10. Polysaccharides and Glycoproteins: Structure and Function
Have you ever wondered why a tree can stand hundreds of feet tall, or how an Olympic sprinter’s muscles access massive amounts of energy in a fraction of a second? The answer lies in the molecular architecture of carbohydrates. Today, we are going to explore a comprehensive slide deck designed specifically for biochemistry and medical students. The core purpose of this deck is to decode the structural and functional diversity of complex carbohydrates, revealing how simple sugar monomers are linked together to create either indestructible cellular walls or highly accessible energy reservoirs. Let’s break down these complex mechanisms together!
Slide 1: Biochemical Concepts and Structures of Polysaccharides and Glycoproteins
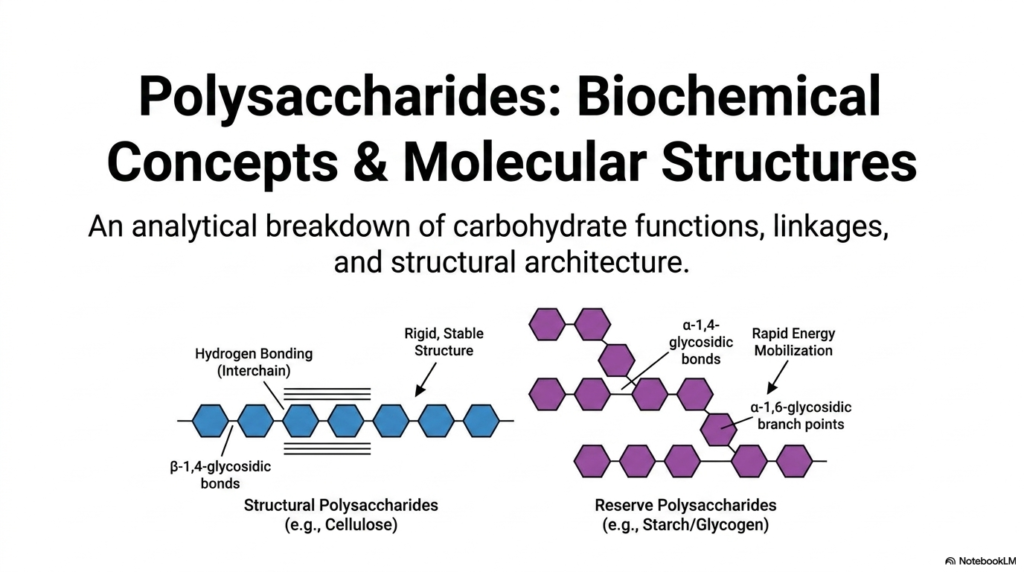
Welcome to our first major topic! When we begin our study of Polysaccharides and Glycoproteins, we immediately see that nature is the ultimate molecular engineer. This first slide provides an essential analytical breakdown of carbohydrate functions, linkages, and structural architecture. As biochemistry students, you must understand that the biological role of Polysaccharides and Glycoproteins is entirely dictated by their three-dimensional conformation. Look closely at the dichotomy presented here. On the left, we have structural polysaccharides like cellulose, which rely on rigid, stable beta-1,4-glycosidic bonds. These linear chains pack tightly together, forming extensive interchain hydrogen bonds that make them incredibly resistant to degradation.
On the right side of the slide, we see reserve polysaccharides like starch and glycogen. Unlike their structural counterparts, these energy-storing molecules utilize alpha-1,4-glycosidic bonds to form main chains, with alpha-1,6-glycosidic branch points scattered throughout. Why does this matter? Branching creates multiple terminal ends, allowing enzymes to rapidly mobilize glucose when a cell desperately needs energy. If you are preparing for medical or advanced biology exams, understanding the distinct behaviors of Polysaccharides and Glycoproteins is absolutely crucial. A simple shift from a beta to an alpha linkage completely transforms the molecule from a rigid support beam into a rapidly accessible battery pack.
Slide 2: Three Biological Imperatives of Polysaccharides and Glycoproteins
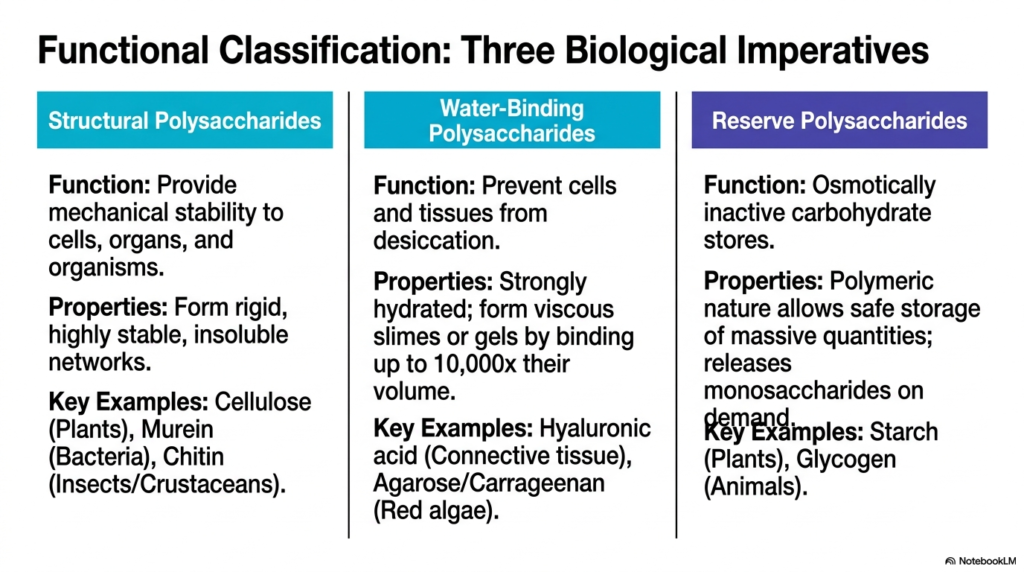
Moving on to slide two, we categorize these macromolecules by their biological functions. The study of Polysaccharides and Glycoproteins reveals three distinct functional domains: structural, water-binding, and reserve. Let’s break these down in plain language. First, structural polysaccharides are designed to provide mechanical stability to cells, organs, and entire organisms. They form highly stable, insoluble networks. We see this in plant cellulose, bacterial murein, and the chitin found in insect exoskeletons.
The second category is uniquely fascinating: water-binding polysaccharides. In the vast landscape of Polysaccharides and Glycoproteins, these are the molecules responsible for preventing cells and tissues from drying out. They are strongly hydrated and can form viscous slimes or gels by binding up to 10,000 times their own volume in water! Hyaluronic acid in your connective tissue is a perfect example. Finally, we have the reserve polysaccharides. Cells need a way to store massive quantities of glucose without disrupting cellular osmosis.
By polymerizing thousands of glucose molecules into giant, osmotically inactive stores like plant starch or animal glycogen, the cell safely hoards energy for a rainy day. Understanding these three functional pillars is the key to mastering the diverse roles of Polysaccharides and Glycoproteins in living systems.
Slide 3: Structural Composition in Polysaccharides and Glycoproteins
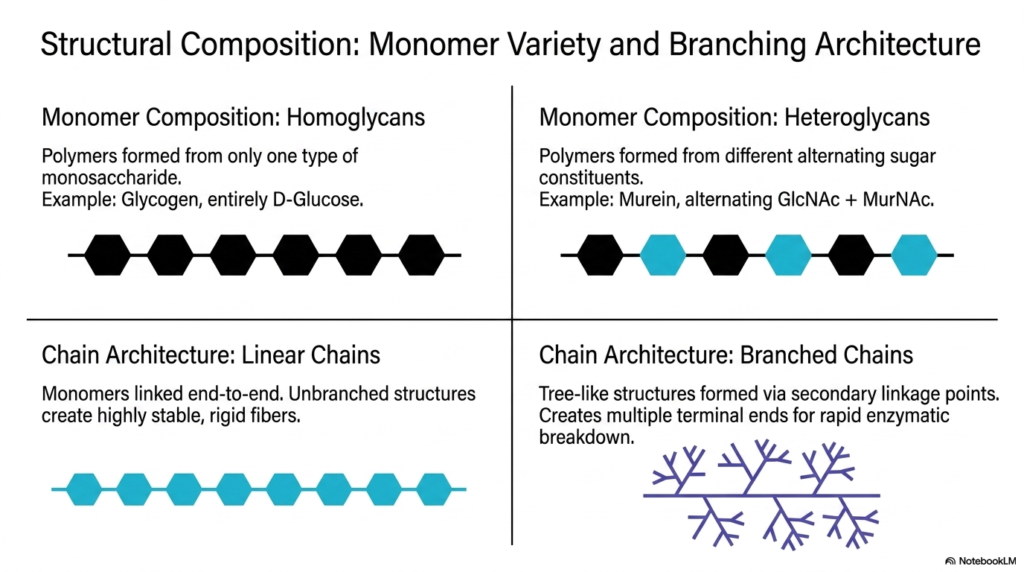
Let’s zoom in on the building blocks! In this slide, we explore the variety of monomers and the branching architecture that define the vast universe of Polysaccharides and Glycoproteins. Think of monomers like Lego bricks. When a polymer is formed from only one type of monosaccharide, we call it a homoglycan. Glycogen is a classic example, as it is composed entirely of repeating D-glucose units. However, many Polysaccharides and Glycoproteins are heteroglycans, meaning they are built from alternating, different sugar constituents. Bacterial murein, which alternates between GlcNAc and MurNAc, is a prime example of this heterogeneous complexity.
Beyond monomer composition, we must look at chain architecture. Linear chains are simply monomers linked end-to-end. Because they lack branches, they can stack tightly together, creating highly stable, rigid fibers perfect for structural support. In contrast, branched chains feature secondary linkage points that create a tree-like structure. From a biochemical perspective, branching is an evolutionary masterpiece. A highly branched molecule has countless “terminal ends.” When a cell needs energy quickly, enzymes can simultaneously attack all these terminal ends, clipping off sugar monomers at incredible speeds. This architectural variety is why Polysaccharides and Glycoproteins can serve such wildly different functions across all kingdoms of life.
Slide 4: Supramolecular Impact of Linkages in Polysaccharides and Glycoproteins
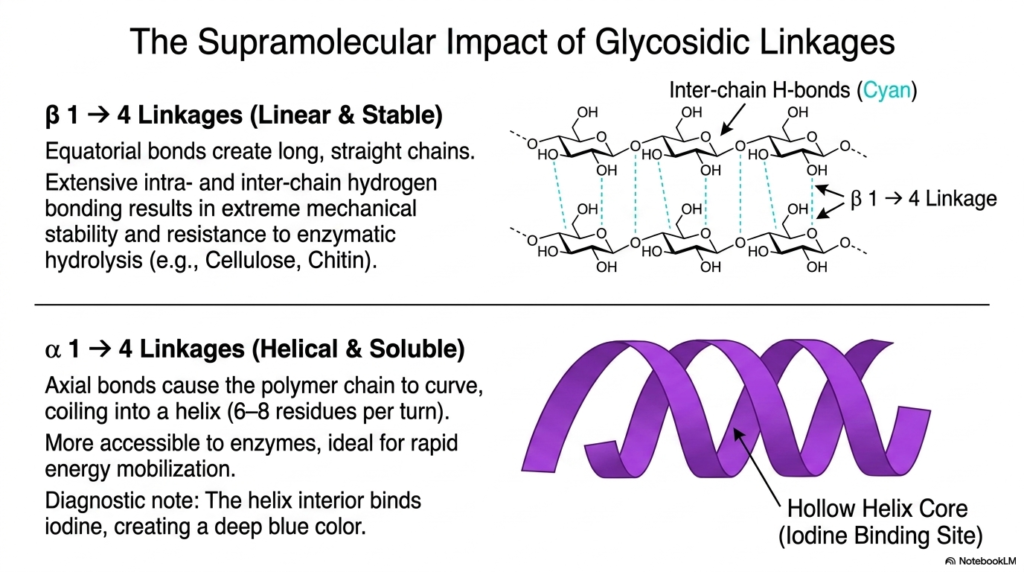
If you want to truly understand biochemistry, you have to appreciate stereochemistry. Slide four illustrates the supramolecular impact of specific glycosidic linkages within the realm of Polysaccharides and Glycoproteins. Let’s compare beta-1,4 linkages with alpha-1,4 linkages. When sugars are joined by beta-1,4 bonds, the equatorial positioning of the bonds forces the polymer to grow in a long, straight line. Because the chains are straight, they can align parallel to one another, allowing for massive networks of intra- and inter-chain hydrogen bonds. This creates a remarkably stable material that is highly resistant to enzymatic hydrolysis, which is exactly why human digestive enzymes cannot break down cellulose!
Conversely, alpha-1,4 linkages feature axial bonds. This slight geometric shift changes everything. Instead of forming a straight line, the polymer chain is forced to curve, eventually coiling into a continuous helix with about 6 to 8 sugar residues per turn. This helical structure is soluble, highly accessible to enzymes, and ideal for energy mobilization. As a fun diagnostic note, the hollow core of this helix is the exact right size to trap iodine molecules, which is why starch turns a deep blue color when exposed to iodine! Mastering these spatial configurations is essential for any student studying Polysaccharides and Glycoproteins, as structure always dictates biological function.
Slide 5: Plant Architecture: Cellulose, Polysaccharides and Glycoproteins
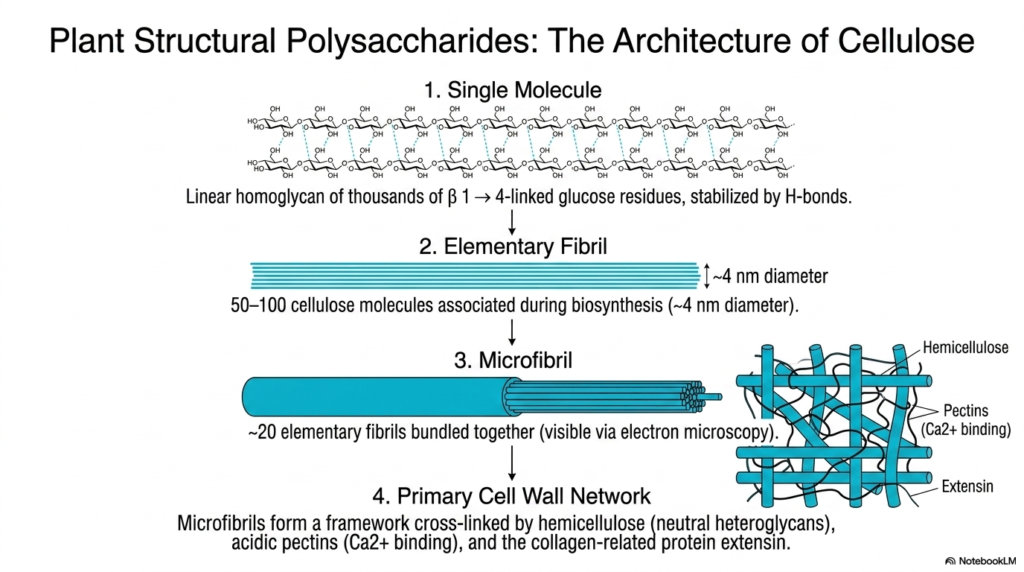
Now let’s apply our structural knowledge to the plant kingdom. This slide walks us through the hierarchical architecture of cellulose, a cornerstone topic in the study of Polysaccharides and Glycoproteins. We start at the microscopic level with a single linear molecule: a homoglycan made of thousands of beta-1,4-linked glucose residues, all stabilized by hydrogen bonds. But nature doesn’t stop there. During biosynthesis, 50 to 100 of these individual cellulose molecules associate with one another to form an “elementary fibril,” which is roughly 4 nanometers in diameter.
The complexity continues to scale up. About 20 of these elementary fibrils are then bundled tightly together to create a microfibril. These microfibrils are so robust that they can actually be visualized using an electron microscope! Finally, these incredibly strong microfibrils are woven together to form the primary cell wall network. They don’t just float randomly; they are extensively cross-linked by hemicellulose (neutral heteroglycans), acidic pectins that bind calcium, and collagen-related proteins called extensins. When you look at the sheer mechanical strength of a giant redwood tree, you are witnessing the phenomenal, scalable power of Polysaccharides and Glycoproteins working in perfect biochemical harmony.
Slide 6: Plant Reserves and Amyloplast Storage – Polysaccharides and Glycoproteins
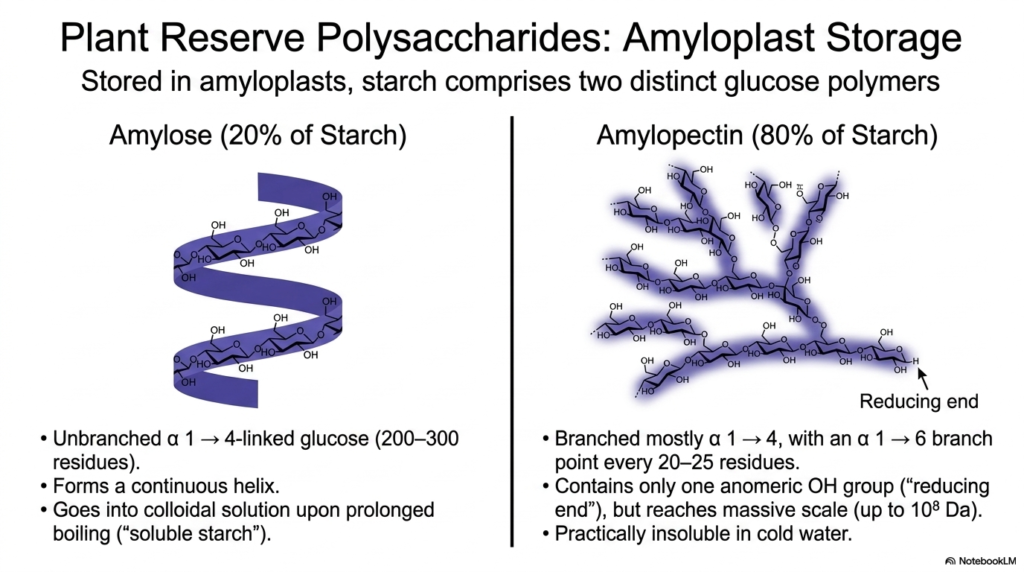
While cellulose provides the physical structure of a plant, the plant also needs a way to store energy. Welcome to the amyloplast! This slide details plant reserve polysaccharides, specifically the breakdown of starch into its two distinct glucose polymers. In our ongoing exploration of Polysaccharides and Glycoproteins, starch serves as the ultimate botanical energy reserve. The first component is amylose, which makes up about 20% of starch. Amylose is an unbranched chain of alpha-1,4-linked glucose that forms a continuous helix. Interestingly, if you boil it long enough, it goes into a colloidal solution known as “soluble starch.”
The remaining 80% of starch is amylopectin. Unlike amylose, amylopectin is heavily branched. It consists mostly of alpha-1,4 linkages but introduces an alpha-1,6 branch point roughly every 20 to 25 residues. This branching allows the molecule to reach a massive scale—up to 10^8 Daltons! Despite its massive size, an entire amylopectin molecule contains only a single anomeric hydroxyl group, known as the “reducing end.” Because of its enormous size and branched complexity, it is practically insoluble in cold water. Understanding the distinct properties of amylose and amylopectin is foundational to studying how plants manage their Polysaccharides and Glycoproteins.
Slide 7: The Animal Dichotomy – Polysaccharides and Glycoproteins
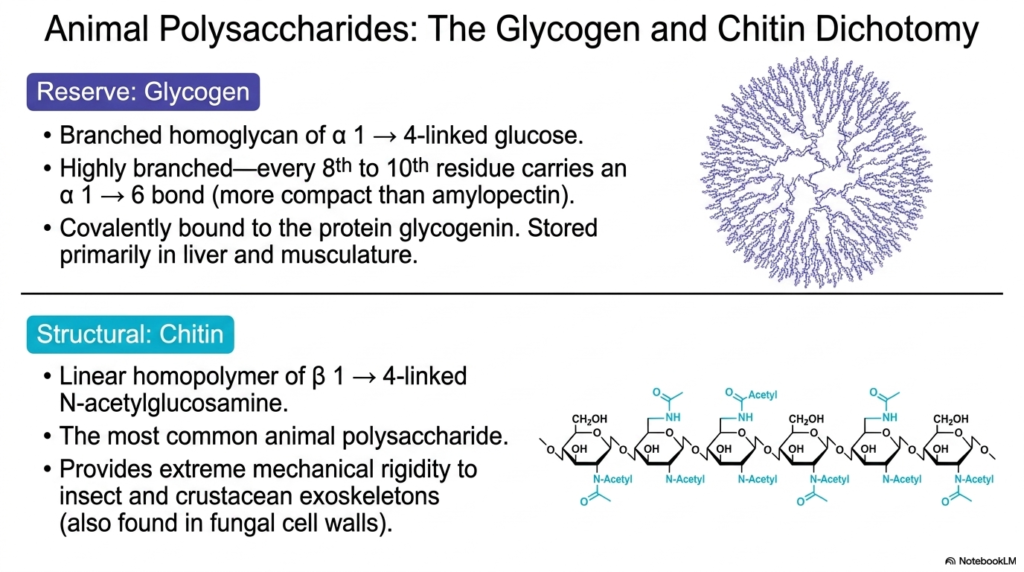
Animals have unique biochemical needs, leading to what we call the glycogen-chitin dichotomy. As we shift our focus to animals in our study of Polysaccharides and Glycoproteins, we see two incredibly specialized macromolecules. First is glycogen, the ultimate animal energy reserve. It is a branched homoglycan of alpha-1,4-linked glucose, very similar to plant amylopectin. However, glycogen is much more highly branched—featuring an alpha-1,6 branch point every 8th to 10th residue! This creates a highly compact, dense sphere of glucose. It is covalently bound to a core protein called glycogenin and is stored primarily in the liver and musculature to provide rapid bursts of energy.
On the structural side, we have chitin. Chitin is a linear homopolymer, but instead of standard glucose, it uses beta-1,4-linked N-acetylglucosamine. This slight chemical modification—the addition of an acetylated amino group—makes chitin incredibly tough. It is the most common animal polysaccharide on Earth, providing extreme mechanical rigidity to the exoskeletons of insects, crustaceans, and, surprisingly, the cell walls of fungi. By comparing glycogen’s fast-burning energy structure with chitin’s armor-like rigidity, we can deeply appreciate the evolutionary versatility of Polysaccharides and Glycoproteins in the animal kingdom.
Slide 8: Bacterial Extracellular Matrices – Polysaccharides and Glycoproteins
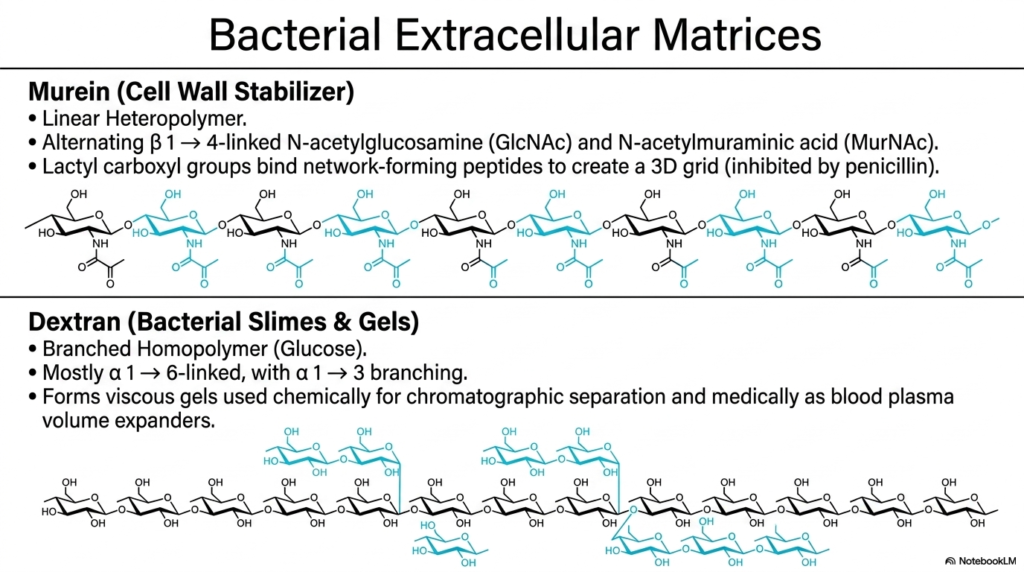
For the medical students reading this, slide eight is particularly high-yield. Here, we examine the bacterial extracellular matrix, a crucial topic in pharmacology and infectious disease. The bacterial world utilizes Polysaccharides and Glycoproteins in highly specialized ways to survive hostile environments. First is murein, also known as peptidoglycan. This is a linear heteropolymer composed of alternating beta-1,4-linked GlcNAc and MurNAc residues. The lactyl carboxyl groups on these sugars bind to network-forming peptides, creating a massive, tough 3D grid that stabilizes the bacterial cell wall. This specific structure is the exact biochemical target of penicillin!
The second major bacterial polysaccharide is dextran, which forms bacterial slimes and gels. Dextran is a branched homopolymer of glucose, but it features a unique linkage pattern: it is primarily alpha-1,6-linked, with alpha-1,3 branching. Bacteria use dextran to form viscous protective biofilms (like the plaque on your teeth!). However, human medicine has brilliantly co-opted this molecule. Because of their gel-forming properties and unique structure among Polysaccharides and Glycoproteins, chemically modified dextrans are used as essential tools in chromatographic laboratory separations and, even intravenously, as blood plasma volume expanders in trauma patients!
Slide 9: Glycosaminoglycans (GAGs) – Polysaccharides and Glycoproteins
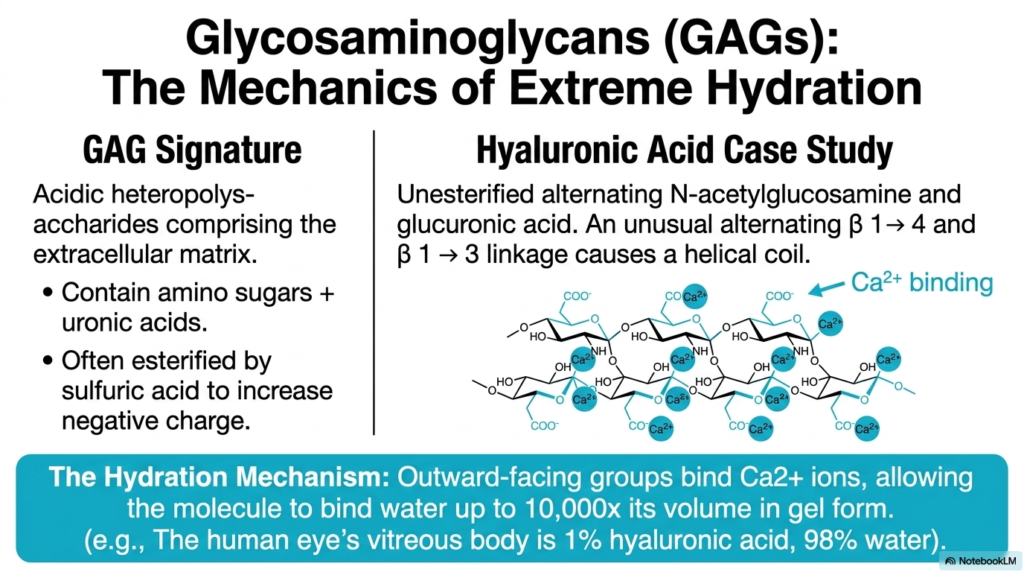
Slide nine introduces us to the mechanics of extreme hydration via Glycosaminoglycans, commonly known as GAGs. When discussing the diversity of Polysaccharides and Glycoproteins, GAGs stand out due to their distinct chemical signature. They are highly acidic heteropolysaccharides that form the structural extracellular matrix in animals. They almost always contain amino sugars paired with uronic acids, and they are frequently esterified by sulfuric acid to massively increase their negative charge.
Let’s look at hyaluronic acid as our prime case study. It is an unesterified polymer of alternating N-acetylglucosamine and glucuronic acid. It utilizes an unusual alternating sequence of beta-1,4 and beta-1,3 linkages, which causes the molecule to coil into a helix. But here is the critical physiological mechanism: the molecule’s outward-facing carboxylate groups bind calcium ions (Ca2+).
The high density of these charged groups creates intense polyanionic repulsion, forcing the molecule to expand outward and trap water. This allows hyaluronic acid to bind up to 10,000 times its own volume in water to form a gel! This class of Polysaccharides and Glycoproteins is the exact reason why the human eye’s vitreous body can maintain its shape while being composed of merely 1% hyaluronic acid and 98% water.
Slide 10: Structural Foundations – Polysaccharides and Glycoproteins
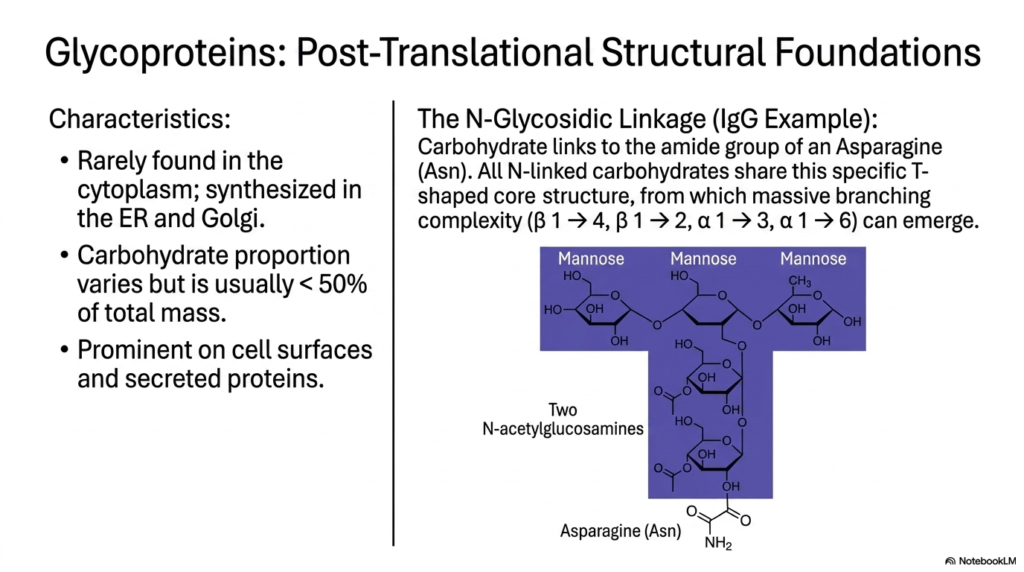
As we move into slide ten, our focus shifts explicitly to glycoproteins and their post-translational structural foundations. While purely carbohydrate polymers are fascinating, the fusion of Polysaccharides and Glycoproteins creates some of the most dynamic molecules in cellular biology. Glycoproteins are rarely found floating freely in the cytoplasm; rather, they are systematically synthesized and folded within the Endoplasmic Reticulum (ER) and the Golgi apparatus. The carbohydrate portion of these molecules varies greatly but usually accounts for less than 50% of the total mass, and they are prominently featured on cell surfaces and secreted proteins like antibodies.
A major focal point for biochemistry students is the N-glycosidic linkage. Using Immunoglobulin G (IgG) as an example, we see that the carbohydrate chain links directly to the amide nitrogen of an Asparagine (Asn) residue. The most remarkable feature of these N-linked Polysaccharides and Glycoproteins is that they all share a universally conserved, T-shaped pentasaccharide core structure. This specific core—comprising mannose and N-acetylglucosamine—serves as the universal foundation from which highly branched patterns (using beta-1,4, beta-1,2, alpha-1,3, and alpha-1,6 linkages) can ultimately emerge to govern cell signaling and immune recognition.
Slide 11: Biosynthetic Pathways of Polysaccharides and Glycoproteins
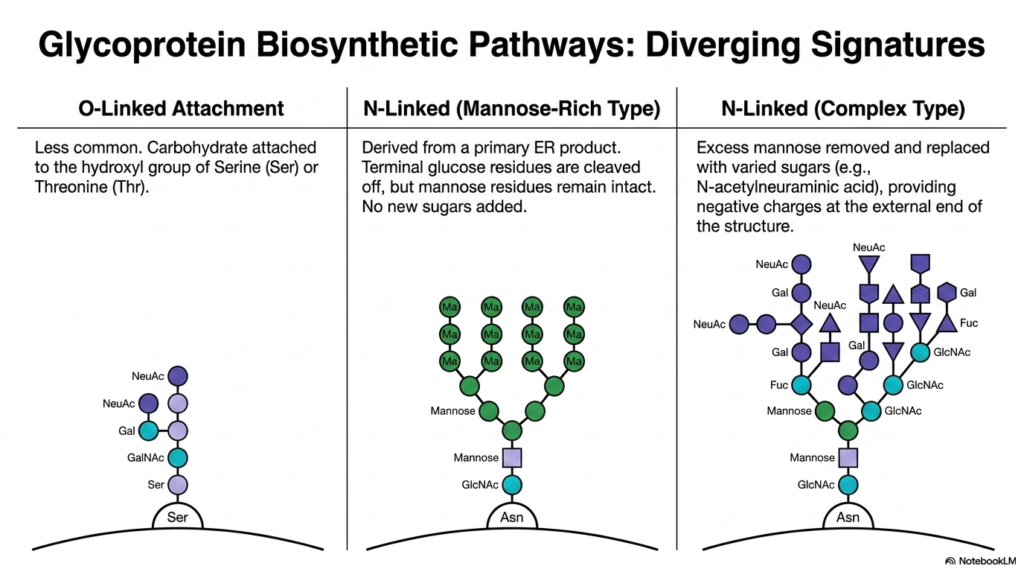
How does a cell actually build these intricate hybrid molecules? Slide eleven maps out the diverging biosynthetic signatures of Polysaccharides and Glycoproteins. We generally classify them by how the sugar chain is attached to the protein. First is the O-linked attachment, which is less common. In this pathway, carbohydrates are attached to the oxygen atom of the hydroxyl group on Serine (Ser) or Threonine (Thr) amino acid residues.
More prominent are the N-linked pathways, which originate from a primary lipid-linked precursor in the ER. There are two major diverging tracks here. The “Mannose-Rich Type” occurs when terminal glucose residues are cleaved off the core structure, but the dense branches of mannose residues remain completely intact, with no new diverse sugars added.
In contrast, the “Complex Type” pathway represents the true structural versatility of Polysaccharides and Glycoproteins. In the Golgi, excess mannose is actively stripped away and replaced with a variety of sugars, such as galactose and N-acetylneuraminic acid (a sialic acid). The addition of sialic acids is particularly critical, as it imparts a strong negative charge at the protein’s external surface, dramatically affecting how the protein interacts with cellular receptors and its survival in the bloodstream.
Slide 12: Synthesis Matrix – Polysaccharides and Glycoproteins
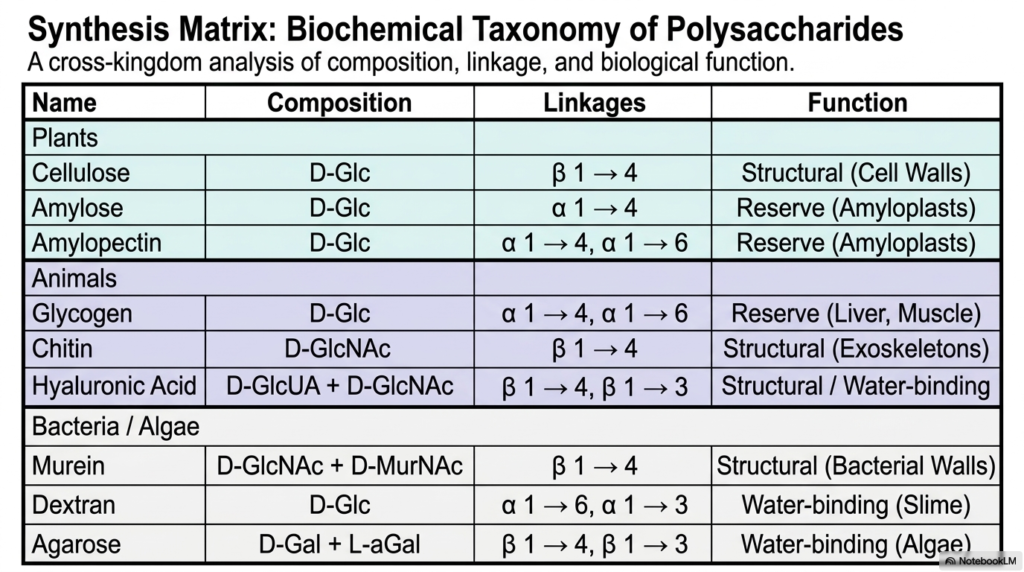
We have covered a tremendous amount of biochemical ground! Our final slide serves as a powerful synthesis matrix, providing a cross-kingdom analysis of composition, linkage, and biological function. If you need a high-yield study guide for Polysaccharides and Glycoproteins, this table is your best friend. It elegantly summarizes the rules we’ve discussed today. Look at the plant kingdom: you see cellulose utilizing beta-1,4 linkages for resilient cell walls, while amylose and amylopectin use alpha-1,4 and 1,6 linkages for amyloplast energy storage.
Shift your eyes to the animal section, and the pattern holds true. Glycogen mimics amylopectin’s alpha-linkage strategy for liver and muscle energy reserves, while chitin mirrors cellulose’s beta-linkages to build tough exoskeletons. Even at the level of bacteria and algae, murein and agarose use beta linkages to form structural walls and water-binding, whereas dextran employs alpha linkages to form slime. The ultimate takeaway from our study of Polysaccharides and Glycoproteins is beautifully clear: across millions of years of evolution, nature has consistently relied on the stereochemistry of glycosidic bonds to determine whether a molecule will serve as an unyielding biological fortress or a rapidly accessible biochemical battery.
Please read our Content Disclaimer Statement.
Check out our social media channels: