17. Understanding Non-Proteinogenic Amino Acids and Their Roles
Medical school and college biochemistry curricula often emphasize a neat, tidy set of twenty molecular building blocks. However, the reality of cellular biology is far more complex and chemically diverse. The core purpose of this slide deck is to explore this hidden biochemical world, detailing the unique structures, origins, and vital physiological functions of molecules existing well outside the standard genetic code.
1. The Expanded Alphabet of Non-Proteinogenic Amino Acids: Beyond the Standard 20
Introductory biology courses emphasize the twenty standard building blocks of life, but this is only a fraction of the story. The true molecular landscape is vastly enriched by Non-Proteinogenic Amino Acids. These specialized molecules provide a level of structural diversity and metabolic complexity that standard protein building blocks alone cannot achieve.
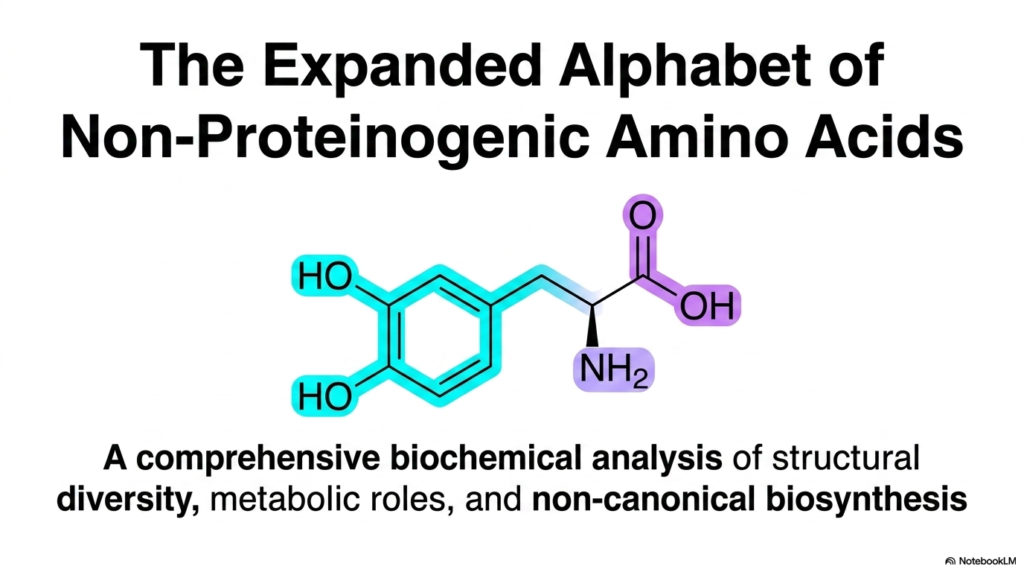
The cover slide features the chemical structure of L-DOPA, a prime example of Non-Proteinogenic Amino Acids. This specific molecule contains a distinct catecholamine ring and serves as an essential precursor to dopamine in the human brain. Its presence underscores that these non-standard molecules are indispensable to complex physiological processes, particularly in neurochemistry.
Understanding Non-Proteinogenic Amino Acids requires a deep dive into their non-canonical biosynthesis. Unlike their standard counterparts, these molecules are not assembled by ribosomes reading messenger RNA. Instead, they are synthesized through intricate enzymatic pathways tailored to specific cellular needs, bypassing the traditional central dogma of molecular biology entirely.
Throughout this presentation, the biochemical analysis will reveal how these unique structures function. From acting as isolated metabolic intermediates to serving as critical signaling molecules, they are the unsung heroes of cellular function. The exploration of these molecules expands the traditional biochemical alphabet, revealing a more nuanced view of life’s molecular toolkit.
The molecular architecture of these compounds often features fascinating chemical variations. While they maintain the fundamental functional groups—an amine and a carboxylic acid—their side chains can incorporate unique ring structures, altered oxidation states, or entirely different elemental compositions. This structural flexibility allows them to interact with enzymes and receptors in highly specific ways.
For medical students, grasping this expanded alphabet is crucial for understanding pharmacology and pathology. Many neurodegenerative diseases, metabolic disorders, and even therapeutic interventions revolve around these non-standard molecules. By mastering this diverse chemical language, future clinicians can better comprehend how physiological systems maintain homeostasis or fail during illness.
2. Defining Non-Proteinogenic Amino Acids: The Uncoded Majority in Biology
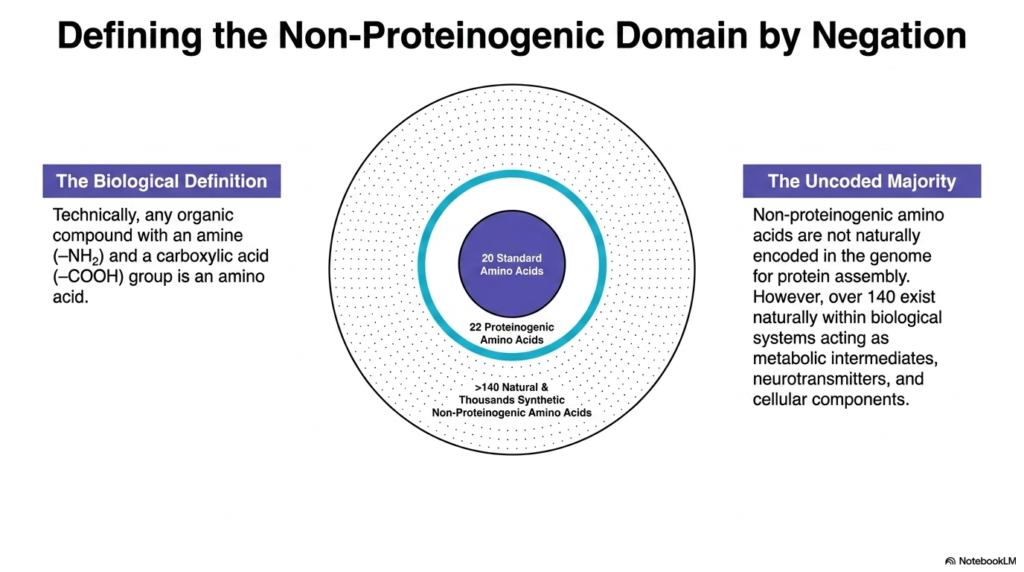
To truly grasp the scope of biochemistry, one must first define Non-Proteinogenic Amino Acids by negation. The biological definition of an amino acid is simply any organic compound possessing both an amine group and a carboxylic acid group. Yet the genome provides dedicated codons for only 20 standard amino acids, leaving a massive uncoded majority.
While twenty-two variants are technically proteinogenic—including rare additions like selenocysteine—the natural world contains over 140 distinct Non-Proteinogenic Amino Acids. Furthermore, synthetic chemistry has generated thousands more in the laboratory. This visual Venn diagram clearly illustrates that the standard genetic code represents merely a tiny central fraction of the possible chemical space available to living systems.
These abundant Non-Proteinogenic Amino Acids are not naturally encoded in the genome for direct incorporation into proteins. Instead of being strung together by ribosomes to form structural proteins or enzymes, they typically exist freely within the cytoplasm or extracellular fluid. Their uncoded status does not diminish their biological importance; rather, it highlights their specialized roles outside of translation.
Within cellular systems, this uncoded majority serves highly specific, dynamic purposes. Many act as vital metabolic intermediates, shuttling chemical groups between different enzymatic pathways. Others function as potent neurotransmitters, facilitating communication between neurons in complex nervous systems. These roles require rapid synthesis and degradation, making them unsuitable for permanent incorporation into stable protein structures.
Medical science continuously discovers new physiological functions for these molecules. Because they are not restricted by the standard genetic code, they offer a vast reservoir of structural diversity. This diversity allows evolutionary processes to utilize them for precise regulatory mechanisms, cellular defense, and complex signaling networks across all domains of life.
Understanding this vast domain helps biochemistry students conceptualize the cell not just as a protein factory, but as a dynamic chemical reactor. The sheer volume of these non-standard molecules proves that life relies on a much broader chemical vocabulary than the DNA sequence alone would suggest.
3. Prebiotic Origins and Non-Proteinogenic Amino Acids: The Meteoritic Alphabet
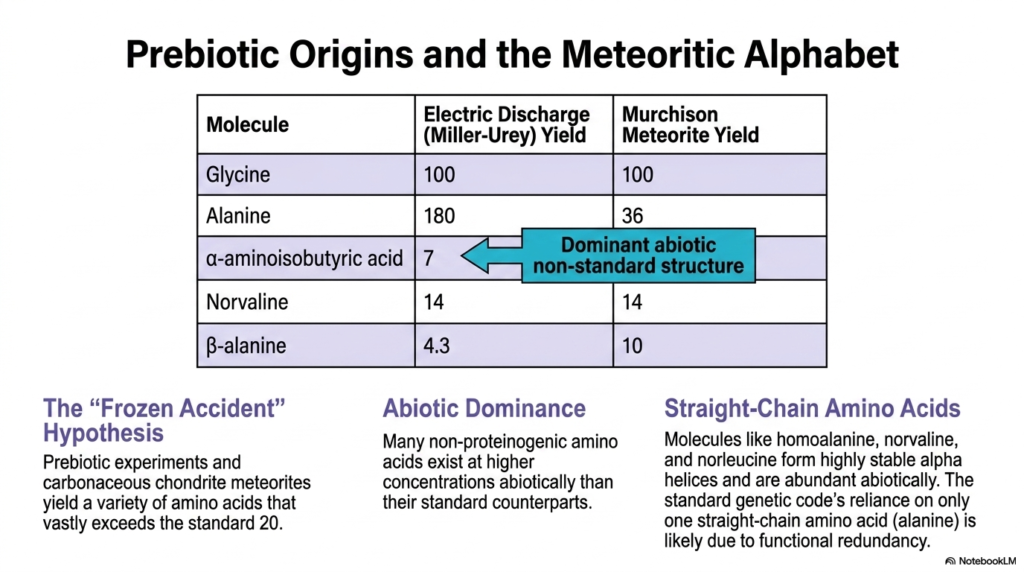
The origins of life on Earth offer profound insights into the nature of Non-Proteinogenic Amino Acids. When examining prebiotic chemistry, researchers look to both laboratory simulations and extraterrestrial samples. Famous experiments, such as the electric discharge studies by Miller and Urey, successfully synthesized organic compounds from primitive atmospheric gases, revealing a surprising diversity of molecules.
Analysis of carbonaceous chondrite meteorites, like the Murchison meteorite, confirms these experimental findings. These ancient space rocks contain a vast array of Non-Proteinogenic Amino Acids, proving that these structures form readily under abiotic conditions. Interestingly, many of these non-standard molecules exist at much higher concentrations in space than the standard twenty found in terrestrial biology.
A key example highlighted in this slide is alpha-aminoisobutyric acid, which represents a dominant abiotic non-standard structure. Along with norvaline and beta-alanine, these Non-Proteinogenic Amino Acids represent a “meteoritic alphabet” of chemical possibilities. Their abundance suggests that Earth’s early chemical soup was teeming with molecular variants that were eventually excluded from the universal genetic code.
This brings up the fascinating “Frozen Accident” hypothesis. If prebiotic environments yielded a variety of amino acids vastly exceeding the standard twenty, why did life settle on its current molecular toolkit? The hypothesis suggests that the standard code was largely a chance event that became locked in place once translation machinery evolved, freezing out the abiotic alternatives.
Furthermore, the dominance of straight-chain amino acids in abiotic environments is notable. Molecules like homoalanine, norvaline, and norleucine easily form highly stable alpha helices. Yet, the standard genetic code relies on only one straight-chain variant: alanine. This is likely due to functional redundancy; life did not need multiple straight-chain variants when one could perform the necessary structural roles perfectly well.
For students studying evolutionary biochemistry, these prebiotic insights are crucial. They demonstrate that the building blocks of life are not universally unique, but rather a selectively curated subset of a much larger chemical universe. The exclusion of certain molecules sheds light on the selective pressures that shaped early cellular life.
4. Shifting the Amine Group: Structural Variations in Non-Proteinogenic Amino Acids
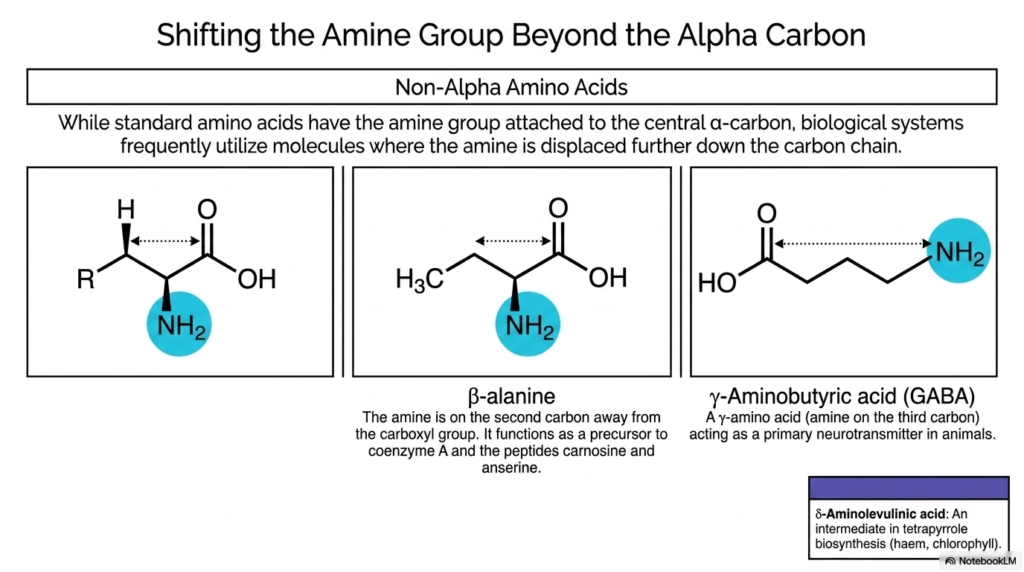
Standard biological building blocks are almost exclusively alpha-amino acids, meaning the primary amine group is attached directly to the central alpha-carbon. However, Non-Proteinogenic Amino Acids frequently break this structural rule. Biological systems expertly utilize non-alpha molecules in which the functional amine is displaced farther along the carbon chain, fundamentally altering the molecule’s biochemical properties.
By shifting the amine group away from the alpha carbon, Non-Proteinogenic Amino Acids lose the ability to form standard peptide bonds within a ribosomal context. Instead of forming rigid protein backbones, these shifted molecules serve specialized, standalone functions. A prominent example is beta-alanine, in which the amine is on the second carbon away from the carboxyl group.
Beta-alanine is a critical non-proteinogenic amino acid variant that functions as an essential precursor. It is required for the biosynthesis of coenzyme A, a fundamental molecule in lipid metabolism and the Krebs cycle. Additionally, it combines with histidine to form the dipeptide carnosine, which acts as a powerful buffer against lactic acid accumulation in muscle tissues.
Moving further down the chain yields gamma-amino acid variants, such as gamma-aminobutyric acid (GABA). In GABA, the amine is located on the third carbon from the carboxyl group. This structural shift transforms the molecule into the primary inhibitory neurotransmitter in the mammalian central nervous system, dictating neuronal excitability and preventing runaway electrical signaling in the brain.
Another crucial example is delta-aminolevulinic acid, featuring an even larger shift. This molecule acts as an indispensable intermediate in tetrapyrrole biosynthesis. Without it, cells could not produce haem for hemoglobin or chlorophyll for photosynthesis, demonstrating how variations in basic carbon backbone structure lead to profoundly different and life-sustaining biological functions.
For medical students, recognizing these structural shifts is vital for understanding pharmacology. Many synthetic drugs are designed to mimic these shifted structures to target specific receptors in the nervous or metabolic systems. Recognizing non-alpha structures is a foundational skill for advanced biochemical analysis.
5. The D-Enantiomers: Non-Proteinogenic Amino Acids in Bacterial Cell Walls
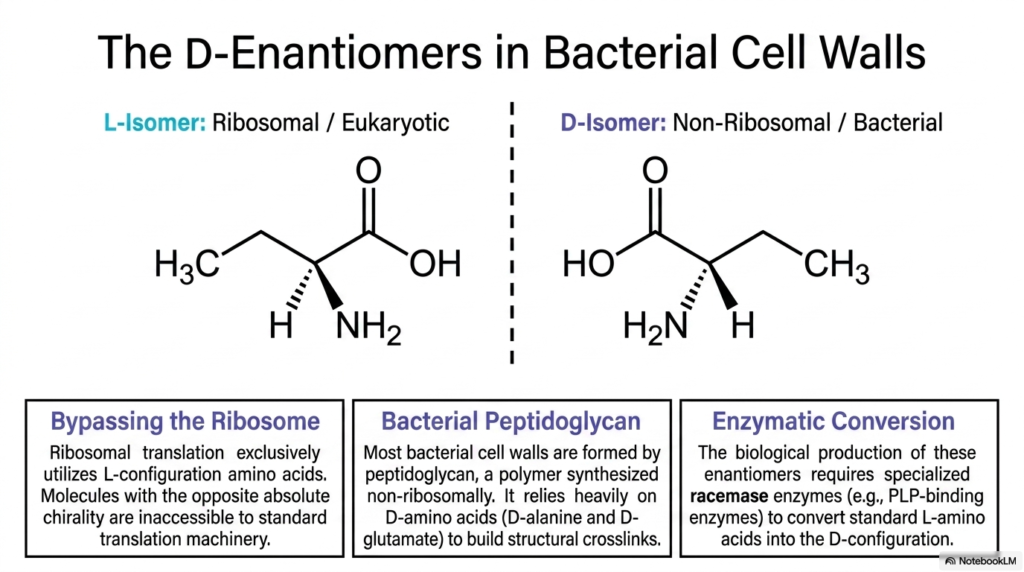
Chirality is a fundamental concept in biochemistry, dictating the three-dimensional geometry of life. Ribosomal translation exclusively utilizes L-configuration isomers to build eukaryotic and prokaryotic proteins. Therefore, their mirror images, the D-isomers, are firmly classified as Non-Proteinogenic Amino Acids because they are completely inaccessible to standard translation machinery and cannot be integrated into normal cellular proteins.
Despite bypassing the ribosome, these D-enantiomer Non-Proteinogenic Amino Acids play a significant role in microbiology, particularly in bacterial cell walls. Bacteria rely heavily on peptidoglycan, a highly durable polymer synthesized non-ribosomally, to maintain structural integrity and resist osmotic pressure. This rigid matrix is extensively crosslinked using specialized D-isomers, providing a unique evolutionary advantage.
The integration of D-isomers, such as D-alanine and D-glutamate, into the peptidoglycan layer protects bacteria from environmental threats. Because most host defense mechanisms, like digestive proteases, are stereospecific for L-isomers, they cannot easily cleave the bonds formed by these Non-Proteinogenic Amino Acids. This structural cleverness makes bacterial cell walls highly resistant to enzymatic degradation by eukaryotic hosts.
Since these molecules cannot be obtained through standard translation, bacteria must synthesize them independently. The biological production of these enantiomers requires specialized enzymes known as racemases. These PLP-binding enzymes actively convert standard L-amino acids into the necessary D-configuration, ensuring a steady supply of structural materials for cell wall biosynthesis and division.
Understanding this stereochemical distinction is paramount for pharmacology and infectious disease management. Many of our most effective antibiotics, including penicillins and cephalosporins, specifically target the enzymes that crosslink these D-isomers. By inhibiting peptidoglycan synthesis, these drugs cause the bacterial cell wall to fail, leading to rapid cell death without harming the host’s L-isomer-based proteins.
Ultimately, the existence of biological D-enantiomers proves that molecular handedness is not just a chemical curiosity, but a crucial determinant of biological function and survival. Medical school curricula place heavy emphasis on this concept when discussing microbial pathogenesis and targeted antimicrobial therapies.
6. Structural Distortion: Incorporating Non-Proteinogenic Amino Acids
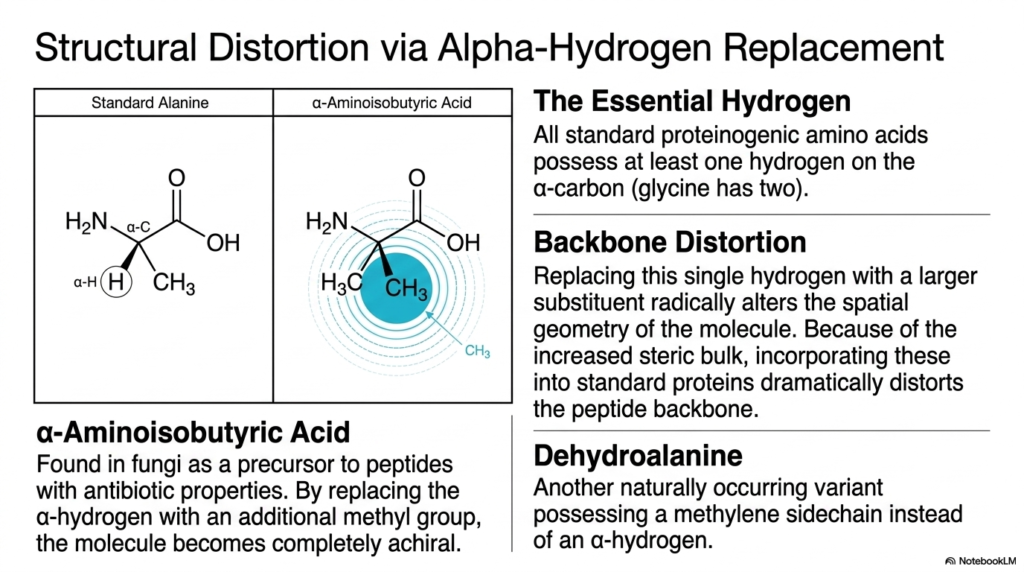
All standard proteinogenic molecules possess at least one vital hydrogen atom on their alpha-carbon, with glycine possessing two. This seemingly minor structural detail is critical for the flexibility and standard folding of proteins. When researchers examine Non-Proteinogenic Amino Acids, they often find fascinating variations where this essential hydrogen is entirely replaced by a larger chemical substituent.
Replacing this single alpha-hydrogen with a bulkier group radically alters the spatial geometry of the molecule. Because of their increased steric bulk, incorporating these Non-Proteinogenic Amino Acids into standard proteins dramatically distorts the delicate peptide backbone. The physical space required by the new substituent forces the adjacent chemical bonds into highly strained, unnatural angles.
A textbook example of this phenomenon is alpha-aminoisobutyric acid. In this specific Non-Proteinogenic Amino Acids variant, the standard alpha-hydrogen of alanine is replaced by an additional methyl group. This simple substitution makes the molecule completely achiral and highly rigid. It is frequently found in fungi and acts as a precursor to specialized peptides with potent antibiotic properties.
The rigidity introduced by these bulky alpha-substituents prevents the formation of standard alpha-helices or beta-sheets. Instead, fungal peptides incorporating these molecules form unique, highly stable structures like 3_10 helices. These distorted structures are highly resistant to degradation by external proteases, giving the fungi a distinct survival advantage in competitive microbial environments.
Another naturally occurring variant mentioned is dehydroalanine. Instead of a standard alpha-hydrogen and a traditional sidechain, dehydroalanine possesses a reactive methylene sidechain double-bonded directly to the alpha-carbon. This unique unsaturated structure makes it a highly reactive electrophile, allowing it to participate in specialized covalent crosslinking within bacterial peptides and certain eukaryotic proteins.
For biochemistry students, these examples highlight the ribosome’s absolute precision and the standard genetic code. The standard twenty building blocks were likely selected during evolution precisely because their steric profiles allow for the versatile, dynamic folding required by complex cellular enzymes, whereas these distorted variants serve niche, hyper-stable roles.
7. Twin Stereocenters: Non-Proteinogenic Amino Acids as Complex Structural Bridges
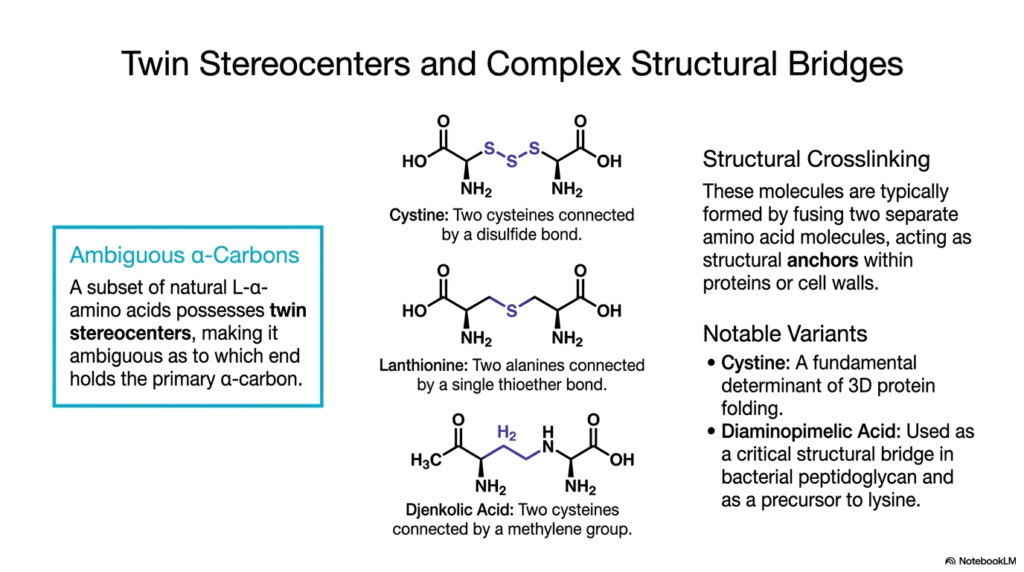
In advanced molecular biology, structural stability is paramount. Certain specialized molecules act as complex structural bridges, fundamentally relying on the presence of twin stereocenters. These particular Non-Proteinogenic Amino Acids are typically formed by fusing two precursor molecules, creating a larger, more complex compound that serves as a robust structural anchor in proteins or cell walls.
A defining hallmark of these structural anchors is the presence of ambiguous alpha-carbons. Because these Non-Proteinogenic Amino Acids are essentially dimers formed from two traditional building blocks, they possess two chiral centers. This dual-center nature makes it chemically ambiguous which end bears the primary alpha-carbon, fundamentally distinguishing it from standard monomeric building blocks.
While cystine is technically formed from standard cysteine residues, its oxidized dimeric form acts much like Non-Proteinogenic Amino Acids in terms of generating higher-order structural rigidity. Connected by a covalent disulfide bond, cystine acts as a fundamental determinant of three-dimensional protein folding, locking complex enzymes and antibodies into their active, functional conformations.
Lanthionine provides a similar bridging function but utilizes a single thioether bond to connect two alanine residues. This unique linkage is commonly found in bacteriocins, which are antimicrobial peptides produced by bacteria. The thioether bridge provides immense stability, allowing these toxic peptides to survive harsh environmental conditions while hunting competing bacterial strains.
Another notable variant is djenkolic acid, where two cysteines are firmly connected by a methylene group. Found naturally in certain Southeast Asian beans, this structural bridge can cause severe toxicity in humans by forming sharp crystals in the urinary tract. This highlights how complex structural bridges can have profound physiological and pathological impacts.
Finally, diaminopimelic acid serves as a critical structural bridge in bacterial peptidoglycan. As a vital precursor to lysine, it has twin stereocenters that enable it to crosslink bacterial cell walls powerfully. Understanding these complex bridges is essential for medical students studying bacterial architecture and the mechanisms of advanced cell-wall targeting antibiotics.
8. Genetically Encoded Exceptions: The Gray Area of Non-Proteinogenic Amino Acids
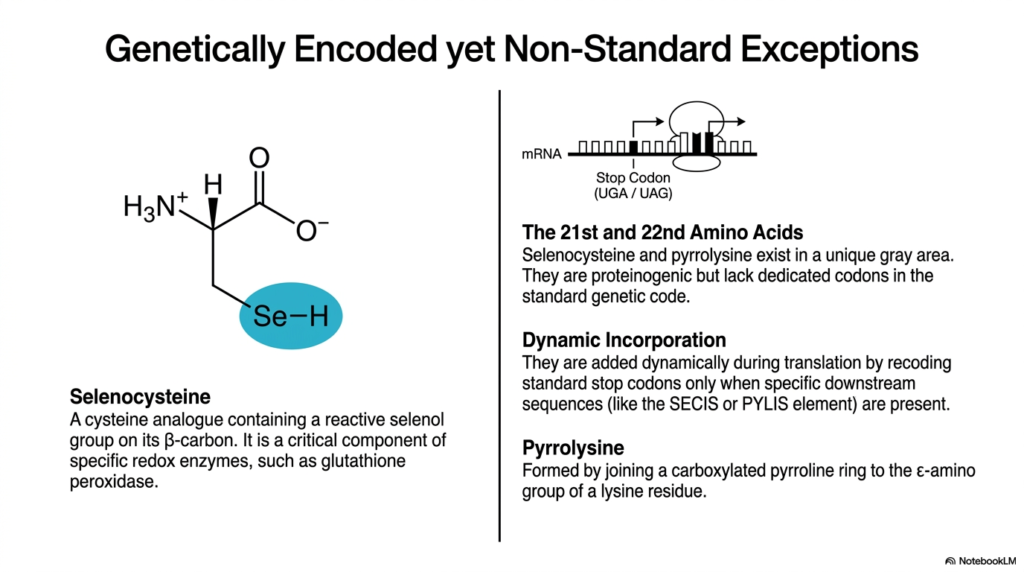
Biochemistry is rarely black-and-white, and the genetic code contains fascinating exceptions. Selenocysteine and pyrrolysine exist in a unique gray area of molecular biology. While they are often discussed alongside Non-Proteinogenic Amino Acids due to their rarity and specialized functions, they are technically proteinogenic. However, they completely lack dedicated, standard codons in the universal genetic code.
Because they lack standard codons, these molecules rely on complex dynamic incorporation mechanisms. They are not assigned to any of the 64 traditional triplet codons. Instead, they behave like Non-Proteinogenic Amino Acids that have somehow hacked the translation machinery. They are added dynamically during active translation by selectively recoding standard stop codons, typically UGA or UAG.
This recoding process only occurs when specific downstream messenger RNA sequences are present. For selenocysteine, a SECIS element forces the ribosome to read a UGA stop codon as an instruction to incorporate the molecule. Without this secondary RNA structure, the ribosome would simply terminate translation. This complex workaround blurs the line between standard building blocks and Non-Proteinogenic Amino Acids.
Selenocysteine is an analog containing a highly reactive selenol group on its beta-carbon. This intense reactivity makes it a critical component of specific redox enzymes, such as glutathione peroxidase, that protect cells from oxidative damage. Its unique chemical properties enable these specialized enzymes to perform catalytic functions that standard sulfur-containing cysteine cannot efficiently achieve.
Pyrrolysine, considered the 22nd amino acid, is equally fascinating. It is formed by joining a carboxylated pyrroline ring directly to the epsilon-amino group of a lysine residue. Found primarily in methanogenic archaea, it is dynamically incorporated via the PYLIS element. This specialized structure is essential for the enzymes that generate methane gas in extreme environments.
For students, these exceptions prove that the genetic code is not entirely frozen. Evolution has found intricate ways to expand the cellular toolkit beyond the standard twenty. Understanding these recoding mechanisms opens the door to advanced applications in bioengineering and synthetic biology.
9. Core Metabolic Intermediates: Non-Proteinogenic Amino Acids in the Urea Cycle
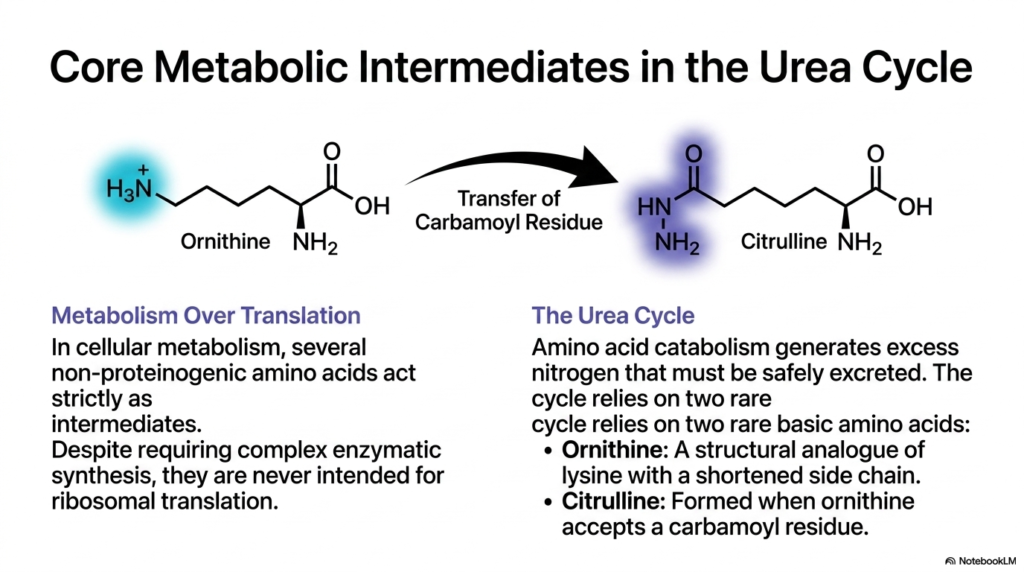
Cellular metabolism prioritizes chemical utility over ribosomal translation. In critical metabolic pathways, several Non-Proteinogenic Amino Acids act strictly as specialized chemical intermediates. Despite requiring complex enzymatic synthesis and significant cellular energy to produce, they are never intended to be strung together by the ribosome into proteins; they serve entirely different physiological purposes.
A prime example of this “metabolism over translation” paradigm is found in the liver. Amino acid catabolism generates toxic excess nitrogen in the form of ammonia that must be safely neutralized and excreted. The hepatic urea cycle relies entirely on rare basic Non-Proteinogenic Amino Acids to shuttle this nitrogen out of the biological system safely and efficiently.
The two most prominent Non-Proteinogenic Amino Acids driving this cycle are ornithine and citrulline. Ornithine acts as a structural analog of lysine but features a shortened aliphatic side chain. It serves as the primary acceptor molecule in the cycle, binding toxic nitrogen compounds within the mitochondria to initiate detoxification.
Once ornithine accepts a carbamoyl group, it is enzymatically converted to citrulline. This biochemical conversion effectively converts toxic ammonia into a safe, water-soluble form. Citrulline is then exported from the mitochondria into the cytosol, where it undergoes further enzymatic reactions to eventually yield urea, which is safely excreted by the kidneys.
Medical students must thoroughly understand these specific intermediates because enzymatic deficiencies within this cycle lead to severe hyperammonemia. If ornithine or citrulline cannot be properly processed, toxic nitrogen rapidly accumulates in the bloodstream, leading to catastrophic neurological damage. Understanding these non-standard molecules is literally a matter of life and death in clinical metabolic genetics.
By acting exclusively as metabolic shuttles, ornithine and citrulline demonstrate the vast chemical utility of molecules outside the genetic code. Their specialized structures enable them to participate in rapid, cyclical chemical reactions that rigid structural proteins cannot carry out.
10. Biosynthesizing Biogenic Amines from Non-Proteinogenic Amino Acids
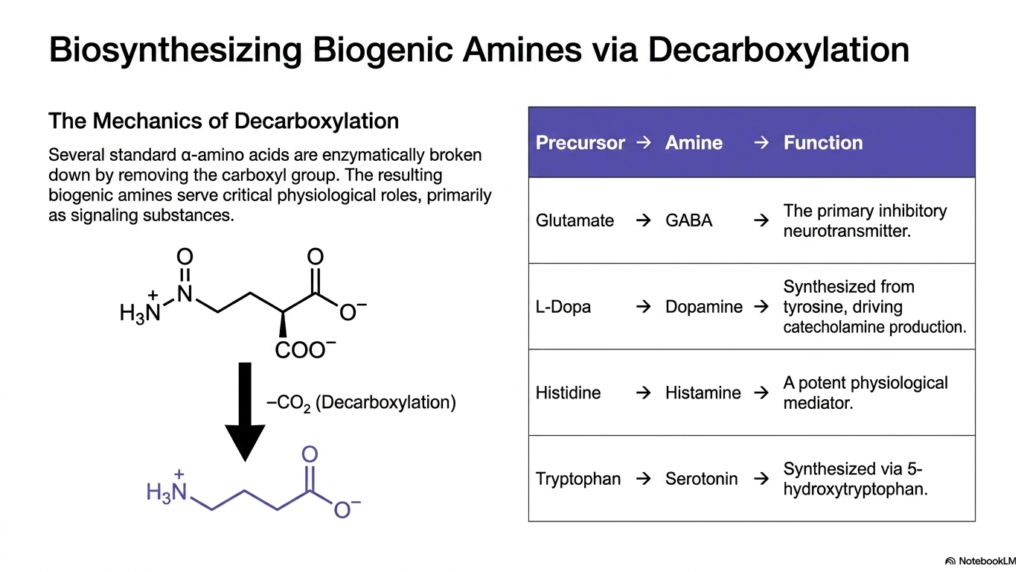
Chemical communication between cells requires rapidly synthesized, highly active signaling molecules. Many of these crucial messengers are formed by converting standard building blocks into potent Non-Proteinogenic Amino Acids. This transformation is typically achieved through a specific enzymatic mechanism known as decarboxylation, which fundamentally alters both the structure and biological function of the precursor molecule.
The mechanics of decarboxylation involve specialized enzymes breaking down standard alpha-amino acids by entirely removing their carboxyl group as carbon dioxide. The resulting biogenic amines are essentially Non-Proteinogenic Amino Acids that have lost their acidic component. This loss transforms them from inert structural building blocks into highly reactive physiological signaling substances within the body.
A classic example is the decarboxylation of glutamate to form GABA. By removing the carboxyl group, the excitatory molecule glutamate is transformed into the primary inhibitory neurotransmitter in the mammalian brain. This specific class of Non-Proteinogenic Amino Acids dictates the delicate balance of neurological activity, preventing overstimulation and conditions such as seizures.
Similarly, the complex precursor L-DOPA is enzymatically decarboxylated to produce dopamine. Dopamine is an essential biogenic amine that drives catecholamine production and regulates mood, reward pathways, and motor control. Its depletion is the primary biochemical hallmark of Parkinson’s disease, making this specific decarboxylation pathway a major focus of neuropharmacology and medical education.
Histidine and tryptophan undergo identical structural modifications. Decarboxylating histidine yields histamine, a potent physiological mediator responsible for inflammatory responses and gastric acid secretion. Meanwhile, tryptophan modification produces serotonin, a critical neurotransmitter that regulates sleep, appetite, and emotional well-being. These transformations highlight the incredible chemical efficiency of biological systems.
For medical and biochemistry students, mapping these precise decarboxylation pathways is absolutely essential for clinical practice. By understanding how standard precursors are stripped of their carboxyl groups to become potent signaling molecules, one gains a foundational understanding of neurochemistry, endocrinology, and the exact pharmacological targets of most modern psychiatric medications.
11. Expanding the Proteome: Non-Proteinogenic Amino Acids via Post-Translational Modification
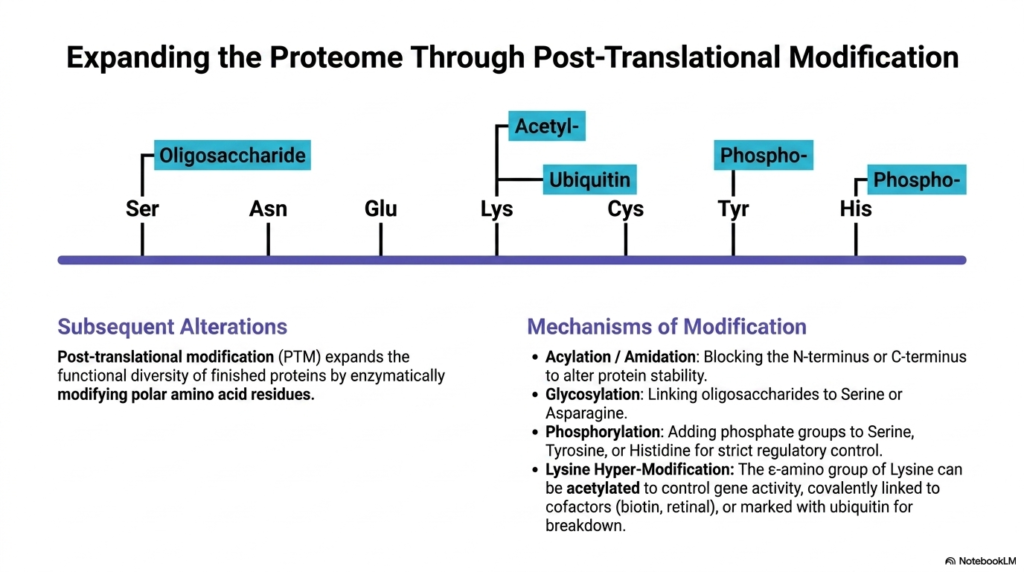
The sheer complexity of human physiology cannot be explained by the standard genetic code alone. The proteome is massively expanded after initial translation through intricate enzymatic processes. By covalently modifying polar residues on finished proteins, cells effectively create custom Non-Proteinogenic Amino Acids directly within the folded protein structure, vastly increasing functional diversity.
Post-translational modification, or PTM, alters the chemical properties of standard building blocks like serine, threonine, and tyrosine. Once modified, these residues behave exactly like distinct Non-Proteinogenic Amino Acids. They gain entirely new properties, such as altered charge, increased steric bulk, or new binding affinities, allowing proteins to dynamically change their function in response to cellular signals.
Common mechanisms include targeted acylation or amidation, which block the N-terminus or C-terminus to alter protein stability and prevent degradation. Glycosylation links complex oligosaccharides to serine or asparagine, facilitating cell-to-cell recognition and immune responses. These modifications demonstrate that the creation of Non-Proteinogenic Amino Acids is not just a metabolic phenomenon but also a major regulatory tool.
Phosphorylation is perhaps the most famous and dynamically vital modification. By adding negatively charged phosphate groups to serine, tyrosine, or histidine, cellular kinases create intense electrostatic changes within the protein. This acts as a molecular switch, turning enzyme activity on or off and driving the complex signal transduction cascades that dictate cellular behavior.
Lysine residues are particularly susceptible to hyper-modification. The epsilon-amino group of lysine can be highly acetylated to control gene activity by altering how tightly DNA wraps around histones. Furthermore, it can be covalently linked to vital cofactors like biotin for carboxylation reactions, or tagged with the bulky protein ubiquitin to mark a damaged protein for immediate destruction.
Biochemistry students must realize that the initial translation product is merely a blank canvas. These post-translational alterations are what truly give proteins their specialized functions, allowing a relatively small genome to produce an incredibly diverse and responsive array of biochemical machinery.
12. High-Impact Structural Modifications: Specialized Non-Proteinogenic Amino Acids
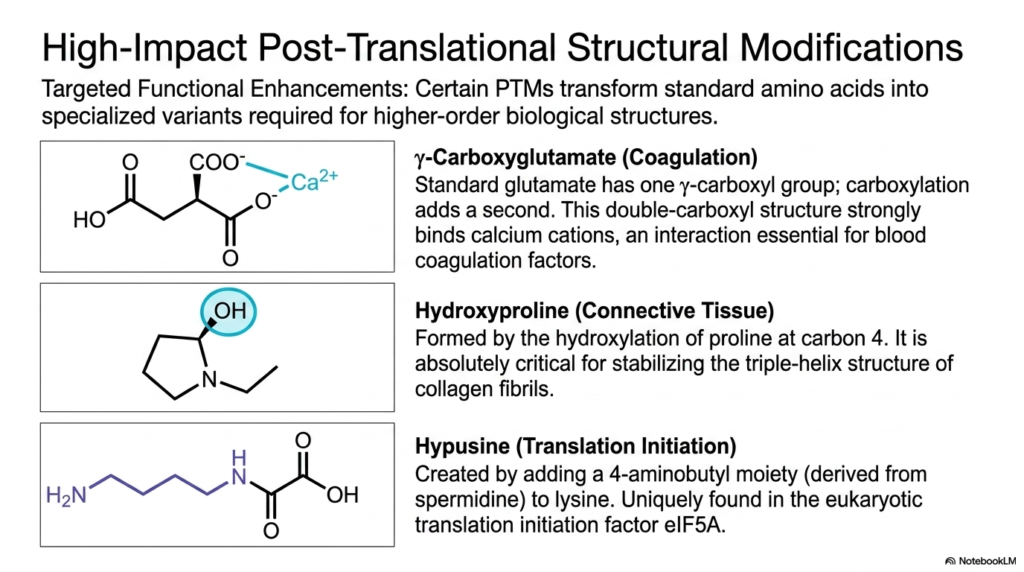
Certain cellular functions require extreme molecular specialization that standard residues simply cannot provide. Through targeted functional enhancements, cells utilize specific enzymes to transform standard building blocks into highly specialized Non-Proteinogenic Amino Acids. These structural modifications are non-negotiable requirements for generating higher-order biological structures and maintaining critical physiological processes, such as tissue integrity and blood clotting.
A classic high-impact example is the synthesis of gamma-carboxyglutamate. Standard glutamate contains one gamma-carboxyl group, but enzymatic carboxylation adds a second. This double-carboxyl Non-Proteinogenic Amino Acids structure is strongly electronegative. It acts as a powerful chelating agent, tightly binding divalent calcium cations, which is an interaction absolutely essential for activating the blood coagulation cascade.
Another profoundly important modification is the creation of hydroxyproline. This molecule is formed by the targeted hydroxylation of proline at carbon 4, requiring vitamin C as an essential cofactor. This specific Non-Proteinogenic Amino Acids variant is absolutely critical for stabilizing the triple-helix structure of collagen fibrils, providing the immense tensile strength required by connective tissues throughout the human body.
A rarer but equally vital modification yields hypusine. This complex structure is formed by adding a 4-aminobutyl moiety, derived entirely from the polyamine spermidine, directly to a standard lysine residue. Hypusine is incredibly unique because it is found exclusively in one specific protein: the eukaryotic translation initiation factor eIF5A, making it essential for cellular replication.
Without these highly targeted structural modifications, human physiology would collapse. A lack of gamma-carboxyglutamate leads to severe bleeding disorders, while a failure to produce hydroxyproline results in scurvy, characterized by massive connective tissue breakdown. These molecules demonstrate how highly specific chemical additions drive macroscopic physiological health.
For medical students, these modified residues represent critical pharmacological targets and disease markers. Understanding their precise structural alterations helps clarify the biochemical mechanisms behind nutritional deficiencies, genetic disorders, and the rational design of modern anticoagulant therapeutics.
13. Exploring Chalcogen Replacements in Non-Proteinogenic Amino Acids
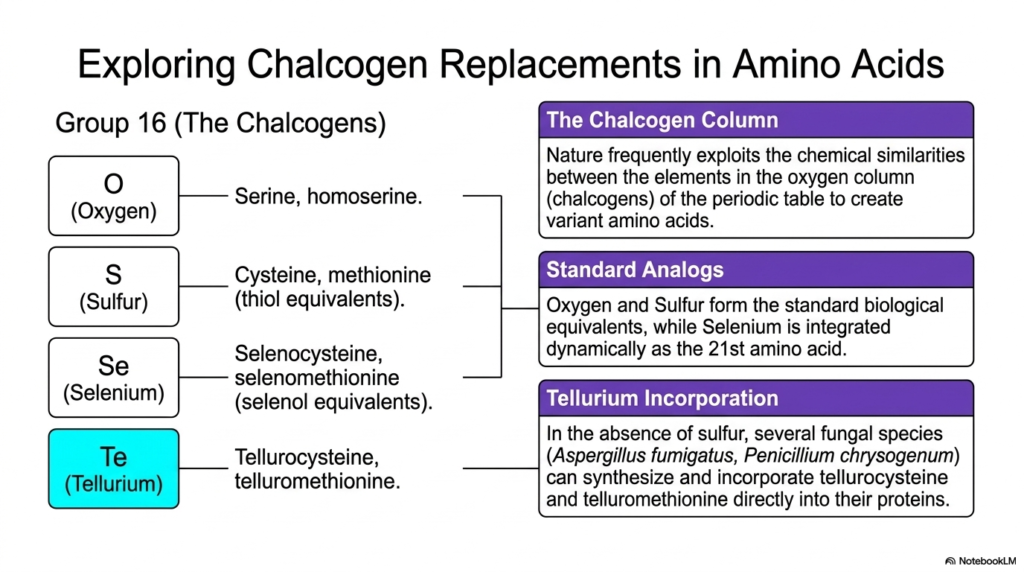
In the periodic table, Group 16 contains the chalcogens: oxygen, sulfur, selenium, and tellurium. Nature frequently exploits the chemical similarities between these vertically aligned elements to create variant molecules. By substituting these heavier chalcogens into the side chains of standard biological molecules, cells can generate highly specialized Non-Proteinogenic Amino Acids with entirely novel reactive properties.
Oxygen and sulfur are the standard biological equivalents, forming the basis for molecules such as serine, homoserine, cysteine, and methionine. However, extending down the chalcogen column reveals fascinating non-standard variants. Selenium is dynamically incorporated to form selenocysteine, essentially one of the rarest Non-Proteinogenic Amino Acids, before being officially recognized as the 21st proteinogenic building block.
The exploration of these elements leads to even more extreme biochemical adaptations. In environments completely devoid of standard sulfur, several fungal species, such as Aspergillus fumigatus, demonstrate incredible metabolic flexibility. They can synthesize tellurium-based Non-Proteinogenic Amino Acids, effectively replacing sulfur atoms with massive tellurium atoms in their biological pathways.
These fungi successfully generate tellurocysteine and telluromethionine. Remarkably, they can incorporate these massive, heavy-metal-containing variants directly into their folded proteins. While this chalcogen replacement severely distorts normal protein geometry due to the sheer size of the tellurium atom, it allows the organism to survive and maintain essential cellular functions in highly nutrient-depleted, toxic environments.
This elemental swapping highlights the incredible chemical adaptability of biological systems. While standard terrestrial life depends heavily on the lighter oxygen and sulfur atoms for hydrogen bonding and disulfide bridges, specialized extremophiles and fungi prove that the molecular machinery of life is surprisingly tolerant of heavy chalcogen substitution.
Biochemistry students studying these replacements gain insight into both evolutionary biology and industrial bioengineering. By understanding how cells tolerate and utilize these heavier analogs, researchers can design novel enzymes with unique catalytic properties, expanding the horizons of synthetic biology and bioremediation strategies.
14. Molecular Mimicry: The Toxic Nature of Certain Non-Proteinogenic Amino Acids
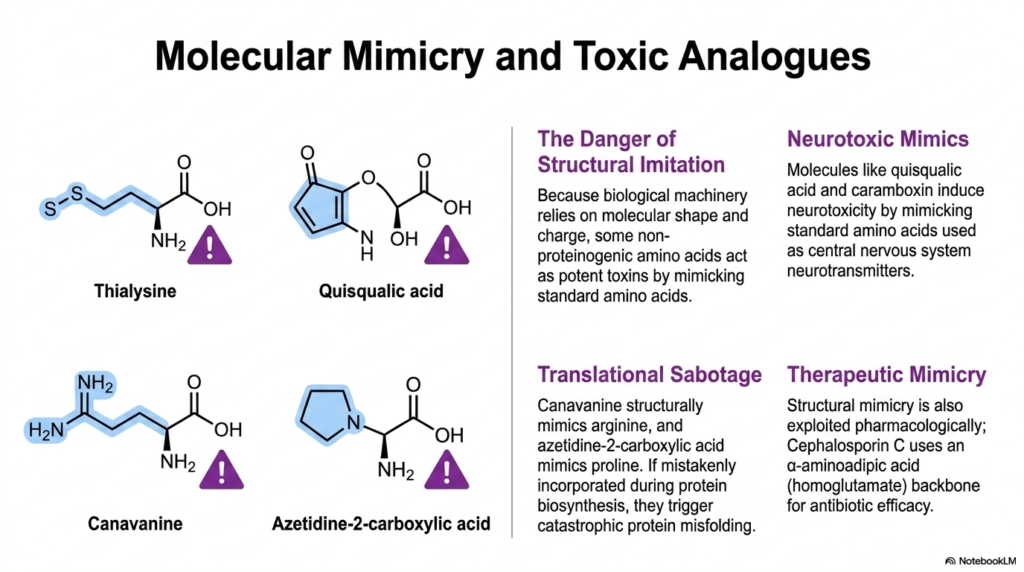
Biological machinery, particularly transfer RNA synthetases and neurological receptors, fundamentally relies on the recognition of highly specific molecular shapes and charges. Because of this reliance on physical architecture, structural imitation poses a severe threat. Some specialized Non-Proteinogenic Amino Acids act as potent toxins precisely because they successfully mimic the three-dimensional structures of standard, essential biological building blocks.
This structural mimicry can lead to catastrophic cellular sabotage. For example, canavanine structurally mimics the basic residue arginine, while azetidine-2-carboxylic acid closely imitates the cyclic structure of proline. If the translation machinery mistakenly incorporates these Non-Proteinogenic Amino Acids into a growing peptide chain, it triggers massive protein misfolding, rapidly leading to cellular death.
Beyond translational errors, molecular mimicry frequently induces severe neurotoxicity. Molecules like quisqualic acid and caramboxin are dangerous Non-Proteinogenic Amino Acids naturally found in certain plants. Because their structures closely resemble endogenous central nervous system neurotransmitters like glutamate, they can aggressively bind to and overstimulate neuronal receptors, causing excitotoxicity, severe seizures, and devastating brain damage.
Plants and fungi often synthesize these toxic analogs as powerful chemical defenses against herbivores and competing microbes. By filling their tissues with these deceptive molecules, they ensure that any organism consuming them will suffer immediate metabolic or neurological breakdown. This represents a fascinating evolutionary arms race fought entirely on a biochemical level.
However, structural mimicry is not always destructive; it is heavily exploited in modern pharmacology. Therapeutic mimicry utilizes this same concept to fight infections. The powerful antibiotic Cephalosporin C utilizes a highly specific alpha-aminoadipic acid backbone—acting as a homoglutamate mimic—to bypass bacterial defenses and achieve highly effective antimicrobial efficacy without harming human host cells.
For medical students, understanding these toxic mimics bridges the gap between toxicology, ecology, and pharmacology. Recognizing how slight structural variations lead to massive physiological disruptions is the core principle behind understanding both natural venoms and the rational design of safe, targeted synthetic drugs.
15. Synthesis and Functional Matrix of Non-Proteinogenic Amino Acids

To consolidate this extensive biochemical analysis, a comparative functional matrix perfectly synthesizes the core domains discussed. The standard proteinogenic molecules serve strictly as translation building blocks, genetically encoded by universal codons. In stark contrast, Non-Proteinogenic Amino Acids operate entirely outside this universal coding system, functioning primarily in structural tuning, metabolic cycling, and intricate cellular signaling.
This matrix categorizes the Non-Proteinogenic Amino Acids into distinct functional domains. Metabolic variants such as citrulline regulate the urea cycle and exhibit structural hallmarks such as twin stereocenters. Meanwhile, biogenic amines, such as dopamine and serotonin, are enzymatically derived via decarboxylation, losing their carboxyl group to become highly reactive cell-signaling powerhouses within complex nervous systems.
The prebiotic domain further rounds out the matrix, reminding us of life’s chemical origins. Molecules like norvaline represent the initial chemical precursors synthesized abiotically in early Earth environments. These specific Non-Proteinogenic Amino Acids are characterized by their straight side chains and often by achiral centers, and they existed freely in meteorites long before the standard genetic code evolved.
The final insight of this presentation is perhaps the most profound for developing biochemists. The 20 standard biological building blocks represent merely the foundational alphabet of life. While they form the structural scaffolding of proteins, they are far too limited to support a dynamic, responsive, and complex physiological organism on their own.
The vast catalog of non-standard variants provides the essential grammar, punctuation, and physiological syntax required for higher-order biological systems to actually function. From sending rapid thoughts across a synapse to successfully crosslinking a protective bacterial wall, these molecules dictate the precise timing and structure of life’s most critical chemical reactions.
Ultimately, medical and college students must view the metabolome as a highly integrated network. The standard twenty build the machines, but the non-standard molecules serve as the fuel, the messengers, and the precise regulatory switches that bring cellular biology to life. Mastering this distinction is the key to advanced biochemical understanding.
Please read our Content Disclaimer Statement.
Check out our social media channels: