27. Enzyme Inhibition Explained: Mechanisms, Types and Kinetics
Imagine a bustling city without traffic lights; pure chaos would ensue instantly. In the cellular world, biological catalysts are the speeding vehicles driving metabolic reactions, and molecular regulators act as the vital traffic signals preventing catastrophe. This comprehensive slide deck provides an in-depth exploration of these precise regulatory mechanisms. Designed for medical and college students, this guide deconstructs complex kinetic models and molecular structures, revealing how biochemical control dictates physiological health and drives targeted pharmacological drug design.
Slide 1: Foundations of Enzyme Inhibition

The study of biochemistry demands a rigorous understanding of molecular regulators. Among the most critical of these mechanisms is Enzyme Inhibition, a process that dictates how biological catalysts are modulated within living systems. The introductory slide sets the stage for this vital topic, highlighting the intersection of molecular mechanisms, mathematical kinetic models, and practical pharmacological applications. By mastering these foundational concepts, students gain the tools to effectively decipher complex metabolic networks.
The visual representation on this slide cleverly illustrates the precise, puzzle-like nature of biological molecules. When exploring Enzyme Inhibition, visualizing the physical interplay between a highly specific protein structure and its regulatory counterpart is absolutely essential. This graphic emphasizes that biological regulation is not merely a conceptual abstraction but a tangible, structural reality governed by molecular geometry. Such visual metaphors are crucial for anchoring the theoretical aspects of kinetic behavior into physical reality.
Furthermore, the overarching themes presented here preview the depth of Enzyme Inhibition required for advanced pharmacology and medical interventions. The intricate dance between catalysts and blocking agents forms the basis for nearly all modern drug design and toxicological studies. Throughout the subsequent materials, the journey will smoothly transition from fundamental binding interactions to complex mathematical proofs, equipping future scientists with robust knowledge.
Understanding the full scope of these mechanisms provides a window into both physiological health and pathological disease states. Without these precise regulatory interventions, metabolic pathways would run unchecked, leading to cellular toxicity or energetic depletion. This opening overview acts as a structured roadmap, promising a rigorous dive into the specific subtypes of regulators that govern cellular life.
Slide 2: Structural Blueprints of Enzyme Inhibition
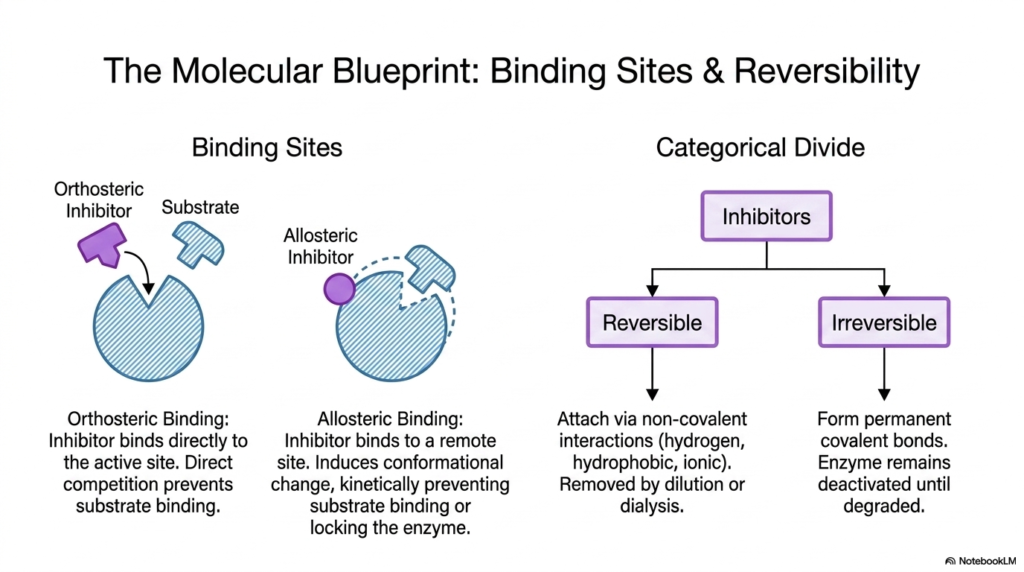
Diving into the mechanics of cellular control, one must first classify the physical locations where regulatory molecules attach. The concept of Enzyme Inhibition is primarily divided into binding-site geography and the permanence of the molecular interaction. The slide elegantly distinguishes between orthosteric binding, in which a molecule directly competes for the active site, and allosteric binding, which occurs at a remote, secondary site on the protein complex.
Understanding these binding sites is critical for mastering Enzyme Inhibition. When an allosteric regulator binds, it induces a conformational change throughout the protein. This structural shift kinetically prevents normal substrate binding or functionally locks the catalyst in an inactive state. This distal control mechanism highlights the incredible flexibility and dynamic nature of biological proteins in aqueous cellular environments.
Beyond spatial geography, the categorical divide between reversible and irreversible mechanisms dictates the physiological lifespan of Enzyme Inhibition. Reversible regulators attach via noncovalent interactions, such as hydrogen bonds, hydrophobic interactions, or ionic interactions. Because these bonds are transient, the blocking molecules can be easily removed by simple cellular dilution or laboratory dialysis, restoring normal catalytic function without requiring new protein synthesis.
Conversely, irreversible regulators form permanent covalent bonds with specific amino acid residues. This severe form of interaction leaves the protein permanently deactivated until the cell degrades and recycles the entire complex. Recognizing this categorical divide is fundamental for pharmacology students, as it determines whether a therapeutic drug will provide temporary relief or long-lasting biological alteration.
Slide 3: Competitive Enzyme Inhibition and Active Site Battles
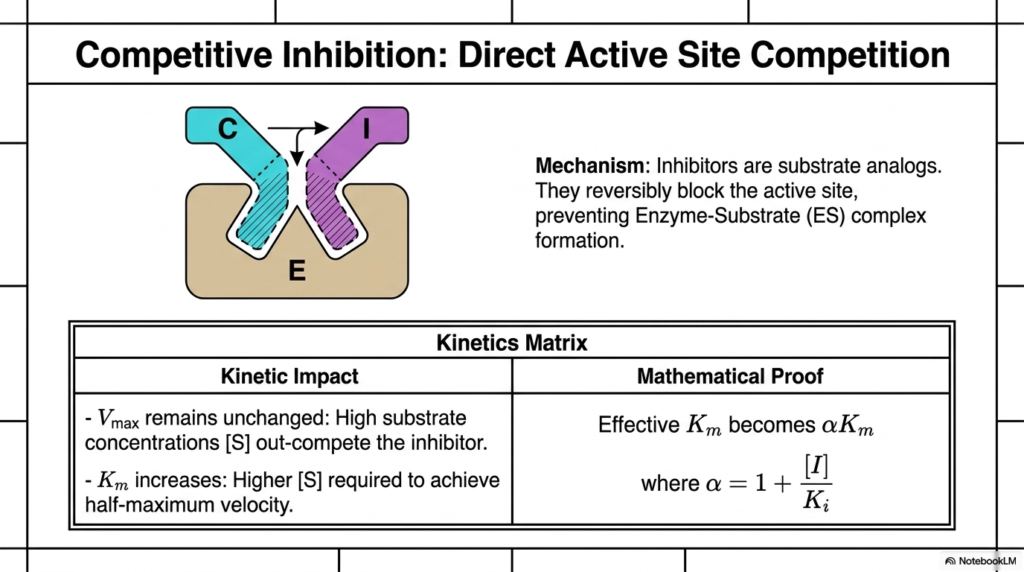
The most intuitive form of molecular regulation is direct active site competition. In Competitive Enzyme Inhibition, the regulatory molecules function as structural analogs to the natural substrate. Because they mimic the native molecule’s shape and charge distribution, they bind reversibly and successfully to the active site, physically blocking the formation of the normal reaction complex. This creates a direct, concentration-dependent molecular tug-of-war.
The kinetics matrix provided on the slide clearly outlines the mathematical impact of this direct competition. Notably, the reaction’s maximum velocity remains unchanged. If the surrounding environment contains an overwhelming concentration of the native substrate, those molecules can outcompete the regulators through sheer numbers, thereby achieving the original maximum catalytic rate.
However, the presence of Competitive Enzyme Inhibition significantly alters the binding affinity metric. The binding constant increases, meaning a much higher concentration of normal substrate is required to achieve half of the maximum reaction velocity. This mathematical shift reflects the diminished probability of a successful catalytic collision when a decoy molecule constantly occupies the target binding pocket.
The mathematical proof provided on the slide introduces the modifying factor alpha, which directly correlates with the concentration of the regulatory molecule. By increasing the effective binding constant by a factor of alpha, researchers can precisely calculate the amount of extra substrate required to overcome the blockade. This exact mathematical relationship is widely used to calculate optimal dosages for competitive pharmaceutical drugs.
Slide 4: Uncompetitive Enzyme Inhibition and the ES Complex
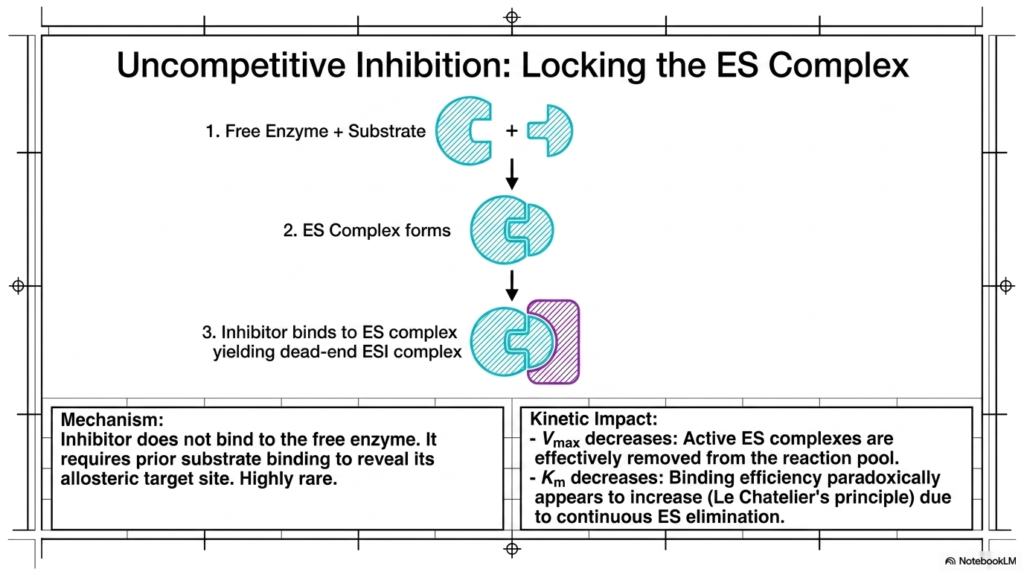
Moving away from direct active-site battles, students encounter a highly rare yet fascinating regulatory mechanism. Uncompetitive Enzyme Inhibition occurs when the regulatory molecule strictly refuses to interact with the free, unbound protein. Instead, this specialized regulator requires the native substrate to bind first, thereby revealing a hidden allosteric target site specifically designed for the blocking agent.
This sequential binding requirement creates a dead-end molecular complex that completely halts catalytic progression. In Uncompetitive Enzyme Inhibition, the active complex is essentially trapped. Because the native substrate is locked inside and the reaction cannot proceed, the overall pool of functional, active reaction centers is effectively removed from the cellular environment, leading to severe kinetic consequences.
The kinetic impact of this phenomenon is highly distinct and mathematically intriguing. First, the reaction system’s maximum velocity decreases significantly. Because the locked dead-end complexes cannot be salvaged by adding more native substrate, the total functional capacity of the biological system is permanently reduced for as long as the regulator remains attached.
Paradoxically, the binding affinity constant also decreases, which normally implies higher efficiency. This occurs due to Le Chatelier’s principle; the continuous elimination of the active complex pulls the equilibrium forward, making it appear as though the initial binding event is highly efficient. This unique signature makes this form of Enzyme Inhibition easily identifiable during advanced kinetic laboratory testing.
Slide 5: Non-Competitive Enzyme Inhibition and Equal Affinity
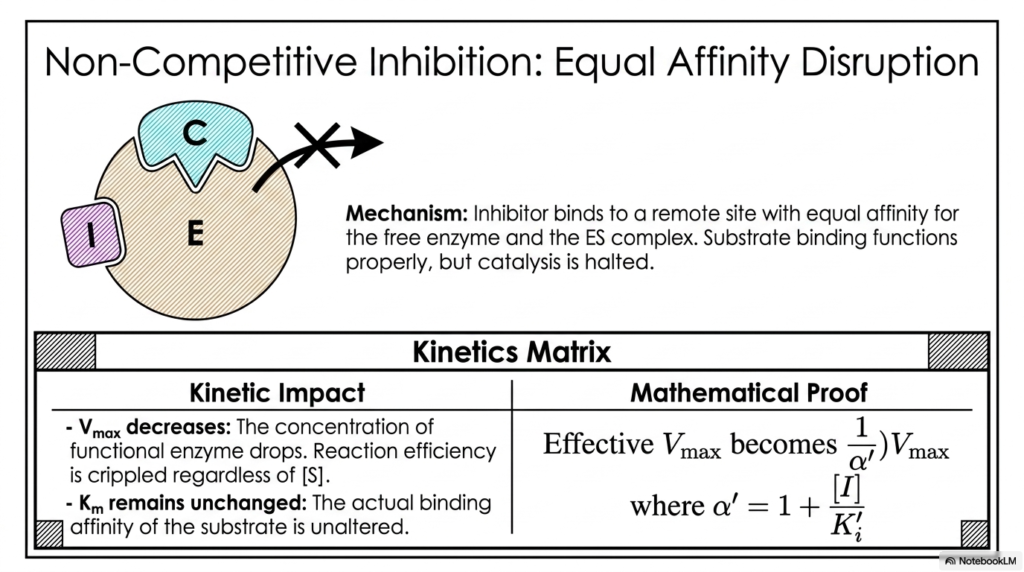
Another critical permutation of remote-site regulation involves molecules that do not care whether the target protein is currently occupied or empty. Non-Competitive Enzyme Inhibition occurs when a regulator binds to an allosteric site with perfectly equal affinity for both the free protein and the actively bound complex. The native substrate can still bind without issue, but the catalytic machinery is completely disabled.
Because the regulatory molecule disrupts the actual chemical reaction rather than the physical docking process, Non-Competitive Enzyme Inhibition drastically cripples reaction efficiency regardless of the surrounding environment. Pouring massive amounts of native substrate into the system will not rescue the reaction rate, as the internal catalytic residues are structurally misaligned and rendered entirely non-functional.
The kinetic matrix on this slide clearly demonstrates this devastating impact. The maximum velocity decreases in proportion to the concentration of the regulatory molecule. The functional protein concentration plummets, mathematically demonstrating that the biological system’s overall catalytic ceiling has been permanently lowered while the regulatory molecules remain bound.
Interestingly, because the initial docking process remains physically unhindered, the binding affinity constant remains entirely unchanged. The mathematical proof modifies only the maximum velocity using the alpha factor, reflecting a pure reduction in catalytic turnover without altering the initial molecular attraction. This distinct kinetic profile allows researchers to easily differentiate this mechanism during pharmacological screening.
Slide 6: Mixed Enzyme Inhibition and Asymmetric Hybrids

In physiological reality, pure regulatory mechanisms are somewhat rare, leading to the importance of hybrid models. Mixed Enzyme Inhibition represents an asymmetric blend of both competitive and non-competitive traits. These regulators bind to allosteric sites and alter the protein’s tertiary structure, but, crucially, their binding affinity varies with the target’s current occupancy state.
This asymmetry is the defining hallmark of Mixed Enzyme Inhibition. The dissociation constant for the free protein is mathematically unequal to the dissociation constant for the occupied complex. This means the regulatory molecule inherently prefers one structural state over the other, introducing a complex layer of mathematical variability into the overall kinetic equation.
The kinetic impact predictably features a decrease in maximum velocity, driven by the non-competitive characteristic that inevitably hampers the active complex. Because the three-dimensional shape is distorted, the catalytic turnover rate inevitably suffers, permanently lowering the reaction ceiling much as a pure non-competitive mechanism would dictate.
However, the binding affinity constant will shift dynamically. It may increase or decrease depending entirely on whether the specific regulator favors the empty state or the occupied state. If competitive dominance prevails, the affinity constant increases; if uncompetitive dominance prevails, the affinity constant decreases. This nuanced form of Enzyme Inhibition requires sophisticated laboratory analysis to accurately diagnose and model.
Slide 7: Allosteric Enzyme Inhibition in Oligomeric Proteins
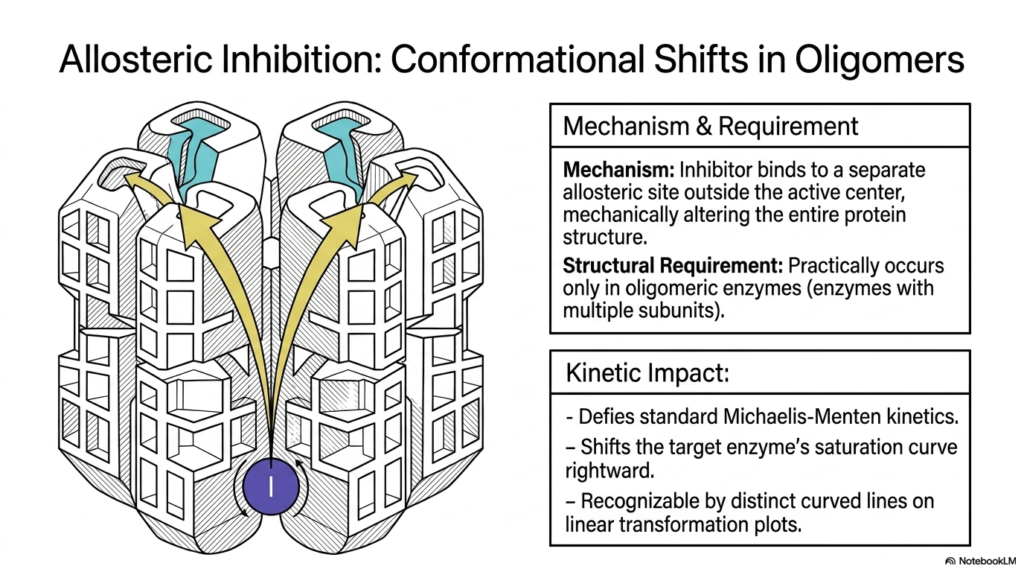
Moving beyond single-subunit models, complex metabolic regulation relies heavily on large, multi-subunit protein complexes. Allosteric Enzyme Inhibition in this context involves regulatory molecules binding outside the active center, but the mechanical alteration ripples through a massive quaternary structure. This structural requirement means that these specific mechanisms occur in practice only in oligomeric complexes with multiple interacting subunits.
When studying Enzyme Inhibition in these massive structures, one observes that binding at a single distant site triggers a cascade of conformational shifts. This cooperative mechanical movement alters the geometry of all associated active sites simultaneously. This allows a single regulatory molecule to exert profound, amplified control over the entire multi-subunit complex, acting as a highly sensitive biological switch.
The kinetic impact of this cooperative behavior is profound, as it defies standard classical kinetic theory. Instead of following predictable hyperbolic curves, these systems exhibit highly complex sigmoidal saturation curves. The regulatory molecule shifts these unique S-shaped curves significantly to the right, indicating a massive decrease in overall systemic affinity.
Recognizing this specific type of Enzyme Inhibition is crucial for medical students analyzing systemic metabolic pathways. When plotting linear transformations of these reactions, the data yields distinct, curved diagnostic lines rather than straight gradients. This unique mathematical signature confirms the presence of cooperative allosteric regulation, a hallmark of critical regulatory checkpoints in human metabolism.
Slide 8: Visualizing Enzyme Inhibition via Hyperbolic Plots
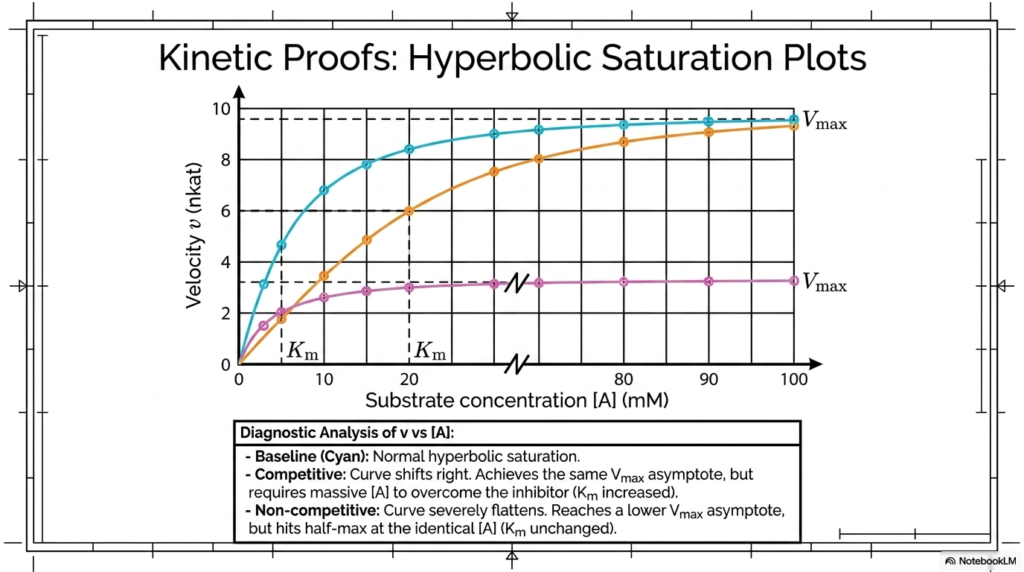
Translating mathematical theory into visual data is a cornerstone of biochemical analysis. This slide demonstrates how researchers utilize hyperbolic saturation plots to diagnose the specific physical nature of molecular regulators. By graphing reaction velocity against substrate concentration, scientists can create clear visual evidence that distinguishes among different mechanisms of Enzyme Inhibition.
The baseline cyan curve represents normal, unhindered catalytic saturation, rapidly climbing before leveling off at an asymptotic maximum. When studying Competitive Enzyme Inhibition, the plotted curve shifts noticeably to the right. While it ultimately achieves the same maximum velocity asymptote, it requires a dramatically higher substrate concentration to overcome the blocking agent.
Conversely, the non-competitive curve behaves entirely differently, providing a stark visual contrast. This curve flattens out severely, reaching a significantly lower maximum velocity asymptote regardless of how much substrate is added to the system. However, it impressively hits its half-maximum mark at the exact same substrate concentration as the normal, healthy baseline curve.
These diagnostic analyses of saturation plots are fundamental laboratory tools. By understanding how Enzyme Inhibition visually distorts these standard hyperbolic curves, biochemistry students can quickly deduce the physical binding mechanics of unknown pharmacological agents. This rapid visual diagnosis is the first critical step in evaluating new therapeutic drug candidates in the laboratory.
Slide 9: Eadie-Hofstee Linearization in Enzyme Inhibition
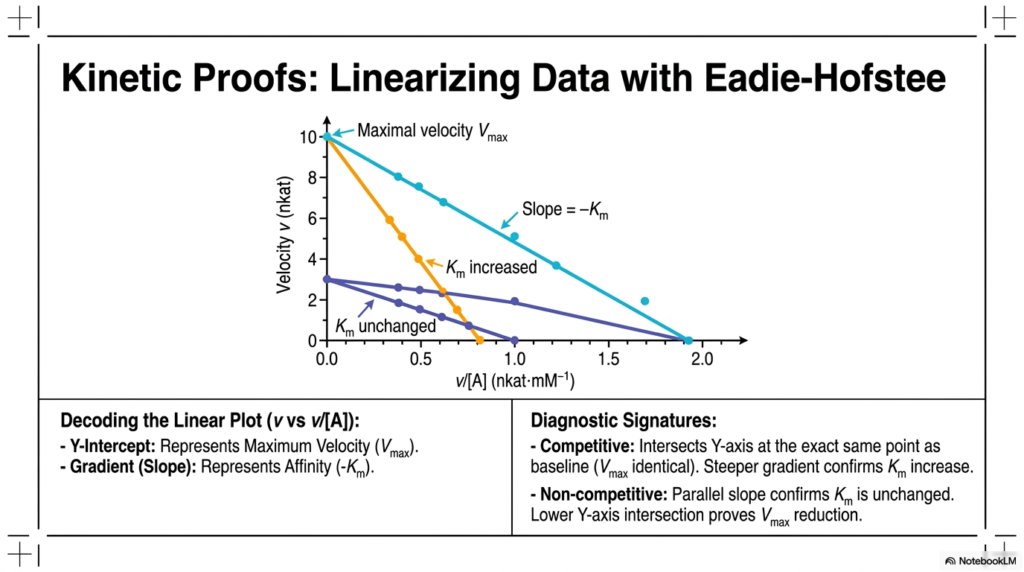
While hyperbolic curves provide a general visual understanding, precise mathematical modeling requires linear data transformations. The Eadie-Hofstee plot elegantly linearizes kinetic data by graphing velocity against the ratio of velocity to substrate concentration. This specific transformation provides distinct diagnostic signatures to identify specific types of Enzyme Inhibition, enabling highly accurate extraction of kinetic constants.
In this linearized format, the key metrics are easily readable straight from the graph’s geometry. The Y-intercept represents the maximum reaction velocity, while the negative slope gradient directly correlates with the binding affinity constant. Decoding these linear plots is a critical skill for students analyzing the precise mathematical impact of Enzyme Inhibition in controlled laboratory settings.
The diagnostic signatures become immediately obvious upon graphing. In a competitive scenario, the linear plot intersects the Y-axis at the same point as the baseline, indicating that the maximum velocity remains identical. However, the significantly steeper gradient visually confirms the pronounced increase in the binding affinity constant, which perfectly matches theoretical predictions.
For a non-competitive scenario, the visual proof is equally striking. The resulting line has a slope perfectly parallel to the baseline, definitively confirming that the binding affinity is unchanged. Simultaneously, the noticeably lower Y-axis intersection unequivocally proves the severe reduction in maximum velocity. This makes Eadie-Hofstee plots an indispensable tool in Enzyme Inhibition research.
Slide 10: The Mathematics Governing Enzyme Inhibition Constants

To truly master the mechanics of molecular regulation, one must dive into the underlying mathematical dissociation constants. This slide rigorously defines the affinity metrics that govern Enzyme Inhibition. The constant Ki represents the specific dissociation metric for a regulator binding to a free protein, while Ki’ denotes the affinity for the actively occupied complex, both measured precisely via laboratory calorimetry.
These distinct dissociation constants are subsequently integrated directly into the modified Michaelis-Menten equation. This expanded mathematical formula represents the foundational calculus of Enzyme Inhibition. By incorporating modifying factors directly into the denominator, the equation accurately predicts how reaction velocity will vary with changing concentrations of regulatory molecules and native substrates.
The modifying factors, alpha and alpha prime, dictate the true kinetic potency of the regulatory molecule. Alpha is calculated by dividing the regulator concentration by its specific dissociation constant. As the concentration of the regulator rises, or if it has an extremely strong binding affinity, these modifying factors increase exponentially, severely dampening the final velocity.
Understanding this mathematical framework is critical for rational drug design. Pharmacologists rely heavily on these precise calculations of Enzyme Inhibition to determine the exact concentration of a drug required to achieve a therapeutic effect without causing systemic toxicity. Mastery of these modifying factors enables the safe and effective dosing of potent biochemical therapeutics.
Slide 11: Reversible Enzyme Inhibition and Transition State Analogs
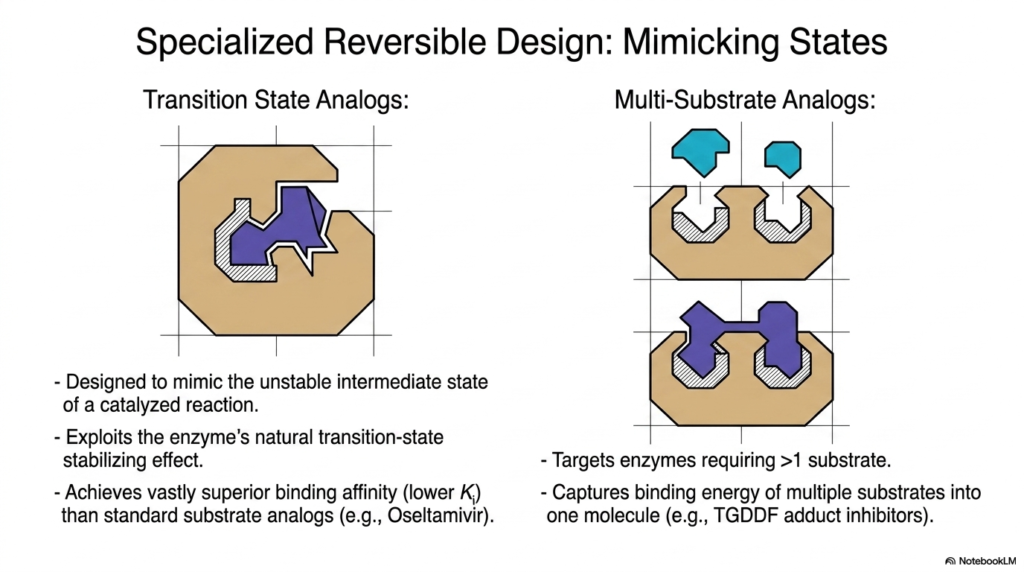
Modern pharmacological design heavily exploits the natural catalytic mechanisms of biological proteins to create highly potent, reversible regulators. This slide highlights Specialized Reversible Design, showcasing how chemists engineer molecules that mimic the unstable intermediate states of catalyzed reactions. By utilizing Enzyme Inhibition principles, scientists can achieve vastly superior binding affinities compared to standard structural analogs.
Transition-state analogs are designed to mimic the incredibly brief, highly unstable structural conformation that occurs mid-reaction. Because biological catalysts have evolved to tightly stabilize this exact intermediate state, they bind to these synthetic analogs with extreme prejudice. This specific form of Enzyme Inhibition results in exceptionally low dissociation constants, yielding highly potent drugs like Oseltamivir.
Another highly sophisticated design strategy involves Multi-Substrate Analogs. These engineered molecules specifically target complex biological reactions that require more than one native substrate to function. By capturing the combined binding energy of multiple substrates and fusing them into a single, cohesive molecule, pharmacologists create incredibly sticky and effective regulatory agents.
These advanced applications of Enzyme Inhibition demonstrate the incredible precision of modern biochemistry. By tricking the biological machinery into binding with molecules that perfectly match their most highly favored structural states, researchers can effectively shut down specific metabolic pathways. This specialized reversible design remains a primary strategy for developing potent antiviral and metabolic medications.
Slide 12: Irreversible Enzyme Inhibition and Covalent Dead-Ends
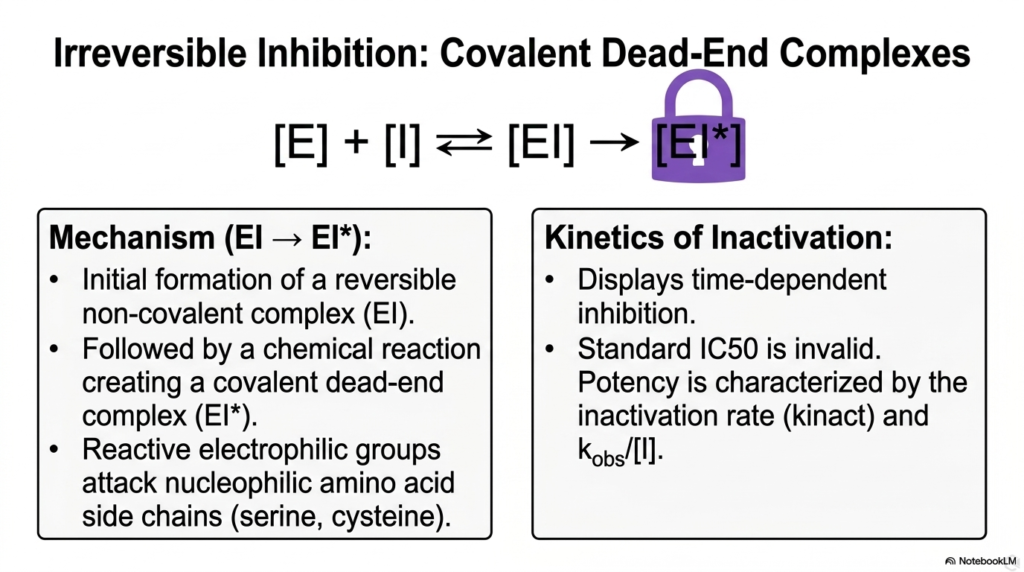
Transitioning from temporary regulatory mechanisms, this slide delves into the severe and permanent consequences of Irreversible Enzyme Inhibition. Unlike reversible counterparts, these dangerous molecules form permanent, covalent dead-end complexes. The mechanism begins deceptively with the initial formation of a standard, reversible non-covalent complex, allowing the regulator to slip neatly into the target active site.
Once nested within the active site, a devastating chemical reaction immediately follows. Reactive electrophilic groups on the regulatory molecule aggressively attack vulnerable nucleophilic amino acid side chains, such as serine or cysteine, lining the protein pocket. This aggressive form of Enzyme Inhibition creates an unbreakable covalent bond, completely and permanently destroying the biological utility of the target complex.
Because this process involves a permanent chemical reaction, the kinetics of inactivation are highly unique. This mechanism displays strict time-dependent inhibition; the longer the target protein is exposed to the regulator, the more complexes become permanently locked. Standard half-maximal inhibitory concentration metrics are entirely invalid for assessing this specific chemical behavior.
Instead, the true potency of this severe Enzyme Inhibition is mathematically characterized by the specific inactivation rate constant. Understanding this time-dependent destruction is vital for toxicologists and medical professionals, as it explains why certain chemical exposures become progressively more lethal over time, requiring immediate physiological intervention before all available cellular machinery is permanently disabled.
Slide 13: Mechanism-Based Enzyme Inhibition and Suicide Substrates
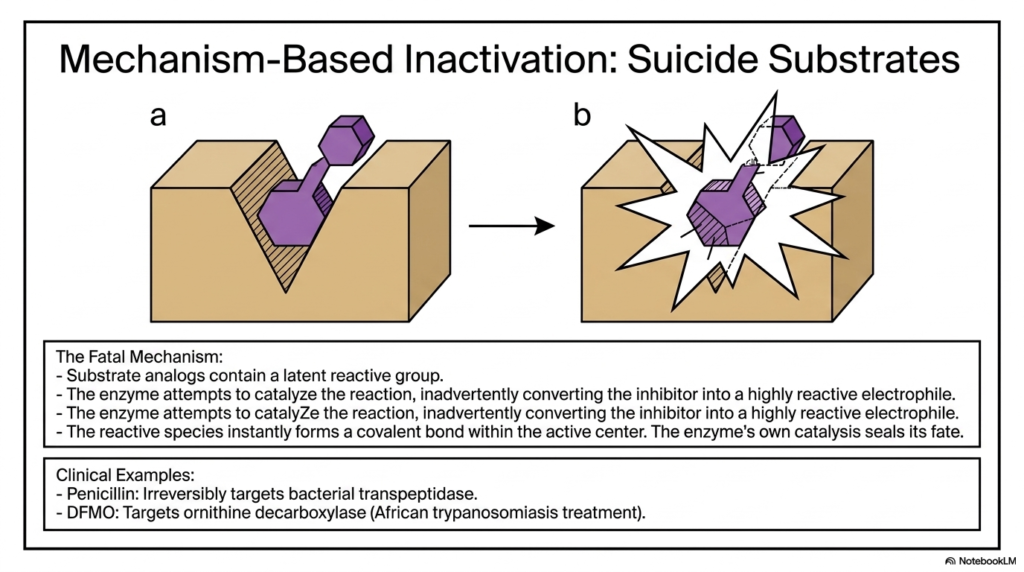
Perhaps the most devious and fascinating regulatory mechanism in biochemistry is Mechanism-Based Inactivation, commonly referred to as the use of suicide substrates. This brilliant form of Enzyme Inhibition utilizes the biological protein’s own catalytic power against itself. These specialized regulators masquerade perfectly as normal, harmless substrates, containing a hidden, latent reactive chemical group.
The fatal mechanism unfolds rapidly once the target protein attempts to catalyze the normal chemical reaction. As the biological machinery acts upon the molecule, it inadvertently converts the disguised regulator into a highly reactive, volatile electrophile. This specific type of Enzyme Inhibition relies entirely on the target protein forcefully triggering its own structural destruction through its natural catalytic cycle.
Once the latent reactive species is generated internally, it instantly forms a permanent covalent bond deep within the active center. The biological catalyst’s own normal functioning seals its irreversible fate. This Trojan-horse approach to Enzyme Inhibition is incredibly specific, as the destructive molecule remains inert until it is forcibly activated by the exact target protein it is designed to destroy.
This elegant, self-destructing mechanism is widely utilized in major clinical therapeutics. Life-saving drugs like Penicillin utilize this exact strategy to permanently target and destroy bacterial transpeptidase complexes. Similarly, drugs like DFMO effectively target critical biological pathways to treat severe parasitic infections, proving that weaponized biological mechanisms save countless human lives daily.
Slide 14: Enzyme Inhibition in Physiological Homeostasis
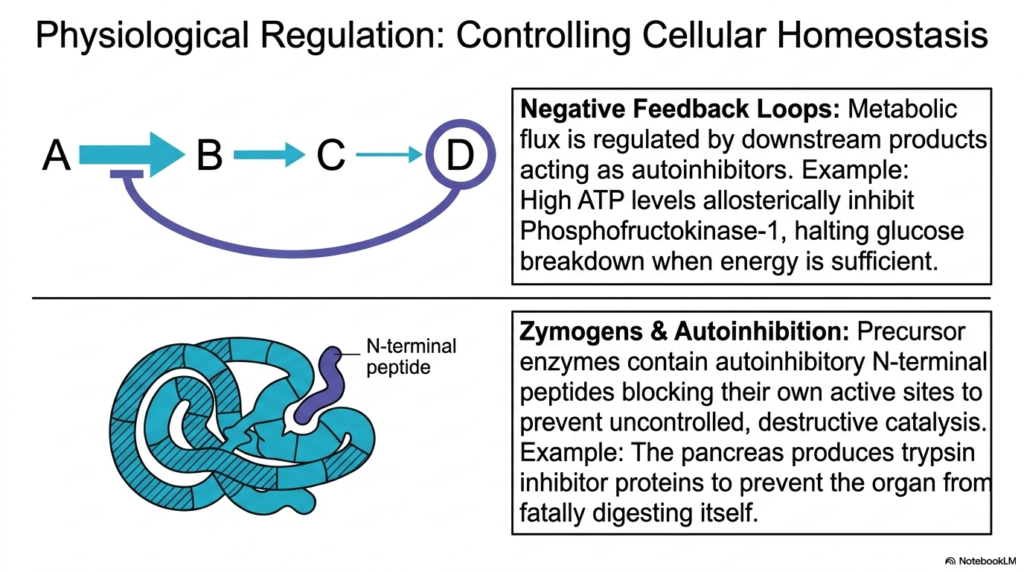
While frequently discussed in the context of pharmacology, natural regulatory mechanisms are constantly occurring to maintain cellular life. This slide highlights how physiological Enzyme Inhibition is paramount for controlling basic biological homeostasis. Without these internal controls, metabolic pathways would wildly consume all available cellular energy, resulting in rapid systemic breakdown and biological death.
A primary example of this internal regulation is the use of negative feedback loops. In these complex systems, metabolic flux is carefully regulated by downstream chemical products acting as natural autoinhibitors. This specific form of Enzyme Inhibition ensures that when sufficient biological energy, such as ATP, accumulates, it naturally binds to and halts the primary upstream catalysts, thereby preserving precious glucose reserves.
Beyond metabolic loops, organisms employ sophisticated structural strategies such as Zymogens to prevent internal destruction. Precursor proteins are synthesized containing built-in autoinhibitory peptide chains that physically block their own active sites. This highly specific Enzyme Inhibition prevents dangerous, uncontrolled catalysis during the delicate manufacturing and transport phases within vital human organs.
A classic medical example of this protective autoinhibition occurs within the human pancreas. The organ produces robust regulatory proteins to temporarily bind and shut down digestive complexes like trypsin. By mastering this localized Enzyme Inhibition, the human body effectively prevents the highly destructive digestive fluids from fatally digesting the very organ that manufactured them.
Slide 15: Clinical Applications of Enzyme Inhibition

The culmination of studying biochemical mechanisms is applying that knowledge to real-world medical and toxicological scenarios. The final slide highlights the global impact of Enzyme Inhibition, noting that nearly one-third of all approved pharmaceutical drugs act as regulatory blockers. These targeted human and pathogen therapeutics form the absolute bedrock of modern clinical medicine.
In the fight against cancer and chronic disease, precision regulation is a primary weapon. Life-saving chemotherapeutics like Methotrexate act through rigorous competitive mechanisms to starve rapidly dividing cancer cells. Similarly, widely used cardiovascular medications employ targeted Enzyme Inhibition to dynamically control blood pressure, showcasing how these microscopic chemical interactions dictate major physiological health outcomes across diverse medical fields.
Conversely, the exact same biochemical principles explain the terrifying lethality of natural toxins and synthetic poisons. Dangerous neurotoxins, ranging from natural plant glycoalkaloids to synthetic weaponized nerve agents like Sarin, weaponize extreme Enzyme Inhibition to trigger catastrophic biological failure. By irreversibly shutting down crucial nervous system regulators, these agents cause rapid, lethal systemic paralysis.
Ultimately, whether utilized as a life-saving antimicrobial drug targeting bacterial cell walls or weaponized as a toxic biological agent, the core mathematical and structural principles remain identical. A deep, comprehensive mastery of Enzyme Inhibition empowers future medical professionals to safely manipulate these powerful biological networks, ensuring they can heal patients while deeply understanding the profound chemical dangers present in nature.
Please read our Content Disclaimer Statement.
Check out our social media channels: