28. Lactate Dehydrogenase: Structure, Isoenzymes, and Mechanism
When an athlete sprints toward the finish line, their muscles scream for energy long after local oxygen stores have been depleted. This biological endurance relies on a highly specific molecular engine to sustain cellular respiration and generate energy under immense stress. The following comprehensive presentation details the structural biochemistry, genetic foundations, and clinical pathology of the enzyme responsible for sustaining life during hypoxic conditions. By breaking down these complex macromolecular interactions, students can fully conceptualize how this critical metabolic gatekeeper functions in human health, disease, and cellular survival.
Slide 1: Introduction to Lactate Dehydrogenase Structural Biochemistry

The introductory visual presents the complex macromolecular assembly of Lactate Dehydrogenase, an absolutely essential metabolic enzyme for cellular survival. The rendered molecular image displays a detailed ribbon diagram of the enzyme’s quaternary structure, highlighting its fundamental tetrameric nature. In this structural model, individual monomeric subunits interact and intertwine to create a highly stable biological machine. Grasping the foundational biochemistry of Lactate Dehydrogenase is the critical first step for medical and university students attempting to understand advanced cellular energy mechanics.
Expanding upon this structural overview, one must recognize that Lactate Dehydrogenase serves as a classic biochemical model for understanding how quaternary protein folding dictates physiological function. The highly detailed cyan and purple helices in the visual emphasize the intricate secondary structures required for metabolic viability. Every structural loop of the peptide backbone is evolutionarily conserved to ensure the catalytic pockets remain functional. By studying the complete architecture of Lactate Dehydrogenase, students can appreciate how microscopic molecular interactions scale up to influence human physiology at the systemic level.
Furthermore, this slide purposefully sets a comprehensive academic agenda by directly linking structural biochemistry to its catalytic mechanism and eventual clinical pathology. This triad of topics is essential because minute structural abnormalities in Lactate Dehydrogenase can drastically impair its overall catalytic efficiency. When human tissues undergo severe stress or cellular death, this intact structural tetramer leaks into the bloodstream. Therefore, mastering the baseline topography of this enzyme provides the necessary framework for interpreting complex diagnostic results in medical laboratories.
Slide 2: Lactate Dehydrogenase as the Gatekeeper of Anaerobic Metabolism
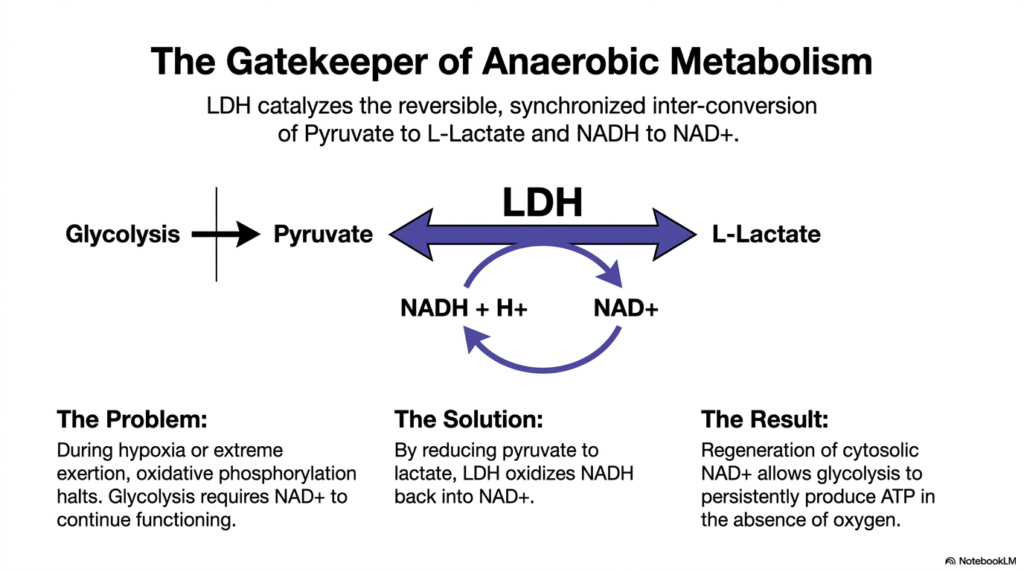
This slide beautifully illustrates why Lactate Dehydrogenase is appropriately titled the gatekeeper of anaerobic metabolism. During severe hypoxia or extreme physical exertion, oxidative phosphorylation in the mitochondria rapidly halts due to oxygen deprivation. Glycolysis, the preliminary pathway for energy production, desperately requires a constant supply of the coenzyme NAD+ to continue producing ATP. Without a mechanism to recycle this coenzyme, cellular energy production would catastrophically collapse, leading to rapid cell death.
To solve this metabolic crisis, Lactate Dehydrogenase catalyzes the reversible, synchronized interconversion of pyruvate to L-lactate. During this highly coordinated reduction reaction, the enzyme simultaneously oxidizes the accumulated NADH back into vital NAD+. This elegant biochemical solution ensures that the cytosolic pool of NAD+ is constantly replenished. Because Lactate Dehydrogenase serves as a metabolic electron sink, glycolysis can continuously produce ATP in the absence of oxygen, sustaining the cell during hypoxic emergencies.
The ultimate result of this reaction is a robust biological failsafe for energy generation. The forward and reverse fluxes through Lactate Dehydrogenase are strictly dictated by the tissue’s immediate metabolic needs and the availability of substrates. By understanding this central enzymatic reaction, students can gain a firm grasp of how cells shift gears between aerobic and anaerobic states, maintaining viability when environmental conditions become hostile to standard oxidative metabolism.
Slide 3: Macromolecular Architecture of the Lactate Dehydrogenase Tetramer
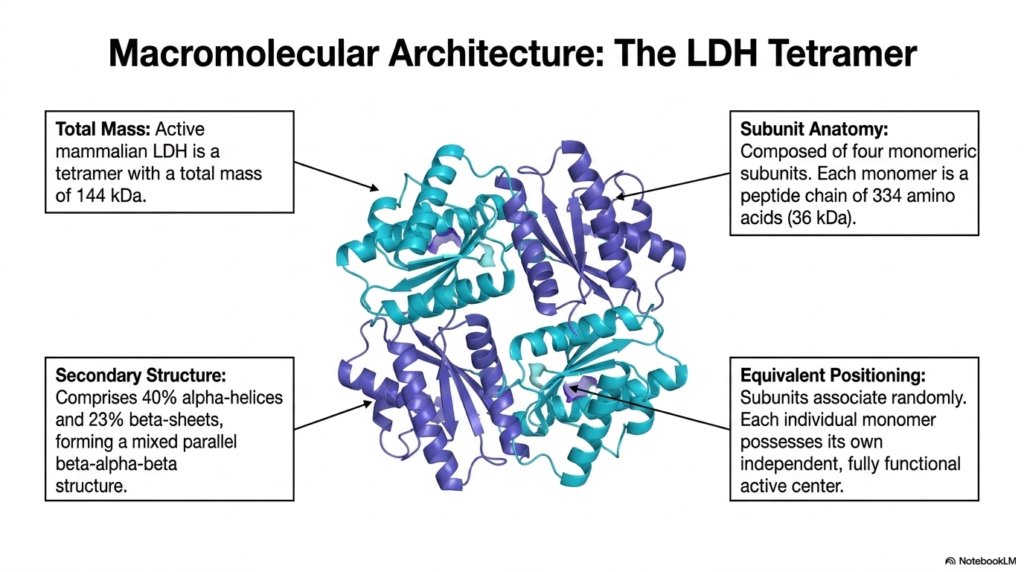
Diving deeper into the physical structure, this slide breaks down the macromolecular architecture of the Lactate Dehydrogenase tetramer. The active mammalian form of this enzyme has a molecular mass of 144 kilodaltons. This massive structure is meticulously assembled from four distinct monomeric subunits, each weighing approximately 36 kilodaltons. Each monomer consists of a lengthy peptide chain of 334 amino acids, folded perfectly to form an active biological catalyst.
The secondary structure of Lactate Dehydrogenase is a masterclass in protein stability, comprising 40% alpha-helices and 23% beta-sheets. These elements fold together to form a highly specific mixed parallel beta-alpha-beta structural motif. This robust configuration provides the necessary thermodynamic stability for the enzyme to operate under fluctuating intracellular conditions. The precise folding of these domains ensures that the enzyme can withstand changes in cellular pH and temperature without instantly denaturing.
A fascinating feature of Lactate Dehydrogenase is the equivalent positioning of its constituent subunits. These monomers associate randomly to form the final tetrameric structure, yet each individual monomer possesses its own fully functional, independent active center. This means a single Lactate Dehydrogenase tetramer can theoretically process four substrate molecules simultaneously. This independent yet cooperative structural alignment maximizes catalytic throughput, ensuring the cell can rapidly convert large amounts of pyruvate under anaerobic stress.
Slide 4: Anatomy of the Lactate Dehydrogenase Catalytic Pocket

The functional heart of Lactate Dehydrogenase lies within its highly specialized catalytic pocket, which relies on three essential amino acid residues and one critical structural loop. The first key player is Arginine-171, a binding residue that features a positively charged guanidinium group. This specific structural component anchors the carboxylate group of the incoming substrate—whether pyruvate or lactate—via strong electrostatic interactions, thereby perfectly orienting the molecule for the impending chemical reaction.
The second major component of the Lactate Dehydrogenase active site is Histidine-195, which is solely responsible for the crucial acid-base catalysis. This specific amino acid acts as the primary proton acceptor or donor, physically changing its electrical charge as it progresses through the catalytic cycle. Arginine-109 works in tandem to stabilize the transition state. During hydride transfer, a highly unfavorable negative charge develops on the substrate’s oxygen atom; Arginine-109 neutralizes this transient instability.
Finally, the catalytic efficiency of Lactate Dehydrogenase is guaranteed by a flexible peptide segment known as Loop 98-111. Upon the successful binding of both the substrate and the coenzyme, this physical loop dynamically swings shut, closing tightly over the active center. This massive conformational change effectively excludes surrounding water molecules from the active site, creating an isolated, non-polar microenvironment. This deliberate exclusion of water is absolutely essential to facilitate the delicate transfer of electrons without unwanted side reactions.
Slide 5: The Lactate Dehydrogenase Catalytic Cycle
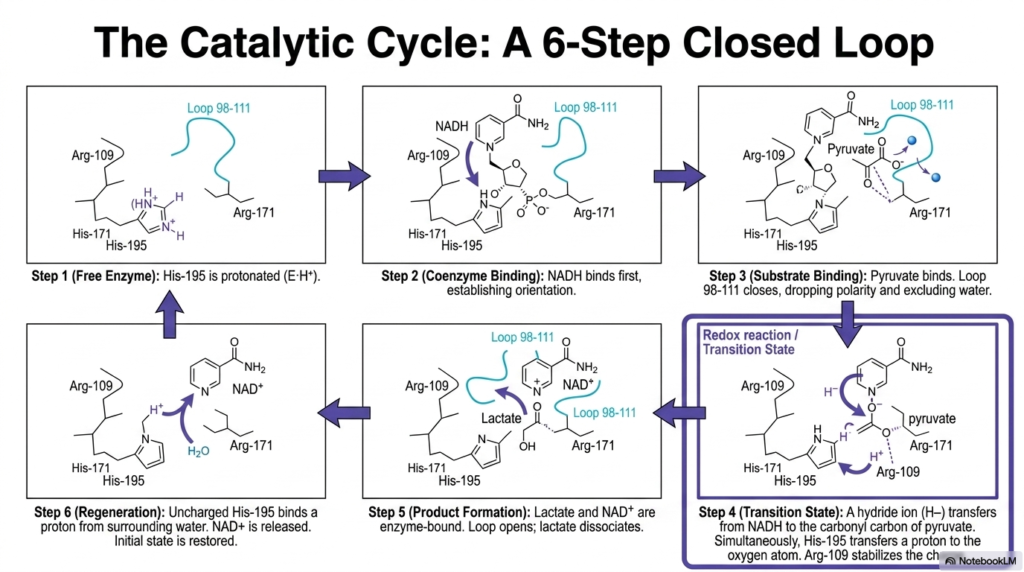
Understanding the kinetics of Lactate Dehydrogenase requires analyzing its elegant six-step closed catalytic cycle. The process begins with the free enzyme in state one, where the crucial Histidine-195 residue is actively protonated and waiting for reactants. In the second step, the coenzyme NADH binds to the active site first. This ordered sequential binding is vital because the massive coenzyme molecule establishes the correct steric orientation required for the subsequent substrate to enter the pocket effectively.
Once the coenzyme is secured, step three involves the binding of pyruvate. It is at this exact moment in the Lactate Dehydrogenase cycle that Loop 98-111 closes aggressively, reducing local polarity and sealing out water. Step four represents the transition state, featuring a highly coordinated redox reaction. A hydride ion transfers directly from the NADH coenzyme to the carbonyl carbon of the pyruvate molecule, while Histidine-195 simultaneously donates its proton to the adjacent oxygen atom, effectively reducing the substrate.
The successful transition state quickly resolves into step five, where the newly formed products, L-lactate and NAD+, briefly remain enzyme-bound. The physical opening of the peptide loop then allows the final L-lactate molecule to dissociate into the cellular cytoplasm. In the final, sixth step of the Lactate Dehydrogenase sequence, the now-uncharged Histidine-195 residue extracts a proton from the surrounding aqueous environment. The NAD+ molecule is released, and the initial baseline state is flawlessly restored.
Slide 6: Genetic Foundations of Lactate Dehydrogenase Subunits
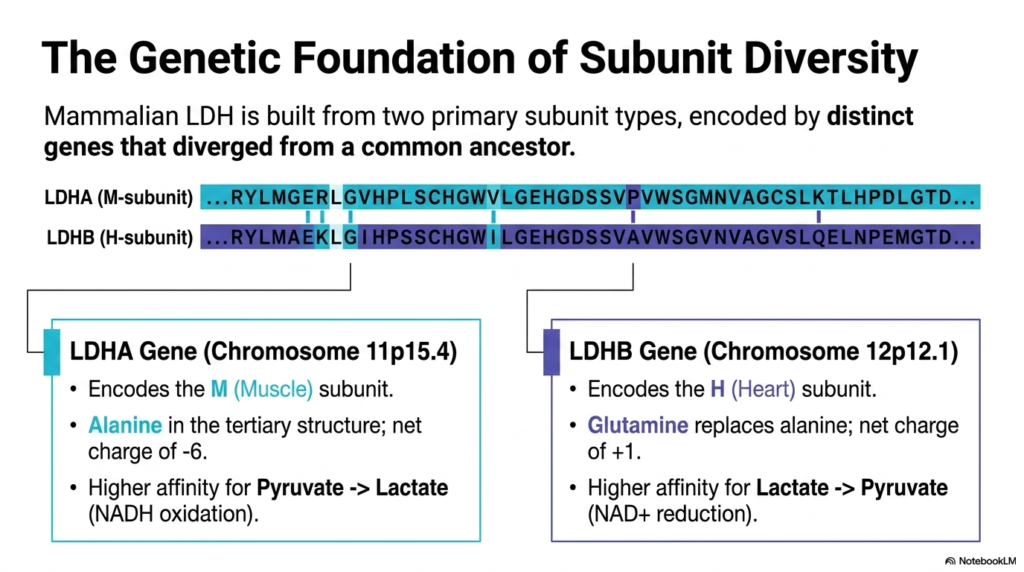
The immense structural diversity of mammalian Lactate Dehydrogenase is entirely rooted in its complex genetic basis. The enzyme is composed of two primary subunit types, each encoded by distinct genes that diverged from a common evolutionary ancestor millions of years ago. By using different genetic instructions, human cells can tailor the specific physical and electrical properties of the resulting enzyme to meet the unique metabolic demands of different tissue types throughout the body.
The first major genetic contributor is the LDHA gene, strategically located on Chromosome 11. This gene encodes the “M” (Muscle) subunit of Lactate Dehydrogenase. A defining biochemical feature of the M subunit is the presence of the amino acid alanine within its tertiary structure, which grants the monomer a distinct net charge of negative six. Consequently, tetramers dominant in M subunits exhibit a much higher catalytic affinity for converting pyruvate into lactate, facilitating rapid NADH oxidation.
Conversely, the LDHB gene, located on Chromosome 12, encodes the “H” (Heart) subunit of Lactate Dehydrogenase. Through minor evolutionary sequence divergence, the H subunit replaces the aforementioned alanine with a glutamine residue. This seemingly tiny substitution drastically alters the overall net charge to a positive one. Because of this altered electrostatic profile, H-dominant subunits demonstrate a much higher affinity for the reverse reaction, preferring to convert lactate back into pyruvate while reducing NAD+.
Slide 7: The Lactate Dehydrogenase Isoenzyme Matrix
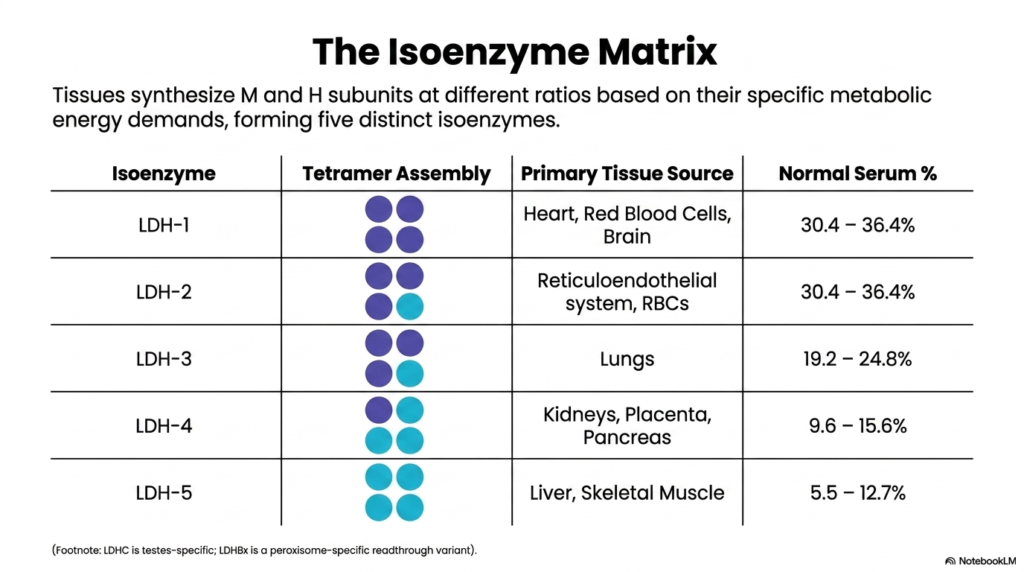
Because tissues synthesize M and H subunits at varying rates, Lactate Dehydrogenase exists in the biological system as a five-part isoenzyme matrix. The random tetrameric assembly of these two distinct monomer types yields five specific isoenzymes, ranging from LDH-1 to LDH-5. Tissues strategically dictate the exact ratio of these newly synthesized subunits based entirely on their local metabolic energy demands, creating a highly customized enzymatic profile that perfectly supports either heavily aerobic or strictly anaerobic cellular environments.
At one end of the spectrum is LDH-1, a tetramer composed of four distinct H subunits. This specific Lactate Dehydrogenase isoenzyme accounts for approximately 30% of normal blood serum and is highly concentrated in the heart, brain, and red blood cells—tissues that rely on constant aerobic metabolism. At the opposite end lies LDH-5, comprising four M subunits. This specific isoenzyme is predominantly localized in the liver and skeletal muscle, which require massive bursts of anaerobic energy.
The intermediate isoenzymes of Lactate Dehydrogenase complete this diverse physiological matrix. LDH-2 contains three H subunits and one M subunit and is the dominant form in the reticuloendothelial system. LDH-3 is evenly split with two of each subunit and is primarily sourced from pulmonary tissues. Finally, LDH-4, containing one H and three M subunits, is predominantly found within the kidneys and pancreas. This strict compartmentalization of isoenzymes is biologically brilliant and clinically invaluable.
Slide 8: Gel Electrophoresis and Lactate Dehydrogenase Separation
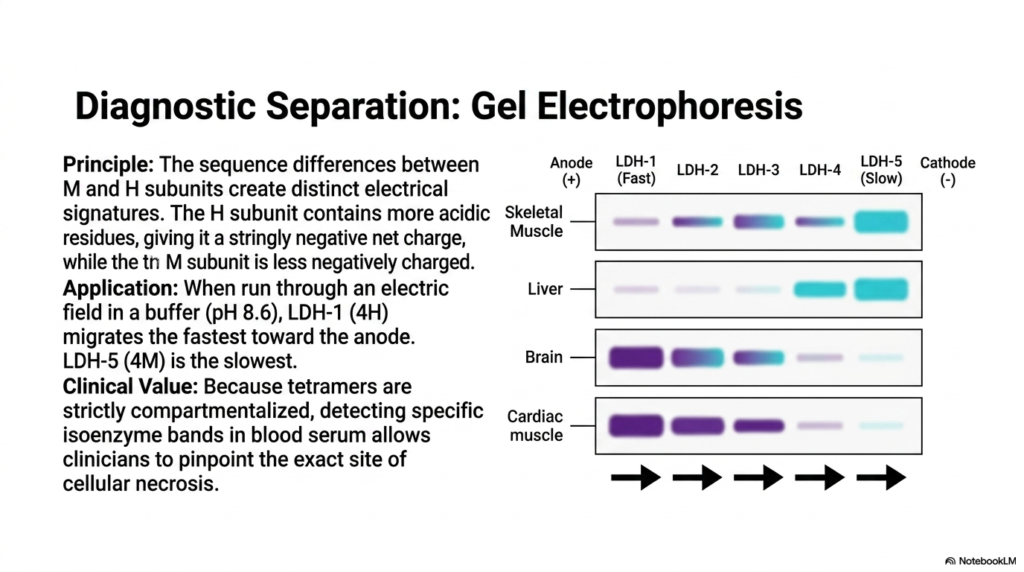
The diagnostic power of Lactate Dehydrogenase relies on our ability to isolate its different forms using gel electrophoresis. The foundational principle of this clinical technique relies heavily on the sequence differences between the M and H subunits. Because the H subunit contains a higher concentration of acidic amino acid residues, it naturally possesses a strongly negative net charge. In stark contrast, the M subunit is significantly less negatively charged, creating distinct electrical signatures for every single tetrameric combination.
When a patient’s serum sample is exposed to an electric field in a standardized buffer solution at pH 8.6, the Lactate Dehydrogenase isoenzymes separate based on their charges. LDH-1, packed with four highly negative H subunits, migrates the absolute fastest toward the positively charged anode. Conversely, LDH-5, composed of four M subunits, migrates the slowest across the gel matrix. The intermediate isoenzymes space themselves out perfectly between these two defined extremes.
The clinical value of separating Lactate Dehydrogenase via electrophoresis cannot be overstated. Because these specific tetramers are strictly compartmentalized within different organs, detecting heavily elevated bands of a specific isoenzyme in blood serum allows astute clinicians to pinpoint the exact anatomical site of cellular necrosis. If the fast-moving band suddenly spikes on a laboratory readout, medical professionals immediately know that specialized aerobic tissues have recently ruptured and spilled their contents into the patient’s bloodstream.
Slide 9: The Cori Cycle and Lactate Dehydrogenase Systemic Metabolism
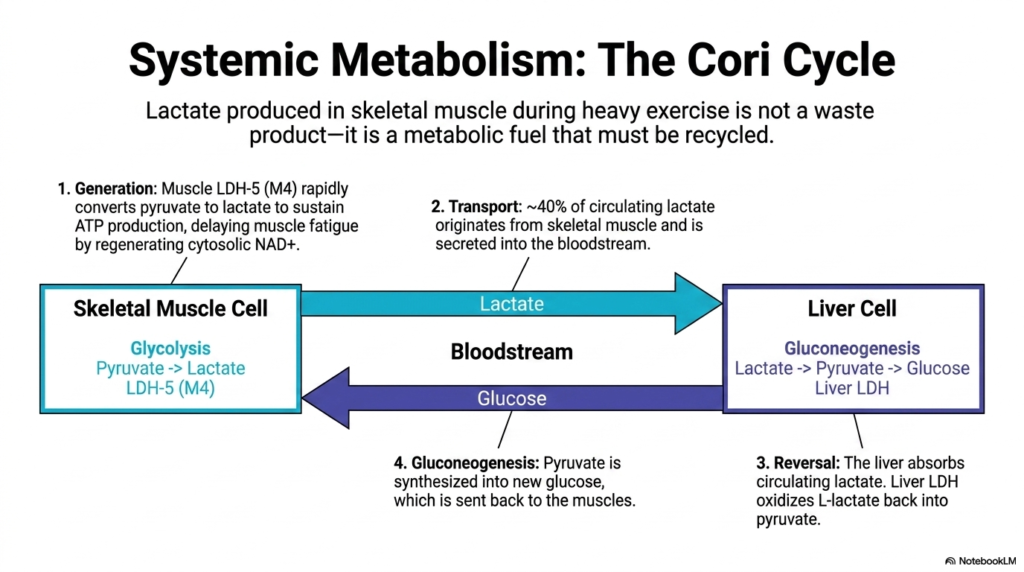
Moving from the micro to the macro, this slide highlights how Lactate Dehydrogenase drives systemic metabolism through the Cori Cycle. It is a common misconception that the lactate produced in skeletal muscle during heavy exercise is merely a toxic waste product. In reality, it is a highly valuable metabolic fuel that must be systematically recycled. During intense exertion, muscle-specific LDH-5 rapidly converts pyruvate to lactate, thereby delaying muscle fatigue by continuously regenerating cytosolic NAD+ to sustain glycolysis.
Once generated, approximately 40% of this circulating lactate originates from these working skeletal muscles and is actively secreted into the human bloodstream. This systemic transport mechanism brings the rich metabolic fuel directly to the liver. Here, specialized hepatic cells absorb the circulating lactate, and liver-specific Lactate Dehydrogenase reverses the reaction. The enzyme efficiently oxidizes the incoming L-lactate back into usable pyruvate, harvesting the molecule’s stored potential for the next phase of systemic energy production.
The final stage of this Lactate Dehydrogenase-driven cycle involves massive hepatic gluconeogenesis. The liver takes the newly formed pyruvate and heavily invests cellular energy to synthesize fresh glucose molecules. This newly manufactured glucose is then secreted back into the bloodstream and shuttled directly to the exhausted skeletal muscles. This elegant, systemic recycling loop ensures that the human body extracts the maximum possible energy from every single glucose molecule during physiological stress.
Slide 10: Modulating Lactate Dehydrogenase Catalytic Activity
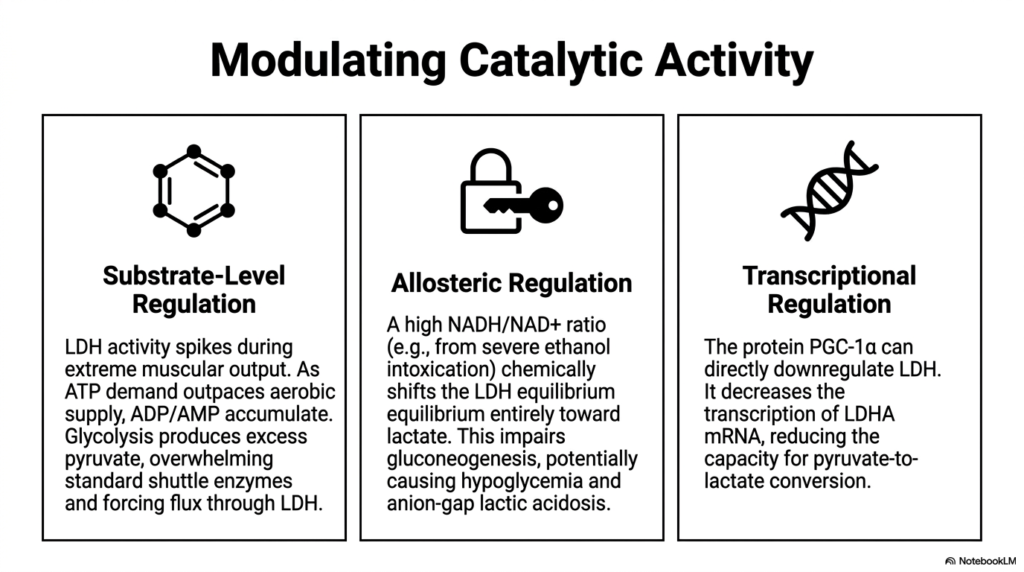
The catalytic activity of Lactate Dehydrogenase is not static; it is heavily modulated by multiple dynamic cellular mechanisms. The first layer is substrate-level regulation, which simply follows the laws of mass action. During extreme muscular output, ATP demand heavily outpaces the standard aerobic supply, causing ADP and AMP to accumulate rapidly. This forces glycolysis into overdrive, producing an immense excess of pyruvate, overwhelming standard mitochondrial shuttle enzymes, and physically driving metabolic flux heavily through the enzyme.
The second critical layer involves the allosteric regulation of Lactate Dehydrogenase. The enzyme’s equilibrium is highly sensitive to the cellular NADH-to-NAD+ ratio. For example, during severe ethanol intoxication, the liver rapidly generates massive amounts of NADH. This chemically shifts the enzyme’s equilibrium entirely toward lactate production. This dangerous biochemical shift actively impairs standard gluconeogenesis, which can tragically lead to severe clinical manifestations like profound hypoglycemia and an anion-gap lactic acidosis.
The final regulatory mechanism governing Lactate Dehydrogenase operates at the deep transcriptional level. Specific cellular proteins, such as PGC-1alpha, possess the unique ability to directly downregulate the genetic expression of this enzyme. By actively decreasing the cellular transcription of LDHA mRNA, the cell intentionally reduces its overall physical capacity for pyruvate-to-lactate conversion. This sophisticated genetic control allows the body to permanently remodel a tissue’s metabolic profile, shifting it away from fermentation and toward oxidative endurance.
Slide 11: Serum Pathology and the Flipped Lactate Dehydrogenase Pattern
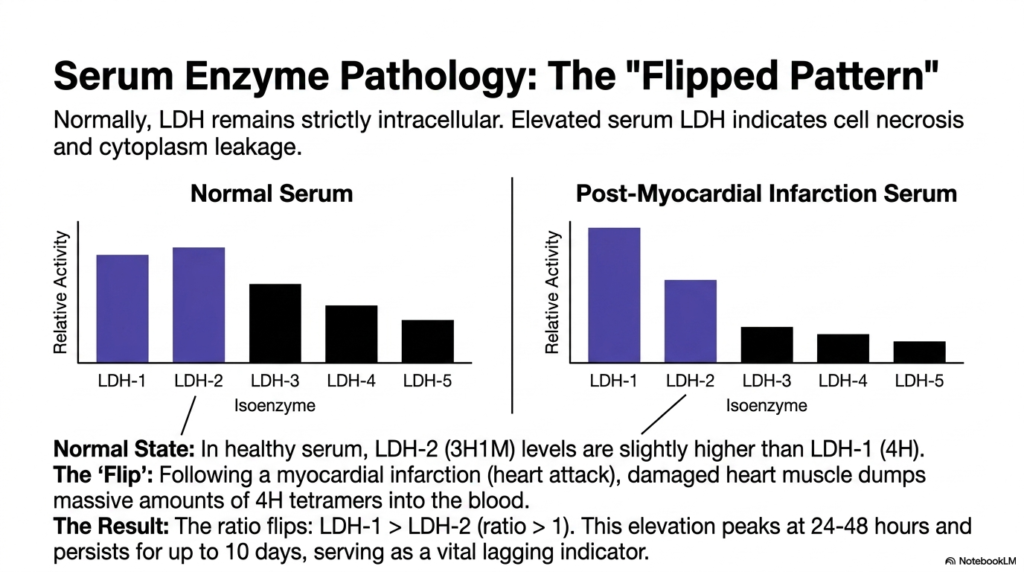
In standard diagnostic medicine, Lactate Dehydrogenase is widely regarded as a premier biomarker for detecting occult tissue pathology. Normally, this massive tetrameric enzyme remains strictly compartmentalized within the intracellular space, performing its metabolic duties away from the blood. However, when an organ experiences severe ischemia or trauma, the resulting cellular necrosis literally tears the cell membrane apart. This catastrophic structural failure causes large quantities of intact cytoplasmic enzymes to leak directly into the surrounding systemic circulation.
The most famous clinical manifestation of this cellular leakage is the “Flipped Pattern” of Lactate Dehydrogenase isoenzymes. In a completely healthy, normal serum sample, the concentration of LDH-2 is predictably slightly higher than that of LDH-1. However, following a massive myocardial infarction, the severely damaged heart muscle aggressively dumps massive quantities of LDH-1 into the bloodstream. This rapid influx drastically alters the standard physiological ratio, causing LDH-1 levels to quickly surpass those of LDH-2.
When a laboratory analysis confirms that the LDH-1-to-LDH-2 ratio is greater than 1, the test has officially “flipped.” This specific elevation of Lactate Dehydrogenase peaks at roughly 24 to 48 hours after the initial cardiac event and notoriously persists in the blood for up to 10 days. Because of this prolonged clearance time, the flipped ratio serves as an absolutely vital lagging clinical indicator, allowing physicians to confidently diagnose heart attacks that occurred several days prior.
Slide 12: Diagnostic Fluid Testing with Lactate Dehydrogenase
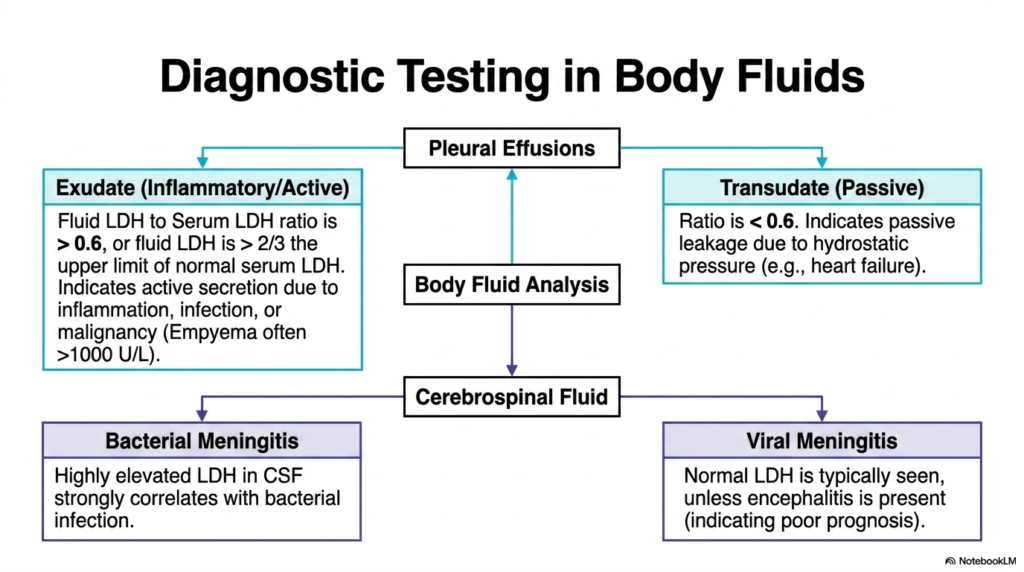
Beyond standard serum analysis, the concentration of Lactate Dehydrogenase is incredibly useful for diagnostic testing in various isolated body fluids. When a patient presents with a pleural effusion—an abnormal collection of fluid surrounding the lungs—physicians must determine if the fluid is an active exudate or a passive transudate. By carefully measuring the specific ratio of fluid enzyme to serum enzyme, clinicians can immediately uncover the underlying pathological mechanism driving the dangerous fluid accumulation.
If the Lactate Dehydrogenase fluid-to-serum ratio is greater than 0.6, or if the absolute fluid value exceeds two-thirds the upper limit of normal serum levels, the effusion is classified as an exudate. This actively implies that the fluid secretion is being driven by severe local inflammation, a raging infection, or an invasive malignancy. Conversely, a ratio below 0.6 indicates a transudate, suggesting that the fluid is passively leaking due to altered systemic hydrostatic pressure, commonly seen in congestive heart failure.
This versatile enzyme is similarly diagnostic for cerebrospinal fluid (CSF) analysis in suspected neurological infections. In cases of severe bacterial meningitis, local tissue destruction leads to markedly elevated levels of Lactate Dehydrogenase in the delicate CSF. Alternatively, if a patient has straightforward viral meningitis, the enzyme levels typically remain normal. Thus, measuring this exact biochemical marker helps desperate emergency physicians rapidly differentiate between a treatable bacterial infection and a viral pathology.
Slide 13: The Warburg Effect and Lactate Dehydrogenase in Cancer
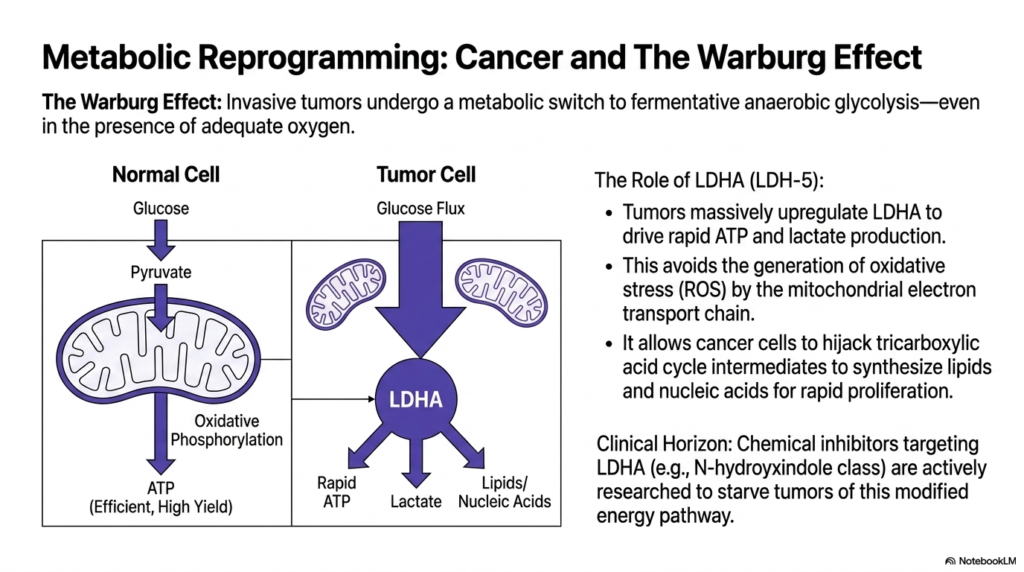
The study of oncology reveals how invasive cancers weaponize Lactate Dehydrogenase through metabolic reprogramming. Discovered nearly a century ago, the famous Warburg Effect describes how aggressive tumors undergo a radical metabolic switch to favor fermentative anaerobic glycolysis, astonishingly doing so even in the presence of totally adequate oxygen supplies. Instead of utilizing the highly efficient mitochondrial oxidative phosphorylation pathway, cancerous cells intentionally short-circuit their metabolism to prioritize sheer speed and raw biomass synthesis.
To execute this malicious metabolic hijacking, tumors massively upregulate LDHA expression, flooding their cytoplasm with Lactate Dehydrogenase. This specific isoenzyme shift intentionally drives the ultra-rapid production of both ATP and lactate. By intentionally avoiding standard mitochondrial respiration, cancer cells cleverly bypass the generation of dangerous reactive oxygen species (ROS) that could otherwise trigger cellular apoptosis. The resulting highly acidic lactate is then exported, creating a toxic microenvironment that severely suppresses the patient’s local immune system.
Furthermore, this immense reliance on Lactate Dehydrogenase allows malignant cancer cells to efficiently hijack standard tricarboxylic acid cycle intermediates. Instead of burning these carbon molecules for energy, the tumor diverts them toward the aggressive synthesis of the massive quantities of lipids and nucleic acids required for rapid cellular proliferation. Recognizing this specific vulnerability, modern pharmacological researchers are actively developing chemical inhibitors targeting the LDHA subunit to actively starve dangerous tumors of their preferred modified energy pathway.
Slide 14: Genetic Deficiencies in Lactate Dehydrogenase
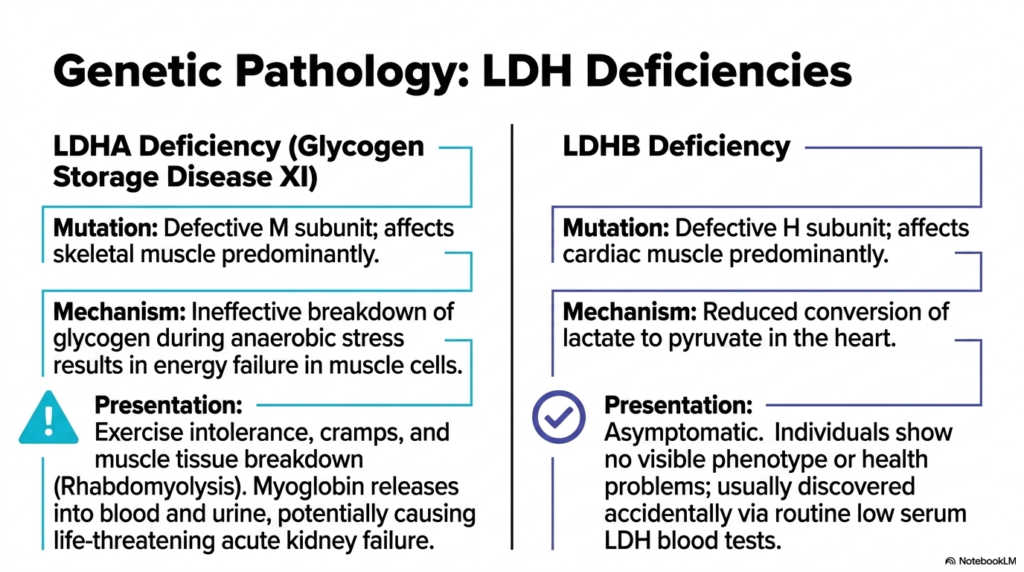
While incredibly rare, inherited genetic deficiencies in Lactate Dehydrogenase provide exceptional insights into human metabolic boundaries. A targeted mutation affecting the LDHA gene results in a defective M subunit, leading to a condition known as Glycogen Storage Disease XI. Because the M subunit is highly expressed in skeletal muscle, this specific genetic pathology predominantly impairs the function of normal muscle tissue. The defective enzymes cause a massive failure to properly break down stored glycogen during routine anaerobic stress.
Patients suffering from an LDHA deficiency present with profound exercise intolerance, agonizing muscle cramps, and dangerous muscle tissue breakdown during intense physical activity. This rapid cellular destruction, termed rhabdomyolysis, actively releases massive amounts of toxic myoglobin directly into the patient’s blood and urine. If not managed carefully by medical professionals, this secondary release triggered by a lack of functional Lactate Dehydrogenase can effortlessly cause life-threatening, irreversible acute kidney failure due to severe tubular blockage.
In stark contrast, a genetic mutation resulting in an LDHB deficiency affects the structural integrity of the H subunit, predominantly impacting healthy cardiac muscle. Because this mutation reduces the efficiency of converting systemic lactate back into pyruvate in the heart, one might expect catastrophic cardiovascular consequences. Surprisingly, patients with this specific Lactate Dehydrogenase deficiency remain entirely asymptomatic. Their unique phenotype is typically discovered only incidentally by clinicians performing routine blood panels that reveal inexplicably low baseline serum enzyme activity.
Slide 15: Synthesizing the Lactate Dehydrogenase System Map
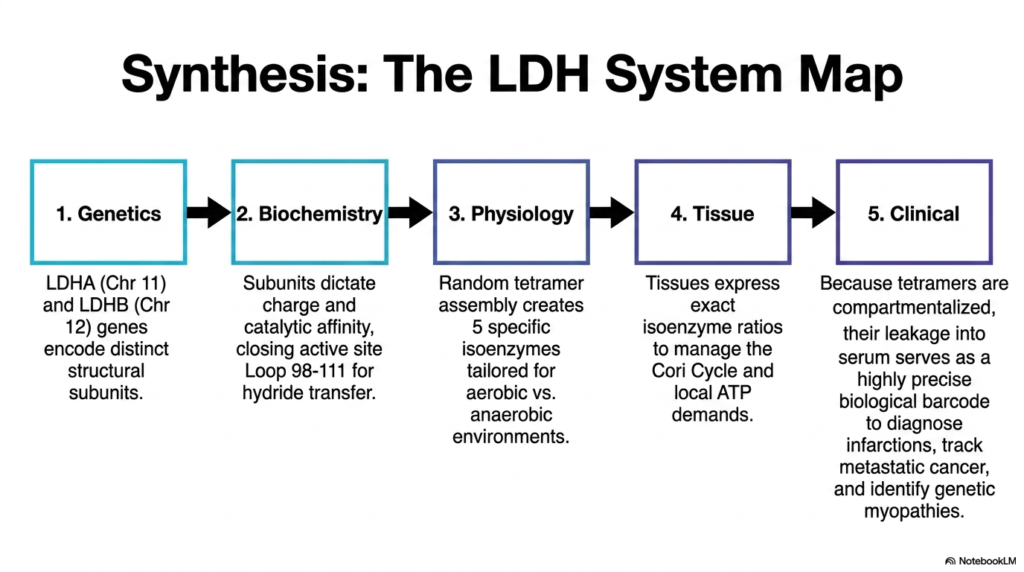
The final slide synthesizes the entire Lactate Dehydrogenase system map, perfectly connecting abstract molecular genetics to tangible clinical realities. At the foundational level, exact genetic instructions housed on Chromosomes 11 and 12 dictate the precise creation of distinct structural subunits. These baseline genetic codes are the ultimate blueprint for the enzyme, determining exactly how the resulting protein will interact with the complex intracellular environment of the human body.
In advanced biochemistry, these exact subunits completely determine the final electrical charge and catalytic affinity of the assembled tetramer. By specifically closing the active site’s protective structural loop around incoming substrates, the enzyme ensures the flawless transfer of vital hydride ions. This highly orchestrated physical biochemistry is precisely what enables Lactate Dehydrogenase to perfectly regulate the massive, systemic Cori Cycle, seamlessly shuttling energetic potential between different organ systems based on their immediate physiological demands.
Ultimately, the strict, deliberate tissue compartmentalization of Lactate Dehydrogenase is what makes it a cornerstone of modern pathology. Because different human tissues express specific isoenzyme ratios to meet their local ATP demands, the sudden leakage of these tetramers into standard blood serum serves as a highly precise biological diagnostic barcode. By carefully deciphering this specific enzymatic barcode, medical professionals can accurately diagnose large infarctions, closely monitor metastatic cancers, and correctly identify rare genetic myopathies in patients.
Please read our Content Disclaimer Statement.
Check out our social media channels: