22. Ribonucleic Acids: Structure and Function
The blueprint of life may be securely stored in our DNA, but the actual, dynamic construction work is carried out by a far more versatile class of molecules. From precisely transmitting genetic codes to actively catalyzing complex chemical reactions, these dynamic structures are the true unsung heroes of cellular machinery. This comprehensive slide deck explores the foundational structural biochemistry and intricate functional mechanisms of these essential biomolecules. The core purpose of this presentation is to equip college and medical school students with a rigorous, mechanistic understanding of how these diverse molecules fold, function, and regulate microscopic life.
Slide 1: Introduction to Ribonucleic acids: Master Regulators of the Cell
The introductory slide of this comprehensive presentation focuses on the fundamental nature of Ribonucleic acids, which serve as the master regulators of cellular life. The visual immediately captures attention with a highly detailed, three-dimensional rendering of a folded molecular structure. This complex configuration highlights the incredible biochemical diversity that these molecules possess, which is absolutely essential for their role in gene expression. The beautifully rendered cyan sugar-phosphate backbone twists and turns in space, demonstrating that these molecules form intricate, highly functional architectures rather than simple linear strands.
To fully appreciate the biochemical role of Ribonucleic acids, students must understand that they are never just passive carriers of genetic information. Instead, they act as dynamic participants that orchestrate the entire, complex process of gene expression. The vivid purple nitrogenous bases depicted in the rendering pair and interact closely to stabilize these elaborate three-dimensional forms. These specific structural intricacies allow the molecules to interact precisely with various cellular components, ensuring that specialized biochemical tasks are executed with remarkable accuracy.
The overarching theme established by this initial visualization is the sheer functional versatility of Ribonucleic acids within living systems. While DNA is frequently celebrated as the ultimate static blueprint of biological life, it is these highly diverse molecular workers that actually interpret, regulate, and execute those vital instructions. The slide successfully establishes a strong foundational understanding that molecular structure directly dictates cellular function, an essential biochemical concept that will be thoroughly explored throughout the entire presentation.
Furthermore, the explicitly highlighted biochemical diversity hints at the numerous specialized classes and subtypes of Ribonucleic acids that operate within the complex cellular environment. By displaying such an intricate structural model right from the very start, the presentation effectively prepares the audience for a deep dive into the precise physical and chemical properties that make these molecules uniquely suited for their essential biological roles. This robust visual foundation is absolutely critical for medical and university students seeking to master molecular biology.
Slide 2: The Chemical Foundation of Ribonucleic acids Versus DNA
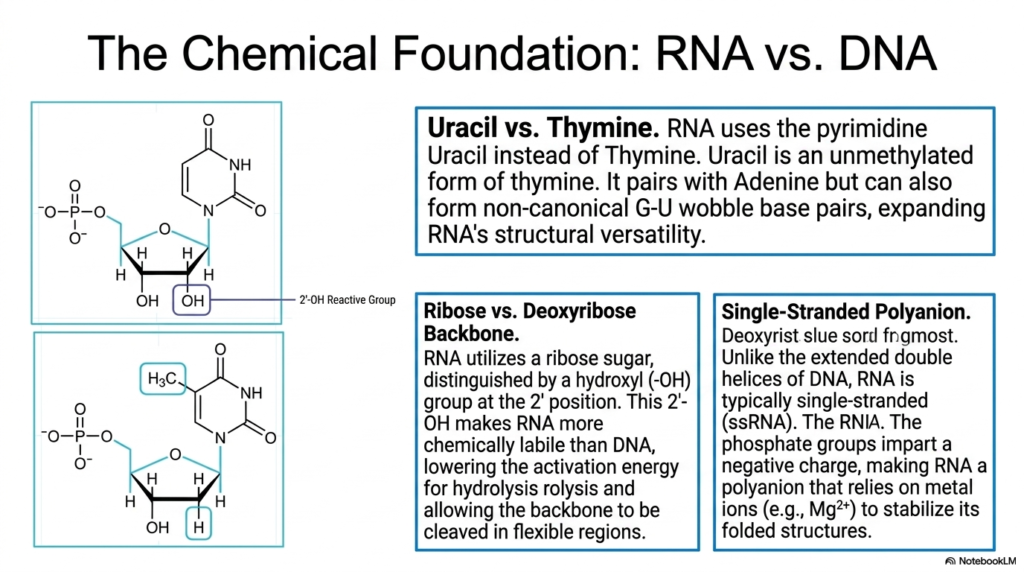
This slide breaks down the foundational chemical differences that distinguish Ribonucleic acids from their more stable genetic counterpart, DNA. The first major distinction highlighted is the use of the pyrimidine Uracil instead of Thymine. The detailed chemical structure reveals that Uracil is an unmethylated form of thymine. While it primarily pairs with Adenine, Uracil can also form non-canonical G-U wobble base pairs, a crucial chemical property that vastly expands the structural versatility of these molecules.
The second crucial chemical distinction involves the pentose sugar backbone of Ribonucleic acids. The visual clearly illustrates the presence of a hydroxyl (-OH) group at the 2′ position of the ribose sugar, which is notably absent in deoxyribose. This highly reactive 2′-OH group makes the molecule significantly more chemically labile than DNA. It effectively lowers the activation energy required for hydrolysis, allowing the backbone to be cleaved in flexible regions during specific biochemical reactions.
A third defining characteristic is the overall physical conformation of Ribonucleic acids as single-stranded polyanions. Unlike the rigid, extended double helices characteristic of DNA, these molecules typically exist as flexible single strands. The prominent phosphate groups detailed in the chemical diagrams impart a strong negative charge across the entire molecule. Because of this potent polyanionic nature, the folding process is highly dependent on environmental factors.
Consequently, the stable folding of Ribonucleic acids relies heavily on the presence of positively charged metal ions, such as magnesium (Mg2+). These metallic cations help neutralize the strong electrostatic repulsion between the negatively charged phosphate groups, allowing the single strand to collapse into its functional three-dimensional shape. Understanding these three core chemical differences—Uracil, the 2′-hydroxyl group, and the polyanionic single-stranded nature—is absolutely essential for students analyzing advanced structural biochemistry.
Slide 3: The Structural Hierarchy of Ribonucleic acids
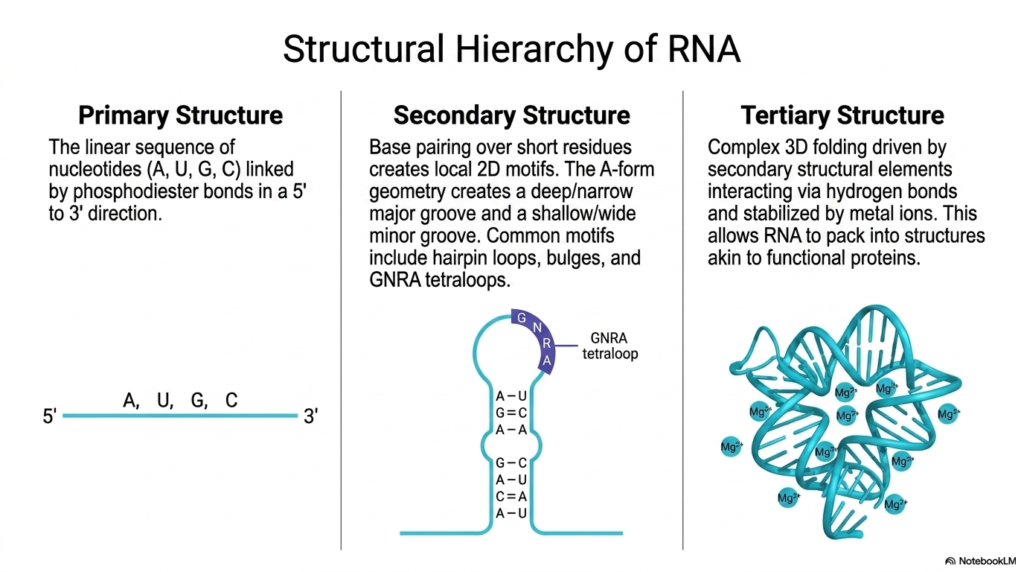
The structural complexity of Ribonucleic acids is beautifully categorized into a distinct hierarchy on this slide, beginning with the primary structure. The leftmost visual demonstrates this fundamental level, which consists of the simple, linear sequence of nucleotides containing Adenine, Uracil, Guanine, and Cytosine. These fundamental building blocks are covalently linked together by strong phosphodiester bonds, strictly following a directional 5′ to 3′ orientation to create the foundational genetic sequence.
Moving to the secondary structure, the presentation illustrates how Ribonucleic acids begin to fold upon themselves. The center diagram shows how localized base pairing over short sequences creates distinct two-dimensional motifs. Common structural elements include hairpin loops, bulges, and specifically highlighted GNRA tetraloops. Furthermore, this localized folding typically adopts an A-form geometry, which is physically characterized by a uniquely deep and narrow major groove coupled with a shallow and wide minor groove.
The pinnacle of this biochemical architecture is the tertiary structure of Ribonucleic acids, showcased on the right side of the slide. This represents the highly complex, three-dimensional folding driven by the various secondary structural elements interacting with one another. These distant regions of the molecule are meticulously drawn together and stabilized via intricate hydrogen bonding networks and the crucial integration of positively charged metal ions like Mg2+.
Ultimately, this advanced hierarchical folding allows Ribonucleic acids to pack tightly into highly defined, stable conformations. The resulting three-dimensional shapes are remarkably akin to the complex architectures seen in functional proteins. By bridging the gap between a simple linear sequence and a sophisticated 3D machine, the molecule gains the extraordinary ability to perform highly specific catalytic and structural roles within the demanding environment of the living cell.
Slide 4: The Core Trinity of Ribonucleic acids: mRNA, tRNA, and rRNA
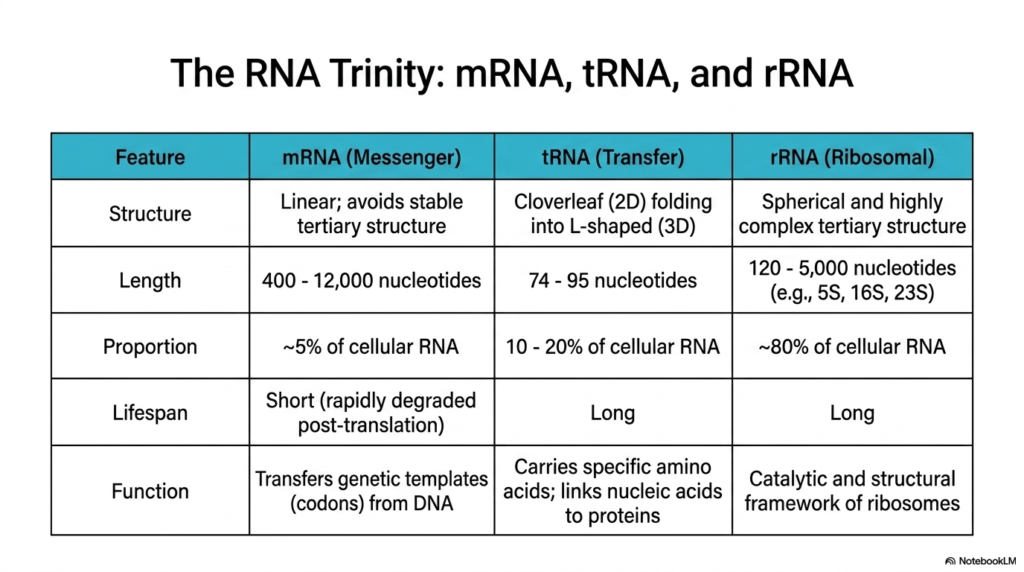
This slide introduces the three fundamental classes of Ribonucleic acids—often referred to as the RNA trinity—through a highly detailed comparative table. The first column examines Messenger RNA (mRNA), which is structurally defined by its linear nature that intentionally avoids stable tertiary folding. Despite accounting for only about 5% of total cellular RNA, these molecules vary wildly in length, ranging from 400 to 12,000 nucleotides, to accurately transfer genetic templates from DNA.
The second class detailed is Transfer RNA (tRNA), representing a highly specialized category of Ribonucleic acids. These remarkably consistent molecules are relatively short, spanning just 74 to 95 nucleotides, and make up roughly 10% to 20% of the cellular pool. Structurally, they are famous for their unique 2D cloverleaf folding that ultimately collapses into a functional 3D L-shape. Their primary role is to serve as reliable adaptors, carrying specific amino acids and linking nucleic acid sequences to growing protein chains.
The third column highlights Ribosomal RNA (rRNA), the most abundant class of Ribonucleic acids, constituting approximately 80% of total cellular RNA. These massive, long-lived molecules span from 120 to over 5,000 nucleotides and fold into spherical, highly complex tertiary structures. Rather than simply carrying information, rRNA provides both the essential structural scaffolding and the primary catalytic framework necessary for the assembly and function of cellular ribosomes.
By directly comparing the structure, length, proportion, lifespan, and function of these molecules, students gain a comprehensive overview of how Ribonucleic acids divide essential cellular labor. The short-lived, rapidly degraded mRNA acts as the fleeting instructional blueprint, while the long-lived tRNA and rRNA form the robust, enduring physical machinery required to continuously synthesize proteins and sustain biological life.
Slide 5: Processing and Maturation of Messenger Ribonucleic acids
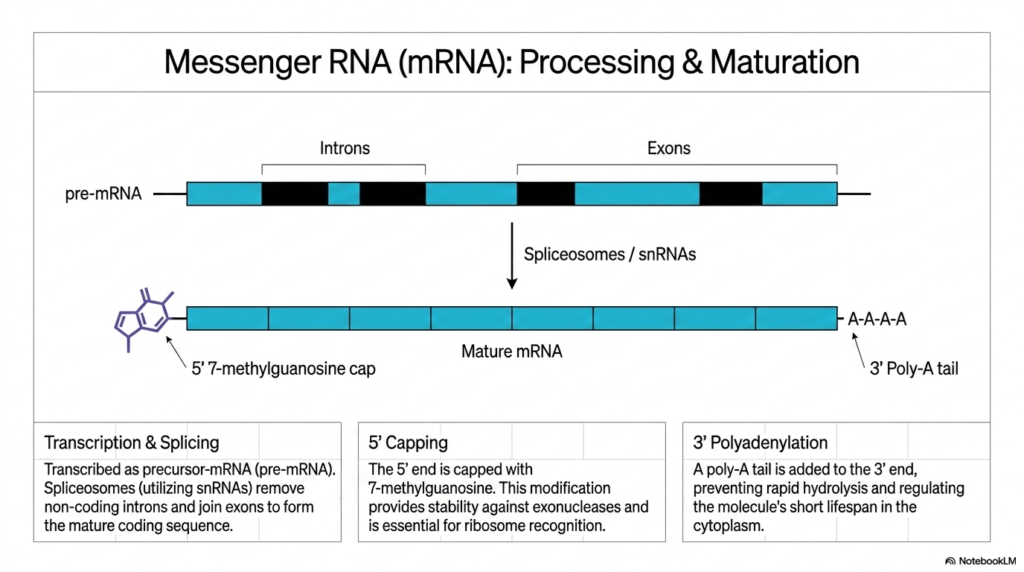
The critical journey from a raw genetic transcript to a highly functional molecule is explored through the processing and maturation of messenger Ribonucleic acids. The central visual diagram tracks this transformation, beginning with a precursor molecule known as pre-mRNA. This initial transcript is composed of alternating coding regions, known as exons, and non-coding intervening sequences, known as introns, which must be surgically addressed before the molecule can be utilized.
To generate a mature sequence, these Ribonucleic acids undergo a highly coordinated process called splicing. A massive molecular machine called the spliceosome, utilizing specialized small nuclear RNAs (snRNAs), precisely targets and removes the non-coding introns. Following this excision, the spliceosome seamlessly joins the remaining exons together. This critical editing process ensures that the final mature mRNA contains an uninterrupted, continuous coding sequence ready for translation.
Simultaneous with splicing, the ends of these messenger Ribonucleic acids are chemically modified to ensure cellular survival. The 5′ end receives a specialized 7-methylguanosine cap. This crucial molecular helmet provides significant physical stability, protecting the delicate transcript against aggressive exonucleases present in the cellular environment. Furthermore, this unique 5′ cap is absolutely essential for the ribosome to successfully recognize and bind to the mRNA.
Finally, the maturation of these Ribonucleic acids is completed at the opposite end through a process known as 3′ polyadenylation. A distinct poly-A tail, consisting of a long chain of adenine nucleotides, is systematically added to the 3′ terminus. This vital modification prevents rapid hydrolysis of the transcript and strictly regulates the molecule’s overall lifespan as it navigates the harsh, enzyme-rich cytoplasm to find a waiting ribosome.
Slide 6: Transfer Ribonucleic acids as the Molecular Adaptors
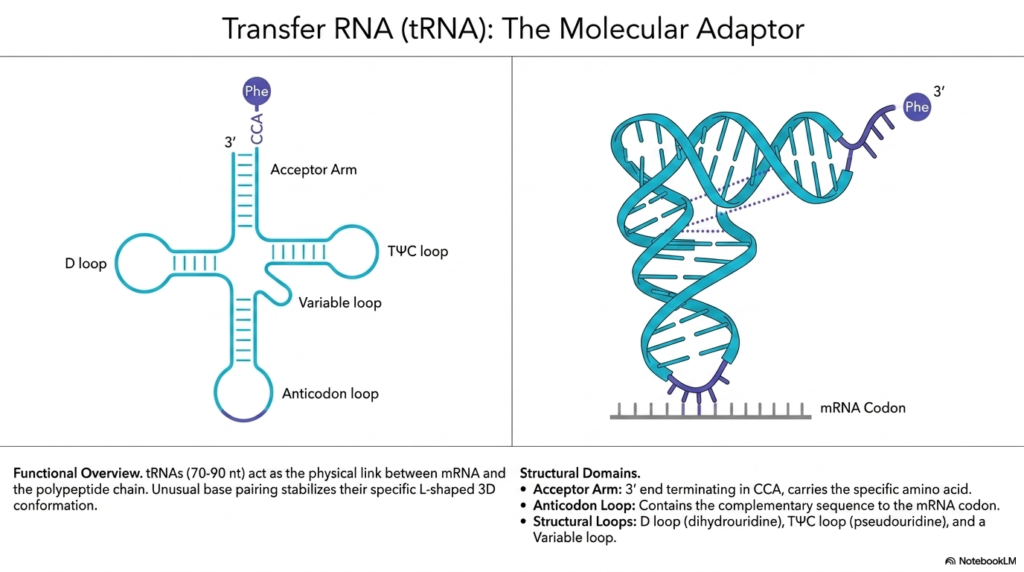
This fascinating slide delves into the physical architecture of transfer Ribonucleic acids, illustrating their vital role as the molecular adaptors of the cell. The visual presents two distinct perspectives: a flattened, two-dimensional cloverleaf model on the left, and a realistic, three-dimensional L-shaped conformation on the right. These highly specialized molecules, typically ranging from 70 to 90 nucleotides in length, act as the direct physical link coordinating the mRNA template with the growing polypeptide chain.
The two-dimensional representation helps decode the distinct structural domains of these transfer Ribonucleic acids. The top features the Acceptor Arm, terminating in a specific CCA sequence that covalently binds and carries the designated amino acid, such as Phenylalanine (Phe). Extending outward are three specialized structural loops: the D loop, the T-psi-C loop containing modified bases, and the highly variable loop that differs subtly between specific tRNA types.
At the very bottom of the two-dimensional model lies the vital Anticodon loop of these Ribonucleic acids. This specific domain contains the crucial complementary nucleotide sequence designed to perfectly match and temporarily bind to the corresponding codon located on the mRNA strand. This exact pairing mechanism is the fundamental chemical basis for accurately translating a genetic nucleic acid code into a physical chain of amino acids.
When folded into its active state, the three-dimensional L-shaped conformation of these Ribonucleic acids is remarkably elegant. The visual demonstrates how unusual base pairing interactions across the molecule stabilize this rigid L-shape. The Acceptor Arm bearing the amino acid points outward at one extreme end, while the Anticodon loop points directly at the mRNA at the other extreme, perfectly positioning the molecule to interface seamlessly within the tight confines of the ribosome.
Slide 7: Deep Dive into Modified Nucleosides of Transfer Ribonucleic acids
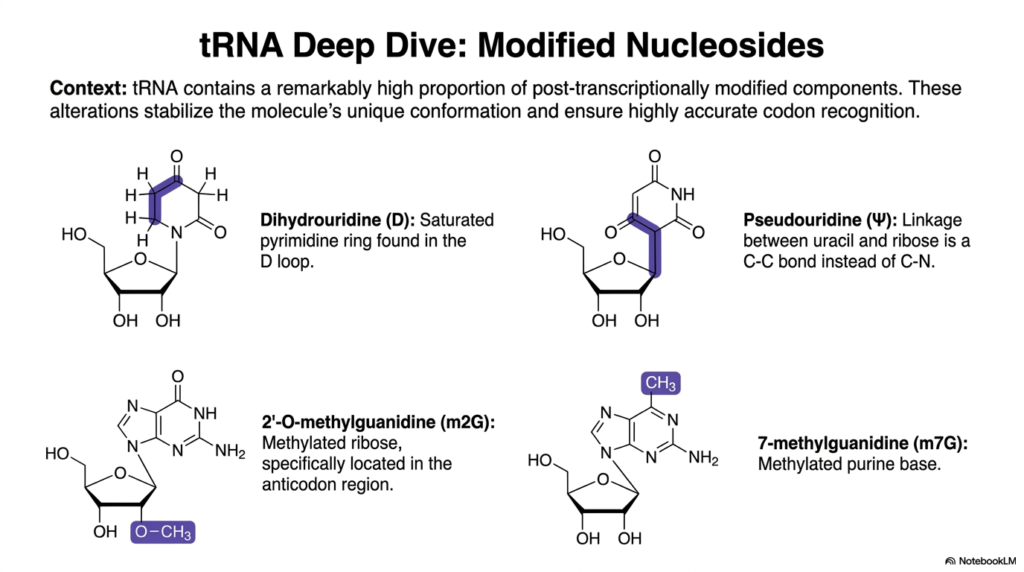
Taking a closer look at molecular fine-tuning, this slide provides a deep dive into the specialized modified nucleosides found uniquely within transfer Ribonucleic acids. The introductory context emphasizes that tRNA contains a remarkably high proportion of post-transcriptionally modified components compared to other transcripts. These specific chemical alterations are not random; they are intentionally engineered by cellular enzymes to stabilize the molecule’s unique 3D conformation and ensure highly accurate codon recognition.
The first two chemical structures presented highlight major modifications involving the pyrimidine ring of these Ribonucleic acids. Dihydrouridine (D) is characterized by a fully saturated pyrimidine ring, a modification that physically bulges and is prominently featured in the molecule’s D loop. Beside it is Pseudouridine (Psi), which features a highly unusual carbon-to-carbon (C-C) bond linking the uracil base directly to the ribose sugar, replacing the standard carbon-to-nitrogen linkage found elsewhere.
The bottom half of the slide focuses on the precise methylation of these Ribonucleic acids. The visual explicitly details 2′-O-methylguanidine (m2G), showcasing a methylated ribose sugar specifically located within the sensitive anticodon region to enhance structural rigidity. Adjacent to it is the structure of 7-methylguanidine (m7G), which represents a heavily methylated purine base that further contributes to the overall stability and functional specificity of the folded transcript.
By analyzing these intricate post-transcriptional modifications, biochemistry students can appreciate the immense complexity of transfer Ribonucleic acids. These chemical tweaks demonstrate that the standard four-letter genetic alphabet is insufficient for the demanding physical stresses placed on tRNA during translation. Instead, the cell actively sculpts and modifies these nucleosides to perfect the molecular machinery required for error-free protein synthesis.
Slide 8: Ribosomal Ribonucleic acids as the Catalytic Core
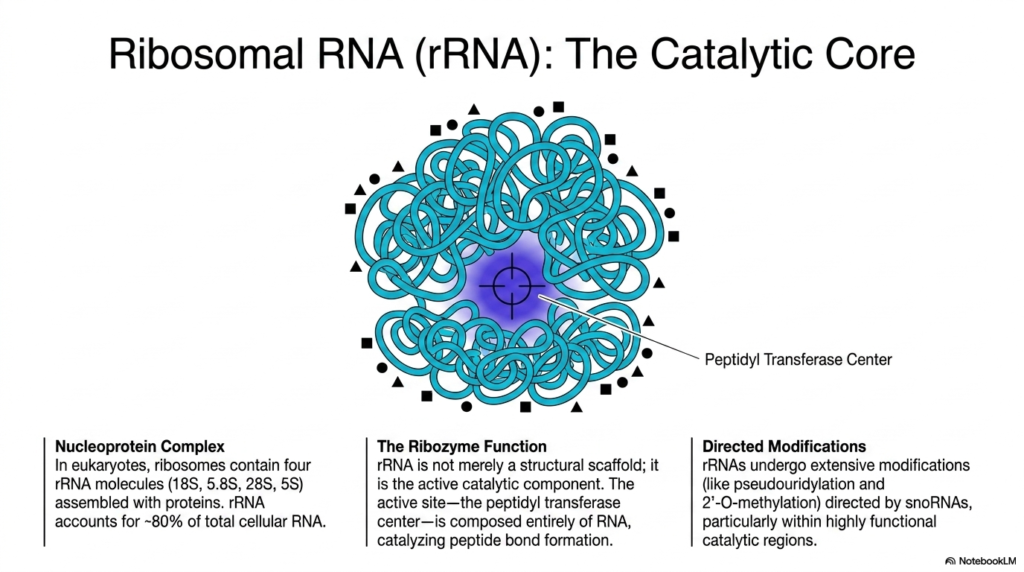
This visually striking slide challenges the traditional view of enzymes by highlighting ribosomal Ribonucleic acids as the true catalytic core of protein synthesis. The central image displays a massive, tangled blue nucleoprotein complex, illustrating how eukaryotic ribosomes are meticulously assembled. This intricate biological machine contains four distinct rRNA molecules—the 18S, 5.8S, 28S, and 5S subunits—which are intricately woven together with stabilizing proteins to account for roughly 80% of total cellular RNA.
The most critical biochemical revelation shown here is the profound ribozyme function of these Ribonucleic acids. The slide explicitly states that rRNA is not merely a passive structural scaffold holding proteins in place. Instead, it is the active catalytic component of the ribosome itself. The prominently highlighted central zone, known as the Peptidyl Transferase Center, is entirely devoid of proteins and composed exclusively of actively folded RNA capable of catalyzing reactions.
It is within this unique, deeply buried RNA pocket that these ribosomal Ribonucleic acids perform their most essential task: catalyzing the formation of peptide bonds between free-floating amino acids. This mechanism proves that RNA itself possesses the powerful chemical ability to drive the fundamental synthesis of all cellular proteins, completely redefining the traditional biochemical hierarchy where only proteins were thought to act as cellular enzymes.
To optimize this immense catalytic power, these Ribonucleic acids undergo extensive, highly directed modifications before full assembly. The text outlines how specialized guide molecules direct site-specific alterations, such as pseudouridylation and 2′-O-methylation, across the massive transcript. These modifications are specifically concentrated within the most highly functional catalytic regions, ensuring the immense molecular machine remains structurally sound and kinetically efficient during rapid protein synthesis.
Slide 9: Regulatory and Processing Classes of Ribonucleic acids
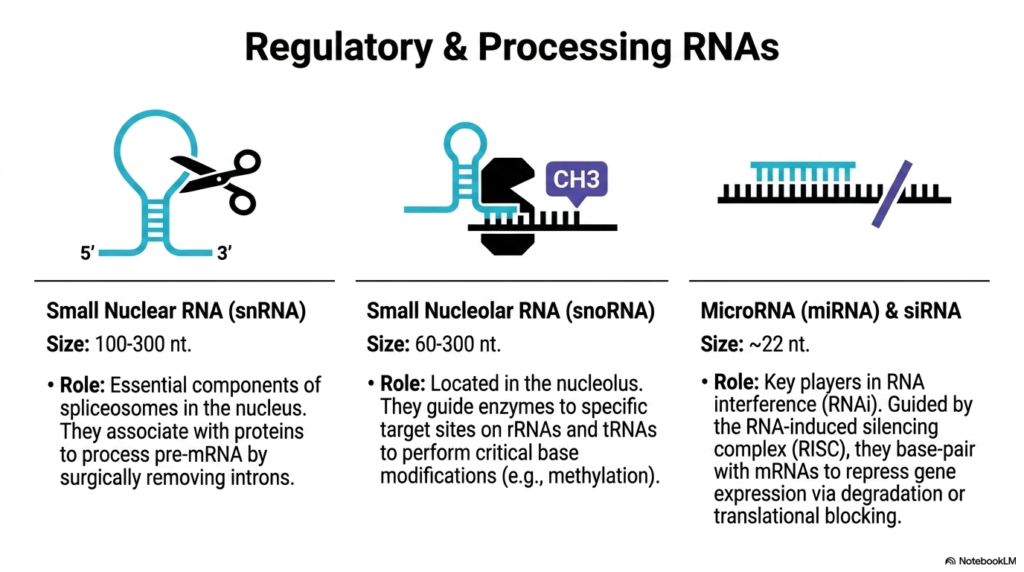
Moving beyond standard protein synthesis, this slide explores the specialized regulatory and processing classes of Ribonucleic acids that actively orchestrate internal cellular operations. The first icon, depicting a pair of molecular scissors, represents Small Nuclear RNA (snRNA). Ranging from 100 to 300 nucleotides in length, these dynamic molecules serve as the essential, active components of spliceosomes located deep within the cellular nucleus. Their primary role is processing pre-mRNA by surgically identifying and removing non-coding introns.
The second crucial class of processing Ribonucleic acids highlighted is Small Nucleolar RNA (snoRNA). Spanning 60 to 300 nucleotides, these specialized molecules are localized specifically within the nucleolus. They function as highly precise molecular guides, binding to specific target sites on maturing rRNAs and tRNAs. By doing so, they accurately direct specialized cellular enzymes to perform the critical chemical base modifications—such as targeted methylation—discussed in earlier slides.
The final section introduces the powerful regulatory mechanisms of microRNA (miRNA) and siRNA, representing incredibly short Ribonucleic acids averaging just 22 nucleotides in length. Despite their minute size, they are the absolute key players in the phenomenon known as RNA interference (RNAi). These tiny transcripts are aggressively utilized by the cellular machinery to hunt down and control specific genetic targets across the cytoplasm.
Operating within the RNA-induced silencing complex (RISC), these short Ribonucleic acids utilize strict complementary base-pairing to bind directly to circulating messenger transcripts. Once attached, they forcefully repress gene expression either by triggering the immediate physical degradation of the target mRNA or by acting as a physical roadblock that completely prevents ribosomal translation. This elegant system provides the cell with profound, immediate control over protein production.
Slide 10: Exploring the Expanded Transcriptome of Ribonucleic acids
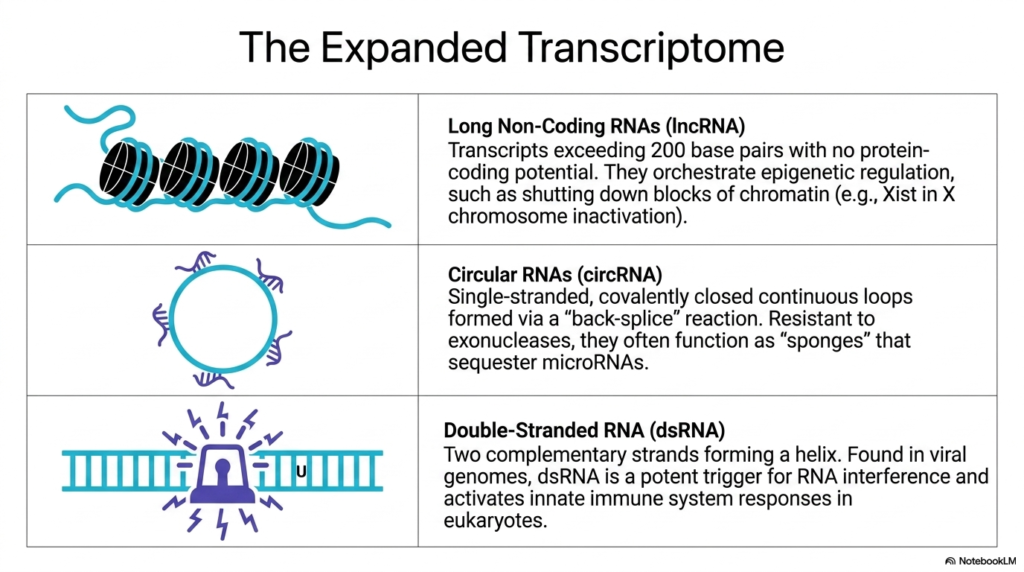
This visually engaging slide invites students to look beyond the classical categories and explore the expanded transcriptome of specialized Ribonucleic acids. The top panel introduces Long Non-Coding RNAs (lncRNA), visually represented by a long cyan strand deliberately wrapping around black cylindrical nucleosomes. These massive transcripts easily exceed 200 base pairs but possess absolutely no protein-coding potential. Instead, they act as master orchestrators of high-level epigenetic regulation within the nucleus.
A prime example of the power of these long Ribonucleic acids is their ability to structurally manipulate genetic expression on a massive scale. The text specifically highlights their role in shutting down entire blocks of chromatin. A famous instance is the Xist transcript, which physically coats and entirely inactivates one of the X chromosomes in female mammals, demonstrating how non-coding sequences can dictate massive, organism-wide genetic silencing.
The middle panel introduces the fascinating architecture of Circular RNAs (circRNA), depicted as a continuous, closed genetic loop. These unique Ribonucleic acids are single-stranded molecules formed via an unusual ‘back-splice’ biochemical reaction that covalently seals their ends together. Because they completely lack exposed terminal ends, they are highly resistant to degradation by cellular exonucleases. They frequently function as molecular ‘sponges’ that attract and securely sequester regulatory microRNAs away from their normal targets.
Finally, the bottom panel warns of the potent biological triggers formed by Double-Stranded RNA (dsRNA). Shown as a rigid double helix emitting alarm signals, these specific Ribonucleic acids consist of two complementary strands locked together. Often found comprising the genomes of invading viruses, their presence in eukaryotic cells acts as a massive red flag. This unique double-stranded structure is a potent trigger that aggressively activates the innate immune system and initiates widespread RNA interference pathways.
Slide 11: Transcription Mechanisms and Synthesis of Ribonucleic acids
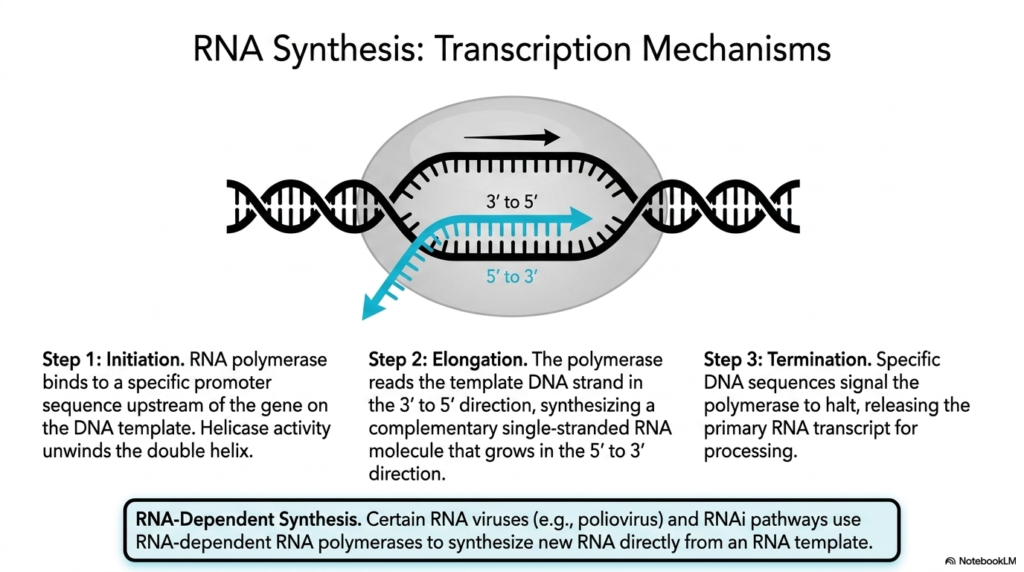
Understanding how genetic codes are mobilized requires analyzing the transcription mechanisms driving the synthesis of Ribonucleic acids. The detailed visual depicts a classic transcription bubble, where the rigid DNA double helix is actively pried apart to accommodate the synthetic machinery. The highly orchestrated process begins with Step 1, Initiation, where a massive RNA polymerase enzyme complex successfully locates and binds tightly to a specific promoter sequence situated upstream of the target gene on the DNA template.
Following successful binding, the synthesis of new Ribonucleic acids moves into Step 2, Elongation. The polymerase complex utilizes its intrinsic helicase activity to unwind the double helix ahead of it. It then reads the exposed template DNA strand strictly in the 3′ to 5′ direction. Matching complementary base pairs, the polymerase seamlessly synthesizes a new, single-stranded RNA molecule that steadily grows longer in the 5′ to 3′ direction.
The fundamental creation of these Ribonucleic acids concludes with Step 3, Termination. The rapidly moving polymerase eventually encounters highly specific DNA sequences that serve as distinct biological stop signs. These specific signals force the polymerase machinery to physically halt its synthetic activity, detach from the DNA template, and finally release the freshly minted primary RNA transcript into the cellular environment, ready for subsequent processing and maturation.
The slide concludes by noting a fascinating alternative pathway known as RNA-Dependent Synthesis of Ribonucleic acids. While standard cellular transcription always relies on a DNA template, certain biological entities bypass this entirely. The text highlights that specific infectious agents, such as the poliovirus, as well as internal cellular RNAi pathways, utilize specialized RNA-dependent RNA polymerases to synthesize brand new transcripts directly from a pre-existing RNA template, showcasing incredible biochemical adaptability.
Slide 12: The RNA World Hypothesis and Evolution of Ribonucleic acids
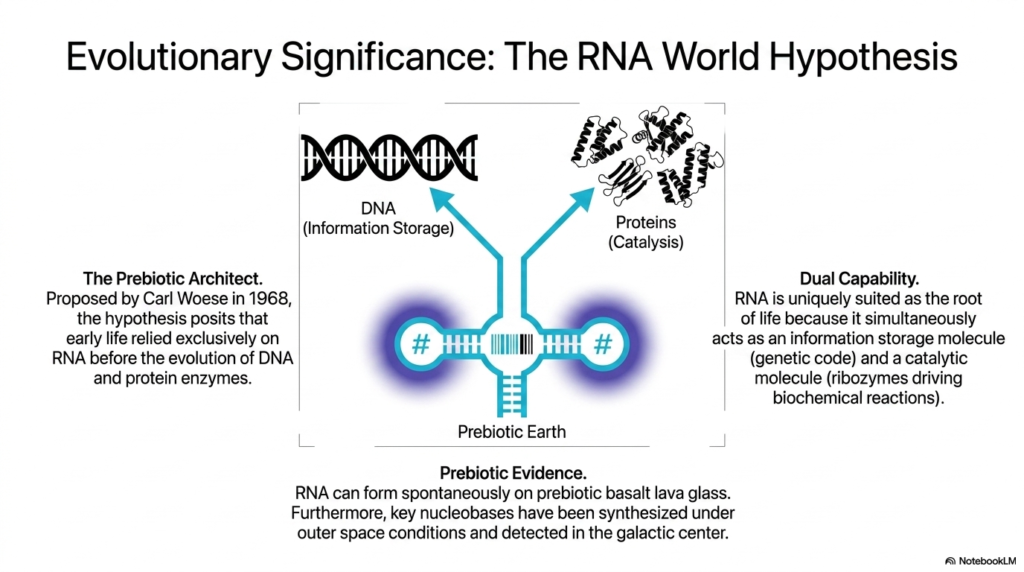
This theoretical slide takes a profound look backward in time to explore the evolutionary significance of Ribonucleic acids through the lens of the famous RNA World Hypothesis. The central diagram visually bridges a critical evolutionary gap, placing a functional RNA molecule exactly midway between DNA, which represents long-term information storage, and complex proteins, which represent active catalysis. This striking visual captures the core premise of how early biological systems may have operated.
The concept, famously proposed by Carl Woese in 1968, positions these ancient Ribonucleic acids as the ultimate prebiotic architects of life. The hypothesis posits a radical idea: before the evolutionary emergence of the stable DNA double helix and before the complex folding of protein enzymes, the earliest forms of life relied entirely and exclusively on RNA molecules to maintain, replicate, and catalyze primitive biological systems on a harsh, ancient Earth.
The fundamental reason these early Ribonucleic acids are perfectly suited as the root of biological life is their astonishing dual capability. As explicitly outlined on the right side of the slide, these unique molecules simultaneously possess the ability to act as reliable information storage molecules, holding the nascent genetic code, while concurrently folding into active ribozymes capable of driving the very biochemical reactions necessary for survival and reproduction.
Supporting this paradigm-shifting theory, the bottom panel presents compelling prebiotic evidence regarding the origins of Ribonucleic acids. Laboratory experiments have remarkably demonstrated that these molecules can form spontaneously on the types of prebiotic basalt lava glass that covered the early Earth. Furthermore, key essential nucleobases required for their synthesis have been successfully created under simulated outer space conditions and even definitively detected floating within the galactic center, hinting at a universal chemical blueprint.
Slide 13: Clinical Horizons and Therapeutics Using Ribonucleic acids
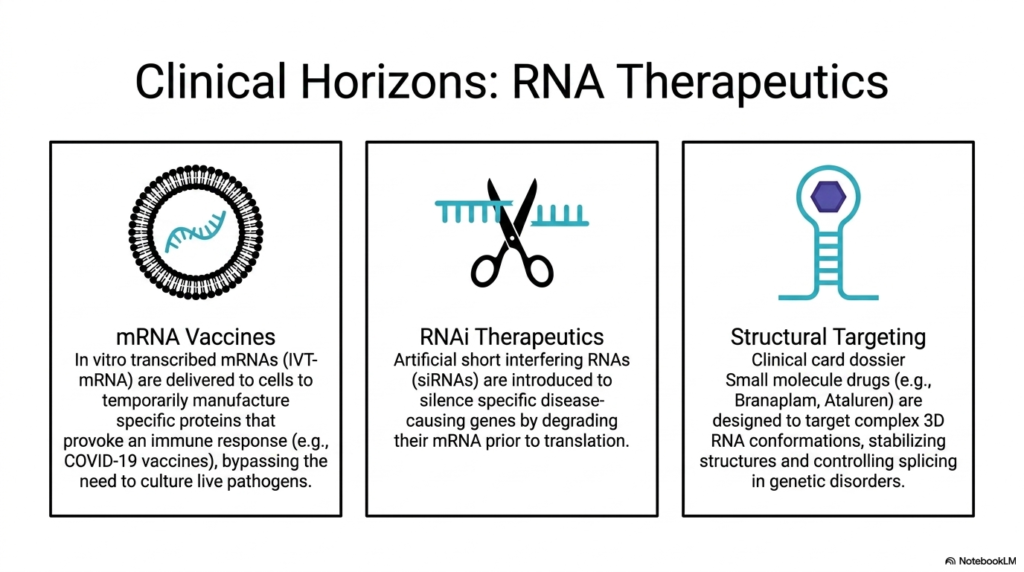
The final slide brilliantly transitions from fundamental structural biochemistry to the cutting-edge clinical horizons of modern therapeutics utilizing Ribonucleic acids. The presentation highlights three major pharmaceutical applications that are actively revolutionizing medical treatments. The first panel focuses on mRNA Vaccines, visually represented by an engineered lipid nanoparticle surrounding a fragile transcript. These sophisticated treatments utilize in vitro transcribed mRNAs (IVT-mRNA) safely delivered directly into a patient’s living cells.
Once successfully delivered, these therapeutic Ribonucleic acids instruct the patient’s own cellular machinery to temporarily manufacture specific, harmless viral proteins. This targeted protein production successfully provokes a robust, highly specific immune response—such as seen in the breakthrough COVID-19 vaccines—completely bypassing the traditional, dangerous need to culture and inject live or attenuated pathogens into the human body.
The second panel explores the immense potential of RNAi Therapeutics, symbolized by a pair of precision molecular scissors targeting specific Ribonucleic acids. This clinical approach introduces highly engineered artificial short interfering RNAs (siRNAs) into the patient’s system. These synthetic molecules are expertly designed to aggressively silence specific, known disease-causing genes by locating and completely degrading their corresponding messenger mRNA transcripts prior to any harmful protein translation occurring.
Finally, the third panel details the innovative field of Structural Targeting of Ribonucleic acids. Rather than introducing new nucleic acids, this approach utilizes specialized small molecule drugs, with clinical examples explicitly including Branaplam and Ataluren. These revolutionary drugs are meticulously designed to locate and target the complex, three-dimensional RNA conformations discussed earlier in the deck. By physically binding to and stabilizing these unique structures, the drugs successfully control and correct erratic gene splicing in severe genetic disorders.
Please read our Content Disclaimer Statement.
Check out our social media channels: