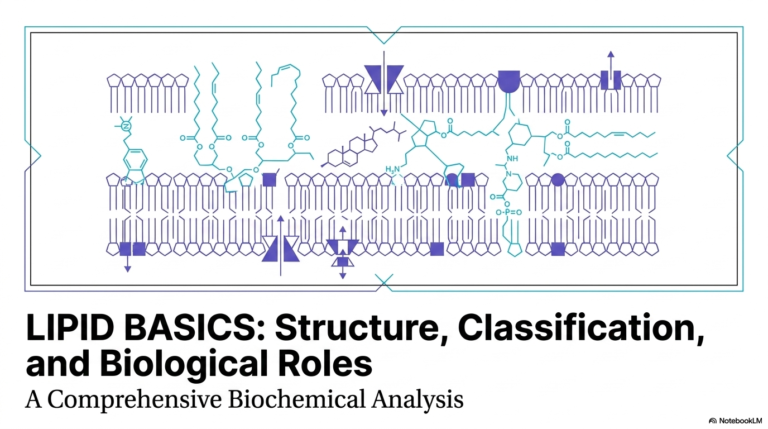
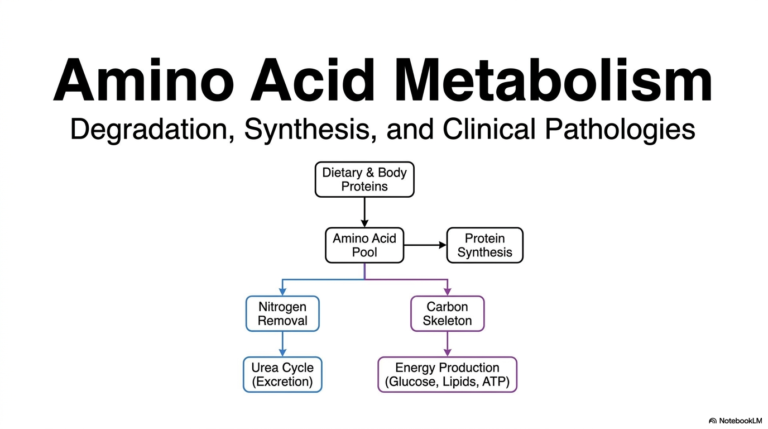
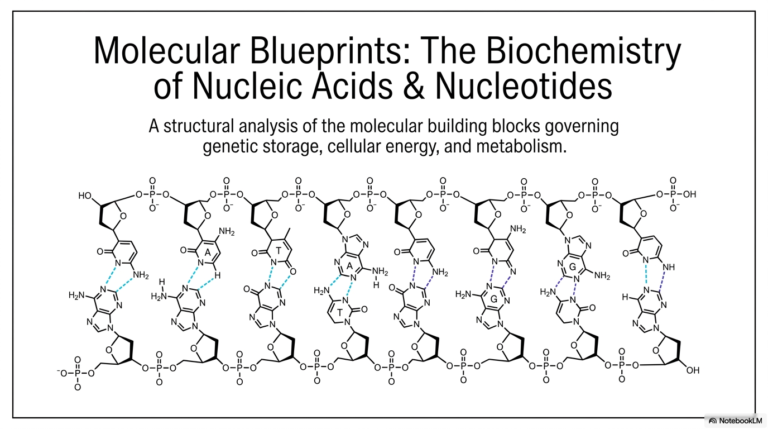
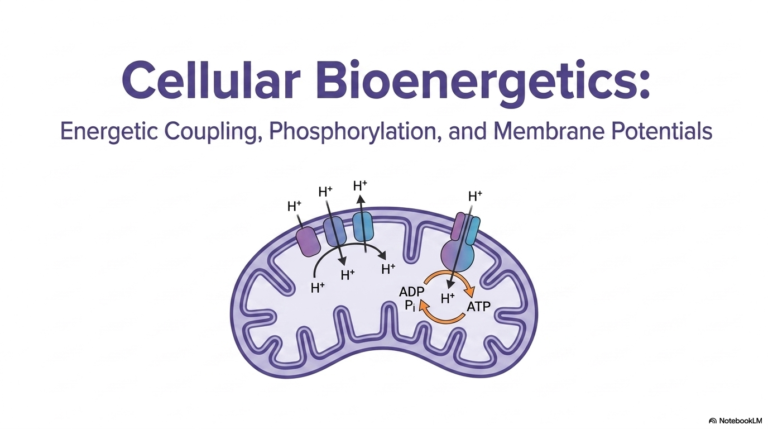
5. Demystifying Catalysis: The Engine of Biochemical Reactions
Imagine if you had to wait thousands of years just for your body to process the breakfast you ate this morning. Without specific molecular accelerators, the biochemical reactions required to sustain human life would be so agonizingly slow that existence itself would be impossible. The core purpose of this slide deck is to demystify these molecular machines. We will explore the energetic barriers that prevent spontaneous combustion in our cells and how biological catalysts gracefully overcome them. This comprehensive guide breaks down the complex biochemical principles of enzymatic action so you can understand exactly how these structures bend the rules of chemistry to keep us alive.
Slide 1: The Foundations of Catalysis
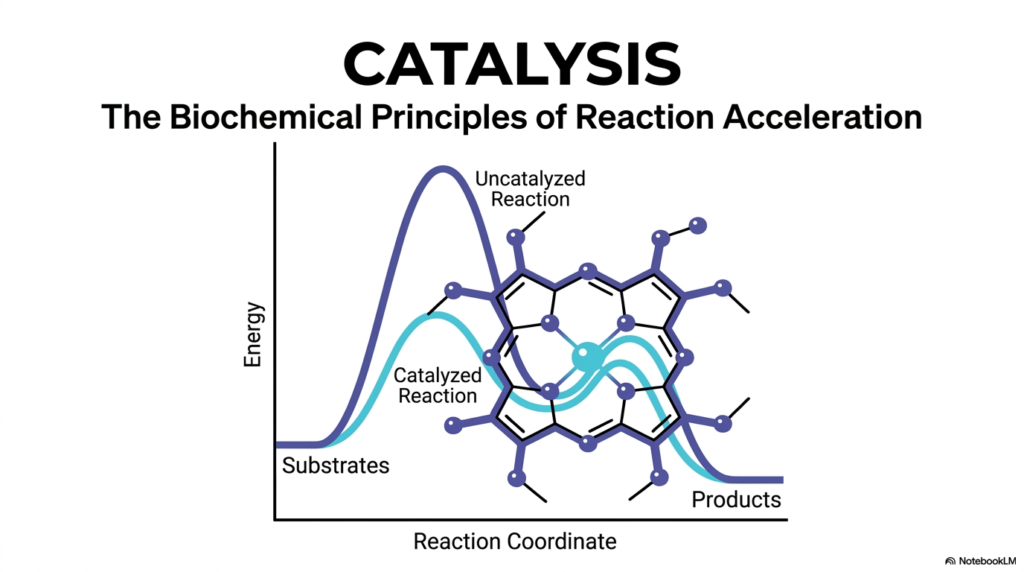
Welcome to the incredible world of accelerating biochemical reactions. The cover slide of our deck introduces the central visual and conceptual themes we will be exploring: the energy landscape of chemical reactions and the complex molecular structures that manipulate it. At the heart of biological survival is catalysis, the process by which specific reactions are sped up to physiological relevance. In the center of the slide, you see a complex molecular structure—a porphyrin ring holding a central ion. This represents the heme group, a recurring structural motif in biochemistry that you will encounter frequently in your medical studies, particularly when studying hemoglobin, myoglobin, and cellular cytochromes.
The graph behind the molecule illustrates the fundamental problem that catalysis successfully solves. You can see two distinct curves representing the energy state of a reaction as it transforms from substrates to products. The taller, dark blue curve represents an agonizingly slow, uncatalyzed reaction, characterized by a massive energetic hurdle. The lower, light blue curve represents the much faster, accelerated pathway. Understanding how biological systems achieve this lower energetic curve is essential for any student of biochemistry.
Without the power of catalysis, the substrates in our cells would remain completely inert, trapped behind insurmountable energy barriers. We will journey through the mechanisms of activation energy, the precarious nature of transition states, and the stunning efficiency of enzymes. By the time you finish studying this deck, you will have a rigorous understanding of reaction acceleration, bridging the gap between abstract physical chemistry and the dynamic processes occurring within every human cell.
Slide 2: Defining Biological Catalysis
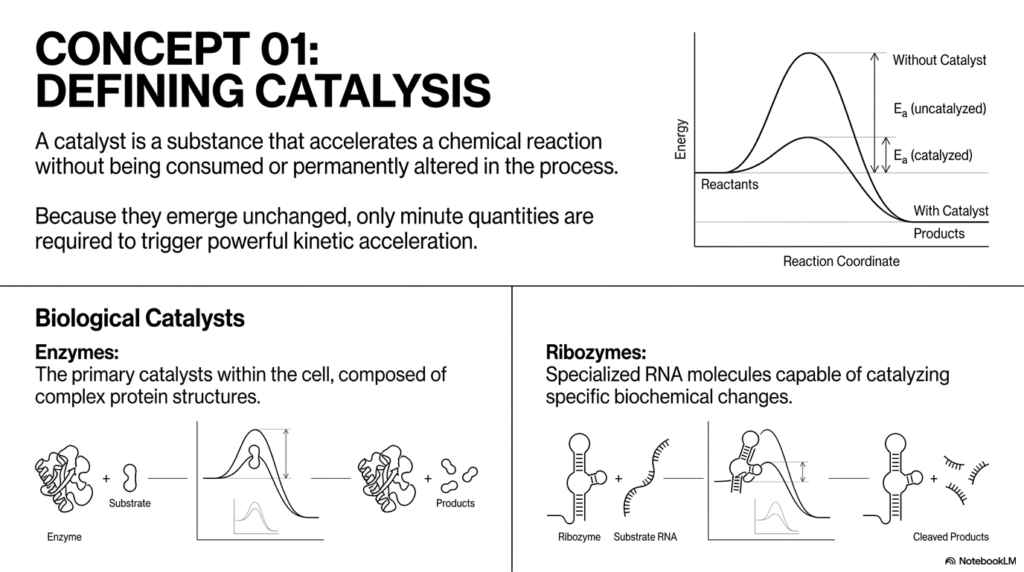
To build a solid biochemical foundation, we must establish a rigorous definition of what a catalyst actually is. According to the slide, a catalyst is a substance that accelerates a chemical reaction without being consumed or permanently altered in the process. This definition highlights the incredible reusability of these molecules. Because they emerge completely unchanged at the end of the reaction cycle, only minute quantities are required to trigger powerful kinetic acceleration. This foundational principle of catalysis explains how trace amounts of certain enzymes can sustain massive metabolic fluxes within a cell.
In the biological realm, we primarily deal with two distinct classes of molecules capable of true catalysis. The most common and versatile are enzymes. Enzymes are the primary catalysts within the cell, composed of complex, perfectly folded protein structures. The slide illustrates how an enzyme physically binds a substrate, facilitates a structural chemical change, and releases the products, returning to its native, ready state. The precise three-dimensional folding of these proteins creates highly specific active sites, enabling remarkable substrate specificity. (Outside of our primary sources, it is worth noting that this specificity is often described by the induced-fit model, where the enzyme undergoes slight conformational changes upon substrate binding. Please verify this supplementary model independently.
However, proteins do not have a monopoly on biological catalysis. The slide also introduces ribozymes: specialized RNA molecules capable of catalyzing specific biochemical changes. The visual demonstrates a ribozyme binding to a substrate RNA molecule and cleaving it into products. The discovery of ribozymes was a massive paradigm shift in molecular biology, proving that nucleic acids could be more than just passive genetic blueprints. They can also act as active molecular machines, accelerating vital intracellular reactions.
Slide 3: Overcoming Activation Energy in Catalysis
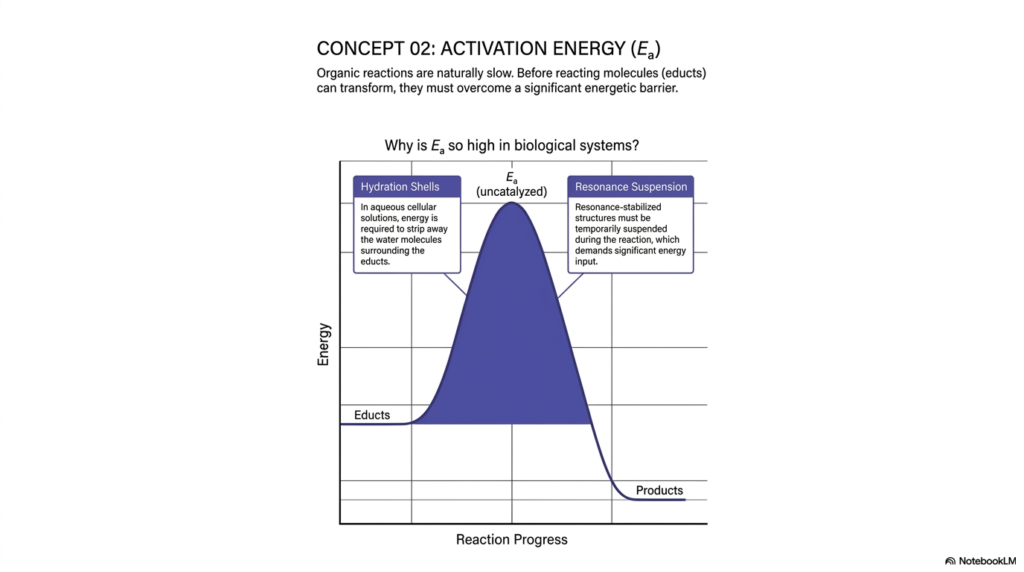
Before we can completely understand how acceleration works, we must understand why reactions are slow in the first place. Concept 02 introduces Activation Energy. The slide explains that organic reactions are naturally sluggish because before reacting molecules—termed educts—can transform into products, they must first overcome a significant energetic barrier. The peak of this tall blue curve represents a massive energy debt that must be paid before the reaction can proceed downhill.
Why is this activation energy so prohibitively high in biological systems? The slide outlines two primary reasons that necessitate robust biological catalysis. First, we must consider hydration shells. Our cells are heavily aqueous environments. Molecules do not float freely in a vacuum; they are surrounded by tightly organized cages of water molecules. To bring two electrons together, cellular energy is required to strip away these surrounding water molecules. Overcoming the intermolecular forces of the hydration shell is a major energetic hurdle that effective catalysis must somehow bypass.
The second major factor is resonance suspension. Many critical organic molecules exist in highly stable, resonance-stabilized states. During a chemical reaction, these stable electronic structures must be temporarily suspended or physically disrupted to form new bonds. Pushing a molecule out of its comfortable, low-energy resonance state requires a substantial energy input. Without catalytic intervention, the ambient thermal energy within a cell is rarely sufficient to overcome the combined barriers of hydration and resonance.
Slide 4: Reaching the Transition State in Catalysis
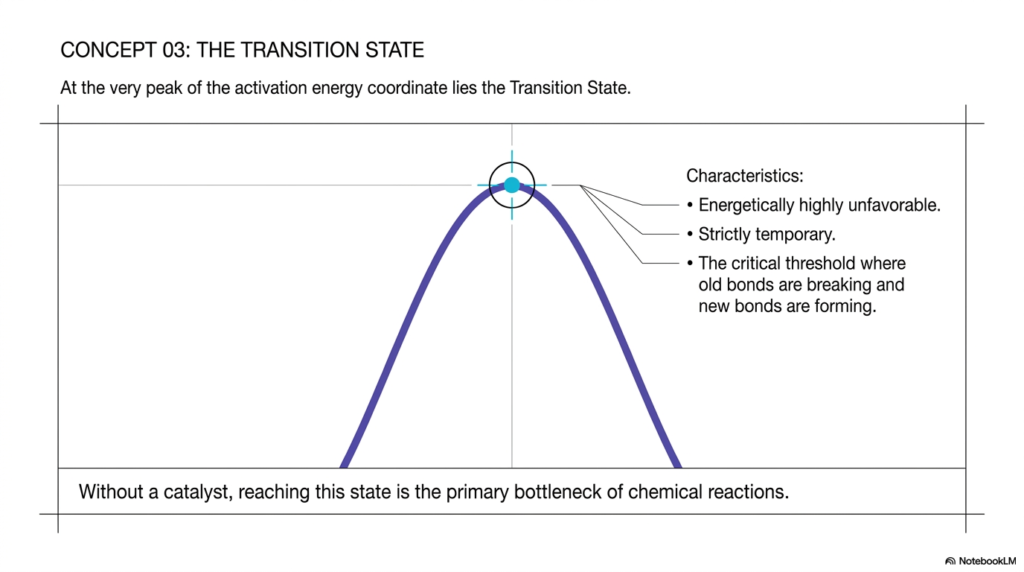
At the very peak of the activation energy coordinate lies the focal point of all chemical reactions: the Transition State. As depicted in the slide, this state is perfectly nestled at the absolute apex of the energy curve. Understanding the transition state is arguably the most critical component of mastering the mechanics of catalysis.
The slide lists three defining characteristics of this state. First, it is energetically highly unfavorable. Molecules naturally resist reaching this state because nature inherently favors the lowest possible energy configurations. Second, it is strictly temporary. The transition state is not a stable chemical intermediate that you can isolate and put in a jar; it is a fleeting, transient moment in time. Third, it is the critical threshold at which old chemical bonds are actively breaking while new ones are simultaneously forming. It represents the ultimate point of no return for the reacting educts.
Without the direct aid of a catalyst, reaching this highly unstable geometry is the primary bottleneck of chemical reactions. The sheer improbability of molecules colliding with sufficient force and in the right orientation to achieve this state makes uncatalyzed reactions impossibly slow. Effective catalysis works precisely by stabilizing this awkward transition state. By physically binding to the educts and forcing them into optimal orientations, catalysts dramatically reduce the energetic cost of reaching this critical threshold.
Slide 5: Forging Alternative Pathways Through Catalysis
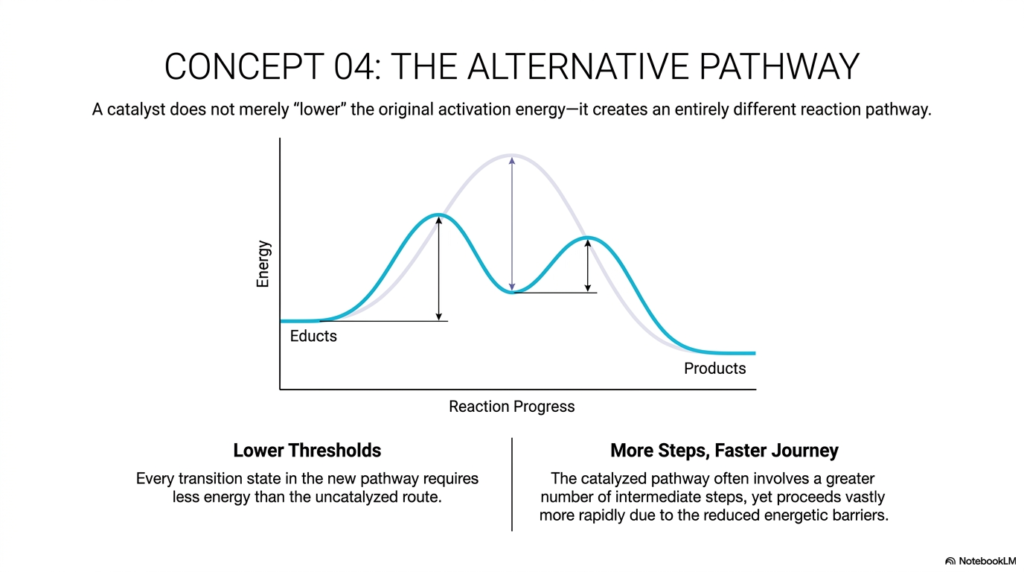
Concept 04 addresses one of the most common conceptual misconceptions in biochemistry. Often, students are taught that a catalyst simply “lowers” the activation energy of a reaction. However, the slide emphatically states that a catalyst does not merely lower the original activation energy barrier—it creates an entirely different reaction pathway. This is a profound and fundamental distinction in the study of catalysis.
If you look closely at the graph on the slide, the purple curve represents the uncatalyzed path, presenting as a single, massive hurdle. The teal curve represents the new catalyzed path. Notice that the teal curve is not just a mathematically shrunken version of the purple one; it actually has two distinct peaks and a small valley in between. This vividly illustrates that the new pathway involves completely different molecular intermediates and a drastically altered mechanism of action. The true essence of catalysis is to provide a safer, more energetic detour.
This alternative pathway features two key hallmarks. First, it has significantly lower thresholds. Every transition state in this new, alternative pathway requires vastly less energy than the towering peak of the uncatalyzed route. Second, it often involves more steps. The slide points out the apparent paradox: the catalyzed pathway often involves more intermediate chemical steps, yet it proceeds vastly more rapidly. This is simply because multiple small energetic hurdles are much easier for a molecular system to clear than a single massive, insurmountable wall. Thus, biological catalysis isn’t simply making the journey physically shorter; it’s about making the journey energetically feasible, step by manageable step.
Slide 6: Thermodynamics vs. Kinetics in Catalysis
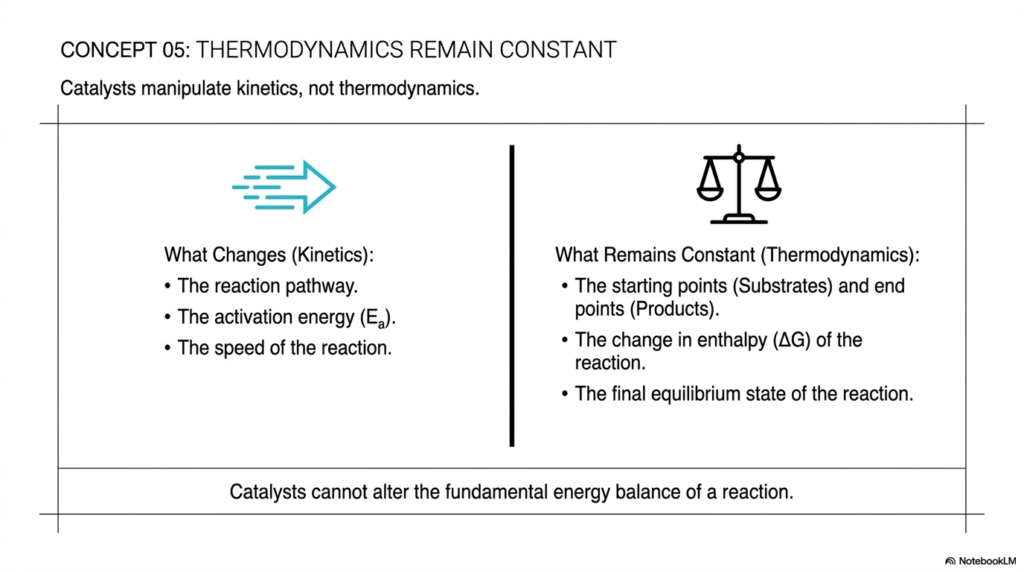
Concept 05 provides a crucial physical boundary for what enzymes can and cannot do, reminding us that thermodynamics remain constant. This slide draws a hard, undeniable line between kinetics (the speed of a reaction) and thermodynamics (the overall energy balance of a reaction). Catalysts are absolute masters of kinetics, but they cannot violate the fundamental laws of thermodynamics.
The left side of the slide outlines what actually changes under the influence of catalysis. A catalyst manipulates the chemical reaction pathway, dramatically lowers the activation energy, and drastically increases the reaction rate. These are exclusively kinetic properties. The catalyst dictates how quickly we reach the destination and the exact molecular route we take to get there.
However, the right side of the slide lists what remains absolutely constant, even under the most potent catalysis. The starting points (the original energy level of the substrates) and the end points (the final energy level of the products) simply do not shift. Consequently, the overall change in the reaction’s enthalpy remains identical. Finally, the equilibrium state of the reaction remains unchanged.
A catalyst will help a cellular system reach equilibrium billions of times faster, but it will never shift the final ratio of products to reactants at that equilibrium. (It is important for medical students to understand that if a reaction is highly endergonic and thermodynamically unfavorable, no amount of catalysis alone can force it to occur; the cell must couple it to an exergonic reaction like ATP hydrolysis.
Slide 7: The Peroxide Problem and Catalysis
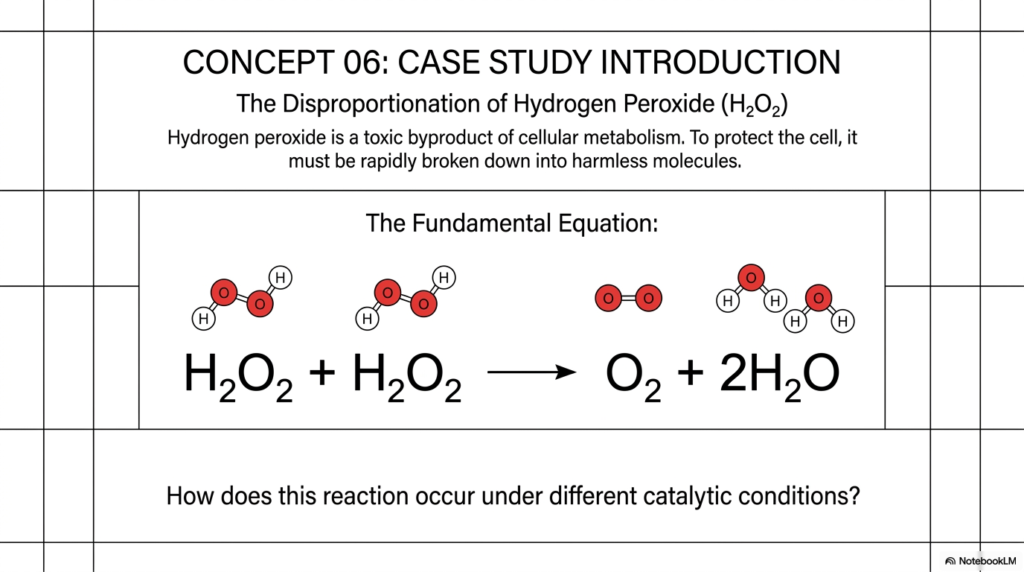
With the theoretical framework securely established, Concept 06 transitions the deck into a concrete, highly relevant biochemical case study: the disproportionation of hydrogen peroxide. This slide introduces the immense clinical and biological stakes that require aggressive catalysis in our cells every second of the day.
Hydrogen peroxide is a notoriously toxic byproduct of cellular metabolism. As our mitochondria efficiently burn fuel to generate energy, they inevitably produce dangerous reactive oxygen species, including hydrogen peroxide. If left unchecked, this peroxide will rapidly wreak havoc on the cell, indiscriminately oxidizing lipids, denaturing vital proteins, and mutating structural DNA. To protect the cell and ensure continuous survival, this toxic molecule must be rapidly broken down into harmless molecules. This is a literal life-or-death scenario where highly efficient catalysis is absolutely mandatory.
The slide presents the fundamental chemical equation we must solve: two molecules of hydrogen peroxide react to form one molecule of molecular oxygen and two molecules of water. Visually, you can easily see the red oxygen atoms and the small white hydrogen atoms elegantly rearranging themselves into safe, biologically compatible products. The critical question posed at the bottom of the slide perfectly sets the stage for the rest of the deck: How does this essential reaction occur under different catalytic conditions? By systematically comparing peroxide breakdown across different states, we will witness the undeniable power and absolute necessity of enzymatic catalysis.
Slide 8: The Baseline: Life Without Catalysis
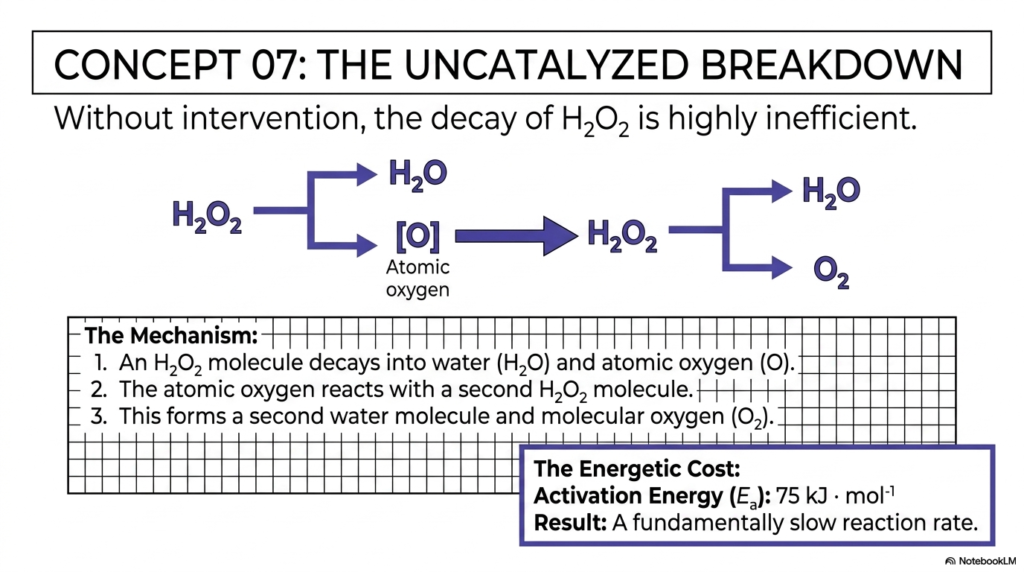
Concept 07 clearly establishes the biochemical baseline by examining the uncatalyzed decomposition of hydrogen peroxide. This illustrates exactly what would happen if our cells suddenly possessed no mechanisms for catalysis. As the slide bluntly states, without direct intervention, the physical decay of hydrogen peroxide is highly inefficient and dangerously slow.
The slide breaks down the uncatalyzed chemical mechanism into distinct steps. First, an individual hydrogen peroxide molecule rapidly decomposes into water and a single isolated atom of oxygen. Second, this highly reactive atomic oxygen must wander the environment to find and react with a second, intact hydrogen peroxide molecule. Finally, this successful collision forms a second water molecule and stable molecular oxygen. The primary issue with this pathway is that the free atomic-oxygen intermediate is highly unstable, making this route energetically prohibitive.
The true problem is neatly quantified at the bottom of the slide: The Energetic Cost. The raw activation energy required for this uncatalyzed route is a steep 75 kJ per mole. Because this energetic barrier is so high, very few molecules at physiological body temperature possess the kinetic energy to naturally overcome it. The result is a fundamentally sluggish, slow reaction rate. If our bodies had to patiently rely on this uncatalyzed pathway, toxic peroxide would accumulate millions of times faster than it could be safely cleared, leading to rapid cellular death. The physiological necessity for rapid catalysis becomes glaringly obvious when we examine the sheer energetic cost of doing nothing.
Slide 9: Inorganic Catalysis with Iodide
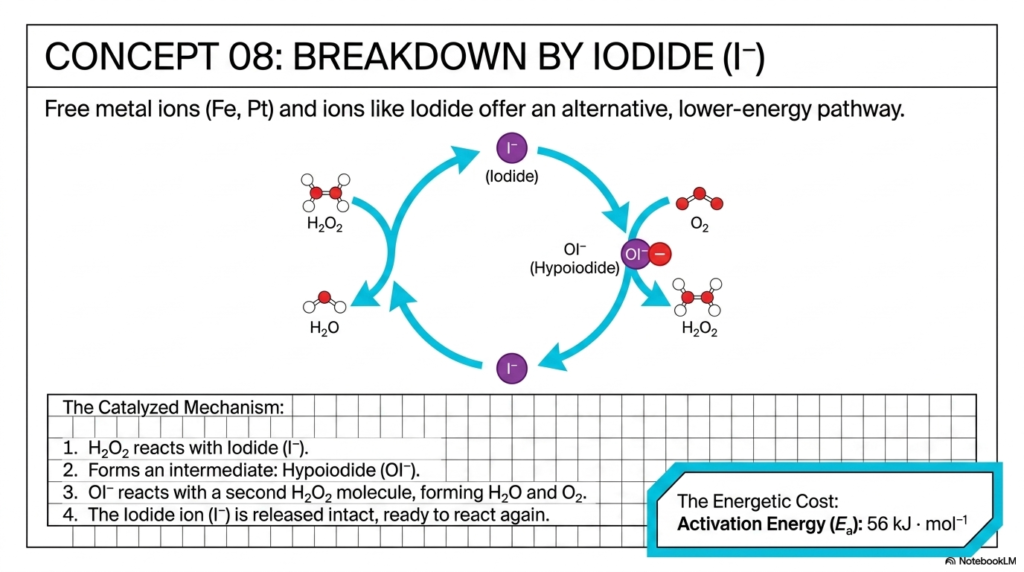
Concept 08 introduces our first, relatively simple molecular solution: inorganic catalysis. The slide demonstrates that free metal ions (such as iron or platinum) and simpler ions, like Iodide, can provide an alternative, significantly lower-energy pathway for the disproportionation of peroxide. This represents a functional, non-enzymatic form of basic catalysis.
The visual provides a beautifully clear, cyclical mechanism for this alternate reaction. First, the hydrogen peroxide reacts directly with the free Iodide ion. Second, this collision successfully forms a crucial intermediate: Hypoiodite. Notice how conceptually different this is from the uncatalyzed pathway: instead of using highly reactive atomic oxygen, we have generated a relatively stable ionic intermediate. Third, this Hypoiodite readily reacts with a second hydrogen peroxide molecule, rapidly forming water and molecular oxygen. Finally, and most importantly for the strict chemical definition of catalysis, the Iodide ion is released completely intact, perfectly ready to initiate the cycle all over again.
By providing this alternative, chemically distinct route, the Iodide catalyst significantly reduces the total energetic cost. The activation energy drops notably from a steep 75 kJ/mol to 56 kJ/mol. This is a substantial reduction, firmly demonstrating that even relatively simple ions can readily catalyze effectively. While it admittedly lacks the extreme structural sophistication of a dedicated protein, iodide perfectly demonstrates the fundamental scientific principle of altering the entire reaction mechanism to effectively bypass towering energy barriers.
Slide 10: The Exponential Math Behind Catalysis

Concept 09 smoothly bridges the conceptual gap between physical chemistry and biological reality by properly introducing Exponential Kinetics. The core mathematical question on the slide is absolutely vital: Why does a seemingly small drop in Activation Energy—from 75 to 56 kJ/mol—matter so incredibly much? The secret to the immense power of catalysis lies firmly in exponential math.
The slide displays a highly simplified, practical version of the Arrhenius equation: v∼e−Ea/RT. This equation explicitly states that the overall reaction rate (v) depends exponentially on the negative activation energy. Because the activation energy variable is positioned squarely in the exponent, the resulting mathematical relationship is not merely linear; it is aggressively exponential. Even a modest, linear reduction in the energy barrier instantly yields an explosive, exponential increase in the number of molecules that can successfully cross that barrier per second.
The bottom of the slide spells out the profound result of the inorganic Iodide catalysis. By actively reducing the activation energy by just 19 kJ/mol, the seemingly simple Iodide ion accelerates the overall reaction by a staggering, massive factor of 2,000. Let that mathematical reality sink in. A relatively modest drop in the required physical energy makes the reaction two thousand times physically faster. This exponential relationship is the true biological engine that drives all cellular efficiency. It beautifully explains how specialized molecules can effortlessly accelerate reactions that would naturally take years, forcing them to occur in mere fractions of a second.
Slide 11: Enter Catalase: Perfecting Catalysis

Concept 10 smoothly transitions from the impressive, yet rudimentary, inorganic catalysts straight to the pinnacle of biological engineering: precise Enzymatic Catalysis. While inorganic catalysts like iodide are admittedly effective and accelerate the reaction 2,000-fold, the slide quickly reminds us that cellular survival actually demands total molecular perfection. Enter the enzyme Catalase.
Catalase is formally introduced as an incredibly potent enzyme, shaped over millions of years of evolution, that effectively protects fragile cells against the toxic oxidative effects of hydrogen peroxide. Unlike the remarkably simple ions, true catalase is a massive, highly structured protein complex. It undeniably represents the ultimate, highly evolved form of biological catalysis. The slide’s faint, artistic blue background image subtly hints at this structural complexity, showing the intricate secondary- and tertiary-folded protein structures that precisely make up the massive body of the enzyme.
The key biological takeaway is boldly stated at the bottom of the slide: Catalase does not merely reduce the activation energy mathematically; it uniquely uses its complex protein architecture to physically and forcibly manipulate the substrate. Simple chemical ions just randomly bump into peroxide. Supreme enzyme catalysis involves aggressively grabbing the substrate, perfectly orienting it, physically excluding water from the sensitive active site, and literally stressing chemical bonds to achieve unmatched efficiency. The precise three-dimensional structure of the protein is what enables this highly controlled, rapid degradation of toxins.
Slide 12: The Heme Group’s Role in Catalysis
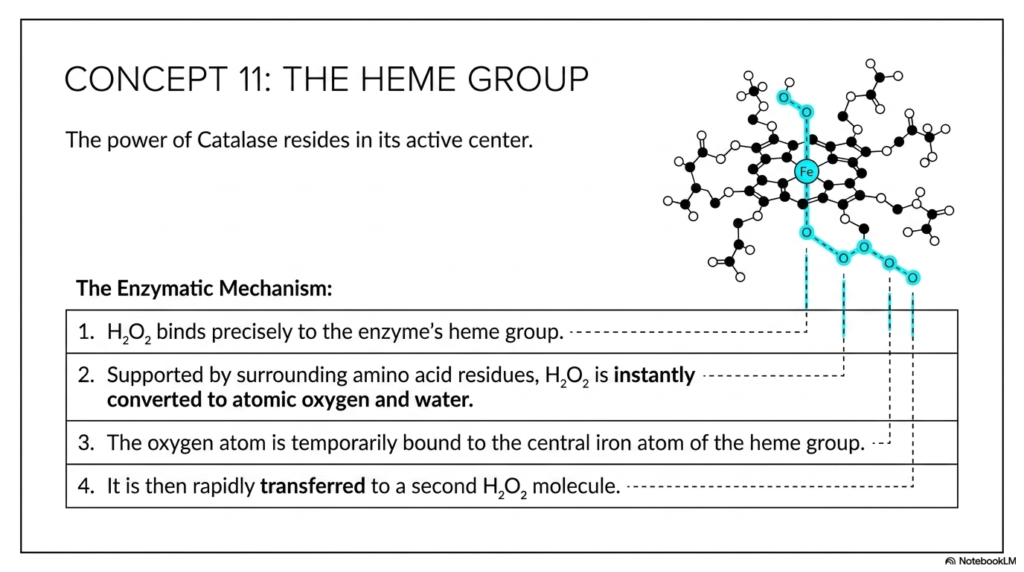
Concept 11 finally zooms in incredibly close on the exact chemical mechanism of Catalase, beautifully revealing that its immense kinetic power primarily resides in its tiny active center: the Heme Group. This slide is a completely breathtaking demonstration of precise structural biology directly driving efficient catalysis. The clear visual boldly shows the incredibly intricate porphyrin ring structure, heavily stabilized by numerous carbon and nitrogen bonds, tightly clutching a single, central Iron atom.
The slide cleanly outlines the remarkably precise enzymatic mechanism in four distinct, sequential steps. First, hydrogen peroxide binds specifically and absolutely perfectly directly to the enzyme’s internal heme group. Second, massively supported by the carefully arranged surrounding amino acid residues of the tight protein pocket, the peroxide is instantly and violently converted to atomic oxygen and plain water. Third, unlike the dangerous, free-floating atomic oxygen found in large amounts in the uncatalyzed route, this specific oxygen atom is safely, tightly, and temporarily bound directly to the central iron atom of the protected heme group. Finally, it is rapidly and safely transferred directly to a second incoming peroxide molecule.
This highly orchestrated, flawless molecular dance is the absolute essence of advanced catalysis. The precisely positioned surrounding amino acid residues mentioned in step 2 are critical; they act like tiny, microscopic hands, continuously pushing and pulling on the substrate’s sensitive electrons to efficiently facilitate rapid bond breaking. The central iron atom acts as a totally perfect temporary chemical storage site for the highly reactive, dangerous oxygen intermediate. This slide stunningly demonstrates how biological enzymes consistently catalyze through precise spatial geometry and highly targeted chemical interactions.
Slide 13: Pushing the Limits of Catalysis

Concept 12 brightly reveals the sheer, almost mind-bending mathematical numbers driving the enzyme’s ultimate performance. Catalase is widely recognized as one of the most efficient enzymes ever known to modern science, routinely pushing the absolute physical boundaries of catalysis.
Let’s carefully look at the stunning metrics. First, the vital Activation Energy is forcefully driven all the way down to a mere 23 kJ/mol. Directly compare this incredibly low number to the towering 75 kJ/mol of the slow, uncatalyzed reaction. Because of the aggressive exponential kinetics we learned about earlier, this unbelievably deep reduction mathematically yields a reaction rate that is roughly 1.3 billion times faster than the entirely uncatalyzed reaction. The simple bar chart clearly visually represents this dramatic, life-saving drop in required kinetic energy.
However, the most genuinely staggering mathematical statistic squarely on the slide is the actual Turnover Rate. A single Catalase molecule can convert up to 100 million molecules of hydrogen peroxide into water and oxygen every second. This reaction rate is so incredibly fast that the massive enzyme is not genuinely limited by how quickly it can perform the actual chemistry, but simply by how surprisingly fast the small substrate molecules can physically diffuse through the ambient water to reach the tight active site. This incredible phenomenon is a profound hallmark of supreme, perfect biological catalysis.
Slide 14: Analyzing the Efficacy of Catalysis
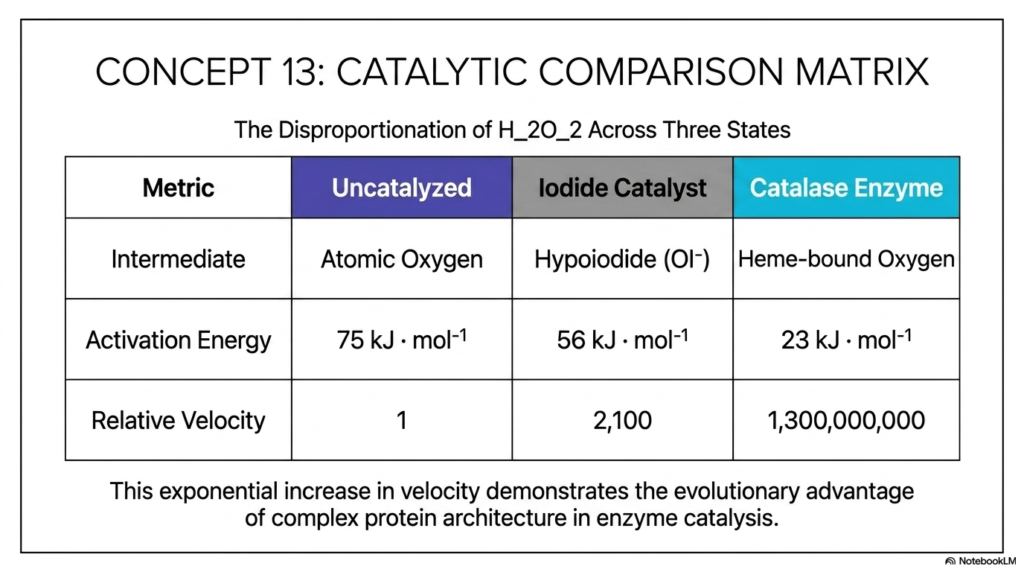
Concept 13 neatly provides a remarkably clear, highly quantitative summary directly via the comprehensive Catalytic Comparison Matrix. This incredibly useful table perfectly encapsulates the core, overarching narrative of the entire slide deck: systematically comparing the specific disproportionation of hydrogen peroxide heavily across three entirely distinct states of intense catalysis.
The organized matrix cleanly tracks three highly specific metrics. First, for the required intermediate chemical state, the dangerously slow uncatalyzed reaction relies heavily on fully free atomic oxygen; the moderate iodide catalyst uses a slightly safer hypoiodite ion; and the supreme catalase uses highly controlled, completely heme-bound oxygen. Second, purely for activation energy, we clearly see a beautiful, progressive drop from 75 kJ/mol down to 56 kJ/mol, and finally to an amazing 23 kJ/mol. Finally, directly looking at relative reaction velocity, if the baseline uncatalyzed reaction has a slow baseline speed of merely 1, simple iodide catalysis operates nicely at 2,100, while perfect catalase operates at a virtually incomprehensible, blinding 1,300,000,000.
The clear text squarely at the bottom neatly summarizes the grand, profound biological takeaway: this incredibly exponential, massive increase in velocity clearly demonstrates the sheer evolutionary advantage of possessing complex protein architecture directly in advanced enzyme catalysis. Over billions of long years, biological cells that evolved large, incredibly complex proteins to flawlessly execute highly specific chemical maneuvers survived, while those that relied strictly on slow, ambient chemistry rapidly perished under massive oxidative stress. This powerful matrix isn’t just a simple chart of numbers; it is literally a mathematical proof securely validating the power of natural selection operating at the absolute molecular level.
Slide 15: The Supreme Paradigm of Catalysis

The final slide, Concept 14, beautifully summarizes the entire deck by broadly outlining the awesome Power of Biological Catalysis as a true paradigm of sheer biological efficiency. It leaves us neatly with three incredibly profound, final educational takeaways directly regarding exactly how advanced enzymes successfully sustain complex human life.
First is clear Pathway Creation. As we have successfully learned, highly effective catalysis does not magically cheat the strictly unyielding, rigid laws of thermodynamics; instead, it intelligently forges entirely new, highly energetically favorable chemical pathways. Powerful enzymes are essentially molecular biological architects, actively building safe chemical tunnels straight through massive thermodynamic mountains. Second is incredible Exponential Returns. strictly because of the Arrhenius equation, even seemingly modest linear physical reductions in the high activation energy can easily yield massive, explosive exponential increases in reaction velocity. This undeniable mathematical reality is effectively the secure foundation of absolutely all biochemical speed.
Finally, the comprehensive deck perfectly concludes with Biological Supremacy. strictly through incredibly precise, highly evolved structural mechanics—perfectly epitomized by the perfectly positioned central iron atom deep within the active heme group—enzymes like Catalase effortlessly push complex chemical reactions to the absolute, unyielding limits of basic physical efficiency. They proudly represent billions of years of intense molecular R&D, resulting in highly advanced molecular machines that operate incredibly close to the theoretical mathematical limits of physics.
As you actively continue your difficult medical and biochemical higher education, please constantly remember that absolutely every metabolic cellular pathway, every vital cellular biological signal, and every single physiological bodily response you intensely study is fundamentally and beautifully driven entirely by the simply breathtaking power, unbelievable speed, and absolute precision of biological catalysis.
Check out our social media channels:
Please read our Content Disclaimer Statement.