustling metropolis with no traffic lights; absolute chaos would instantly ensue. Similarly, a living cell heavily relies on vast, deeply interconnected biochemical signaling networks to actively maintain absolute internal order and survive. This comprehensive slide deck explores the fundamental molecular mechanisms that govern these essential biochemical pathways. By carefully breaking down transcriptional control, enzyme interconversion, and complex allosteric modulation, this overview clearly illustrates how biological life adapts to physiological demands. For students, mastering these dynamic concepts is absolutely crucial for understanding human health.
Slide 1: Introduction to Metabolic Regulation
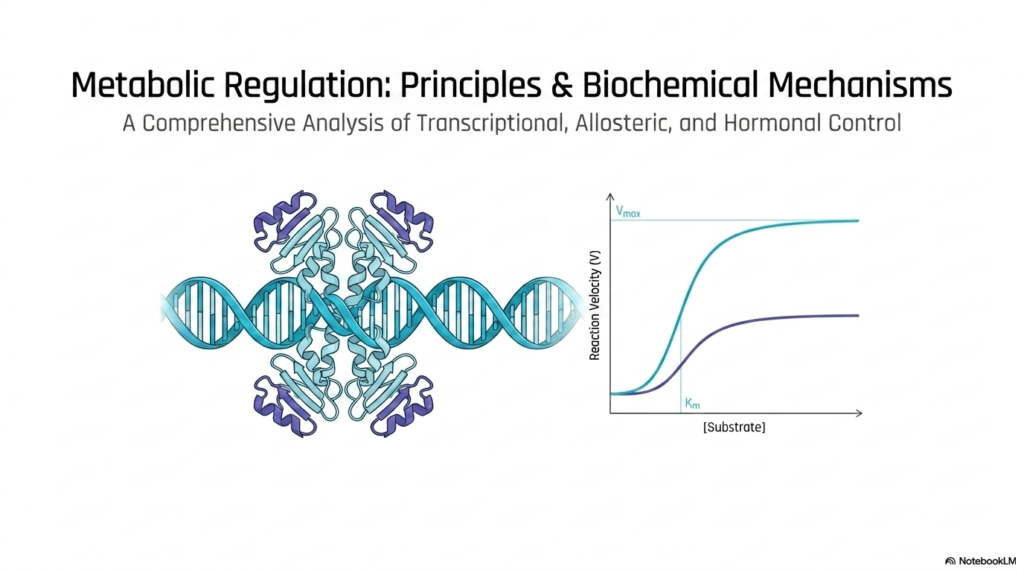
The opening visualization perfectly sets the stage for understanding complex biological control systems. At the heart of all cellular function lies metabolic regulation, a highly dynamic process that ensures biochemical pathways operate efficiently and respond instantly to the organism’s immediate physiological needs. The imagery prominently features a DNA double helix entwined with structural regulatory proteins, beautifully symbolizing the foundational role of genetic control in these biochemical processes. This visual elegantly captures how biological life is fundamentally governed by precisely timed molecular interactions.
Adjacent to the genetic structure, the slide presents a crucial biochemical graph showing reaction rate versus substrate concentration. This graph clearly contrasts standard hyperbolic Michaelis-Menten kinetics with the unique sigmoidal curve characteristic of complex allosteric enzymes. Understanding this distinct kinetic behavior is vital for mastering metabolic regulation, as it visually demonstrates how certain key enzymes can act as highly sensitive molecular switches. The sigmoidal nature reveals cooperative binding, allowing enzymes to rapidly ramp up their catalytic activity over a narrow range.
By highlighting both genetic architecture and enzymatic kinetics, the slide outlines a highly comprehensive analysis of cellular control. The synthesis of transcriptional, allosteric, and hormonal control forms the essential triad of metabolic regulation. College and medical students must grasp that these are not isolated physiological phenomena, but rather heavily interconnected layers of a highly evolved biological network. Each layer offers a distinct degree of sensitivity and response time, ultimately maintaining the delicate balance of homeostasis required for cellular survival.
A deep dive into these exact principles reveals how the biological cell completely avoids chaotic overproduction and wasteful depletion of resources. Without effective control, the simultaneous operation of opposing metabolic pathways would quickly lead to a futile, deadly cycling of cellular energy. The structural complexes and kinetic behaviors introduced in this visual serve as the fundamental vocabulary for the deeper mechanisms explored in subsequent sections. This comprehensive overview perfectly prepares the learner to transition from basic principles to specific, intricate biological pathways.
Slide 2: The Temporal Hierarchy of Metabolic Regulation
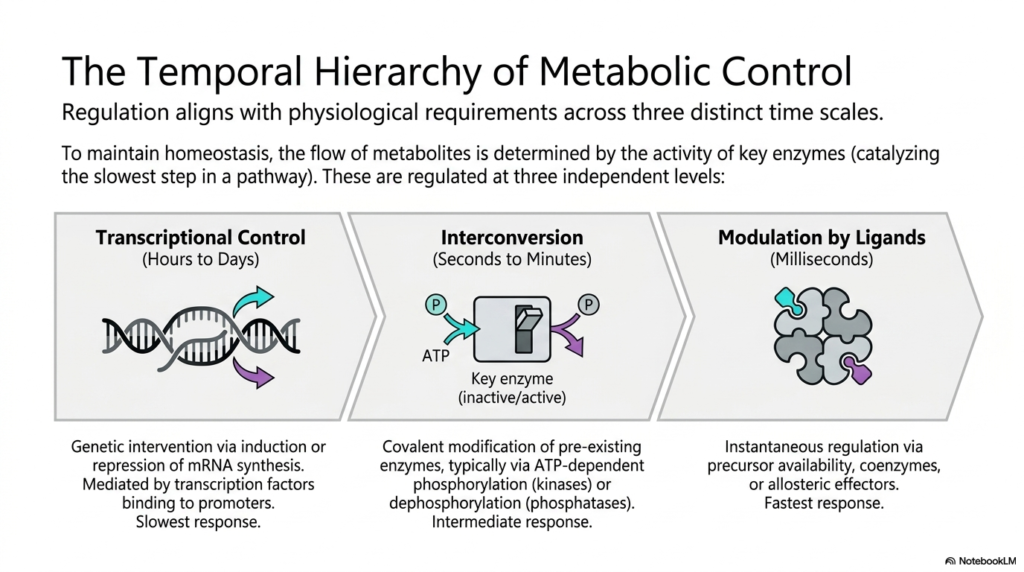
Cellular homeostasis relies entirely on the precise flow of chemical metabolites, which is strictly governed by the activity of key enzymes. These highly crucial enzymes typically catalyze the slowest, rate-limiting step in any given biochemical pathway. To ensure maximum efficiency, metabolic regulation operates across three distinct time scales, carefully aligning cellular responses with precise physiological requirements. This precise temporal hierarchy completely prevents wasteful energy expenditure while allowing the organism to adapt to both immediate environmental shifts and long-term developmental changes.
The slowest, most foundational layer of metabolic regulation involves complex transcriptional control, which unfolds over hours or even days. This sophisticated process entails genetic intervention by directly inducing or repressing messenger RNA synthesis deep within the nucleus. Specialized transcription factors bind to specific promoter regions of DNA, dictating whether the cellular machinery will produce more of a specific enzyme. Because it requires synthesizing completely new protein structures from scratch, this response provides long-lasting biological adjustments.
Operating on a much faster intermediate time scale of seconds to minutes is the highly efficient process of enzyme interconversion. This rapid form of metabolic regulation fundamentally relies on the targeted covalent modification of pre-existing enzymatic proteins. Typically driven by ATP-dependent phosphorylation by specific kinases or dephosphorylation by phosphatases, this mechanism physically alters the target enzyme’s conformation. By simply adding or removing a charged phosphate group, the cell can instantly toggle a key enzyme between its active and inactive physiological states.
The absolute fastest cellular response occurs in mere milliseconds through the direct modulation of enzymes by various ligands. This highly instantaneous tier of control relies heavily on precursor availability, localized coenzyme levels, or the rapid binding of small allosteric effectors. By carefully utilizing these distinct, independent levels of temporal control, the biological system creates a highly robust network. The seamless integration of slow genetic shifts, rapid covalent modifications, and instant allosteric adjustments ensures that the cell can survive fleeting nutrient fluctuations.
Slide 3: Ligand Modulation in Metabolic Regulation
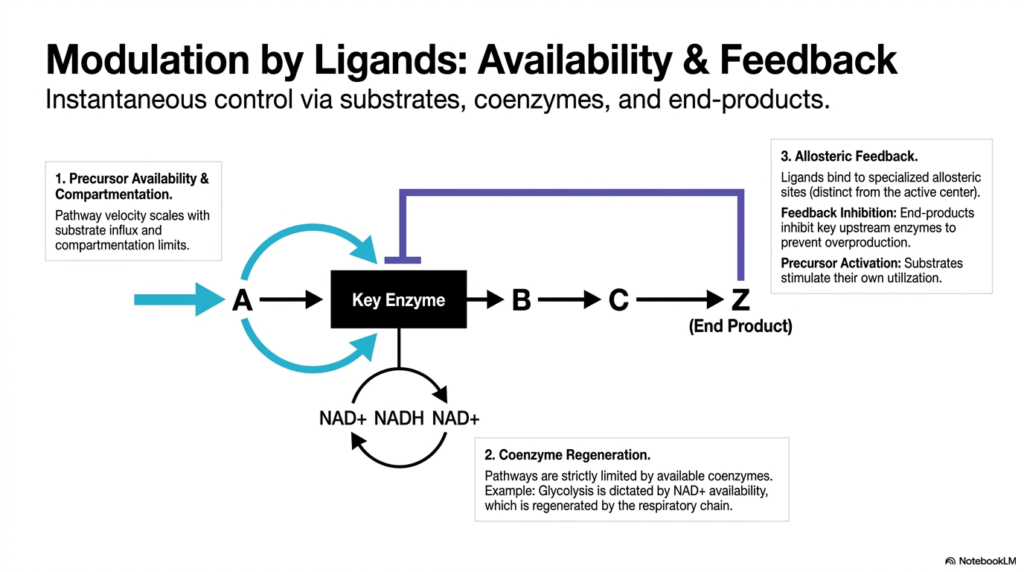
Instantaneous cellular control heavily relies on the direct modulation of enzymes by various biological ligands, specifically including substrates, crucial coenzymes, and terminal end-products. The overall velocity of any metabolic pathway fundamentally scales with the immediate availability of its precursors and the physical limits of internal cellular compartmentation. When substrates are highly abundant, they can naturally stimulate their own enzymatic utilization, a powerful effect known as precursor activation. This localized form of metabolic regulation ensures that key enzymes only operate when raw materials are sufficient.
Furthermore, these complex metabolic pathways are strictly physically limited by the available intracellular pool of specific coenzymes. A classic, highly relevant example is glycolysis, which is entirely dependent on the immediate availability of the NAD+ coenzyme. Because NAD+ must be continuously regenerated by the highly complex respiratory chain, the overall metabolic flux is tightly coupled to the cell’s oxidative capacity. This dependency represents a passive but highly effective method of metabolic regulation, linking the breakdown of glucose directly to immediate energy states.
Beyond simple chemical availability, the biological cell actively utilizes highly sophisticated allosteric feedback loops. In these complex systems, specific small-molecule ligands bind to specialized allosteric sites that are physically entirely distinct from the enzyme’s catalytic active center. Through negative feedback inhibition, the final end-products of a pathway can directly bind to and completely inhibit the key upstream enzymes. This specific form of metabolic regulation acts as a powerful emergency brake, instantly shutting down an entire multi-step pathway to prevent toxic overproduction.
The true biological beauty of these specialized regulatory loops lies in their instantaneous nature and incredibly high degree of chemical specificity. By explicitly targeting the first committed step of a biochemical pathway, allosteric effectors provide a highly efficient, fail-safe mechanism to actively reroute metabolites toward alternative biological needs. This highly localized ligand control allows individual organelles to make rapid micro-adjustments on the fly, perfectly complementing the much slower, systemic shifts orchestrated at the broader hormonal and genetic levels across the entire living organism.
Slide 4: Allosteric Kinetics in Metabolic Regulation
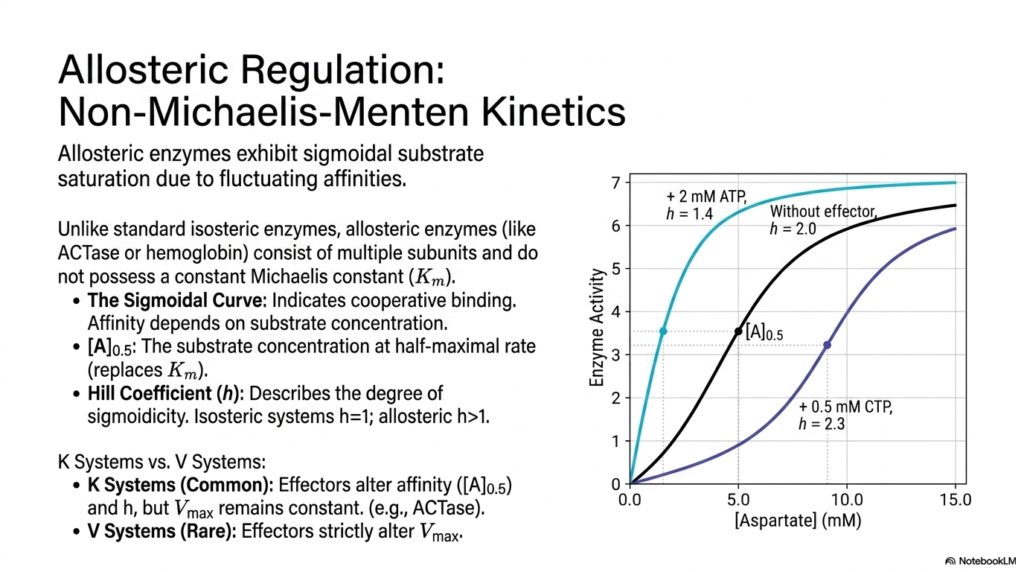
Traditional isosteric enzymes closely follow standard Michaelis-Menten kinetics, reliably producing a highly predictable hyperbolic curve as substrate concentration increases. However, highly complex allosteric enzymes deviate markedly from this standard model, instead exhibiting a distinctive sigmoidal substrate saturation curve. This defining kinetic characteristic arises directly from the complex quaternary structural architecture of these proteins, which consist of multiple physically interacting subunits. Understanding this sigmoidal behavior is fundamentally crucial for mastering metabolic regulation at the systemic biological level.
The presence of a sigmoidal curve clearly indicates highly cooperative binding among the enzyme’s subunits. When one single substrate molecule binds, it successfully induces a profound conformational change that dramatically increases the specific binding affinity of the remaining vacant sites. Because they lack a constant Michaelis constant, biochemists use [A]0.5 to denote the substrate concentration at the half-maximal reaction rate. The exact degree of sigmoidicity is formally described by the Hill Coefficient, which is greater than one in these specific complex systems.
These highly sensitive allosteric enzymes are typically categorized into two highly distinct regulatory classes: K Systems and V Systems. K Systems are exceedingly common in normal metabolic regulation. In these biological setups, allosteric effectors physically alter the enzyme’s intrinsic affinity for the target substrate, thereby shifting the [A]0.5 value and the Hill Coefficient, while the reaction’s absolute maximum velocity remains constant. A classic biological example is Aspartate Carbamoyltransferase, in which effectors dynamically shift the enzyme’s activation threshold.
In stark contrast, V Systems are considered relatively rare in standard biological contexts. In these highly specialized cellular cases, the direct binding of allosteric effectors strictly alters the maximum velocity of the targeted enzymatic reaction without significantly impacting the underlying substrate affinity. The overarching systemic importance of both distinct systems in metabolic regulation cannot be mathematically overstated. By shifting these vital kinetic parameters, allosteric enzymes act as highly sensitive metabolic switches, capable of rapidly turning a biological pathway on or off.
Slide 5: ACTase as a Model of Metabolic Regulation
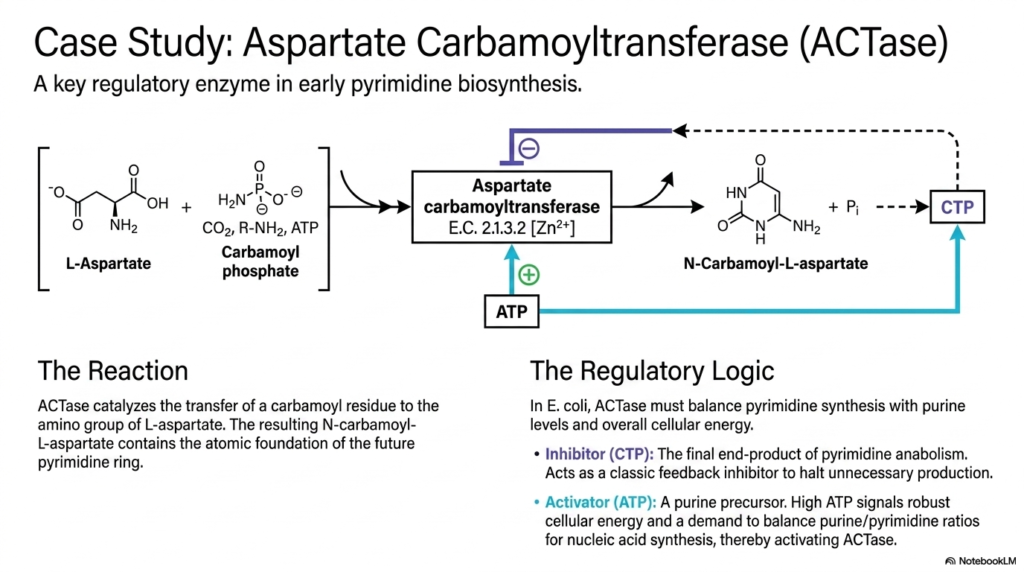
Aspartate Carbamoyltransferase, widely known by the acronym ACTase, is a premier biological case study for understanding complex allosteric control mechanisms. This key regulatory enzyme catalyzes the first committed step in early pyrimidine biosynthesis in specific bacteria such as E. coli. The highly specific chemical reaction directly transfers a carbamoyl group to the amino group of L-aspartate, thereby creating N-carbamoyl-L-aspartate. This newly formed molecule contains the complete atomic structure required for the eventual formation of the complex pyrimidine ring.
The underlying regulatory logic governing ACTase perfectly and beautifully illustrates the broader, highly complex principles of metabolic regulation. The massive enzyme is specifically tasked with heavily balancing pyrimidine synthesis with both existing purine levels and the overall energy state of the bacterial cell. The final complex end-product of this specific pyrimidine anabolic pathway is Cytidine Triphosphate (CTP). As intracellular CTP concentrations naturally rise, the molecule acts directly as a classic, powerful feedback inhibitor, binding to regulatory sites to halt further unnecessary biological production.
Conversely, the high concentration of Adenosine Triphosphate (ATP) acts as a potent activator of the same enzyme. ATP is a vital purine precursor and is the primary chemical energy currency of the living cell. High levels of ATP strongly signal robust cellular energy and a pressing biological demand to balance the purine-to-pyrimidine ratio. This dynamic, highly sensitive interplay between ATP and CTP showcases how metabolic regulation seamlessly integrates multiple, strongly conflicting cellular biochemical signals.
Through this sophisticated, highly evolved structural mechanism, the massive enzyme autonomously and continuously interprets the exact metabolic environment. When cellular energy is incredibly abundant, and purines severely outweigh pyrimidines, the system rapidly accelerates pyrimidine generation. When pyrimidines eventually accumulate to biologically sufficient levels, the pathway is immediately throttled by the product. This highly elegant system of metabolic regulation ensures that the bacterial cell consistently and efficiently maintains the exact stoichiometric ratios of nucleotides required for replication and overall cellular survival.
Slide 6: Conformational Shifts in Metabolic Regulation
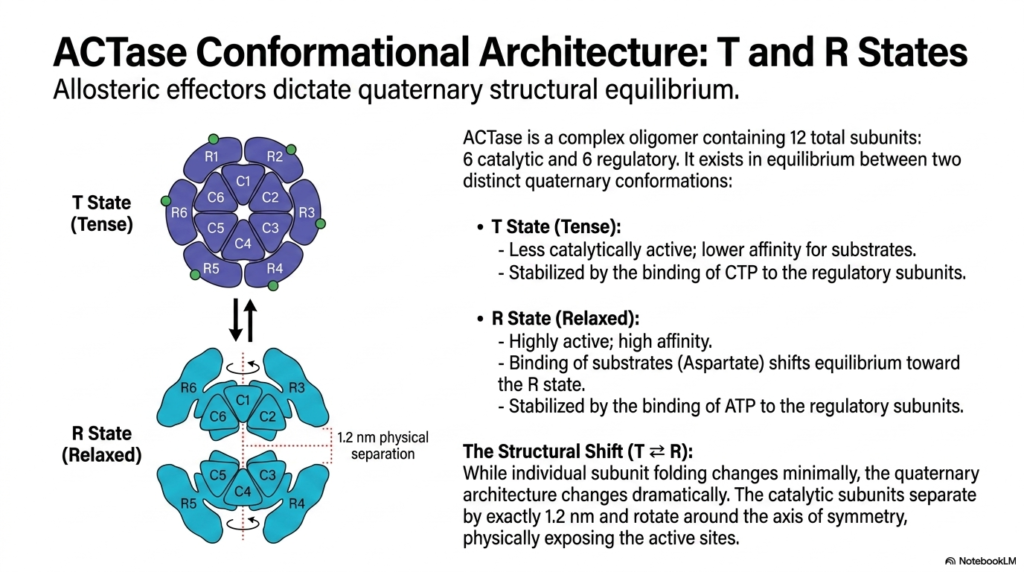
The truly remarkable kinetic behavior of ACTase is entirely physically dependent on its incredibly complex conformational architecture. The massive biological enzyme is a large oligomer composed of twelve structural subunits: six active catalytic subunits responsible for the chemical reaction, and six peripheral regulatory subunits that selectively bind allosteric effectors. The entire structure exists in a highly delicate structural equilibrium between two totally distinct quaternary conformations. This structural dynamism is a hallmark of metabolic regulation at the atomic-level biological level.
In the complete absence of activating substrates, or when tightly bound to the powerful inhibitor CTP, the enzyme strongly favors the T State, commonly known as the “Tense” state. The T State is clearly characterized by a significantly less catalytically active internal conformation and a massively lower affinity for its target substrates. CTP binds highly specifically to the regulatory subunits, heavily stabilizing this compact physical structure. This physical lockdown is a major structural mechanism of metabolic regulation, actively suppressing the biochemical pathway continuously to preserve energy.
The direct biological introduction of specific substrates, particularly Aspartate, strongly shifts the equilibrium toward the R State, widely known as the “Relaxed” state. The R State is a highly biologically active conformation that boasts a strong chemical affinity for incoming substrates. Furthermore, the direct binding of ATP to the external regulatory subunits helps stabilize this highly relaxed architecture. The critical transition from the T to the R state actively represents a highly dramatic physical structural shift within the overall protein architecture.
During this highly functional transition, the specific catalytic subunits literally and physically separate by exactly 1.2 nanometers and physically rotate around their central axis of symmetry. This profound mechanical movement exposes the buried active sites, instantly making them readily accessible for rapid, continuous substrate processing. This dramatic 1.2-nanometer biological expansion is the true physical embodiment of metabolic regulation, perfectly illustrating how tiny chemical signals can reliably induce massive mechanical movements to successfully toggle an enzyme’s biological function off and on.
Slide 7: Structural Domains Governing Metabolic Regulation
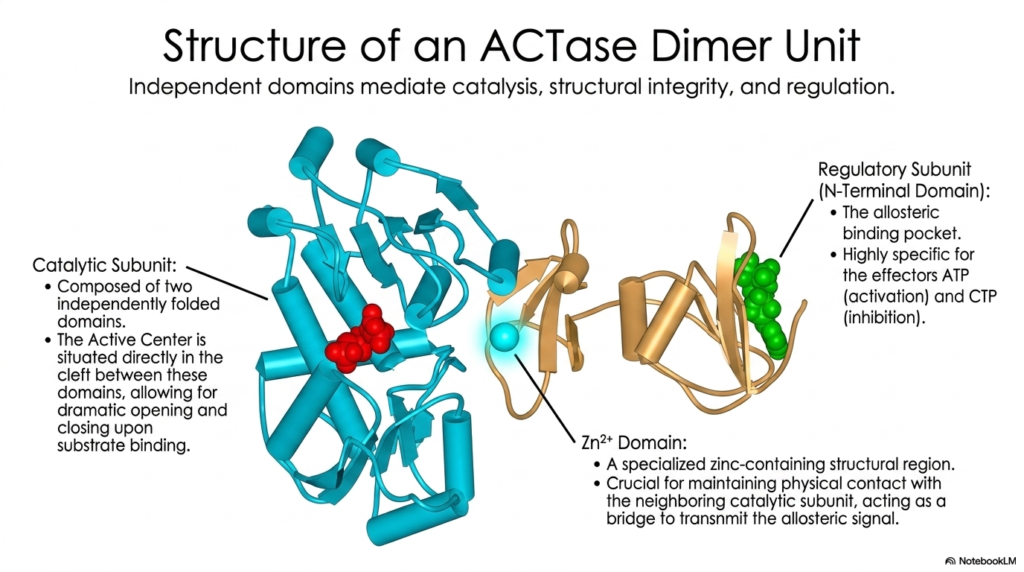
To fully appreciate mathematically how structural integrity dictates complex enzymatic function, students must closely examine the highly specific architecture of an individual ACTase dimer. The massive protein continuously utilizes highly specialized, completely independent domains to seamlessly mediate catalysis, fiercely maintain structural stability, and flawlessly execute regulatory commands. The main catalytic subunit itself is carefully composed of two completely independent folded biological domains. The active center is positioned directly within the molecular cleft between these structures, ready for immediate enzymatic action.
Tightly attached to this powerful catalytic machinery is the specific regulatory subunit, which prominently features a massive N-Terminal Domain. This exact specific structural domain houses the highly critical allosteric binding pocket. The pocket is highly specific for the enzyme’s distinct allosteric effectors, strictly recognizing ATP for biological activation and CTP for targeted inhibition. The deliberate physical separation of the active center and the allosteric pocket is a major defining structural feature of complex metabolic regulation, allowing for precise control without interference.
Carefully bridging the vast physical gap between the distant regulatory and catalytic subunits is a highly specialized zinc-containing structural region widely known as the Zinc Domain. The continuous presence of the single zinc ion is absolutely biologically crucial for fiercely maintaining direct physical contact with the neighboring catalytic subunit. It serves as an essential, highly stable physical bridge, specifically tasked with rapidly transmitting the allosteric signal generated by chemical ligand binding through the protein’s internal biological structure to ensure a proper systemic cellular response.
Without the extreme structural integrity explicitly provided by the rigid Zinc Domain, the highly critical biochemical signals from ATP or CTP would completely fail to reach the physically distant active site. The intricate, highly evolved design of the specific dimer unit beautifully highlights the incredibly precise atomic engineering consistently found in complex biochemical systems. Each unique structural domain has a highly specific, irreplaceable function, working in perfect concert to enable the cell to continuously exert exquisite metabolic regulation over its internal pathways.
Slide 8: Genetic Mechanisms of Metabolic Regulation
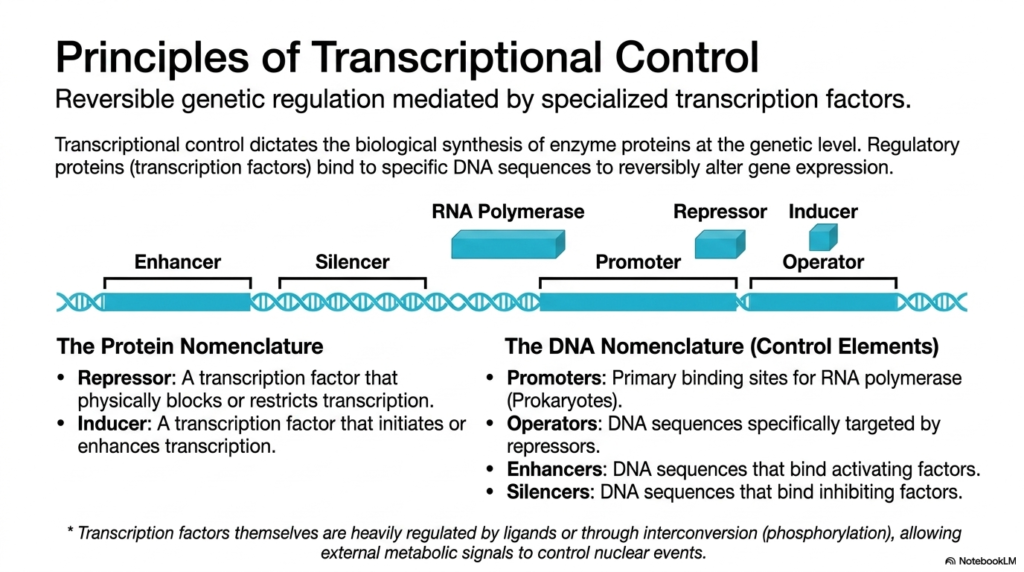
Moving well beyond instantaneous, rapid enzymatic shifts, long-term metabolic regulation is entirely dependent on the highly complex principles of nuclear transcriptional control. This specific genetic mechanism directly dictates the complex biological synthesis of the enzyme proteins themselves at the foundational genetic level. Reversible genetic regulation is continuously mediated by highly specialized regulatory proteins commonly known as transcription factors. These specific factors bind to unique DNA sequences to forcibly and reversibly alter the expression of targeted genes based on the cell’s immediate needs.
A strong biological grasp of this complex genetic process requires a thorough understanding of the specific protein nomenclature involved in metabolic regulation. A cellular repressor is a powerful transcription factor that heavily physically blocks or severely restricts the biological transcription of a targeted gene. Conversely, an inducer is a highly specialized transcription factor specifically designed to initiate or significantly enhance gene transcription. Their biological activity is often heavily regulated by specific cellular ligands or through direct covalent interconversion signaling cascades.
On the complex genetic side, highly precise structural DNA control elements dictate exactly where these specific proteins physically intervene. Promoters are the primary binding sites for the enzyme RNA polymerase, particularly in certain prokaryotic organisms, marking the precise physical starting point for genetic transcription. Operators are highly specific DNA sequences that are selectively targeted and tightly bound by specific repressor proteins. This structural arrangement ensures that, when a repressor is active, it can completely block the polymerase from advancing.
In addition to standard promoters and operators, highly complex eukaryotic systems frequently actively utilize distal enhancers and silencers. Enhancers are vast, distal DNA sequences that actively bind potent activating factors to massively supercharge gene transcription, while silencers strongly bind specific inhibitory factors to forcefully shut the biological process down. The careful coordination between these varied DNA elements and their corresponding biological transcription factors forms a highly complex network of metabolic regulation, ensuring that specific enzymes are only synthesized when explicitly required by the organism.
Slide 9: The Matrix of Metabolic Regulation in Genetics
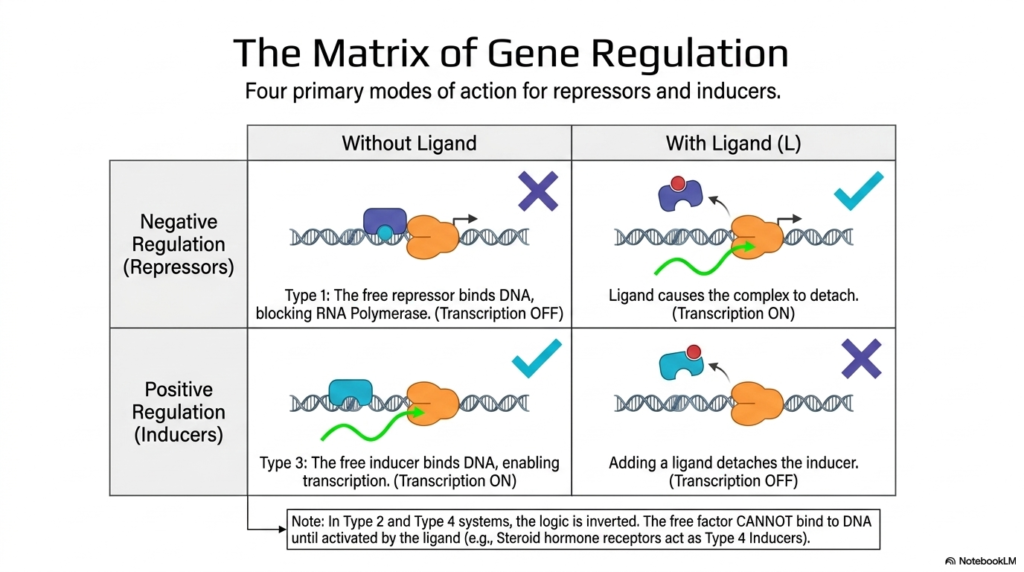
Complex genetic control systems can be systematically categorized into exactly four primary modes of biological action, forming a comprehensive, highly predictive matrix of gene regulation. The biological system systematically categorizes these mechanisms primarily into targeted negative regulation, which relies heavily on repressors, and positive regulation, which actively utilizes inducers. Deeply understanding this exact matrix is crucial for all medical students studying metabolic regulation, as it clearly illustrates how the presence or complete absence of specific small-molecule ligands can operate effectively.
In highly specific Type 1 negative regulation, the complex biological system actively employs a massive default lock. In the absence of a specific ligand, the free repressor protein binds tightly and naturally to the target DNA operator sequence. This binding aggressively forms a massive physical barrier that strongly blocks RNA Polymerase and effectively shuts down biological transcription. When the specific ligand is introduced, it tightly binds to the repressor, actively causing a massive conformational change that violently forces the complex to detach from the DNA entirely.
Conversely, highly complex Type 3 positive regulation aggressively utilizes a specific inducer to successfully drive metabolic regulation. In this specific biological scenario, the fully free inducer protein naturally binds directly to DNA, enabling continuous transcription in the baseline default state. However, the direct cellular addition of a highly specific ligand to the biological system tightly binds to the inducer and violently forces it to detach from the targeted genetic sequence, instantly halting RNA polymerase activity.
It is absolutely biologically vital to closely note that this precise logic can be completely naturally inverted in highly specific Type 2 and Type 4 biological systems. In these inverted, highly specialized architectures, the transcription factor cannot physically bind to the target DNA until it is fully biologically activated by a highly specific external ligand. A highly classic, medically relevant example involves specific steroid hormone receptors, which function powerfully as Type 4 inducers in metabolic regulation, initiating profound physiological shifts systemically.
Slide 10: The Lactose Operon in Metabolic Regulation
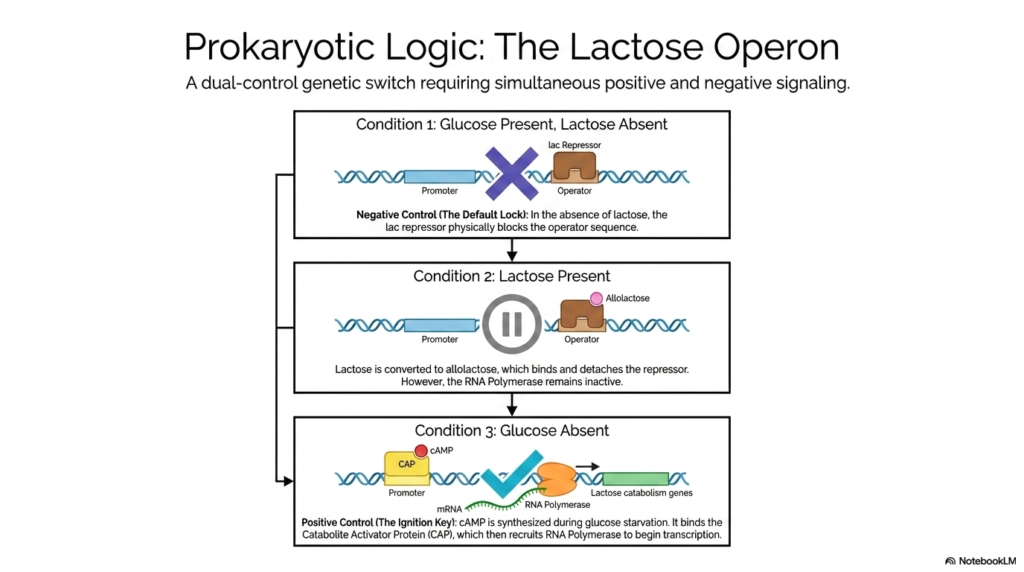
The classic prokaryotic Lactose Operon clearly represents an absolute structural masterpiece of advanced biological computing, functioning as a highly integrated, dual-control genetic switch. This classic, well-studied biological system requires simultaneous positive and strong negative signaling to function, providing a truly perfect educational model for understanding multi-layered metabolic regulation. The complex operon actively governs the genes necessary for lactose catabolism, but to aggressively maximize energy efficiency, these genes must be activated only under extremely specific environmental conditions.
The biological system’s powerful negative control mechanism actively acts as a massive default physical lock. In the complete and total absence of lactose, the lac repressor remains highly active and physically binds tightly to the operator sequence, completely restricting access for the enzyme RNA polymerase. However, when lactose is abundant, it is rapidly converted by enzymes into a specific isomer, known as allolactose. Allolactose is the specific chemical ligand that binds to the repressor, removing the physical block.
The truly massive biological ingenuity of this precise form of metabolic regulation lies entirely in its powerful secondary positive control mechanism, which specifically serves as the highly complex genetic ignition key. When standard glucose is highly naturally abundant, the specific operon remains largely dormant, even if lactose is highly present. It is only during periods of severe glucose starvation that the bacterial cell produces large amounts of the critical signaling molecule cAMP. This crucial molecule rapidly binds to the Catabolite Activator Protein, driving activation.
The highly complex lactose operon clearly demonstrates that basic metabolic responses are tightly guarded by extensive redundancy. The complex cellular machinery will fiercely not expend massive amounts of critical energy synthesizing lactose-digesting enzymes if a highly preferred, completely easily metabolized energy source is readily available. By aggressively requiring both the physical removal of a repressor and the heavy binding of an activator, this genetic switch perfectly showcases the sheer massive elegance of metabolic regulation at the absolute genetic level.
Slide 11: DNA Bending and Metabolic Regulation
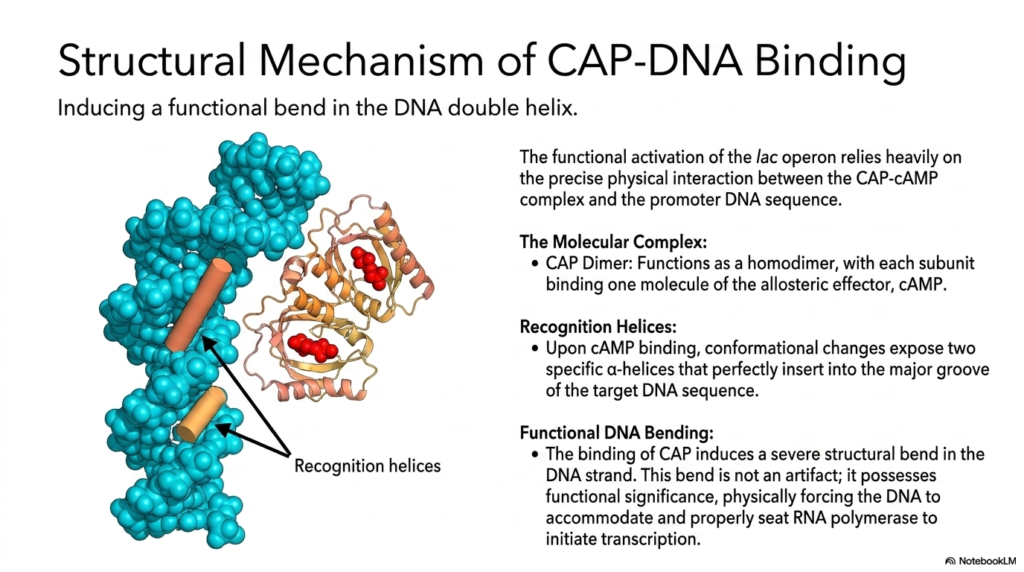
The highly functional activation of the complex lac operon offers a truly fascinating, deeply educational glimpse into the precise physical mechanics of complex metabolic regulation, operating at the atomic level. Biological activation relies entirely on the precise, highly specific physical interaction between the CAP-cAMP molecular complex and the target promoter DNA sequence. The Catabolite Activator Protein continuously functions strictly as a massive homodimer in the cellular environment, with each subunit binding one molecule of the vital allosteric effector cAMP.
Upon rapidly binding the crucial effector cAMP, the massive CAP dimer instantly undergoes a highly significant conformational shift. This profound structural-biological change completely exposes two highly specific alpha-helical recognition helices on the protein complex’s vast exterior. These specific recognition helices are perfectly chemically contoured to insert deeply into the vast major groove of the exact target DNA sequence, showcasing a massive, highly specific structural hallmark of efficient genetic metabolic regulation.
The most dramatic, biologically profound consequence of this specific protein-DNA structural interaction is the resulting massive structural deformation of the genetic code itself. The tight binding of the massive CAP dimer actively induces a completely severe, massive, nearly 90-degree structural bend in the rigid physical DNA double helix. Medical students must understand that this extreme, dramatic bend is absolutely not an incidental structural artifact; it has profound functional significance in supercharging gene transcription by altering the DNA’s physical landscape.
By violently and aggressively bending the rigid DNA strand, the massive CAP complex effectively wraps the entire genetic sequence completely around the massive incoming RNA polymerase enzyme. This extreme mechanical action aggressively forces the target DNA to seamlessly accommodate and properly seat the massive polymerase enzyme, drastically increasing its overall binding affinity to the exact promoter region. This profound mechanical interaction clearly demonstrates that metabolic regulation relies on precise, forceful cellular interactions to function properly.
Slide 12: Systemic Hormonal Metabolic Regulation
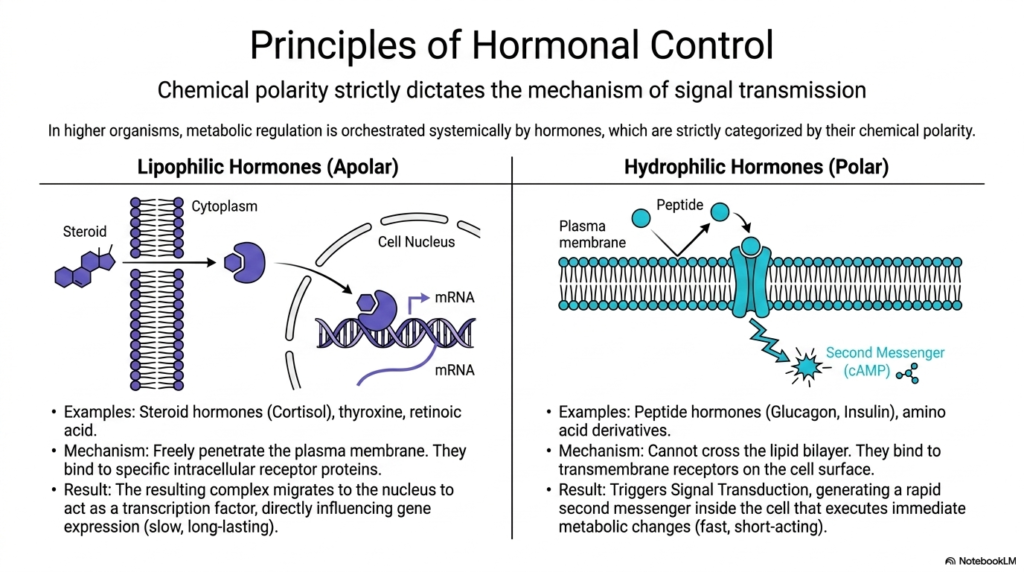
While rapid allosteric and highly complex genetic controls actively manage highly localized cellular internal environments, metabolic regulation in vastly complex higher organisms must absolutely be seamlessly orchestrated systemically across multiple independent tissues. This utterly massive biological coordination is actively and continuously executed by specific hormones, which are powerful chemical messengers that travel seamlessly throughout the entire bloodstream. A fundamental biological principle establishes that the specific chemical polarity of these distinct molecules strictly dictates their exact mechanism of systemic biological signal transmission and overall cellular physiological impact.
Specific lipophilic hormones, notably powerful steroid hormones such as systemic cortisol, vital thyroxine, and retinoic acid, are completely apolar and highly hydrophobic. Due to their distinct chemical nature, these molecules can easily and freely penetrate the rigid lipid bilayer of the plasma membrane without requiring any surface transport proteins. Once deep inside, they bind specifically and tightly to internal receptor proteins, allowing the resulting complex to act as a potent transcription factor for metabolic regulation, driving very slow, extremely long-lasting genetic changes.
Conversely, highly critical hydrophilic hormones are highly polar molecules that cannot readily cross the highly impermeable lipid bilayer. This absolutely distinct biological category explicitly includes vital peptide hormones, such as rapid glucagon and systemic insulin. To actively and successfully exert massive metabolic regulation, they absolutely must bind to highly complex, highly specific transmembrane receptors located on the cell surface. This external physical binding event heavily instantly triggers a massively complex internal process widely known as rapid cellular signal transduction.
This vast, complex biological process completely and safely generates a massive, rapid surge of a highly specific small internal second messenger, specifically cyclic AMP (cAMP), deep inside the cell. These tiny internal second messengers rapidly and completely trigger immediate, massively systemic cellular metabolic changes by activating specific rapid kinase cascades. Understanding this massive chemical structural dichotomy is essential for all highly trained medical professionals who actively and continuously study systemic metabolic regulation and associated endocrine pathology.
Slide 13: Liver Homeostasis and Metabolic Regulation

The massive systemic liver continuously functions as the central, indispensable biological hub for systemic glucose homeostasis in the human body. To maintain stable blood sugar levels, this vast organ continuously balances the massive synthesis of stored internal glycogen with its extremely rapid breakdown and release. This highly critical, overarching process of metabolic regulation is strictly orchestrated by a powerful, finely tuned hormonal triad. These three hormones constantly monitor and forcefully direct the liver’s vast biochemical resources to ensure the organism’s total survival.
Extremely powerful, rapid-acting systemic glucagon, a highly essential peptide hormone, is aggressively released by the pancreas purely during dangerous periods of profoundly low blood glucose. It acts via a highly efficient, massive internal cAMP signaling cascade to fiercely drive rapid enzyme interconversion. The primary, profound physiological result of this intense metabolic regulation is the rapid, highly aggressive activation of systemic cellular glycogen breakdown. Simultaneously, the pathway completely inhibits glycogen synthesis, creating a rapid, powerful one-way flood of vital glucose directly from the liver into the bloodstream.
Acting in complete direct structural opposition to this physiological process permanently stands systemic insulin, another immensely powerful and critical peptide hormone. Systemic insulin actively and effectively executes its own distinct, precise form of systemic metabolic regulation by rapidly reducing intracellular cAMP levels via specific phosphodiesterase enzymes. The ultimate physiological result successfully protects and violently activates crucial glycogen synthase. This specific action aggressively drives the liver to immediately safely store vast amounts of excess circulating post-meal glucose as internal cellular glycogen reserves for future biological use.
When the liver’s internal cellular glycogen energy reserves are thoroughly exhausted under extremely severe, prolonged physiological stress, the body rapidly deploys cortisol. As a highly powerful lipophilic steroid hormone, cortisol completely avoids rapid interconversion and actively relies entirely on extremely slow metabolic regulation through nuclear-targeted genetic transcriptional control. It effectively and massively upregulates the biological synthesis of completely new vital gluconeogenic enzymes from raw amino acids. This ultimately gives the entire liver the long-term biological capacity to create completely new glucose molecules from scratch.
Slide 14: Cascades in Metabolic Regulation
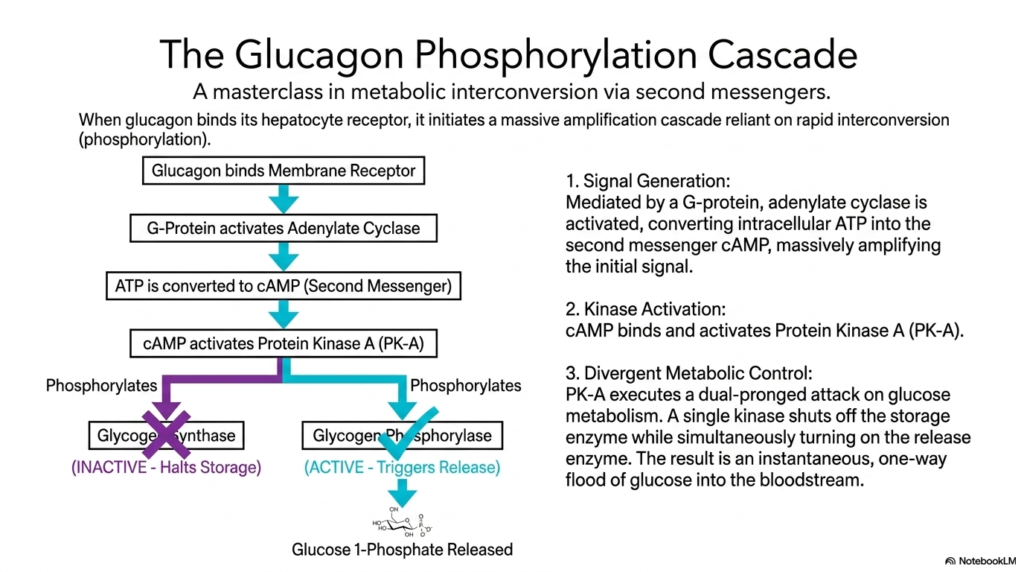
The glucagon signaling pathway is a masterclass in cellular control, showcasing metabolic regulation via rapid second messengers. When the powerful peptide hormone glucagon binds precisely to its highly specific external receptor on the hepatocyte membrane, it instantly initiates a massive biological cascade. This sophisticated mechanism allows a single microscopic hormone molecule circulating in the vast bloodstream to trigger an overwhelming biochemical response deep inside the target cell. Understanding this signal transduction pathway is vital for medical students studying metabolic disorders.
Once activated by the hormone-receptor interaction, a specialized membrane-bound G protein seamlessly stimulates the crucial enzyme adenylate cyclase. This highly aggressive enzyme rapidly converts vast intracellular pools of standard ATP directly into the vital second messenger, cyclic AMP (cAMP). This single, highly localized metabolic regulatory event massively amplifies the initial hormonal signal. By rapidly generating thousands of cAMP molecules, the cell effectively bridges the gap between the extracellular environment and its internal biochemical machinery, preparing for an immediate physiological response.
Upon rapid generation, this massive surge of cAMP binds to and fully activates Protein Kinase A (PK-A). This master enzyme actively executes a highly specific, totally exact dual-pronged chemical metabolic regulation. PK-A functions by aggressively attaching charged phosphate groups to specific target enzymes in a process known as covalent interconversion. By precisely executing this chemical modification, PK-A manages to control two completely opposing biochemical pathways simultaneously, rapidly and highly efficiently maximizing the liver’s overall chemical and biological output during metabolic stress.
The true physiological brilliance of this specialized form of metabolic regulation lies in its elegant divergence. The single activated kinase simultaneously turns off the cellular storage enzyme, glycogen synthase, and turns on the cellular release enzyme, glycogen phosphorylase. This perfectly coordinated dual action ensures that the liver does not wastefully attempt to store and release glucose simultaneously. The ultimate result is a rapid, instantaneous, and highly powerful one-way flood of vital glucose directly into the bloodstream to rescue the organism.
Slide 15: The Unified Model of Metabolic Regulation
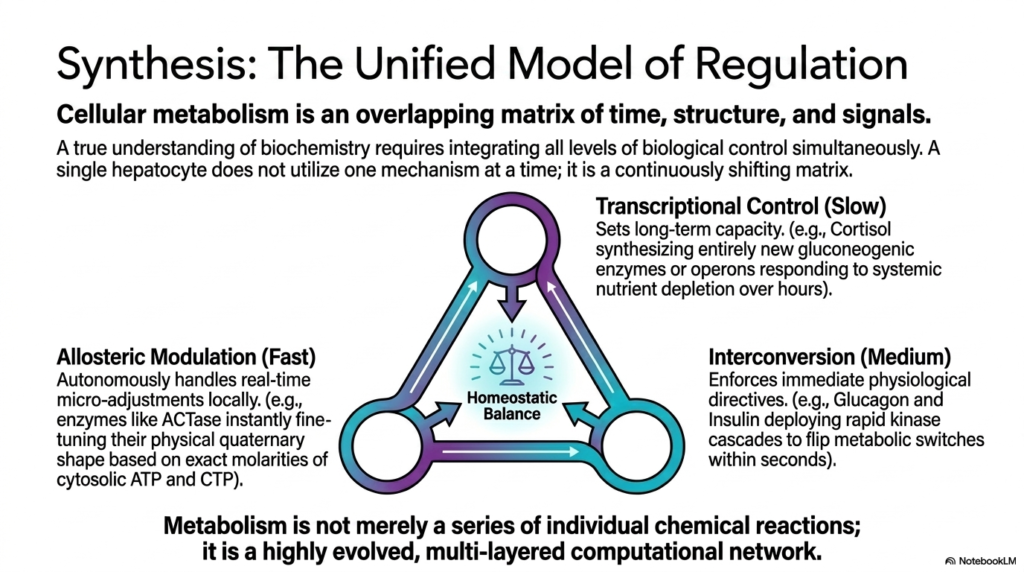
To truly achieve a profound understanding of complex biological biochemistry, students must completely integrate all aspects of metabolic regulation. Cellular metabolism is absolutely not a simple, linear sequence of individual chemical reactions occurring in isolation. Instead, it is a highly evolved, deeply complex, and fully overlapping matrix of time, structural biology, and chemical signals. A single living hepatocyte does not simply use one mechanism of control at a time; it exists in a continuously shifting matrix, balancing immense inputs to maintain perfect cellular homeostasis.
At the foundational core of this unified biological model is the slow, highly deliberate process of transcriptional control. This form of metabolic regulation determines the cell’s long-term metabolic capacity. For instance, when the systemic body is under prolonged severe stress, steroid hormones like cortisol direct the nucleus to synthesize entirely new gluconeogenic enzymes from scratch. While this specific process takes hours to unfold, it fundamentally rescales the cell’s entire baseline biochemical machinery, allowing it to easily survive extended periods of deep systemic nutrient depletion.
Layered entirely directly upon this foundational baseline is the intermediate process of enzyme interconversion. This rapid, medium-term form of metabolic regulation actively enforces immediate physiological directives sent from the whole organism. When the pancreas releases powerful systemic peptide hormones, such as insulin or rapid-acting glucagon, they quickly activate intracellular kinase cascades to flip specific metabolic switches within the cell within mere seconds. This ensures that the cell’s baseline machinery functions exactly in accordance with the immediate physiological needs of the surrounding environment.
Finally, operating flawlessly at the absolute fastest localized level, allosteric modulation functions as the cell’s real-time autonomous micro-adjustment system. This exceptionally rapid mechanism of metabolic regulation allows key structural enzymes like ACTase to instantly fine-tune their physical quaternary shape based entirely on the exact localized molarities of cytosolic substrates and chemical energy molecules like ATP. By seamlessly integrating these three vastly different levels of systemic biological control, the living organism transforms basic chemical reactions into a remarkably robust, highly evolved computational network for biological survival.
Please read our Content Disclaimer Statement.
Check out our social media channels: