45. Lipid Metabolism: Biochemical Pathways and Processes
Have you ever wondered how the human body seamlessly transitions from burning off a heavy meal to surviving overnight fasts without running out of energy? The secret lies in the elegant choreography of our cellular machinery. This slide deck explores the fundamental biochemistry governing how fats are synthesized, stored, and broken down to fuel the body. By mastering these pathways, medical and college students will unlock a deeper understanding of human bioenergetics and metabolic health.
Slide 1: Introduction to Lipid Metabolism Pathways
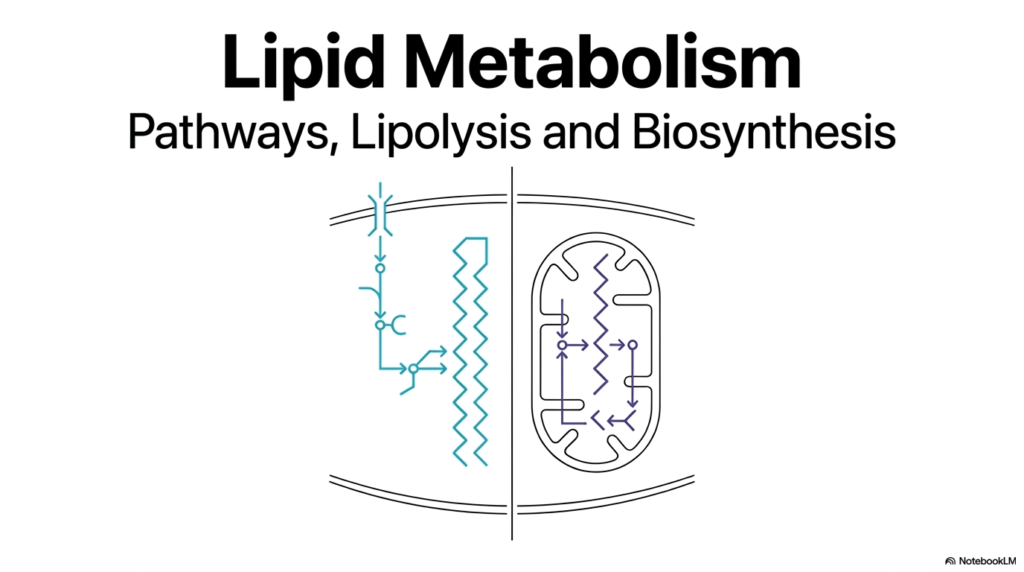
When embarking on the study of biochemistry, few topics are as dynamically complex and medically relevant as Lipid Metabolism. This introductory slide sets the stage for an overarching journey into how cells manage their most concentrated energy reserves. It visually contrasts the dual nature of these pathways by showcasing the cellular boundaries where these vital reactions occur. The left side of the graphic represents the cytoplasmic space, suggesting anabolic processes in which fatty acids are synthesized and elongated from smaller precursors.
Conversely, the right side of the illustration highlights the mitochondrion, the powerhouse of the cell, where the catabolic breakdown of fats takes place. Understanding the spatial separation of these pathways is the first critical step in mastering Lipid Metabolism. By compartmentalizing synthesis in the cytoplasm and degradation in the mitochondria, the cell prevents these opposing reactions from occurring simultaneously. This brilliant evolutionary design ensures metabolic efficiency and prevents a futile cycle of constant building and breaking.
The visual representation introduces students to the overarching themes of pathways, lipolysis, and biosynthesis. As one dives deeper into Lipid Metabolism, it becomes evident that these processes are not isolated events. They are part of a highly integrated system that responds continuously to the energy demands of the entire organism. This slide serves as a foundational roadmap for the detailed biochemical mechanisms that follow.
Whether the body is fasting or in a state of plenty, these pathways act as the ultimate regulators of energy homeostasis. Students must appreciate this big-picture view before zooming into the specific enzymatic reactions. This macroscopic perspective makes the intricate molecular details much easier to digest and retain.
Slide 2: Systemic Integration in Lipid Metabolism
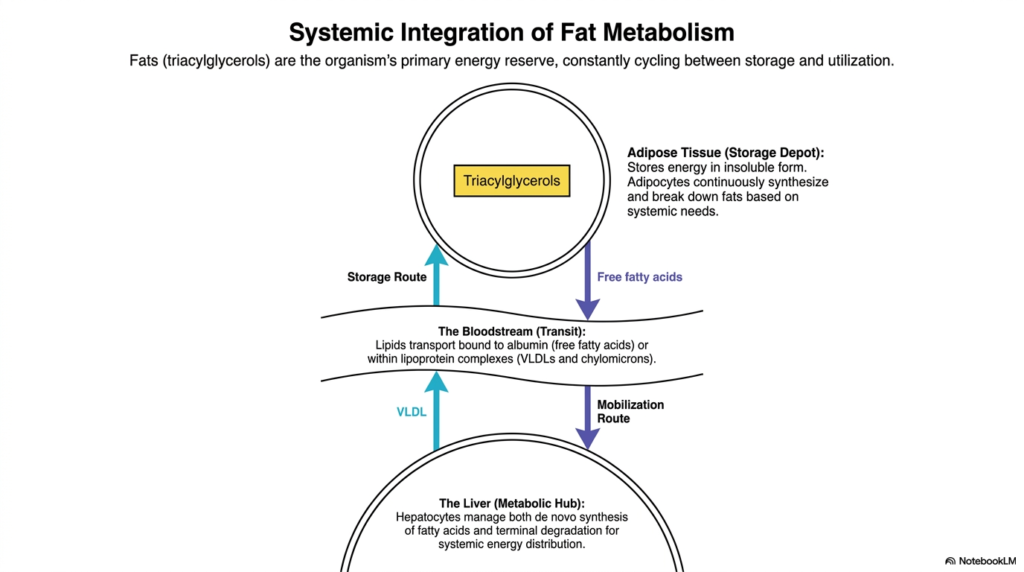
To truly understand Lipid Metabolism, one must first appreciate how fat is managed on a systemic, whole-body level. Fats, primarily in the form of triacylglycerols, serve as the organism’s premier energy reserve. This slide beautifully illustrates the continuous, dynamic cycling of these lipids between storage depots and the tissues that utilize them. Adipose tissue serves as the body’s primary storage depot, storing energy in an insoluble, highly concentrated form. Adipocytes are far from static; they are highly active cells that continuously synthesize and break down fats in response to the organism’s systemic energy needs.
The bloodstream serves as the grand transit highway for these lipid molecules. Because fats are inherently hydrophobic, they do not readily dissolve in the watery environment of blood plasma. Therefore, Lipid Metabolism relies on specialized transport mechanisms. Free fatty acids are shuttled securely when bound to the protein albumin, ensuring they reach their destination. Meanwhile, more complex lipids, such as triacylglycerols, are packaged into highly organized lipoprotein complexes, such as Very Low-Density Lipoproteins (VLDLs) and chylomicrons, which navigate the circulatory system to deliver their cargo.
At the center of this intricate web sits the liver, acting as the ultimate metabolic hub. Hepatocytes play a masterful role in Lipid Metabolism by managing both the de novo synthesis of fatty acids and their terminal degradation. When energy is abundant, the liver synthesizes fats and packages them into VLDLs for peripheral storage. During energy deficits, it orchestrates the distribution of fuel.
This systemic integration ensures that no tissue is left without the energy it requires to function. The delicate balance between the storage and mobilization routes highlights the exquisite regulatory control the body maintains over its lipid reserves, a concept central to both biochemistry and clinical medicine.
Slide 3: Adipocyte Lipogenesis and Lipid Metabolism
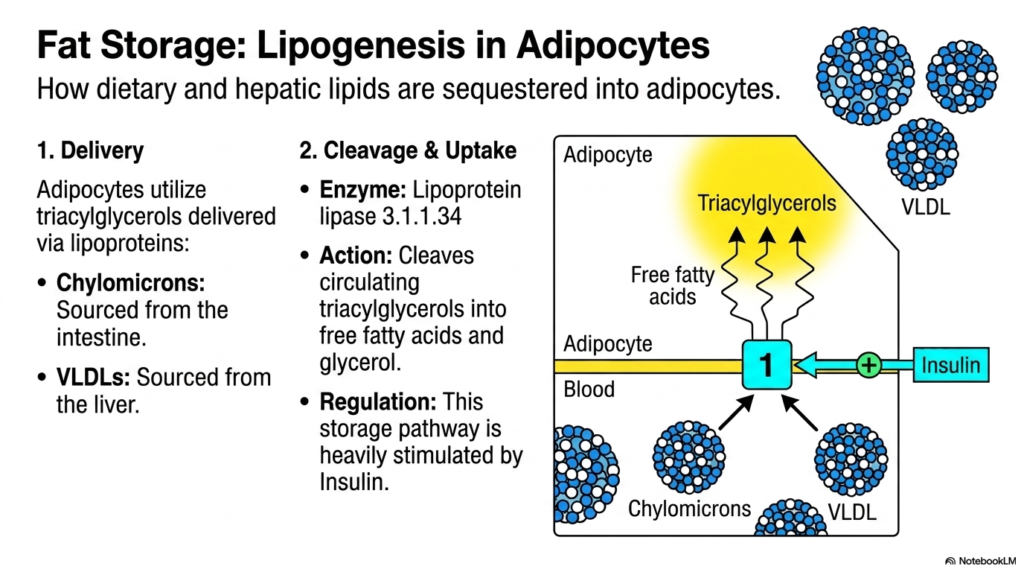
Focusing on the storage aspect of Lipid Metabolism, this slide details the precise mechanisms by which adipocytes sequester dietary and hepatic lipids. The process of lipogenesis in fat cells is a masterclass in cellular efficiency and nutrient capture. Triacylglycerols arrive at the adipose tissue via the bloodstream, packaged neatly in lipoproteins. Chylomicrons deliver lipids sourced directly from the intestine after a meal, while VLDLs transport fats that were synthesized in the liver. However, these bulky complexes cannot cross the adipocyte membrane directly.
This biological hurdle is overcome by a crucial enzyme known as Lipoprotein lipase 3.1.1.34. Positioned strategically at the endothelial surface of the blood vessels supplying adipose tissue, this enzyme orchestrates a vital step in Lipid Metabolism. It catalyzes the cleavage of the circulating triacylglycerols, breaking them down into free fatty acids and glycerol. This cleavage is absolutely essential, as only the free fatty acids are capable of being transported across the cell membrane and into the interior of the adipocyte for long-term storage.
Once inside the adipocyte, these free fatty acids are rapidly reassembled back into triacylglycerols, forming the massive lipid droplets characteristic of fat cells. The regulation of this entire storage pathway is a cornerstone of Lipid Metabolism and metabolic health. Insulin, the hormone of plenty, heavily stimulates this process.
When blood glucose is high, insulin signals the adipocytes to upregulate Lipoprotein lipase activity, thereby maximizing the extraction and storage of fats from the bloodstream. This insulin-driven sequestration lowers circulating lipid levels and ensures that excess energy is safely tucked away for future use, demonstrating the elegant coordination between carbohydrate abundance and fat storage.
Slide 4: Fat Mobilization in Lipid Metabolism
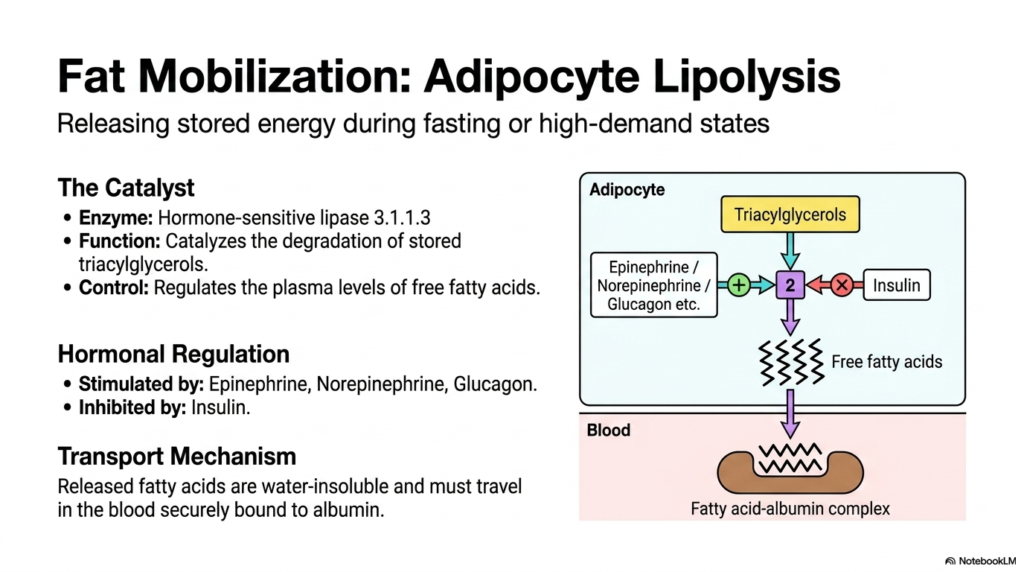
When the body enters a fasting state or faces high energy demands, Lipid Metabolism shifts dramatically from storage to mobilization. This slide unpacks the catabolic process of adipocyte lipolysis, where stored energy is rapidly liberated to fuel the organism. The primary catalyst for this energetic rescue mission is Hormone-sensitive lipase 3.1.1.3. Unlike the lipase responsible for fat storage, this intracellular enzyme acts directly on the massive lipid droplets within the adipocyte, catalyzing the degradation of stored triacylglycerols into their constituent free fatty acids and glycerol molecules.
The activity of Hormone-sensitive lipase is tightly controlled by systemic hormonal signals, ensuring that fat mobilization only occurs when absolutely necessary. This precise hormonal regulation is a fascinating aspect of Lipid Metabolism. The pathway is powerfully stimulated by catabolic hormones such as Epinephrine, Norepinephrine, and Glucagon. These chemical messengers alert the adipocyte to a systemic energy deficit, prompting the rapid breakdown of fat. Conversely, Insulin acts as a powerful inhibitor of this enzyme, effectively halting lipolysis when blood sugar levels are high and energy is abundant.
Once the triacylglycerols are cleaved, the newly freed fatty acids face a significant transport challenge. Because they are highly water-insoluble, they cannot freely diffuse through the aqueous environment of the bloodstream. To solve this, Lipid Metabolism relies on a dedicated carrier protein. The released fatty acids are immediately bound to albumin, a ubiquitous blood protein.
This fatty acid-albumin complex securely chaperones the hydrophobic lipids through the circulation, delivering them safely to energy-hungry tissues like skeletal muscle and the heart, where they will undergo oxidation to generate life-sustaining ATP.
Slide 5: Preparation for Oxidation in Lipid Metabolism
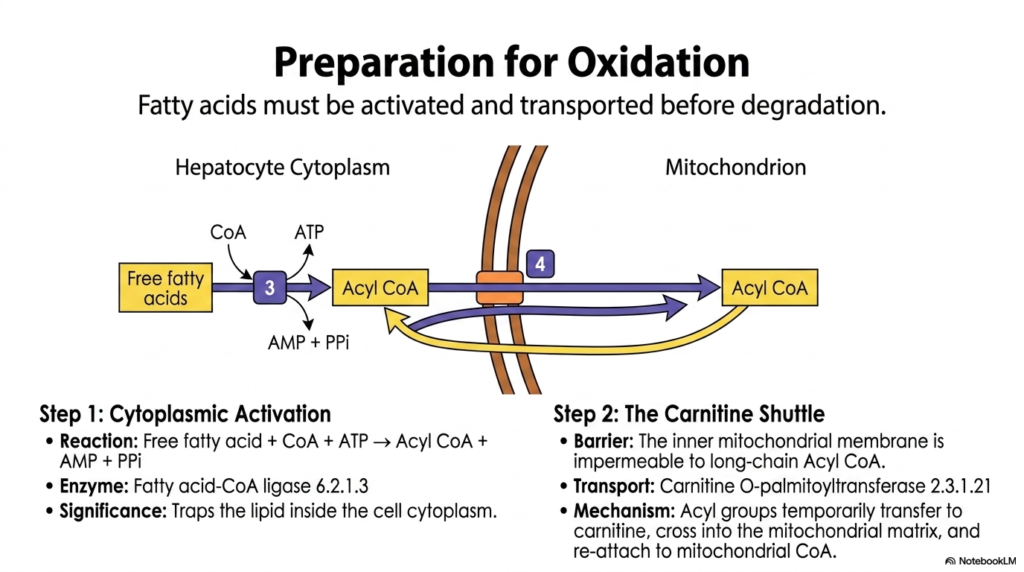
Before fatty acids can be burned for fuel, they must undergo a critical preparatory phase, a fundamental concept in Lipid Metabolism. This slide outlines the essential steps required to activate and transport fatty acids into the mitochondrial matrix, where they are degraded. The journey begins in the hepatocyte cytoplasm with Step 1: Cytoplasmic Activation. Here, a free fatty acid reacts with Coenzyme A (CoA) and ATP to form a high-energy molecule called Acyl CoA, releasing AMP and pyrophosphate (PPi) in the process.
This activation reaction is catalyzed by the enzyme Fatty acid-CoA ligase 6.2.1.3. The significance of this initial step in Lipid Metabolism cannot be overstated. By attaching the bulky CoA group, the cell effectively traps the lipid inside the cytoplasm, preparing it for the next phase of its metabolic journey. However, the newly formed Acyl CoA faces a formidable barrier: the inner mitochondrial membrane, which is strictly impermeable to long-chain Acyl CoA molecules. The fatty acid cannot reach the site of oxidation without specialized assistance.
This necessitates Step 2: The Carnitine Shuttle, an elegant transport mechanism central to Lipid Metabolism. The enzyme Carnitine O-palmitoyltransferase 2.3.1.21 facilitates this delicate transfer. It temporarily moves the acyl group from CoA onto a smaller carrier molecule called carnitine.
This newly formed acyl-carnitine complex can easily cross the inner mitochondrial membrane. Once inside the mitochondrial matrix, the acyl group is swiftly transferred back to a resident mitochondrial CoA molecule. Carnitine is then recycled back to the cytoplasm, ready to shuttle the next activated fatty acid into the mitochondria for energy production.
Slide 6: Beta-Oxidation Energy Harvesting in Lipid Metabolism
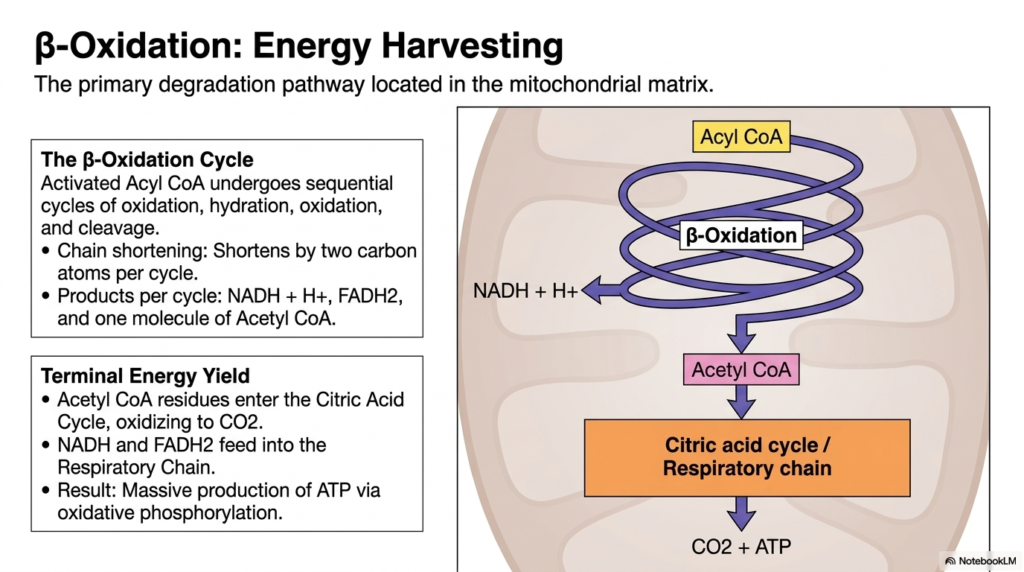
Once the activated fatty acid reaches the mitochondrial matrix, it undergoes the primary degradation pathway of Lipid Metabolism known as Beta-Oxidation. This slide breaks down the repetitive, high-yield energy-harvesting cycle that sustains the body during prolonged fasting. The Beta-Oxidation cycle is a sequential, four-step process involving oxidation, hydration, a second oxidation, and finally, cleavage. With each turn of this metabolic wheel, the activated Acyl CoA molecule is systematically chopped down, shortening the carbon chain by exactly two carbon atoms per cycle.
The biochemical products generated per cycle are incredibly valuable for cellular energy production. Every round of cleavage yields one molecule of NADH, one molecule of FADH2, and one molecule of acetyl-CoA. These products represent the true energetic payoff of Lipid Metabolism. The NADH and FADH2 molecules are high-energy electron carriers that immediately enter the mitochondrial Respiratory Chain, driving ATP production through oxidative phosphorylation. The sheer volume of reduced coenzymes produced here makes fat a vastly superior energy source compared to carbohydrates.
The terminal energy yield from Beta-Oxidation is staggering. The Acetyl-CoA residues generated by the cleavage steps do not go to waste; they enter the Citric Acid Cycle (Krebs cycle), where they are completely oxidized to carbon dioxide.
This secondary oxidation of Acetyl CoA generates even more NADH and FADH2, which again fuel the respiratory chain. Ultimately, the complete Beta-Oxidation of a single lipid molecule produces a large amount of ATP, providing the sustained energy required to keep vital organs functioning during times of nutritional scarcity.
Slide 7: Ketogenesis as the Overflow Valve in Lipid Metabolism
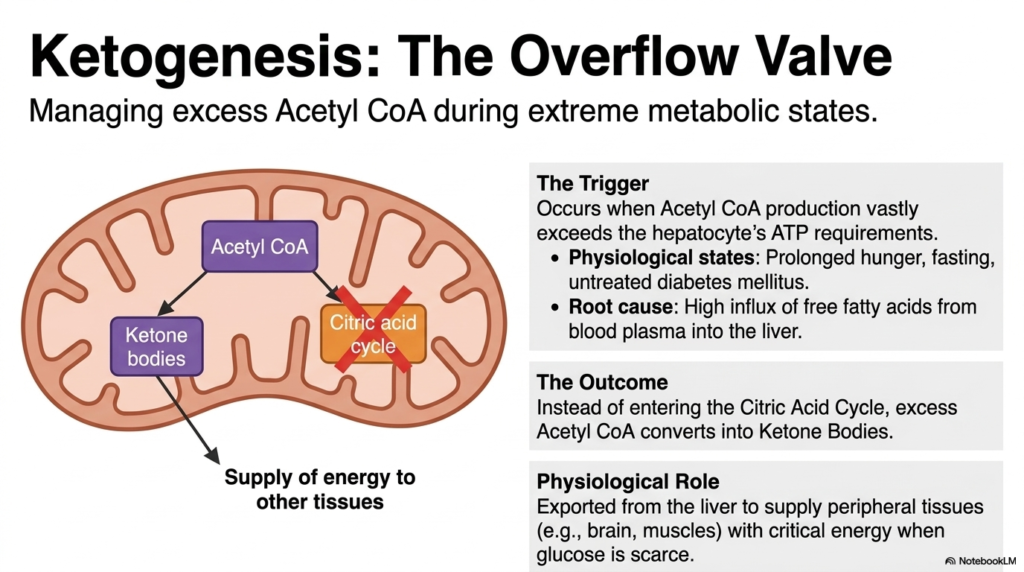
Ketogenesis represents a fascinating adaptive survival mechanism within Lipid Metabolism. This slide explains how the liver manages massive excesses of Acetyl CoA during extreme physiological stress. The trigger for ketogenesis occurs when the production of Acetyl-CoA from beta-oxidation far exceeds the hepatocyte’s immediate ATP requirements. This metabolic bottleneck typically arises during prolonged periods of hunger, extended fasting, or pathological conditions such as untreated diabetes mellitus. The root cause is a massive influx of free fatty acids mobilized from adipose tissue flooding into the liver.
Normally, Acetyl-CoA enters the Citric Acid Cycle for complete oxidation. However, in these extreme states, the cycle becomes overwhelmed, and intermediates are depleted for gluconeogenesis. Consequently, Lipid Metabolism employs an ingenious overflow valve. Instead of entering the stalled Citric Acid Cycle, the accumulating excess Acetyl CoA is diverted and enzymatically converted into Ketone Bodies within the liver mitochondria. This diversion prevents the toxic buildup of Acetyl CoA and allows the liver to continue beta-oxidation uninterrupted.
The outcome of this process is profoundly important for systemic survival. The newly synthesized ketone bodies are released from the liver into the bloodstream. Their physiological role in Lipid Metabolism is to supply peripheral tissues with a critical alternative fuel source.
Crucially, tissues like the brain and skeletal muscles, which heavily rely on glucose, can easily absorb and utilize these ketone bodies for energy. By exporting ketone bodies, the liver essentially packages the energy derived from fats into a water-soluble format that can keep the central nervous system functioning when circulating glucose is dangerously scarce.
Slide 8: De Novo Lipogenesis Origins in Lipid Metabolism
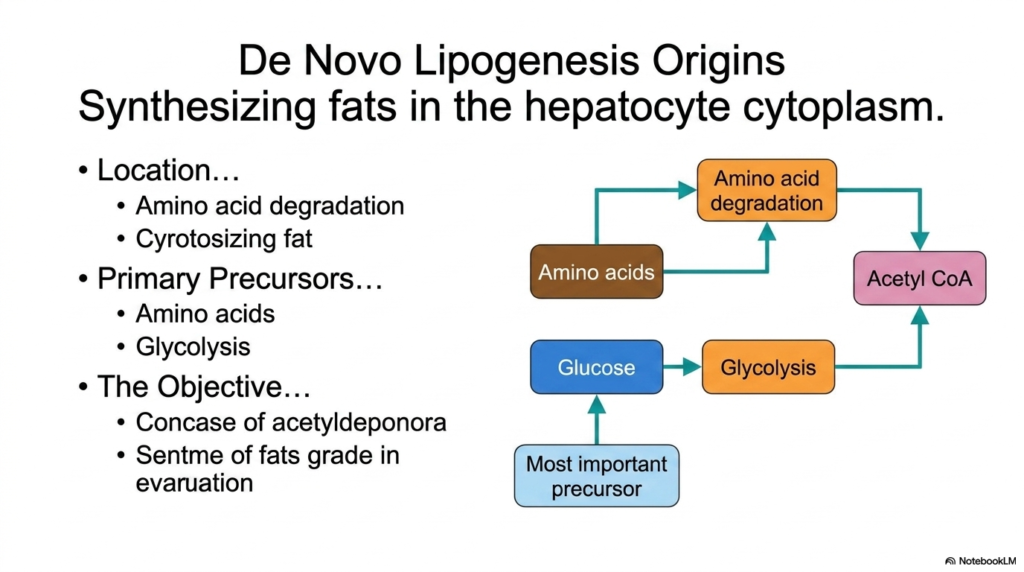
Shifting focus from degradation to creation, this slide introduces the origins of fat synthesis, a pathway essential to anabolic Lipid Metabolism. De Novo Lipogenesis is the biochemical process by which the body synthesizes brand new fatty acids from non-lipid precursors. This vital constructive process occurs in the hepatocyte cytoplasm. Unlike beta-oxidation, which occurs within the mitochondria to harvest energy, the cellular machinery requires a distinct spatial environment to build fats, ensuring that synthesis and degradation do not awkwardly interfere with one another.
The primary precursors for synthesizing these fats originate from the breakdown of other macronutrients. Through the degradation of amino acids and the process of glycolysis, the cell ultimately funnels carbon atoms into a common molecular currency: Acetyl CoA. Glucose acts as the most important precursor in this aspect of Lipid Metabolism. When carbohydrate intake exceeds the body’s immediate energy needs, glycolysis ramps up, producing an excess of Acetyl-CoA that cannot be fully consumed by the Citric Acid Cycle.
The objective of De Novo Lipogenesis is to capture this surplus energy. The cell effectively condenses these acetyl-CoA molecules, transforming the easily accessible energy of carbohydrates and amino acids into the dense, long-term storage format of lipids.
This pathway highlights the incredible metabolic flexibility of the human body. By converting excess dietary sugars and proteins into fat, the organism ensures that no valuable caloric energy is wasted. This foundational understanding of Lipid Metabolism is crucial for grasping how diets high in carbohydrates can ultimately lead to increased adiposity and fatty liver disease.
Slide 9: The Key Regulatory Enzyme in Lipid Metabolism
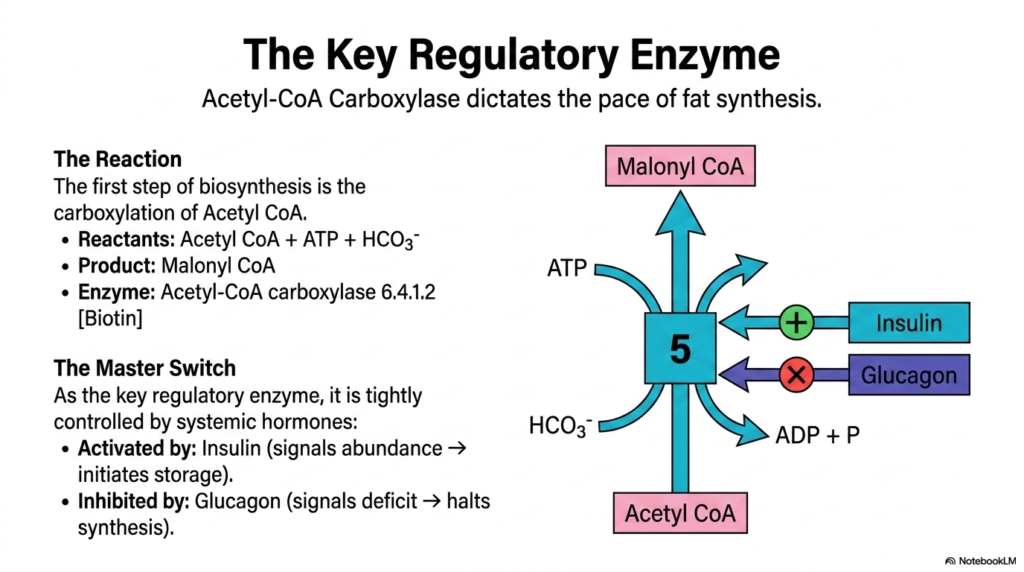
Every major biochemical pathway features a primary point of control, and in Lipid Metabolism, that critical checkpoint is managed by Acetyl-CoA Carboxylase. This slide highlights the key regulatory enzyme that dictates the overall pace of fat synthesis. The reaction catalyzed here is the very first committed step of fatty acid biosynthesis. The enzyme facilitates the carboxylation of Acetyl-CoA by adding a bicarbonate (HCO3-) group, using ATP-derived energy to produce Malonyl-CoA. This reaction also relies on the vital cofactor biotin to transfer the carbon dioxide group.
Because this is the rate-limiting step of fat synthesis, Acetyl-CoA Carboxylase acts as the master switch of anabolic Lipid Metabolism. Its activity is tightly governed by systemic hormonal signals that reflect the organism’s overall energy status. The enzyme is powerfully activated by Insulin, which serves as the classic signal of biochemical abundance. When you consume a meal and blood sugar rises, insulin binds to hepatocyte receptors, initiating a cascade that activates this enzyme, thereby kickstarting the production of Malonyl CoA to store the excess energy as fat.
Conversely, the pathway must be halted immediately during periods of energy deficit. Glucagon, the hormone of fasting, strongly inhibits Acetyl-CoA Carboxylase. This inhibition is a protective mechanism in Lipid Metabolism to prevent the cell from wasting precious ATP on building fats when the body desperately needs to mobilize stored energy instead.
This elegant push-and-pull regulation ensures that fatty acid biosynthesis only occurs when physiologically appropriate, seamlessly aligning cellular chemistry with the systemic nutritional state.
Slide 10: The Elongation Cycle in Lipid Metabolism
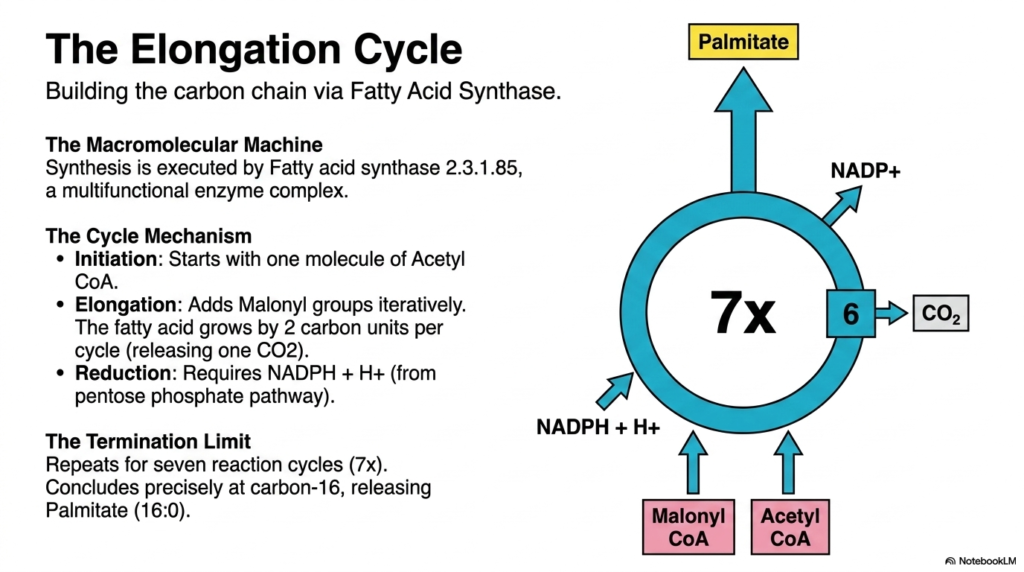
Once Malonyl-CoA is generated, the cell is ready to begin constructing the actual carbon chain, a fascinating mechanistic process in Lipid Metabolism. This slide details the Elongation Cycle, showcasing how molecular precursors are meticulously stitched together. The synthesis is executed by a massive, multifunctional enzyme complex known as Fatty acid synthase 2.3.1.85. This macromolecular machine is an incredible feat of biological engineering, containing all the necessary catalytic domains to repetitively add carbon units to the growing lipid chain without ever releasing the intermediate products.
The cycle mechanism is highly ordered and perfectly repetitive, exemplifying the efficiency of Lipid Metabolism. Initiation always starts with a single primer molecule of Acetyl CoA. From there, the elongation phase begins by iteratively adding two-carbon units derived from malonyl-CoA. During each addition, the previously attached carboxyl group is released as carbon dioxide, which thermodynamically drives the reaction forward. This constructive process requires substantial reducing power, supplied by NADPH + H+, generated primarily by the pentose phosphate pathway.
The reduction steps utilize the NADPH to remove oxygen and establish the fully saturated, highly reduced hydrocarbon tail characteristic of energy-dense lipids. This continuous loop is not indefinite; it is subject to a strict termination limit governed by the enzyme’s structure.
The Fatty acid synthase complex repeats these reaction steps exactly seven times. This precise enzymatic countdown concludes when the carbon chain reaches 16 carbons. At this exact point, a thioesterase domain cleaves the final product, releasing the fully formed Palmitate (16:0) into the cytoplasm.
Slide 11: Modification and Export in Lipid Metabolism
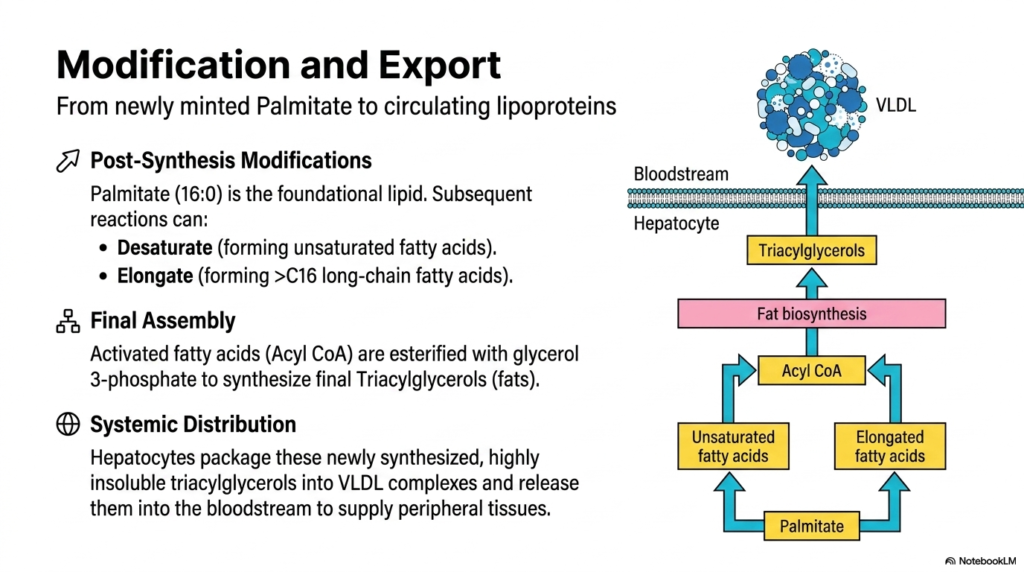
The release of Palmitate from the fatty acid synthase complex is not the end of the anabolic journey in Lipid Metabolism. This slide explores the crucial post-synthesis modifications and the ultimate export of these newly minted lipids. Palmitate (16:0) serves as the cell’s foundational lipid precursor. However, biological membranes and varied storage depots require a diverse array of fatty acids. Consequently, subsequent enzymatic reactions in the endoplasmic reticulum can modify Palmitate. Desaturase enzymes can introduce double bonds to form unsaturated fatty acids, while elongase enzymes can add further carbon units to create long-chain fatty acids greater than 16 carbons.
Once the desired array of fatty acids is created, they must undergo final assembly. A key rule of Lipid Metabolism is that free fatty acids are inherently toxic and detergent-like if left to accumulate in the cytoplasm. To neutralize them and prepare them for transport, these activated fatty acids (acyl-CoA) are sequentially esterified onto a glycerol 3-phosphate backbone. This multi-step esterification process ultimately synthesizes the final, biologically inert Triacylglycerols, commonly known as fats.
The final hurdle is systemic distribution. The liver cannot indefinitely store the massive quantities of fat it produces without suffering lipotoxicity. Therefore, a critical function of hepatic Lipid Metabolism involves packaging these highly insoluble newly synthesized triacylglycerols.
Hepatocytes meticulously wrap these lipids, along with cholesterol and apolipoproteins, to form Very Low-Density Lipoprotein (VLDL) complexes. These VLDLs are then secreted across the cell membrane and directly into the bloodstream, securely transporting the liver’s newly manufactured energetic bounty to peripheral tissues and adipose depots.
Slide 12: Diagnostic Comparison Matrix of Lipid Metabolism
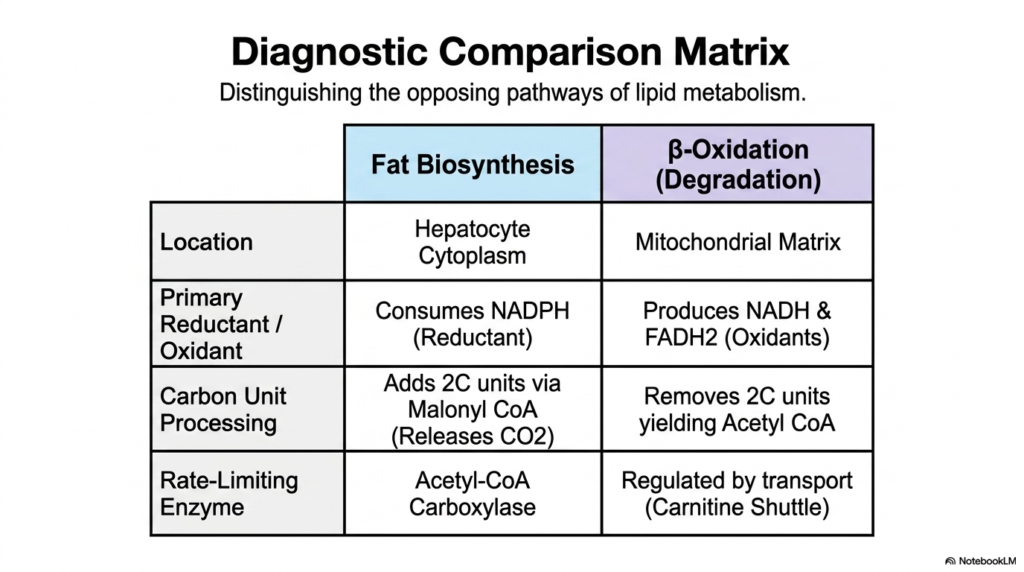
Mastering the intricate details of Lipid Metabolism requires distinguishing between its opposing constructive and destructive pathways. This slide provides a brilliant diagnostic comparison matrix that directly contrasts fat biosynthesis with beta-oxidation. By analyzing these pathways side by side, the elegant evolutionary logic that prevents their simultaneous operation becomes starkly apparent. First, the spatial location is strictly segregated: biosynthesis occurs exclusively in the hepatocyte cytoplasm, whereas degradation via beta-oxidation is confined to the mitochondrial matrix, ensuring physical separation of the opposing enzymes.
Furthermore, the chemical currency used by these pathways reveals a fundamental dichotomy in Lipid Metabolism. Fat biosynthesis is a highly reductive, anabolic process that consumes massive amounts of NADPH as its primary reductant. In stark contrast, beta-oxidation is a catabolic, oxidative process that continuously strips electrons from the lipid chain, producing the powerful oxidants NADH and FADH2 to fuel the respiratory chain. The handling of carbon units also mirrors one another: synthesis adds two-carbon units via Malonyl-CoA (releasing CO2), while degradation systematically removes two-carbon units, yielding Acetyl-CoA.
Finally, the matrix highlights the distinct rate-limiting bottlenecks that govern these twin pillars of Lipid Metabolism. The pace of biosynthesis is strictly dictated by the activity of the enzyme Acetyl-CoA Carboxylase.
Conversely, the rate of beta-oxidation is largely regulated by transport kinetics, specifically the activity of the Carnitine Shuttle, which dictates how quickly fatty acids can even enter the mitochondria. This comparative view solidifies a deep, functional understanding of how cellular lipid dynamics operate.
Slide 13: Regulatory Integration of Lipid Metabolism
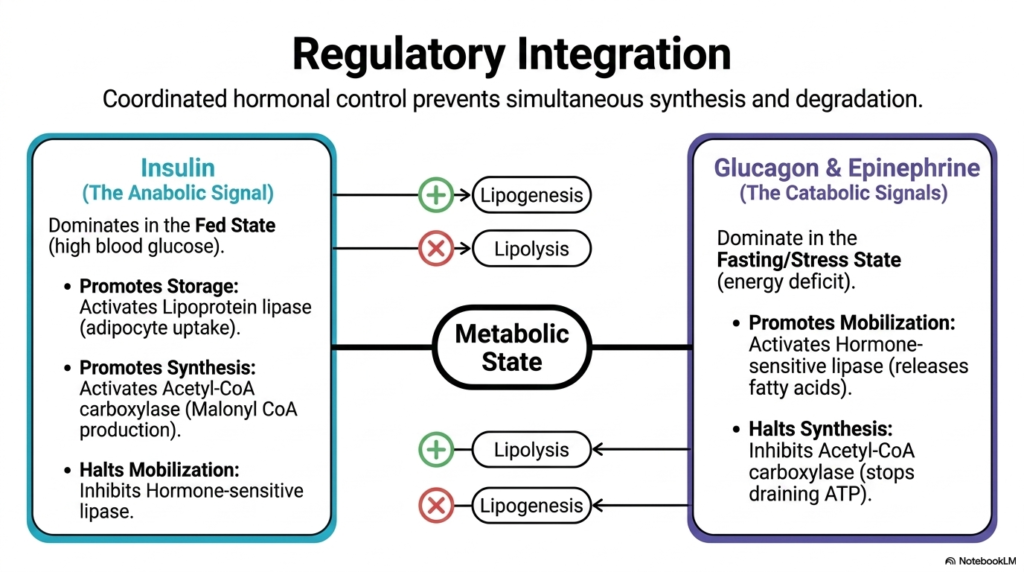
The final slide synthesizes the entire physiological orchestration of these biochemical processes, highlighting the overarching regulatory integration of Lipid Metabolism. Because synthesizing and degrading fats simultaneously would result in a massive, lethal waste of cellular ATP, the body utilizes coordinated hormonal control to strictly segregate these states. The organism’s metabolic state dictates the predominant hormonal signal, ensuring that anabolic and catabolic pathways act as opposing sides of a metabolic seesaw, never operating at maximum capacity simultaneously.
Insulin acts as the undisputed master of the fed state. When blood glucose is high, this potent anabolic signal dominates Lipid Metabolism. Insulin profoundly promotes storage by activating lipoprotein lipase to maximize adipocyte lipid uptake. Simultaneously, it promotes de novo synthesis by strongly activating Acetyl-CoA Carboxylase to produce Malonyl-CoA. Crucially, to prevent futile cycling, insulin also halts mobilization by strongly inhibiting hormone-sensitive lipase. This coordinated three-pronged attack ensures that times of caloric abundance result exclusively in the building and sequestration of fat reserves.
On the flip side, Glucagon and Epinephrine rule the fasting or stress states. When the body faces a severe energy deficit, these catabolic signals take command of Lipid Metabolism. They reverse the metabolic flow by heavily promoting mobilization, activating hormone-sensitive lipase to unleash a flood of free fatty acids from storage.
Concurrently, they halt synthesis by phosphorylating and inhibiting Acetyl-CoA Carboxylase, instantly stopping the drain on cellular ATP. This exquisite push-and-pull hormonal integration guarantees that the organism’s lipid machinery is always perfectly tuned to its immediate survival and energetic needs.
Please read our Content Disclaimer Statement.
Check out our social media channels: