9. Monosaccharides and Disaccharides: Structure and Examples
When the general public hears the word “sugar,” their minds naturally gravitate toward sweet snacks, empty calories, or modern dietary guidelines. But in the demanding, awe-inspiring world of advanced biochemistry, sugars are the intricate, three-dimensional architectural scaffolds that quite literally hold cellular life together. From the rapid firing of your neuronal networks to the stable preservation of your DNA across countless generations, carbohydrate structures dictate biological destiny.
The core purpose of this detailed slide deck is to break down the highly complex world of carbohydrate chemistry into digestible, logical blueprints. We are going to thoroughly explore the stereochemistry, functional modifications, and powerful enzymatic bonds that transform simple carbon rings into dynamic biological machines. Let’s dive deeply into the subject and decode the structural secrets of cellular life!
Slide 1: Biochemical Blueprints of Monosaccharides and Disaccharides
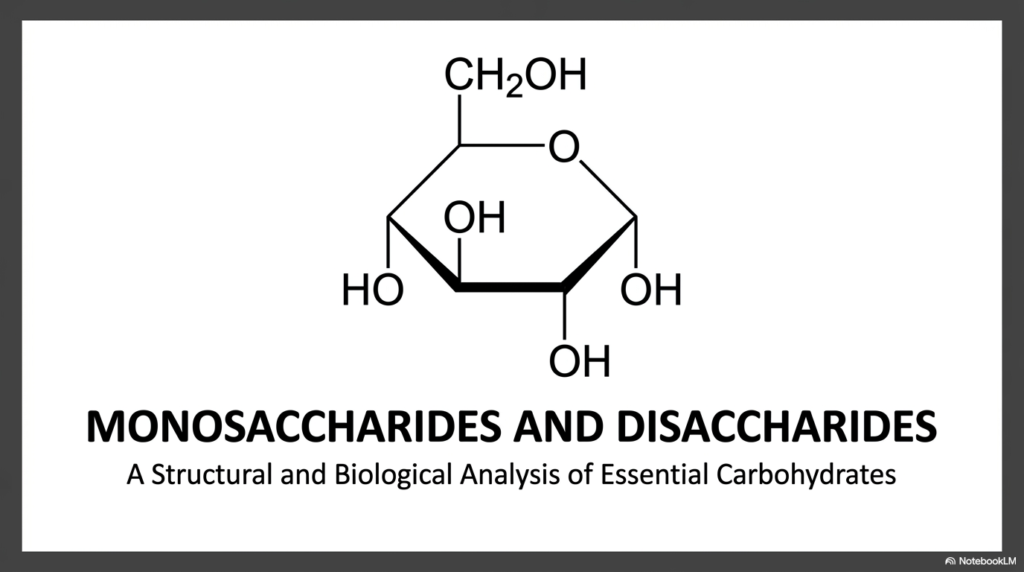
Welcome to our visual journey! When you look at the title slide, you are looking at the foundational architecture of life itself. The rigorous study of Monosaccharides and Disaccharides goes way beyond simply memorizing flat metabolic pathways; it is about clearly visualizing three-dimensional molecular blueprints. This initial image perfectly sets the stage for our comprehensive biological and structural analysis. If you want to truly master advanced biochemistry, you have to start thinking in dynamic 3D.
The intricate skeletal structure highlighted centrally on the slide showcases the precise spatial distribution of highly reactive hydroxyl groups. In the complex realm of Monosaccharides and Disaccharides, the exact orientation of these functional groups changes absolutely everything. A single flip can turn a vital, life-saving energy source into an indigestible structural fiber.
Let’s dive deeper into what this visual actually represents for your future careers. In your introductory textbooks, you often see flat, lifeless hexagons. But biological reality is incredibly dynamic. The core purpose of rigorously examining Monosaccharides and Disaccharides is to recognize them as highly reactive, precise spatial entities. Every single carbon atom, every oxygen bridge, and every stereochemical angle is purposefully designed by evolutionary nature. Throughout this comprehensive slide deck, we will systematically break down the structural and biological analysis of these essential carbohydrates.
You will quickly learn to appreciate that the intense study of Monosaccharides and Disaccharides is fundamentally the study of life’s biological scaffolding. Whether it is the rapid burst of energy that powers a beating heart cell or the rigid structural matrix that gives a towering redwood tree its immense physical strength, it all starts right here with these foundational carbon rings. By the time we finish this visual and scientific journey, you will clearly see exactly why mastering the chemical architecture of Monosaccharides and Disaccharides is absolutely crucial for your future medical and scientific success. Let’s decode these structures together!
Slide 2: Structural Taxonomy in Monosaccharides and Disaccharides
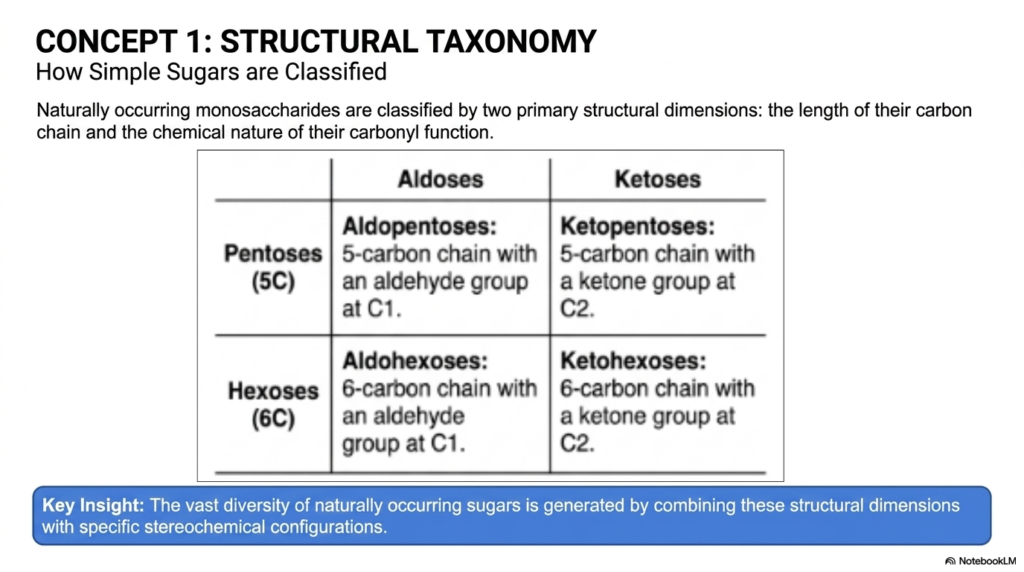
To make sense of the vast, intimidating world of Monosaccharides and Disaccharides, we first need a highly reliable system of structural organization. Think of this slide as your biological filing cabinet. Concept 1 formally introduces the structural taxonomy used by chemists to cleanly classify simple sugars. To communicate effectively as a medical professional or laboratory researcher, you must speak the specialized language of Monosaccharides and Disaccharides fluently. Naturally occurring monosaccharides are primarily organized across two major structural dimensions.
First, we carefully look at the absolute length of their carbon chain. In human and plant biochemistry, we primarily focus on pentoses, five-carbon sugars that play critical structural and genetic roles. Then we have hexoses, our massive six-carbon heavyweights that robustly drive cellular energy production and create immense global biomass.
The second essential dimension in accurately classifying Monosaccharides and Disaccharides is the precise chemical nature of their carbonyl function. If a sugar has a terminal aldehyde group—meaning the reactive carbonyl is securely locked at the very end of the carbon chain at the C-1 position—it belongs to the aldose family. On the flip side, if the biological sugar contains an internal ketone group, typically positioned explicitly at the C-2 carbon, it is universally designated as a ketose.
Why does this fundamental taxonomy matter for your advanced studies? The key insight highlighted on the slide is that incredible, awe-inspiring biological diversity arises simply from elegantly combining these basic structural dimensions with highly specific stereochemical configurations. When you look at an unknown carbohydrate structure in a clinical diagnostic lab, you should immediately and instinctively ask yourself: Is this an aldohexose? Is it a ketopentose? This rigorous structural taxonomy is the universal foundation for literally everything we do in the advanced study of Monosaccharides and Disaccharides.
By mastering this classification matrix early in your training, you will be able to predict how different sugars will behave in an aqueous cellular environment and how they will interact with specific metabolic enzymes.
Slide 3: Aldopentoses and the Genetics of Monosaccharides and Disaccharides
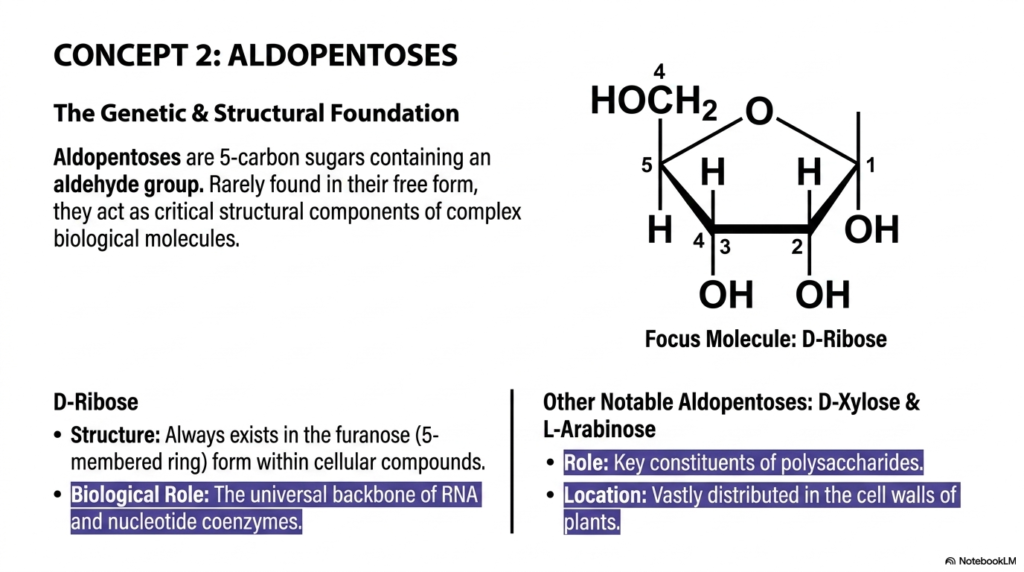
Moving dynamically to Concept 2, we are zooming in on a highly specific and foundational class of molecules: the powerful aldopentoses. As we continue to rapidly explore the vast, diverse catalog of Monosaccharides and Disaccharides, it becomes immediately clear that evolutionary biology uses highly specific sugar shapes for highly specific functional tasks. Aldopentoses are unique, precisely shaped five-carbon sugars containing a highly reactive aldehyde functional group. While you will very rarely find these particular sugars floating freely in human blood plasma or flowing plant sap, their underlying structural importance within the broader family of Monosaccharides and Disaccharides is absolutely monumental. These are not your typical daily dietary sugars; they are the immortal genetic architects of all known cellular life.
The absolute, undisputed star of this particular slide is D-Ribose. From a strict structural standpoint, when D-ribose is incorporated into living cellular compounds, it almost exclusively exists in its furanose form, a compact, highly stable five-membered ring. Its overarching biological role is genuinely staggering. It serves as the universal, unyielding molecular backbone of RNA and as a critical component of nucleotide coenzymes like ATP, NADH, and FADH2. Simply put, without the precise chemical geometry of this highly specific member of the Monosaccharides and Disaccharides family, cellular genetics, rapid protein transcription, and biological energy transfer would instantly cease to exist.
Beyond our genetic hero D-ribose, this slide also highlights other highly notable aldopentoses, specifically D-xylose and L-arabinose. While these might not appear regularly on your standard metabolic blood panels, they are incredibly important in the advanced botanical study of Monosaccharides and Disaccharides. They serve as essential constituents of massive structural polysaccharides and are widely distributed in the incredibly rigid cell walls of terrestrial plants worldwide. For those of you heading into agricultural science, modern pharmacology, or advanced plant biochemistry, understanding these structural sugars is crucial. They literally dictate how immense trees stand tall against the relentless pull of gravity and even fundamentally alter how human gastrointestinal tracts process complex dietary fibers.
Slide 4: Aldohexoses: The Powerhouse Monosaccharides and Disaccharides
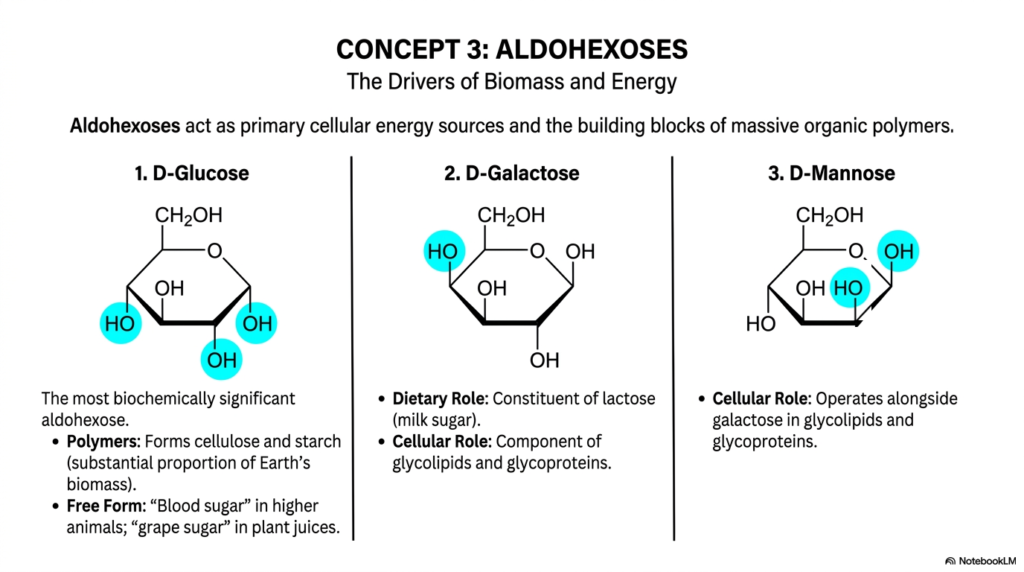
Concept 3 formally brings us face-to-face with the absolute metabolic heavyweights of cellular biology: the mighty aldohexoses. In any comprehensive, serious academic discussion regarding Monosaccharides and Disaccharides, the aldohexoses rightfully and undeniably take center stage. These six-carbon, aldehyde-containing sugars serve as robust primary cellular energy sources and fundamental, endlessly repeating building blocks of large organic polymers. Thoroughly understanding the dynamic physiological behaviors of these highly specific Monosaccharides and Disaccharides is a non-negotiable core requirement for truly mastering both human disease physiology and complex plant metabolism.
Let’s carefully and systematically dissect the three primary structural examples cleanly shown on the slide. First, we have D-Glucose, which is unquestionably the single most biochemically significant aldohexose on Earth. It is universally known as “blood sugar” in higher mammals and “grape sugar” in plant juices. Its incredible enzymatic ability to endlessly polymerize produces cellulose and starch, which together make up a substantial proportion of our planet’s total terrestrial biomass.
Next up is D-Galactose, which differs chemically from standard glucose only at a single carbon atom—it is what we formally call a C-4 epimer. It plays a crucial dietary role as a core constituent of milk sugar and serves as a vital, identifying signaling component of complex glycolipids and glycoproteins embedded in human cellular membranes.
Finally, we systematically examine D-Mannose, another crucial epimer of glucose that is structurally altered specifically at the C-2 position. D-mannose works closely alongside galactose in highly specialized cellular roles, particularly in the proper folding and formation of complex glycoproteins. When carefully considering the vast clinical and medical relevance of Monosaccharides and Disaccharides, understanding these seemingly minor stereochemical differences is absolutely critical for your future medical training. Just one single hydroxyl group pointing the wrong way completely changes how the human body metabolizes the fuel! The precise hormonal regulation of D-glucose is the central physiological mechanism in successfully managing diabetes mellitus, a devastating disease you will actively treat daily.
Slide 5: Ketoses as Metabolic Reserves in Monosaccharides and Disaccharides
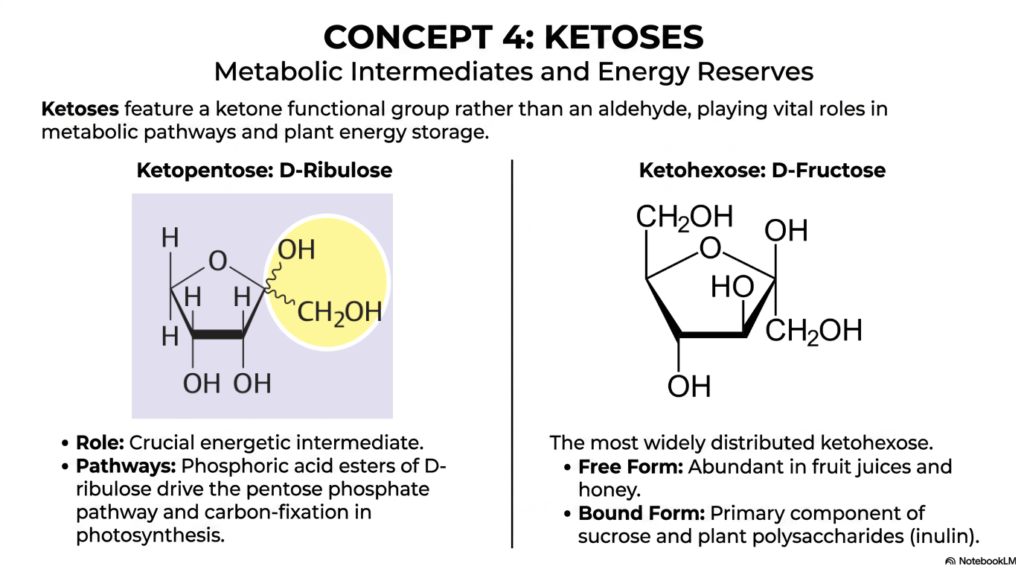
In Concept 4, we structurally pivot away from the highly common aldehydes and focus our sharp scientific attention entirely on the versatile ketoses. As we rapidly and methodically continue exploring the vast, intricate catalog of Monosaccharides and Disaccharides, it is absolutely vital to explicitly recognize exactly how functional group placement chemically alters a molecule’s long-term metabolic destiny. Ketoses boldly feature an internal ketone functional group—typically locked firmly at the C-2 position—rather than a standard terminal aldehyde. This subtle yet profound chemical shift elegantly confers distinct roles on them as crucial metabolic intermediates and major biological energy reserves within the dynamic, fascinating realm of Monosaccharides and Disaccharides.
The slide visually highlights two major biological players that you absolutely need to commit to memory: D-Ribulose, a highly specific ketopentose, and D-Fructose, a ketohexose. Let’s closely examine D-Ribulose first. It is an absolutely crucial, high-energy intermediate for almost all plant life on Earth. When actively phosphorylated into phosphoric acid esters by cellular enzymes, D-ribulose vigorously drives the complex pentose phosphate pathway and serves as the primary, irreplaceable molecule for carbon fixation during plant photosynthesis. It is a stunning, undeniable biochemical fact that every single carbon atom currently in your human body was, at some ancient evolutionary point, captured by a plant actively using a chemical derivative of this exact sugar!
On the other hand, D-Fructose is the most widely distributed ketohexose in nature. In its free-circulating molecular form, it is highly abundant and incredibly sweet to the human palate, found predominantly in wild fruit juices and natural honey. However, in its tightly bound, highly polymerized form, D-fructose serves as the primary structural component of common plant polysaccharides, such as inulin and everyday table sugar. For any ambitious medical student actively analyzing the clinical impact of Monosaccharides and Disaccharides, understanding human fructose metabolism is a high-yield topic. Because of its highly unique ketose structure, it actively bypasses major regulatory enzymes in human glycolysis, leading to immense clinical implications for modern obesity and metabolic syndrome.
Slide 6: Deoxyaldoses and Information Storage in Monosaccharides and Disaccharides
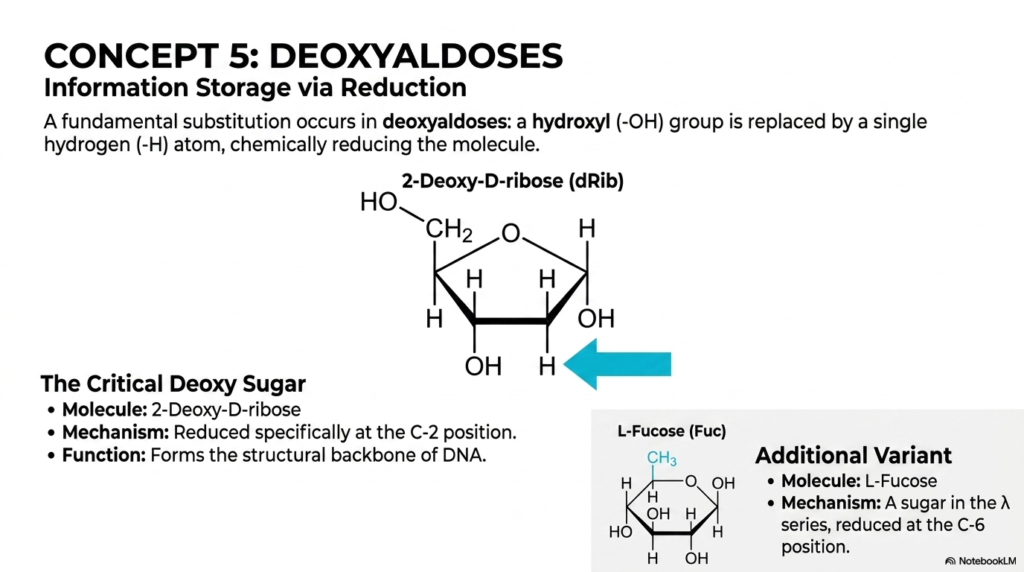
Concept 5 smoothly and elegantly introduces a highly subtle but structurally transformative chemical modification: the evolutionary creation of deoxyaldoses. When thoroughly and comprehensively studying the full functional spectrum of Monosaccharides and Disaccharides, we frequently see that cellular biology incredibly often subtracts specific atoms to expertly create entirely new, highly specialized biological functions. In a deoxyaldose, a fundamental chemical substitution occurs in which a highly reactive hydroxyl (-OH) group is directly and permanently replaced by a single, highly stable hydrogen (-H) atom, thereby reducing the entire molecule. This intentional, enzyme-driven chemical reduction is a brilliant evolutionary adaptation uniquely found within the broad, diverse class of Monosaccharides and Disaccharides.
The absolute most critical, world-changing deoxy sugar clearly shown here on the slide is 2-Deoxy-D-ribose. In this highly specific, legendary molecule, the chemical reduction happens exclusively at the C-2 position. Why is this structurally and medically important for all biology? The permanent removal of that highly reactive oxygen atom drastically and permanently increases the long-term chemical stability of the delicate five-membered sugar ring. Because of this newly added durability, 2-Deoxy-D-ribose was actively chosen by nature over billions of years to flawlessly form the unyielding, immortal structural backbone of DNA. The long-term, multi-generational storage of complex genetic information strongly demands a highly robust molecule, and this specific modification ensures that our genetic code does not violently degrade inside the nucleus.
Furthermore, the slide importantly presents an additional, highly specialized cellular variant: L-Fucose. This is a complex, deeply fascinating sugar that is explicitly reduced at the C-6 position, where it permanently terminates with a chemically stable methyl group rather than a standard hydroxyl. Among all the rigorously studied Monosaccharides and Disaccharides, L-Fucose is highly unique because it operates primarily in complex cellular recognition and sophisticated immunological signaling pathways. It is a very common, absolutely necessary component firmly found on the outer protective surfaces of human red blood cells, directly contributing to the ABO blood group antigens. Understanding these missing oxygen atoms is critical for mastering both human genetics and safe hospital blood transfusions!
Slide 7: Acetylated Amino Sugars in Monosaccharides and Disaccharides
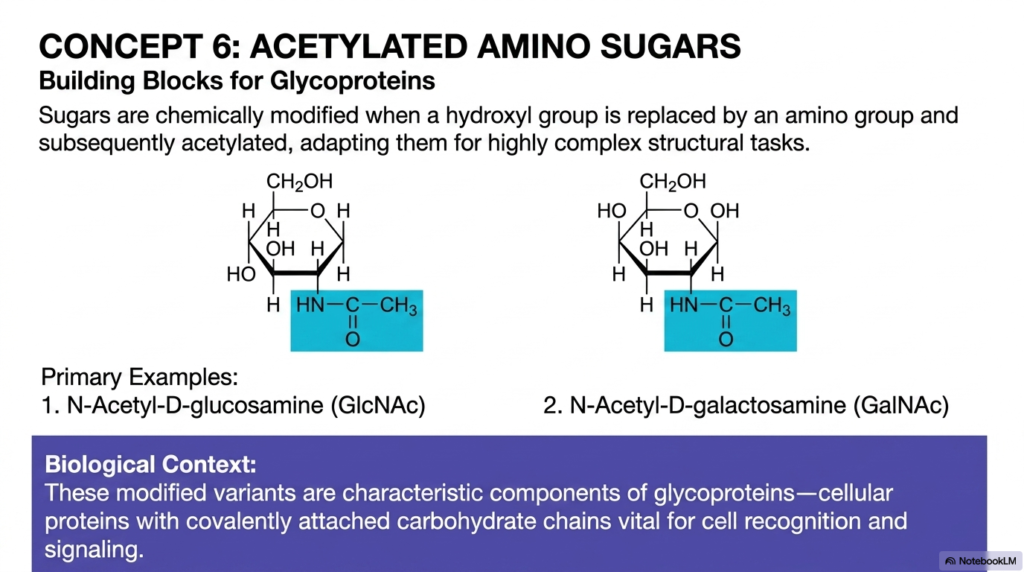
Concept 6 boldly takes us deep into the endlessly fascinating realm of highly complex chemical modifications with acetylated amino sugars. To successfully and reliably build advanced, resilient biological structures using Monosaccharides and Disaccharides, living cells frequently and intentionally modify simple sugars to perfectly adapt them for highly specialized, physically demanding structural tasks. In these incredible, robust biological variants, a standard hydroxyl group is first biochemically replaced by a functional amino group, and then subsequently heavily acetylated by highly specific cellular enzymes. These rapid, sequential enzymatic steps create incredibly durable molecules that easily represent the undisputed structural elite of all Monosaccharides and Disaccharides.
Our primary, high-yield clinical examples here are N-Acetyl-D-glucosamine, commonly abbreviated as GlcNAc in the clinical literature, and N-Acetyl-D-galactosamine, widely known as GalNAc. By permanently and tightly adding the bulky, nitrogen-containing acetyl group, the basic, underlying chemical properties of the hexose ring are entirely and irreversibly transformed. In their natural, dynamic biological context, these heavily modified variants are the characteristic, endlessly repeating building blocks of glycoproteins. Glycoproteins are highly complex, essential cellular proteins deliberately adorned with covalently attached carbohydrate chains. They actively act as the protective, highly interactive barrier on the outside of your human cells, playing absolutely vital, irreplaceable roles in cell-to-cell recognition and targeted human immunity.
For anyone rigorously and seriously studying the clinical applications of Monosaccharides and Disaccharides, these modified molecules are highly relevant in active hospital settings. They are definitely not just simple cellular markers. In the human body, these tough, highly adapted sugars actively form a major, structural, shock-absorbing part of the resilient cartilage in our active joints and the protective, incredibly thick mucus continuously lining our harsh gastrointestinal tract. By biochemically replacing a simple oxygen atom with a bulky, resilient acetylated nitrogen group, the human body successfully prevents rapid enzymatic degradation, giving these vital components an incredibly long, highly functional lifespan.
Slide 8: Acidic and Alcoholic Modifications in Monosaccharides and Disaccharides
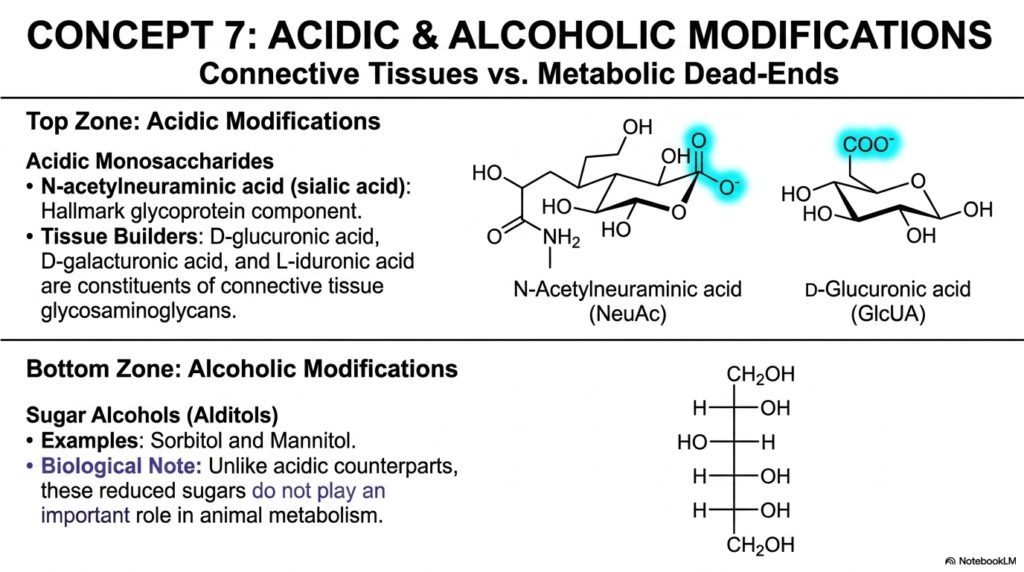
Concept 7 systematically divides our focused scientific attention clearly between two entirely distinct, wildly opposing chemical modifications: acidic and alcoholic alterations. The diverse, incredibly expansive evolutionary toolkit of Monosaccharides and Disaccharides relies heavily on such deliberate chemical alterations to build complex, highly functional mammalian bodies. By actively adding massive negative electrical charges or fully chemically reducing the sugars into basic alcohols, cellular biology effortlessly creates highly specialized molecular tools. Thoroughly understanding these directly contrasting modifications provides deep, highly actionable insight into exactly how Monosaccharides and Disaccharides function in both dense tissue architecture and highly specific, sometimes highly dangerous, metabolic pathways.
The highly visual “Top Zone” clearly features critical acidic modifications. Here, we meticulously analyze N-acetylneuraminic acid (commonly known clinically as sialic acid) and D-glucuronic acid. These highly acidic monosaccharides carry a strong negative electrical charge at standard human physiological pH. Sialic acid is an absolute hallmark component of glycoproteins, frequently capping the vulnerable ends of sugar chains on cell surfaces to successfully prevent premature degradation by circulating, destructive blood enzymes. D-glucuronic acid, on the other hand, is a massive, incredibly strong biological tissue builder. It is a critical, continuously repeating constituent of connective tissue glycosaminoglycans, the spongy matrix molecules that make up our flexible cartilage and joint fluid.
Conversely, the “Bottom Zone” elegantly highlights the exact opposite: aggressive alcoholic modifications producing highly reduced sugar alcohols known universally to biochemists as alditols, such as sorbitol and mannitol. The highly specific biological note on the slide is incredibly striking for the broader clinical context of Monosaccharides and Disaccharides: unlike their vital acidic counterparts, these fully reduced sugars typically do not play an important structural or energetic role in normal, healthy animal metabolism. In fact, they are very often considered dangerously toxic metabolic dead ends. For example, in poorly controlled clinical diabetes, excess blood glucose is pathologically converted rapidly into sorbitol, leading directly to tragic diabetic cataracts and excruciating peripheral nerve damage.
Slide 9: The Glycosidic Bond Uniting Monosaccharides and Disaccharides
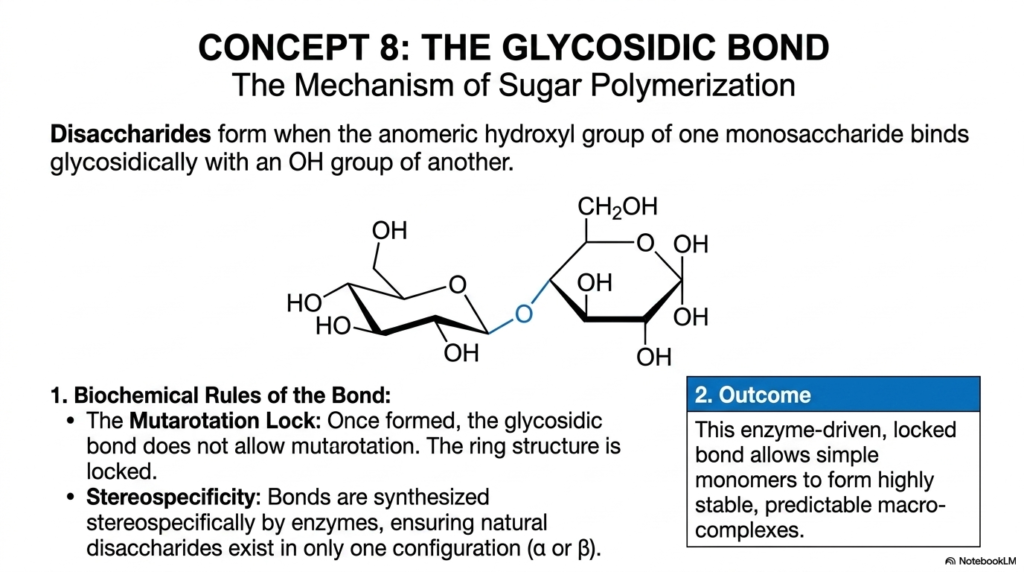
Concept 8 strictly and rigorously focuses on the single most functionally important chemical reaction in all of carbohydrate biochemistry: the enzymatic formation of the glycosidic bond. This specific, incredibly powerful mechanism is the absolute defining structural feature that literally separates simple Monosaccharides and Disaccharides from one another. Complex disaccharides structurally and functionally form when the highly reactive anomeric hydroxyl group of one individual monosaccharide strongly and permanently binds glycosidically with an -OH group of another sugar molecule. If you truly want to deeply and thoroughly understand the vast architecture of Monosaccharides and Disaccharides, you must deeply understand the intense intricacies of this precise enzymatic polymerization process.
There are two fundamental, absolutely unbreakable biochemical rules to this bond formation that you must aggressively master as medical and biochemistry students. First is the highly critical concept of the “Mutarotation Lock.” When a simple sugar is floating freely in an aqueous bodily solution, its anomeric carbon can spontaneously open and close, dynamically flipping back and forth between alpha and beta configurations.
However, once the strong glycosidic bond forms, the sugar ring is chemically locked in place at that specific anomeric carbon forever. Second is the incredibly important principle of absolute stereospecificity. The intricate cellular enzymes that actively synthesize these chemical bonds are fiercely specific, cleanly ensuring that natural disaccharides exist permanently in only one precise, highly functional configuration—either pure alpha or pure beta.
The ultimate biological and structural outcome of this intense, focused chemical reaction is massive, world-building structural stability. This enzyme-driven, tightly locked bond seamlessly allows simple, chaotic, floating monomers to efficiently organize into highly stable, predictably shaped macro-complexes. When rigorously studying the complex digestion of Monosaccharides and Disaccharides in human physiology, you will explicitly learn that the exact stereochemical geometry of the glycosidic bond strictly dictates exactly how the human body extracts energy. A simple microscopic difference between an alpha versus a beta linkage literally dictates what is nutritious food and what is completely undigestible dietary fiber!
Slide 10: Maltose and Starch Breakdown in Monosaccharides and Disaccharides
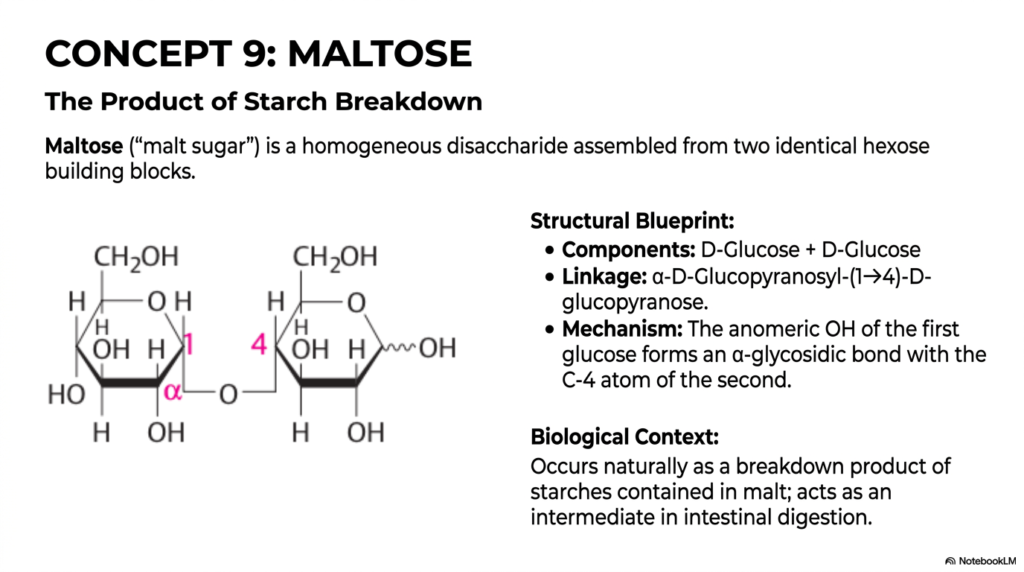
In Concept 9, we directly, practically, and elegantly apply our newly acquired knowledge of the glycosidic bond to Maltose, a classic, incredibly high-yield biological disaccharide. When closely and meticulously observing the natural, enzyme-driven digestion of Monosaccharides and Disaccharides within the human gastrointestinal tract, maltose consistently stands as a vital, absolutely inescapable intermediate biochemical step. Often referred to commonly in the global commercial food industry simply as “malt sugar,” maltose is a remarkably homogeneous, functionally straightforward disaccharide. It is cleanly assembled purely from two completely identical hexose building blocks. Among the major dietary Monosaccharides and Disaccharides we regularly consume, it is undeniably one of the absolute simplest to understand both structurally and mechanistically.
Let’s carefully analyze the clearly provided, highly detailed structural blueprint. Maltose is composed entirely of a single D-Glucose molecule perfectly and securely paired with another identical D-Glucose molecule. The exact chemical linkage securely connecting them is formally an α-D-Glucopyranosyl-(1→4)-D-glucopyranose bond. Mechanistically, this means that the tightly locked anomeric -OH group of the first glucose molecule forms a strict alpha-glycosidic bond with the C-4 atom of the second glucose molecule.
Because the bond is specifically formed in the alpha configuration, the structural oxygen bridge spatially points “down” relative to the strict orientation of the standard sugar ring. Importantly, the second glucose molecule permanently retains a free, unlocked anomeric carbon on its far right side, meaning it acts functionally as a highly reactive reducing sugar.
The true, undeniable biological context of maltose is incredibly important for clinical medical students. Maltose does not typically exist in large, free quantities in nature. Rather, it occurs naturally as a direct, highly specific enzymatic breakdown product of highly complex starches found in agricultural grains and seeds. In human digestive physiology, it acts as a highly critical, fleeting intermediate. When salivary or pancreatic amylase attacks a starchy potato, it creates manageable maltose, which is then cleanly cleaved at the intestinal brush border to release free, highly energetic glucose for your waiting cells.
Slide 11: Lactose and Mammalian Nutrition in Monosaccharides and Disaccharides
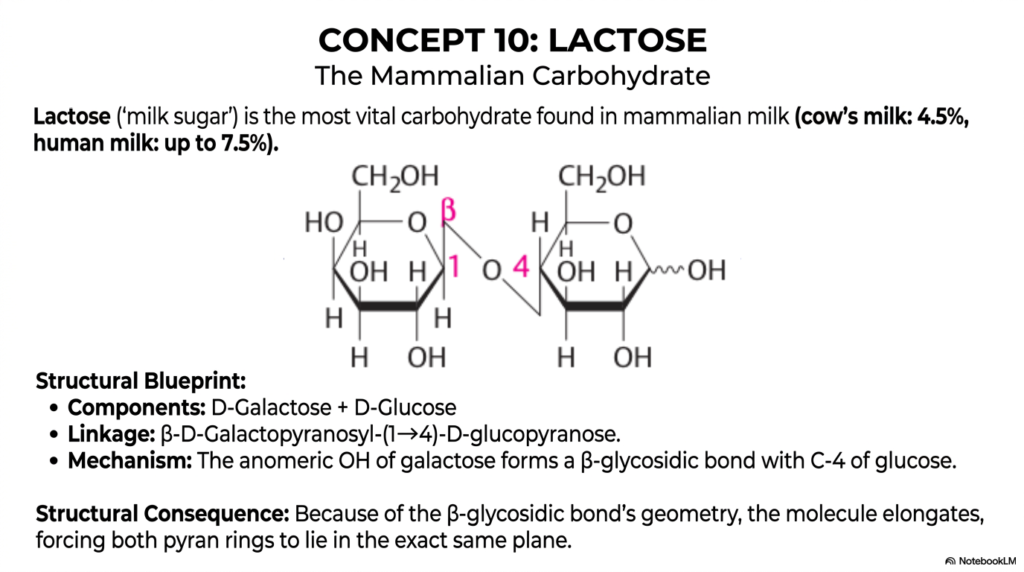
Advancing swiftly to Concept 10, we encounter Lactose, an absolutely fascinating, medically relevant case study in the highly diverse world of Monosaccharides and Disaccharides. Frequently referred to in clinical pediatrics and nutritional contexts as ‘milk sugar,’ lactose is the most vital, energy-dense carbohydrate naturally found in mammalian milk. It efficiently provides essential, easily accessible energy for rapidly developing infants across various mammalian species.
To give you some quantitative, real-world context beautifully shown on this slide, standard cow’s milk contains about 4.5% lactose, while human breast milk is exceptionally rich in this carbohydrate, containing an incredible up to 7.5%. For college and medical students alike, completely understanding how this specific sugar operates is a fundamental stepping stone to mastering the broader, highly complex chemistry of all Monosaccharides and Disaccharides.
Let’s carefully break down its elegant, highly specific structural blueprint so you can vividly see the molecular mechanics at play. Lactose is a highly specific disaccharide synthesized cleanly from two distinct monosaccharide components: D-galactose and D-glucose. The critical, highly specific enzymatic linkage reliably uniting these two carbohydrate rings is technically classified as a β-D-galactopyranosyl-(1→4)-D-glucopyranose bond. In plain, highly accessible English, the highly reactive anomeric hydroxyl group of the galactose molecule forms a precise, stereospecific beta-glycosidic bond directly with the fourth carbon of the glucose molecule. As we rigorously continue to structurally classify various Monosaccharides and Disaccharides, you will quickly notice a recurring, unbreakable rule: molecular stereochemistry always rigidly dictates physiological function.
The ultimate, inescapable structural consequence of this specific beta-glycosidic bond geometry is profound and chemically unyielding. The β(1→4) bond forces the overall molecule to elongate, thereby causing both bulky pyranose rings to lie rigidly in the same flat spatial plane. This highly rigid planar geometry is absolutely essential for the specific brush border enzyme lactase to properly recognize and cleave the molecule during normal human digestion. When adult patients lack this specific enzyme, the large, intact molecule wreaks absolute havoc in the colon, leading directly to the incredibly common, uncomfortable clinical presentation of lactose intolerance.
Slide 12: Sucrose and Plant Transport Forms of Monosaccharides and Disaccharides
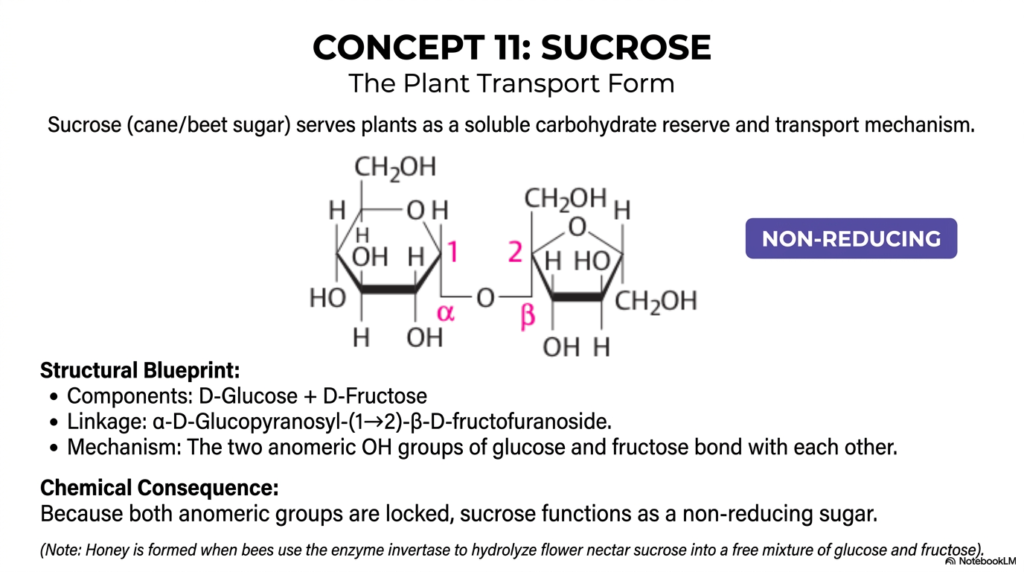
Concept 11 dramatically and importantly introduces Sucrose, arguably the most famous, heavily discussed, and widely consumed carbohydrate in the entire civilized world. In the broad, highly complex, and deeply nuanced study of Monosaccharides and Disaccharides, sucrose is incredibly chemically unique and biochemically fascinating. Common table sugar, largely derived commercially from massive quantities of sugar cane or sugar beet, serves wild plants as a highly soluble, incredibly dense carbohydrate reserve and as their primary, incredibly secure internal transport mechanism.
While complex animals aggressively use freely available, highly reactive blood glucose to rapidly transport energy throughout their dynamic bodies, plants rely heavily on the structurally stable, highly unreactive sucrose molecule, making it an absolutely essential subject within the extensive chemistry of Monosaccharides and Disaccharides.
The highly detailed structural blueprint of sucrose quickly reveals exactly why it is biochemically so utterly special. Its two core monosaccharide components are D-Glucose and D-Fructose. The specific, highly powerful linkage powerfully uniting them is an incredibly complex α-D-Glucopyranosyl-(1→2)-β-D-fructofuranoside double bond. Notice exactly how intricate and mathematically precise that specific chemical nomenclature sounds!
Mechanistically, the highly reactive two anomeric -OH groups of both the glucose and the fructose molecules bond directly and strongly with each other. The C-1 of the glucose binds flawlessly to the C-2 of the fructose. This specific, tightly double-locked connection has a massive chemical consequence: because both anomeric groups are firmly locked strictly inside the newly formed glycosidic bond, sucrose completely functions as a non-reducing sugar.
When scientifically and rigorously categorizing Monosaccharides and Disaccharides, this distinct non-reducing status is sucrose’s absolute, undeniable defining biochemical trait. It successfully prevents the massive molecule from spontaneously opening up and dangerously reacting with other vital cellular proteins or structures. This extreme, brilliant chemical stability allows plants to actively utilize it for highly safe, long-distance sap transport. As an incredibly engaging biological aside, delicious honey is magically formed exactly when foraging bees use a highly specific enzyme called invertase to safely hydrolyze this highly stable flower nectar sucrose into a free, chaotic, and incredibly sweet mixture of simple glucose and fructose.
Slide 13: Disaccharide Synthesis and the Matrix of Monosaccharides and Disaccharides

Concept 12 finally and brilliantly offers us a master comparative matrix, beautifully and highly efficiently summarizing the highly complex synthesis of these essential dietary sugars. As we thoughtfully and rigorously conclude our intensive, multi-slide exploration of Monosaccharides and Disaccharides, it is critical to momentarily step back and view these important, life-giving molecules side by side. This brilliant, cleanly formatted table provides a rapid-fire, extremely high-yield review of Maltose, Lactose, and Sucrose, structurally solidifying their highly unique physiological roles within the vast, complex family of Monosaccharides and Disaccharides. For highly dedicated medical or graduate students aggressively cramming for high-stakes, career-defining board exams, this single comprehensive summary slide is absolutely pure, unadulterated gold.
Let’s systematically and carefully review the matrix together to permanently lock in these critical biochemical concepts. Maltose, neatly composed of glucose and glucose, actively utilizes an α(1→4) glycosidic bond. Crucially, it remains a reactive reducing sugar solely due to its free anomeric carbon, serving as a vital biological intermediate in human intestinal starch digestion.
Lactose, gracefully uniting galactose and glucose, highly uniquely uses a β(1→4) bond. It also firmly remains a reducing sugar, and powerfully functions as the primary, highly energy-dense carbohydrate delicately formulated in mammalian milk. Finally, Sucrose, elegantly combining glucose and fructose, features a unique, tightly locked α(1→2)β double bond. As we just discussed, this entirely locks both anomeric carbons simultaneously, permanently rendering it completely non-reducing.
The overarching, highly critical synthesis insight proudly displayed at the absolute bottom of the slide brings all these highly complex biochemical concepts wonderfully together. While all three of these major, daily dietary Monosaccharides and Disaccharides are carefully assembled via enzymatically formed, highly stereospecific glycosidic bonds, Sucrose distinctly and purposefully stands entirely apart from the rest. By tightly binding both reactive anomeric carbons together simultaneously, it intentionally sacrifices its chemical reducing capability to magically become a highly stable, non-reactive chemical transport vehicle. This expertly crafted comparative matrix teaches us that, in the incredible world of Monosaccharides and Disaccharides, the ultimate biological destiny of any sugar is determined solely by its atomic bonds.
Please read our Content Disclaimer Statement.
Check out our social media channels: