44. Diabetes Mellitus: Insulin Biosynthesis and Metabolic Deficiency
How does a single, tiny protein molecule manage the energy balance of an entire human organism? The answer lies in its precise, three-dimensional architecture. The core purpose of this introductory slide is to establish the disease’s foundational biochemistry by visualizing insulin’s complex folded structure. Understanding this molecular blueprint is the first step in decoding the systemic failures that occur when this critical hormone is missing or ignored by the body.
Slide 1: The Molecular Architecture of Diabetes Mellitus: Why Protein Structure Matters

At the very heart of Diabetes Mellitus is a profound disruption in cellular communication. The molecular structure displayed represents insulin in its hexameric form. This is a tightly packed, highly stable arrangement that allows the human pancreas to store massive quantities of the hormone safely. Six insulin monomers bind together around central zinc ions. However, for the body to actually utilize this hormone, these stable hexamers must rapidly dissociate into active monomers upon release into the bloodstream.
When the body requires energy management, the active insulin monomers bind to specialized receptors on the surface of target cells. Without this functional protein interaction, the biochemical gates that permit glucose to enter cells remain tightly locked. A diagnosis of Diabetes Mellitus essentially means this elegant molecular system has collapsed. The failure happens either because the vital protein is no longer synthesized or because the target cells have grown entirely resistant to its biochemical signal.
For future healthcare professionals, grasping the molecular foundations of Diabetes Mellitus is absolutely essential. It directly bridges the gap between abstract molecular biology and the severe clinical symptoms patients experience. By thoroughly examining this structural biochemistry, the medical community gains the deep mechanistic insight needed to engineer synthetic insulins, develop targeted pharmacological therapies, and create better diagnostic tools for managing this pervasive disease.
Slide 2: The Endocrine Paradigm of Diabetes Mellitus: Orchestrating Systemic Metabolism
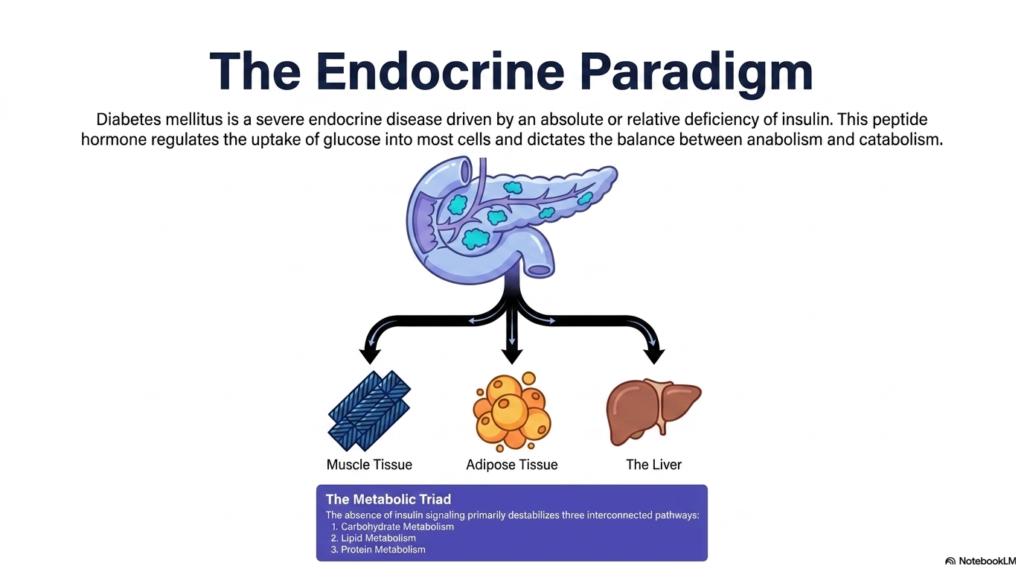
Imagine a master conductor suddenly abandoning an orchestra mid-symphony; biological chaos would instantly follow. This slide illustrates that exact scenario, with the core purpose of explaining how the absence of insulin disrupts the entire endocrine paradigm. It highlights the triad of primary tissues that rely on this peptide hormone, showing how the absence of a single chemical messenger can trigger a multi-organ crisis.
Diabetes Mellitus is fundamentally defined as a severe endocrine disease driven by an absolute or relative deficiency of insulin. This crucial peptide hormone acts as the master regulator of systemic energy, dictating the delicate balance between anabolism and catabolism. When the pancreas fails to secrete adequate insulin, or when tissues stop responding to it, the body loses its primary mechanism for driving glucose into cells. Consequently, energy cannot be utilized or stored properly.
The illustration highlights the three major metabolic players impacted by Diabetes Mellitus: muscle tissue, adipose tissue, and the liver. Under normal physiological conditions, insulin signals muscle and fat cells to take up glucose from the blood, while simultaneously telling the liver to halt new glucose production. Without this signal, these three tissues undergo a radical biochemical shift. They essentially behave as if the body is starving, completely ignoring the abundant glucose circulating in the bloodstream.
Understanding this tissue triad is critical for comprehending the full scope of Diabetes Mellitus. The absence of insulin signaling does not merely affect blood sugar; it profoundly destabilizes three interconnected pathways. Carbohydrate metabolism spirals out of control, lipid metabolism breaks down into unregulated fat breakdown, and protein metabolism shifts toward muscle degradation. This systemic cascade reveals why this condition is far more than just a simple blood sugar problem.
Slide 3: Cellular Biosynthesis in Diabetes Mellitus: How Beta Cells Build Insulin
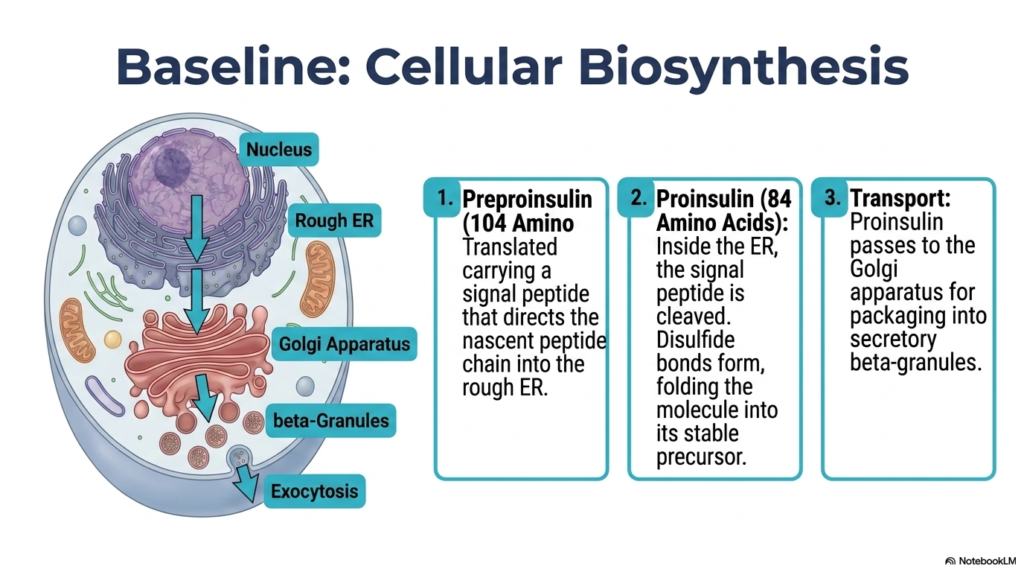
Before a hormone can exert its regulatory power over the body, it must be meticulously constructed from scratch on a cellular assembly line. The core purpose of this slide is to track the fascinating biochemical biosynthesis of insulin within the pancreatic beta cell. By mapping this baseline cellular process, students can better understand where the production pipeline fails in the disease state.
The assembly of insulin begins in the nucleus, but the actual physical construction takes place at the rough endoplasmic reticulum. The cellular machinery translates the genetic code into preproinsulin, a nascent 104-amino-acid precursor molecule. This molecule carries a specific signal peptide that acts as a molecular address tag, directing the growing peptide chain directly into the interior lumen of the endoplasmic reticulum. In cases of advanced Diabetes Mellitus, the cellular stress on this production machinery can lead to significant beta-cell dysfunction.
Once inside the endoplasmic reticulum, the signal peptide is rapidly cleaved away, resulting in a molecule known as proinsulin. This 84-amino-acid structure then undergoes critical folding driven by the formation of disulfide bonds. These covalent linkages are absolutely vital for maintaining the structural integrity of the final hormone. If genetic mutations disrupt this specific folding process, the resulting structural defects can directly trigger forms of Diabetes Mellitus by preventing the creation of functional insulin.
Following successful folding, proinsulin is encapsulated and transported to the Golgi apparatus. Here, the cellular machinery packs the precursor molecules into secretory beta-granules. This compartmentalization is the final preparatory step before the hormone is chemically refined. Understanding this intricate transport and packaging system is essential for biochemists studying Diabetes Mellitus, as any bottleneck or defect in this pathway severely compromises the body’s ability to maintain energy homeostasis.
Slide 4: Peptide Maturation in Diabetes Mellitus: From Proinsulin to Active Hormone
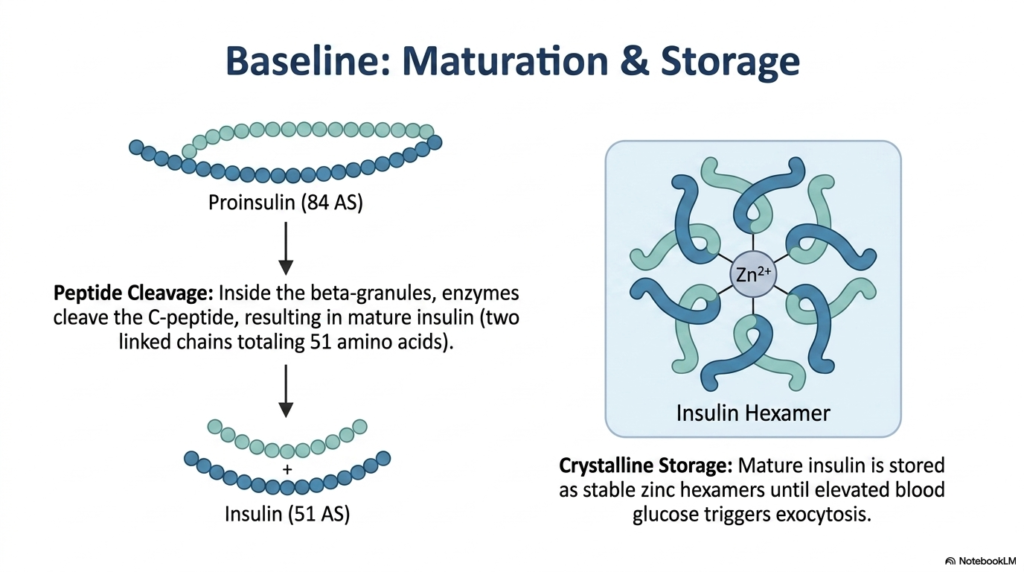
A newly manufactured tool is often useless until it is properly sharpened and securely stored for future use. The core purpose of this slide is to illustrate the final maturation and storage phases of insulin prior to its cellular release. It breaks down the enzymatic cleavage that activates the hormone, providing a crucial window into beta cell function.
Inside the secretory beta-granules, specialized endopeptidase enzymes go to work on the folded proinsulin molecule. These enzymes perform a highly specific biochemical surgery, cleaving away the connecting C-peptide sequence. This excision leaves behind the mature insulin molecule, which consists of two distinct polypeptide chains—the A chain and the B chain—linked together by the previously formed disulfide bridges, totaling 51 amino acids. In clinical evaluations of Diabetes Mellitus, measuring this byproduct is highly valuable.
The excised C-peptide is co-secreted into the bloodstream in a one-to-one ratio with mature insulin. Because C-peptide degrades much more slowly than insulin, clinicians frequently measure it to accurately assess a patient’s residual endogenous pancreatic function. For patients with Diabetes Mellitus, a low or absent C-peptide level strongly indicates total beta-cell failure, whereas a normal or high level suggests that insulin resistance is the primary driver of the metabolic dysfunction.
Once fully matured, insulin does not immediately enter the bloodstream. Instead, it is stored in a highly stable, crystalline form within the beta-granules. The molecules coordinate around zinc ions to form tightly packed hexamers. These hexamers remain safely stored until a glucose influx signals the cell to undergo exocytosis. When researching Diabetes Mellitus, understanding this zinc-dependent storage mechanism is critical for pharmacologists designing synthetic, long-acting insulin formulations for patient therapy.
Slide 5: Divergent Pathogenesis in Diabetes Mellitus: Type 1 Versus Type 2
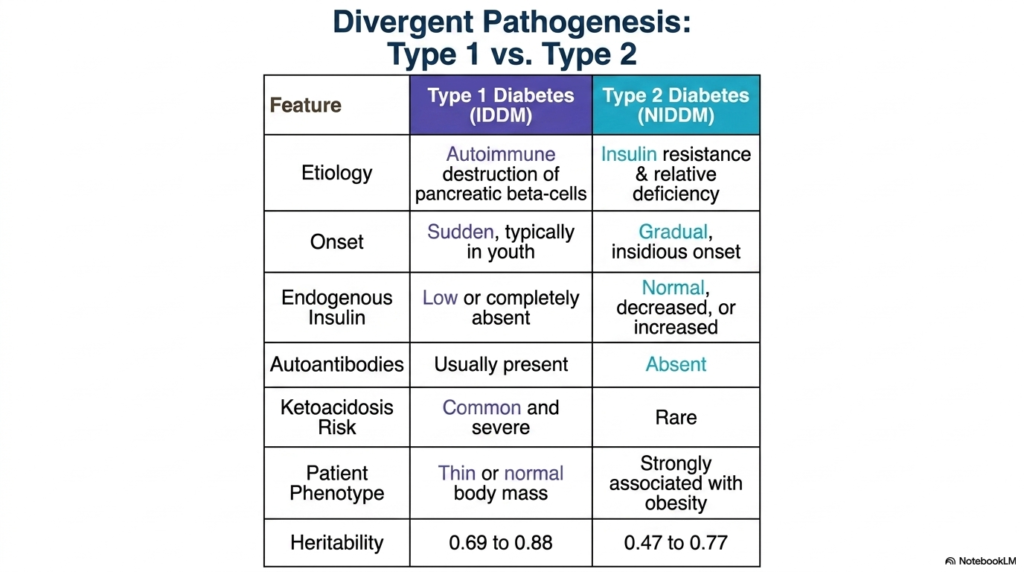
Not all metabolic failures share the same origin story, even if the clinical endpoint looks remarkably similar. The core purpose of this slide is to clearly contrast the distinct etiological and biochemical pathways underlying the two primary forms of this disease. By examining this comparative table, students can differentiate the underlying mechanisms of metabolic collapse.
Type 1 Diabetes Mellitus is characterized by a sudden, complete loss of insulin production. The etiology is fundamentally autoimmune; the body’s own immune system erroneously identifies pancreatic beta cells as foreign and systematically destroys them. Consequently, endogenous insulin levels drop to zero. Patients usually present with a thin or normal body mass and require immediate, lifelong exogenous insulin therapy to survive. The presence of circulating autoantibodies is a definitive diagnostic marker of this specific pathogenesis.
In stark contrast, Type 2 Diabetes Mellitus is primarily driven by peripheral tissue resistance to insulin, paired with a gradual, relative decline in insulin secretion. The onset is highly insidious, often developing silently over many years. In these patients, endogenous insulin levels may be normal or even markedly elevated in the early stages, as the pancreas attempts to overcome cellular resistance. This form of the disease is heavily associated with obesity, lifestyle factors, and metabolic syndrome.
Both divergent pathways inevitably lead to the severe systemic condition known as Diabetes Mellitus, but they carry different acute risks. Because Type 1 features a total absence of insulin, the biochemical brakes on lipid breakdown are completely removed, making patients highly susceptible to common and severe diabetic ketoacidosis. Type 2 patients, retaining a small degree of insulin signaling, rarely develop ketoacidosis, though they still suffer the long-term consequences of chronic hyperglycemia.
Slide 6: Carbohydrate Dysregulation in Diabetes Mellitus: The Hyperglycemic Cascade
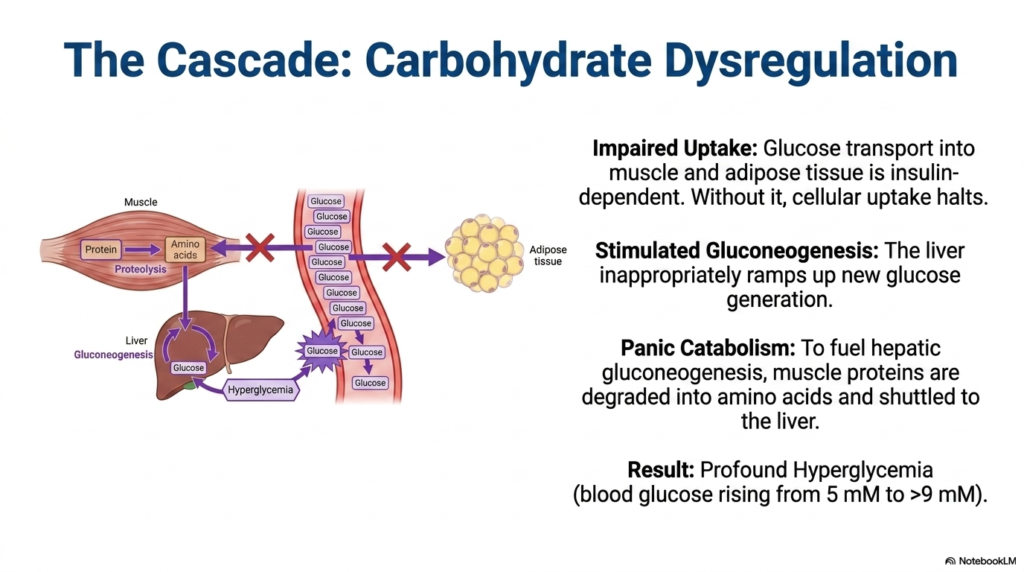
What happens to a biological engine when its primary fuel supply is locked outside the combustion chamber? The core purpose of this slide is to map the immediate biochemical consequences of insulin deficiency on carbohydrate metabolism and tissue starvation. It visually traces the cascade that inevitably leads to profound and dangerous hyperglycemia.
The most immediate consequence of a lack of insulin is massively impaired glucose uptake. Transport proteins, specifically GLUT4 in muscle and adipose tissue, require an active insulin signal to translocate to the cellular membrane. Without this hormonal key, the transport mechanisms halt entirely. Glucose pools uselessly in the bloodstream while the muscle and fat cells experience acute intracellular starvation. This cellular hunger is a defining biochemical hallmark of uncontrolled Diabetes Mellitus.
Sensing this deep cellular starvation, the body initiates desperate counter-regulatory measures. The liver, lacking the inhibitory signal of insulin, inappropriately ramps up gluconeogenesis, synthesizing brand new glucose molecules and dumping them into the already saturated bloodstream. Simultaneously, panic catabolism sets in. Muscle tissue begins rapidly degrading its own structural proteins into free amino acids. These amino acids are shuttled directly to the liver to fuel even more gluconeogenesis, exacerbating the crisis of Diabetes Mellitus.
The ultimate result of this multi-tissue miscommunication is profound hyperglycemia. The blood glucose concentration, normally tightly regulated around 5 millimolar, rapidly climbs to 9 millimolar or higher. The body is essentially cannibalizing its own muscle mass to produce fuel that it cannot even absorb. Understanding this vicious cycle is absolutely critical for managing Diabetes Mellitus, as it illustrates why patients experience severe muscle wasting and persistent high blood sugar simultaneously.
Slide 7: Renal Threshold Overflow in Diabetes Mellitus: The Mechanics of Osmotic Diuresis
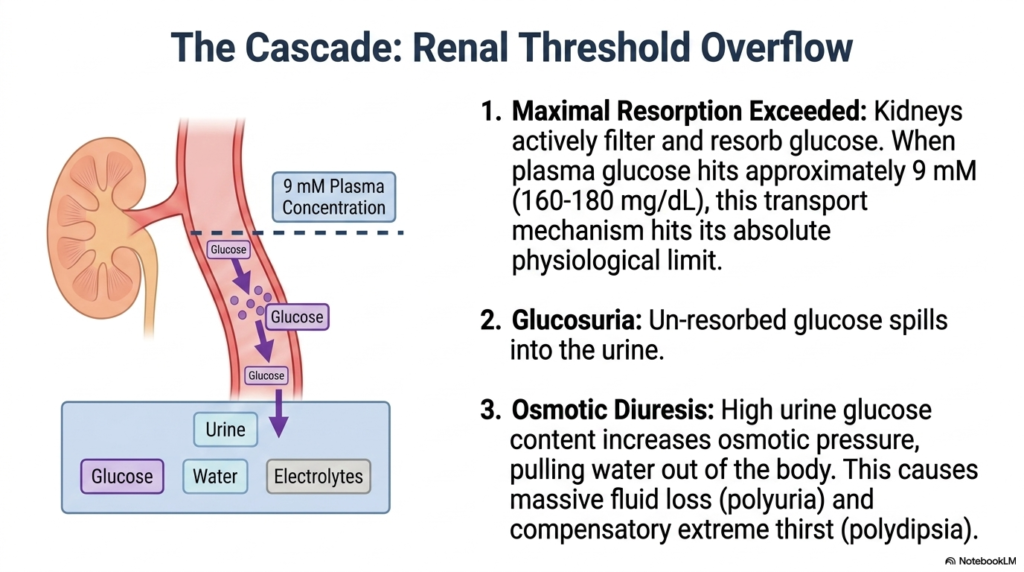
The human kidney is a phenomenal biological filter, but every filtration system has an absolute mechanical breaking point. The core purpose of this slide is to logically explain the renal consequences of severe hyperglycemia and the classic, visible symptoms it produces. It bridges the gap between microscopic blood chemistry and macroscopic patient presentation.
Under normal conditions, the kidneys actively filter and completely resorb all circulating glucose back into the bloodstream. However, in uncontrolled Diabetes Mellitus, this elegant transport mechanism hits its absolute physiological limit. When plasma glucose concentration reaches approximately 9 millimolar (roughly 160 to 180 milligrams per deciliter), the renal transport proteins become entirely saturated. The system simply cannot pump the massive volume of sugar back into the blood fast enough.
Once this maximal resorption capacity is exceeded, the unresorbed glucose spills over into the urine, a clinical condition known as glucosuria. This is a definitive historical and biochemical marker of Diabetes Mellitus. The presence of heavy glucose in the urinary tract radically alters the local osmotic balance. Because glucose is an osmotically active solute, it acts like a chemical sponge, drawing water out of surrounding tissues and into the renal tubules.
This fluid shift triggers profound osmotic diuresis. The patient experiences massive, uncontrollable fluid loss, leading to the classic symptom of polyuria, or excessive urination. To compensate for this rapid and dangerous dehydration, the brain’s thirst centers are intensely stimulated, causing polydipsia, or extreme thirst. For a student studying Diabetes Mellitus, grasping this osmotic mechanism perfectly explains why patients present with unending thirst and constant urination despite drinking gallons of water.
Slide 8: Lipid Dysregulation in Diabetes Mellitus: The Collapse of Fat Storage
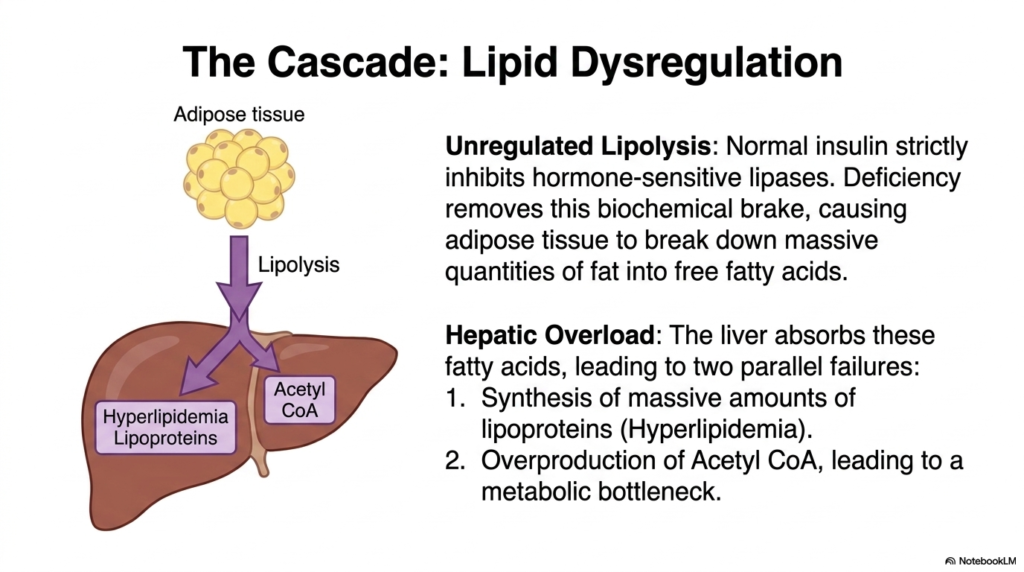
Adipose tissue is not merely a passive storage depot; it is a highly active metabolic organ constantly awaiting hormonal signals. The core purpose of this slide is to explore how the loss of insulin signaling unleashes unregulated and catastrophic fat breakdown. It illustrates the dangerous biochemical shift from carbohydrate reliance to toxic lipid overload.
In a healthy state, normal baseline insulin strictly inhibits the activity of hormone-sensitive lipases within fat cells, effectively keeping stored triglycerides safely locked away. A severe insulin deficiency removes this biochemical brake. Adipose tissue, sensing a false state of starvation, begins to rapidly and massively break down stored fat into circulating free fatty acids. This unregulated lipolysis is a major pathogenic driver in untreated Diabetes Mellitus.
These mobilized free fatty acids flood the bloodstream and are rapidly taken up by the liver, triggering two parallel metabolic failures. First, the liver attempts to repackage these lipids, leading to the synthesis of massive amounts of lipoproteins. This generates profound systemic hyperlipidemia, which significantly increases the patient’s cardiovascular risk. Lipid dysregulation is therefore a central, life-threatening component of the overall pathology of Diabetes Mellitus.
Secondly, the liver begins to aggressively oxidize these fatty acids, leading to an immense overproduction of Acetyl-CoA. This creates a severe metabolic bottleneck within the hepatic mitochondria. The normal energy cycles are completely overwhelmed by the sheer volume of lipid breakdown products. For biochemistry students, understanding this hepatic overload in Diabetes Mellitus is critical, as it sets the stage for the most dangerous acute complication of the disease: systemic ketoacidosis.
Slide 9: Ketogenesis and Acidosis in Diabetes Mellitus: The Danger of Metabolic Starvation
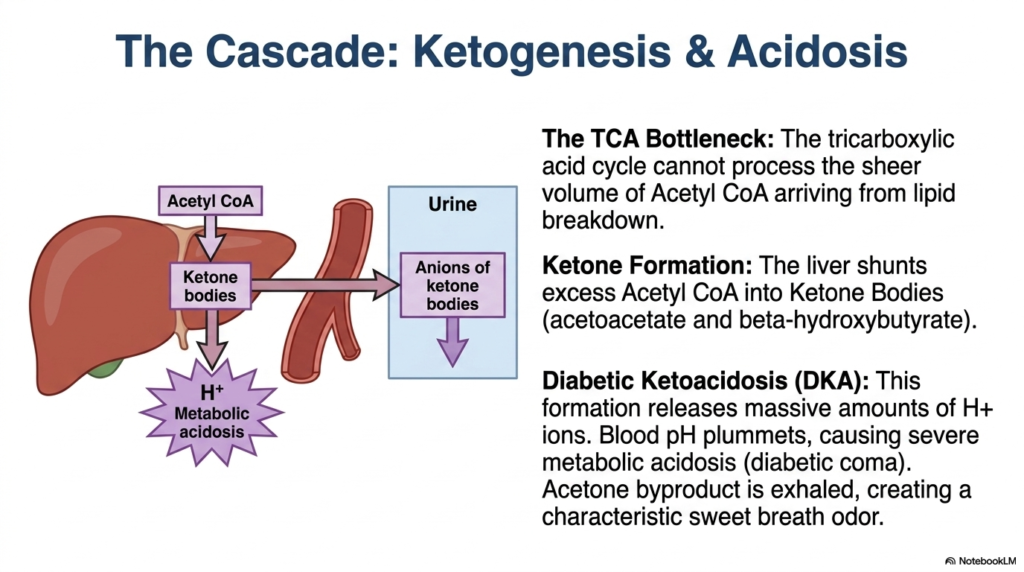
Desperate biochemical times call for desperate cellular measures, but these physiological backup plans can easily turn deadly. The core purpose of this slide is to detail the metabolic shift toward ketone body production and its severe acidic consequences. It explains the mechanics behind a life-threatening medical emergency driven by lipid overload.
The massive influx of free fatty acids into the liver generates an overwhelming amount of Acetyl CoA. The tricarboxylic acid (TCA) cycle, the cell’s standard metabolic engine, lacks the capacity to process the sheer volume of this molecule produced by unregulated lipid breakdown. Faced with this massive bottleneck, the liver is forced to activate an alternative biochemical shunt. This pathway is a defining characteristic of severe, uncontrolled Diabetes Mellitus.
To clear the backlog, the liver shunts excess Acetyl-CoA into the synthesis of ketone bodies, primarily acetoacetate and beta-hydroxybutyrate. While ketones can serve as an emergency fuel source for the brain, their rapid overproduction creates a catastrophic problem. The formation of these specific molecules releases massive quantities of hydrogen ions into the bloodstream. In the context of Diabetes Mellitus, this overwhelms the body’s natural pH-buffering systems.
As hydrogen ions accumulate, the blood pH plummets, plunging the patient into severe metabolic acidosis, clinically known as Diabetic Ketoacidosis (DKA). If left untreated, this acidic environment depresses the central nervous system, leading to a diabetic coma and eventual death. Additionally, the spontaneous breakdown of acetoacetate produces acetone gas, which is exhaled through the lungs, giving patients with severe Diabetes Mellitus a highly characteristic, sweet, fruity breath odor.
Slide 10: Systemic Complications of Diabetes Mellitus: The Long-Term Toll on Microvasculature
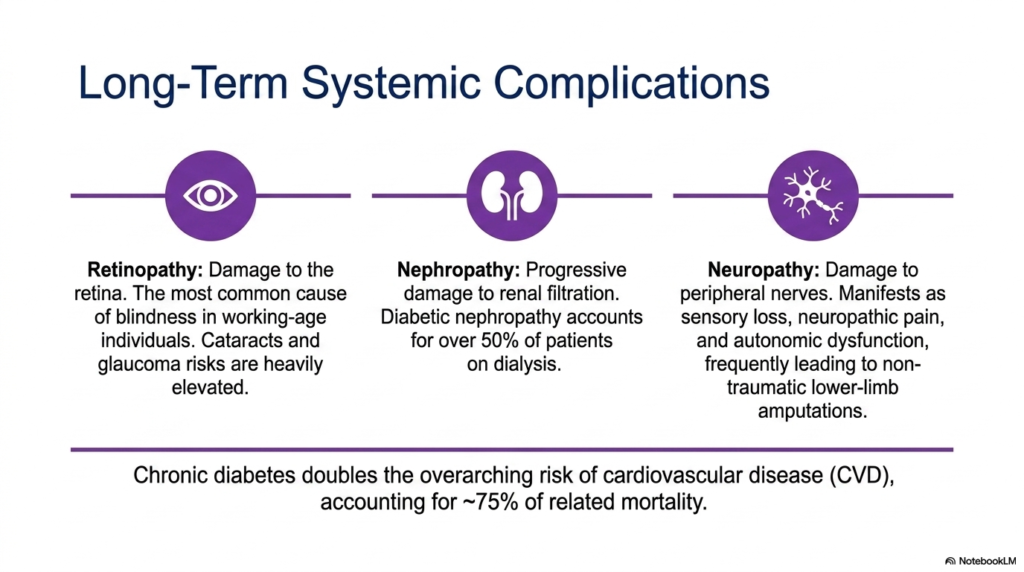
A chronic, unrelenting flood of sugar in the bloodstream acts like a slow-moving toxin to the body’s most delicate tissues. The core purpose of this slide is to outline the devastating long-term consequences of uncontrolled hyperglycemia across multiple organ systems. It highlights the systemic, overarching morbidity that patients face over a lifetime.
The microvascular damage caused by Diabetes Mellitus is particularly severe in the eyes, leading to diabetic retinopathy. High blood glucose damages the fragile capillaries of the retina, causing leaks, swelling, and abnormal vessel growth. This progressive damage makes it the most common cause of non-traumatic blindness in working-age individuals worldwide. Furthermore, the overarching metabolic stress dramatically elevates the risks for developing cataracts and glaucoma much earlier in life.
Similarly, the kidneys’ intricate filtration network is highly vulnerable to sugar toxicity. Diabetic nephropathy represents progressive, irreversible damage to the renal glomeruli, steadily reducing filtration efficiency. As the damage compounds over decades, patients often progress to end-stage renal disease. Strikingly, the chronic vascular damage wrought by Diabetes Mellitus accounts for over half of all patients currently requiring life-sustaining dialysis in the medical system today.
Beyond the eyes and kidneys, peripheral nerves suffer immensely. Diabetic neuropathy manifests as severe sensory loss, agonizing neuropathic pain, and dangerous autonomic dysfunction. Because patients lose protective pain sensation in their extremities, minor cuts frequently ulcerate and become severely infected, frequently leading to non-traumatic lower-limb amputations. Ultimately, chronic Diabetes Mellitus doubles the overarching risk of cardiovascular disease, which tragically accounts for roughly three-quarters of all disease-related mortality.
Slide 11: Diagnostic Biochemistry of Diabetes Mellitus: Quantifying Metabolic Failure
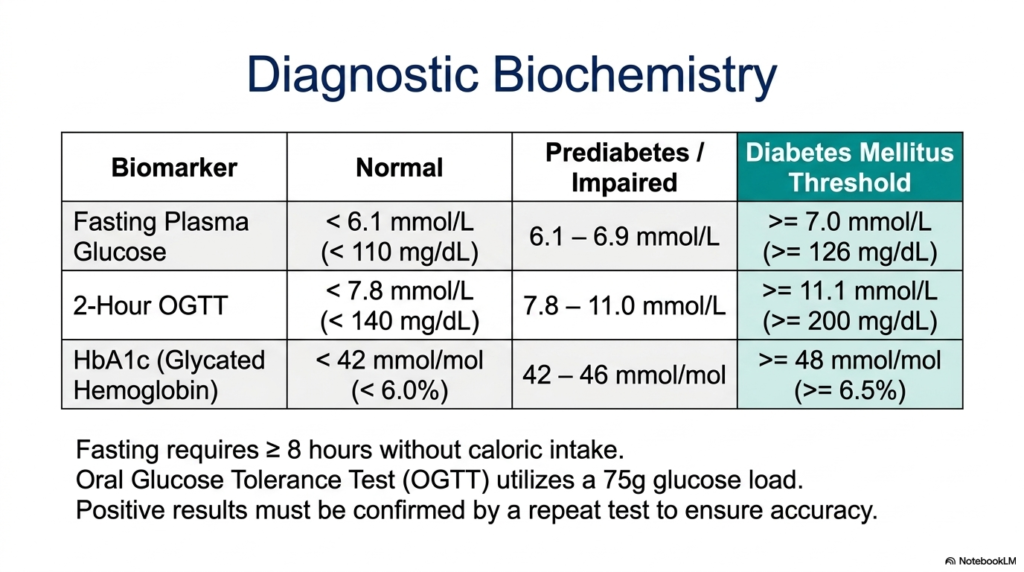
How does the modern medical field precisely measure an otherwise invisible metabolic collapse? The core purpose of this slide is to thoroughly decode the clinical biomarkers used to definitively diagnose and monitor this complex endocrine disorder. It translates abstract biochemical failures into strict numerical thresholds used by physicians worldwide.
The most fundamental metric is the Fasting Plasma Glucose test. This requires the patient to fast for at least 8 hours, ensuring that no recent caloric intake affects the reading. A normal fasting baseline is below 6.1 millimolar. However, if the plasma glucose reads at or above 7.0 millimolar (126 milligrams per deciliter), it indicates a profound failure of basal insulin signaling. This threshold is a primary diagnostic criterion for the identification of active Diabetes Mellitus.
Alternatively, the 2-Hour Oral Glucose Tolerance Test (OGTT) dynamically stresses the metabolic system. The patient ingests a heavy 75-gram glucose load, and clearance is measured two hours later. A healthy pancreas easily clears this challenge, maintaining levels below 7.8 millimolar. If the reading remains at or above 11.1 millimolar (200 milligrams per deciliter), it proves the cellular gates cannot open. This severely impaired clearance definitively confirms a diagnosis of Diabetes Mellitus.
Finally, the HbA1c test measures glycated hemoglobin, offering a long-term biochemical snapshot. Because glucose irreversibly binds to red blood cells over their three-month lifespan, an HbA1c level of 6.5 percent or higher indicates chronic, sustained hyperglycemia. For patients managing Diabetes Mellitus, this specific biomarker is invaluable, as it accurately reflects their metabolic control over time. All positive results across these three modalities must be carefully confirmed by repeat testing.
Slide 12: The Metabolic Map of Diabetes Mellitus: Synthesizing the Systemic Breakdown
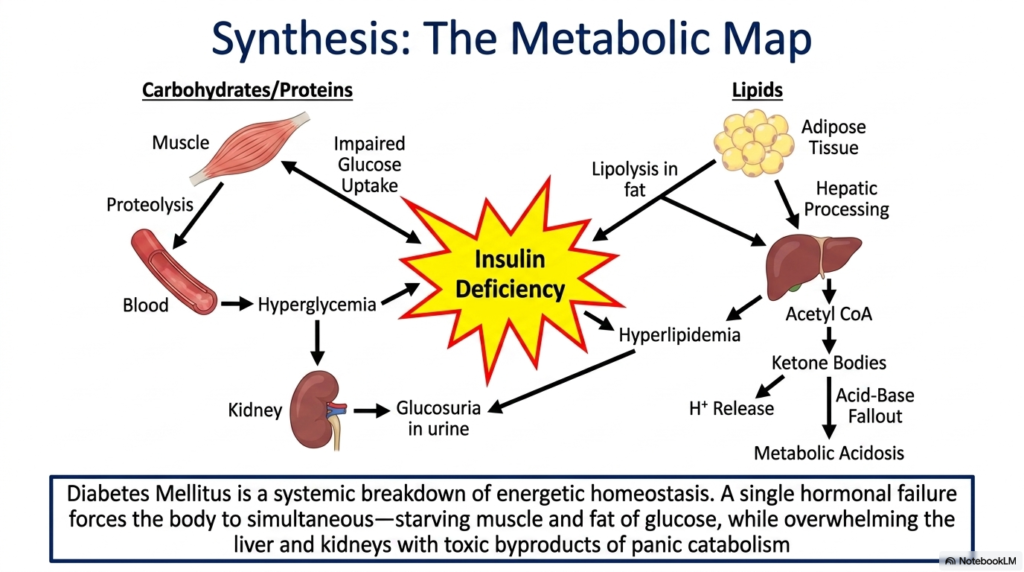
To truly comprehend a complex disease, one must zoom out from individual molecules and view the entire interconnected landscape of human physiology. The core purpose of this final slide is to powerfully synthesize all previously discussed biochemical pathways into one comprehensive metabolic map. It serves as a visual capstone for the entire lecture.
The diagram perfectly illustrates that Diabetes Mellitus is not a localized organ issue, but rather a systemic breakdown of energetic homeostasis. A single hormonal failure—insulin deficiency—forces the body into simultaneous, contradictory states. The muscle and adipose tissues are literally starving for glucose because uptake is severely impaired. Concurrently, the liver is tricked into panic mode, generating excess sugar and processing massive amounts of lipids from rapid fat breakdown.
This singular deficiency triggers a multipronged biochemical disaster. The resulting hyperglycemia overflows the kidneys, causing urinary glucose and water loss. Simultaneously, the lipid overflow leads to systemic hyperlipidemia and floods the liver with Acetyl CoA. The hepatic processing of this toxic overload generates dangerous ketone bodies, dropping the blood pH and causing severe metabolic acidosis. Diabetes Mellitus fundamentally rewires the body to destroy its own tissues.
Ultimately, understanding this map is the key to understanding the patient. The fatigue, the thirst, the weight loss, and the dangerous acidity are not isolated symptoms; they are deeply connected branches of the same broken biochemical tree. By mastering the molecular mechanisms of Diabetes Mellitus, future scientists and clinicians are fully equipped to intervene, repair the broken pathways, and ultimately save the lives of those suffering from this devastating disease.
Please read our Content Disclaimer Statement.
Check out our social media channels: