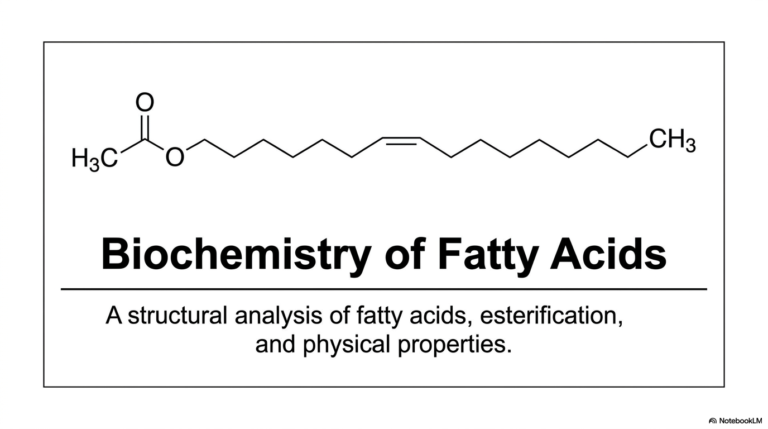
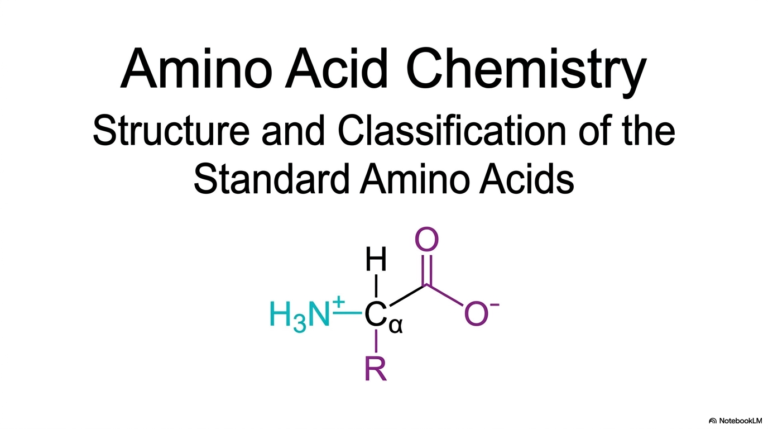
18. Understanding the Peptide Bond: Structure and Thermodynamics
Proteins are the microscopic machines that keep living organisms functioning, but their immense complexity stems from a beautifully simple set of chemical rules. Imagine trying to build an infinitely diverse array of complex structures using only a single type of locking hinge; this is precisely what biology achieves at the molecular level. The core purpose of this comprehensive slide deck is to explore the structural mechanics, thermodynamics, and conformational rules that govern protein architecture. By mastering these fundamental biochemical concepts, students and medical professionals will discover how simple atomic linkages dictate the intricate, life-sustaining folded shapes of all biological macromolecules.
Slide 1: Introduction to the Peptide Bond and Protein Architecture
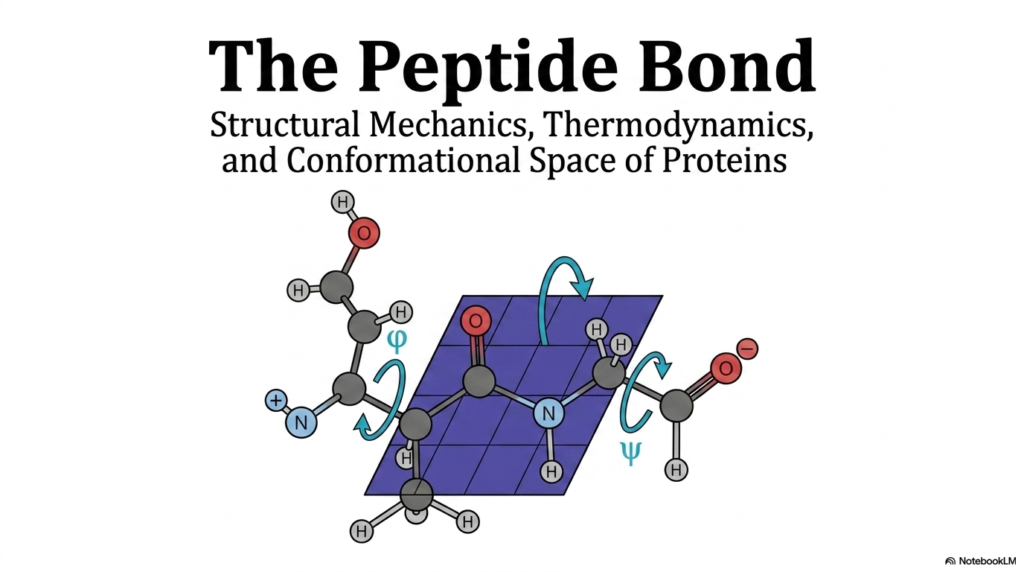
The foundation of all protein structure begins with a highly specific chemical linkage. The introductory slide visually introduces the peptide bond, highlighting its central role in linking individual amino acids into long polypeptide chains. A detailed molecular diagram showcases the specific atoms involved in this connection, specifically emphasizing the planar nature of the linkage. The illustration also introduces the crucial phi and psi dihedral angles that dictate how the backbone can twist and fold. By visualizing the peptide bond in this manner, students can instantly recognize its rigid, plate-like structure amidst the flexible joints of the larger molecule.
Understanding the physical properties of the peptide bond is universally essential for any student of advanced biochemistry. This covalent linkage is not merely a static connection; it possesses unique electronic characteristics that fundamentally restrict the molecule’s physical movement. The slide sets the stage to explore how structural mechanics and thermodynamics interact in this seemingly simple connection. These restricted movements create a defined conformational space, meaning proteins can only fold into specific, mathematically predictable shapes. The rigorous study of the peptide bond is essentially the study of how molecular constraints give rise to the boundless diversity of life.
The visual representation prominently features the characteristic repeating backbone sequence. Carbon, oxygen, nitrogen, and hydrogen atoms are arranged in a highly specific geometry that forms the primary structure of all known proteins. The slide effectively foreshadows the complex interplay of forces that will be discussed in later sections. It visually demonstrates that the peptide bond forms a rigid plane, effectively locking six atomic players into a two-dimensional arrangement. This immense rigidity forces all conformational flexibility onto the adjacent single bonds, represented by the curved arrows indicating rotation.
For medical and scientific professionals, mastering these structural mechanics provides vital insight into everything from enzyme kinetics to structural diseases. The foundational geometry established by the peptide bond dictates the entire three-dimensional architecture of the resulting protein chain. As the educational presentation progresses, these introductory concepts will be expanded upon to explain how thermodynamics and molecular kinetics govern protein synthesis and degradation. The initial visual serves as a critical anchor point for comprehending the intricate molecular choreography required to sustain biological life.
Slide 2: The Peptide Bond and Proteins as Macromolecular Workhorses
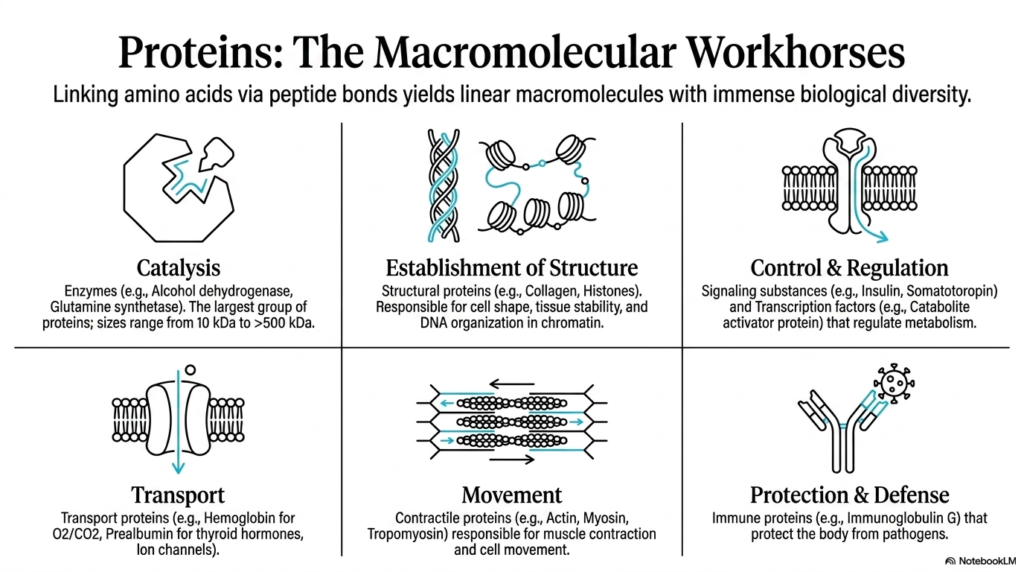
The second slide contextualizes the critical importance of these macromolecules in biological systems. By linking amino acids via peptide bonds, organisms generate linear macromolecules that exhibit an immense array of biological diversity. The presentation highlights six primary functional categories that proteins fulfill within living cellular systems. From serving as robust structural pillars to acting as dynamic signaling molecules, the versatility of these polymers is absolutely staggering. This functional diversity is entirely dependent on the structural integrity provided by the backbone linkages holding the individual amino acid building blocks firmly together.
Catalysis is one of the most vital functions, with enzymes like alcohol dehydrogenase accelerating chemical reactions that are absolutely necessary for survival. Without the stable framework provided by the peptide bond, these massive protein structures, some exceeding five hundred kilodaltons in size, would immediately unravel. Structural proteins such as collagen and histones rely on these robust chemical linkages to maintain cellular shape, stabilize delicate tissues, and organize DNA into chromatin. Furthermore, transport proteins like hemoglobin demonstrate how specific backbone folding patterns enable the precise carrying of oxygen and carbon dioxide throughout the complex human bloodstream.
The slide also details the indispensable roles of proteins in movement, control, and biological protection. Contractile proteins, including actin and myosin, form the mechanical basis for muscle contraction and cellular motility. Regulatory proteins, such as insulin and various transcription factors, delicately orchestrate complex metabolic pathways and gene expression profiles. In the realm of protection and defense, immune proteins like immunoglobulins rigorously guard the body against invading pathogens. The peptide bond is the fundamental chemical unit that makes every one of these diverse biological functions physically possible.
Understanding this immense functional diversity highlights exactly why the underlying molecular chemistry is so carefully regulated by the living cell. The linear sequence of amino acids, joined end-to-end, ultimately determines whether a newly formed polypeptide chain becomes a rigid structural fiber or a highly specific catalytic enzyme. The incredible stability of the peptide bond ensures that these molecular workhorses can operate reliably under violently fluctuating physiological conditions. This slide reinforces the idea that all high-level biological complexity arises from straightforward, repetitive chemical interactions at the microscopic level.
Slide 3: Anatomy and Nomenclature of the Peptide Bond
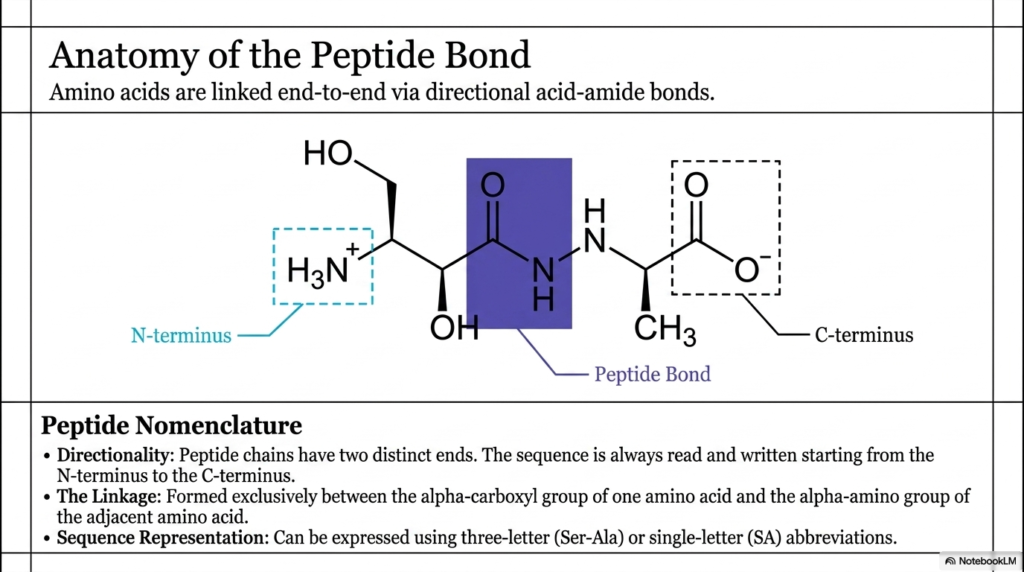
The third slide breaks down the specific molecular anatomy of this crucial biochemical linkage. Amino acids are linked end-to-end via directional acid-amide bonds, creating a distinct physical polarity across the entire macromolecule. This molecular polarity manifests as two distinct ends: an N-terminus with a free amino group and a C-terminus with a free carboxyl group. The formation of the peptide bond always occurs in a highly specific orientation, which directly dictates how the biological machinery reads and synthesizes the chain. This universal directionality is not just a chemical curiosity; it is a fundamental property that influences how the molecule ultimately folds.
A detailed molecular structure on the slide highlights the exact atomic location of the linkage. It is formed exclusively between the alpha-carboxyl group of one amino acid and the alpha-amino group of the adjacent amino acid. The shaded region on the visual emphasizes the newly formed peptide bond, illustrating the precise arrangement of carbon, oxygen, nitrogen, and hydrogen atoms. Recognizing this specific core structure is crucial for students learning to precisely identify protein backbones within larger, more complex molecular diagrams. The precise atomic connectivity underpins the rigid geometric properties that characterize the backbone.
Beyond the strict chemical structure, the slide also introduces essential nomenclature rules utilized by biochemical scientists worldwide. Because peptide chains possess distinct functional ends, the sequence of amino acids is universally read and written starting from the N-terminus and proceeding rapidly toward the C-terminus. This standard scientific convention reflects the actual biological direction of synthesis occurring within the cellular ribosome. To communicate these sequences efficiently, researchers utilize specific sequence representations. The amino acids connected by a peptide bond can be expressed using either three-letter abbreviations, like Ser-Ala, or more compact single-letter abbreviations.
Understanding these standardized naming conventions is strictly mandatory for analyzing scientific literature and utilizing massive biochemical databases. The distinct anatomical features of the backbone ensure that the resulting molecular interactions are highly specific and structurally sound. By mastering the anatomy of the peptide bond, dedicated researchers can accurately predict how different functional groups will behave when incorporated into a growing polypeptide chain. This localized atomic anatomy ultimately scales up to permanently define the macroscopic properties of the mature protein, emphasizing the importance of understanding the macromolecule at its most basic structural level.
Slide 4: Synthesis and Formation of the Peptide Bond
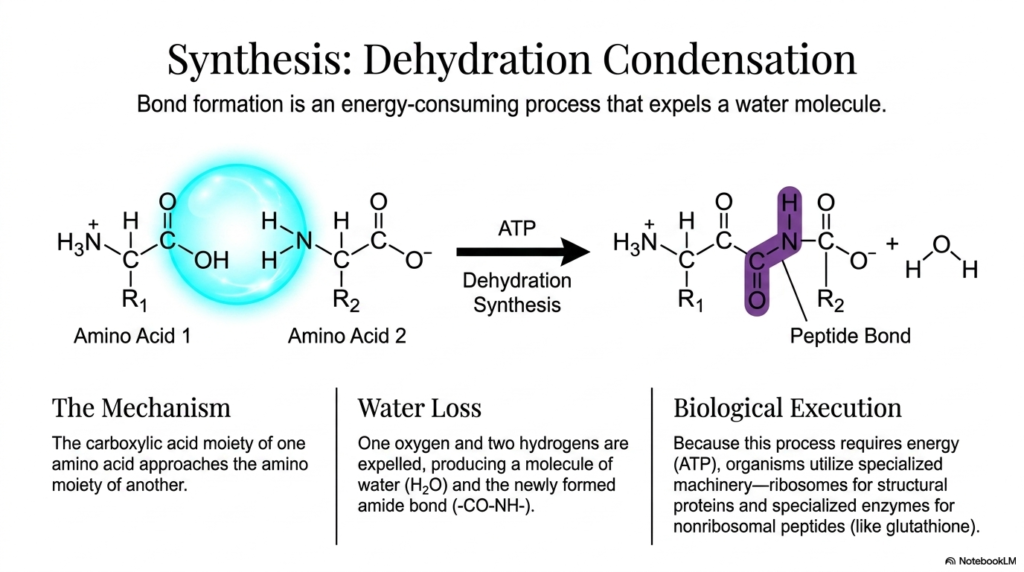
The fourth slide delves into the complex biochemical processes required to link free amino acids. The formation of a peptide bond does not occur spontaneously; it is a highly energy-intensive process known as dehydration condensation. During this specialized reaction, the carboxylate group of one amino acid approaches the amino group of another. This carefully orchestrated spatial alignment allows the chemical reaction to proceed under normal biological conditions. Understanding this precise condensation mechanism is highly vital for students exploring how molecular building blocks are assembled into functional biological polymers.
Because the formation of the peptide bond is highly thermodynamically unfavorable, the cell must invest substantial cellular energy, typically in the form of ATP, to drive the reaction forward. This energy investment facilitates the crucial dehydration step, in which one oxygen atom and two hydrogen atoms are forcibly expelled from the reacting molecules. This expelled material rapidly forms a single water molecule, leaving behind the newly synthesized amide linkage. The targeted removal of water is a classic, highly effective biochemical strategy for forcing two smaller molecules to join to form a much larger, more complex structure.
In biological systems, the execution of this specific synthesis requires specialized and massive molecular machinery. Organisms primarily use ribosomes to catalyze peptide bond formation during the translation of messenger RNA into structural and functional proteins. These massive ribosomes act as incredibly precise biological factories, bringing amino acids together and dramatically lowering the activation energy of the condensation reaction. For specialized, nonribosomal peptides like glutathione, entirely distinct enzymatic systems handle the structural assembly outside of the traditional ribosomal pathway.
The slide clearly highlights that this synthesis is fundamentally endergonic, meaning it actively absorbs energy rather than spontaneously releasing it. By expending precious ATP to forge the peptide bond, the living cell intentionally pushes the molecular system into a substantially higher energy state. This expensive investment is absolutely necessary to create the durable structural frameworks that living organisms require. The resulting chemical linkage is remarkably stable, effectively ensuring that the newly formed protein chain can withstand the harsh, fluctuating conditions of the busy cellular cytoplasm.
Slide 5: The Thermodynamic Paradox of the Peptide Bond

The fifth slide introduces a fascinating chemical contradiction widely known as the energy paradox of the peptide bond. This compelling paradox contrasts the complex thermodynamics of protein synthesis against the remarkably stable kinetics of the fully formed linkage. While forging these atomic connections requires a massive input of cellular energy, breaking them takes an astonishingly long time under standard physiological conditions. This dual nature is exactly what makes cellular proteins both incredibly expensive for the cell to produce and highly reliable once deployed. Students must thoroughly reconcile these two opposing physical forces to fully grasp protein stability.
From a strict thermodynamic perspective, forming a peptide bond is highly unfavorable and definitely endergonic. Joining two free-floating molecules into a single, heavily restricted chain drastically decreases the thermodynamic entropy of the local system. Because the entire universe naturally trends toward chaos and disorder, this synthesis requires direct coupling with ATP hydrolysis to forcefully drive the reaction forward. Conversely, the degradation or hydrolysis of the bond is highly favorable and wonderfully exergonic. Breaking the protein chain apart permanently releases between eight and sixteen kilojoules per mole of Gibbs free energy, as the natural physical laws prefer the broken, disordered state.
Despite this inherent thermodynamic preference for complete destruction, the kinetics of the peptide bond tell a completely different, highly stable story. Once properly formed, the linkage is incredibly stable due to a massive activation energy barrier that entirely prevents rapid spontaneous hydrolysis. Under standard physiological conditions, sitting at twenty-five degrees Celsius, the spontaneous half-life of a single bond ranges from three hundred and fifty to six hundred years. This remarkable kinetic stability effectively ensures that the structural integrity of biological tissues and specialized cellular machines is maintained throughout an organism’s entire operational lifespan.
To overcome these deeply inherent physical properties, biological systems rely entirely on highly specialized biological catalysts. The synthesis reaction occurs rapidly only because massive molecular machines, such as ribosomes, artificially lower the activation energy required for successful formation. In stark contrast, the rapid, necessary cleavage of a peptide bond during digestion or cellular recycling requires highly specific digestive enzymes to ensure the reaction occurs on a biologically useful timescale. This delicate, beautiful balance between thermodynamic drive and kinetic constraint is a masterpiece of evolutionary engineering that enables complex biological life to persist.
Slide 6: Resonance and the Rigid Plane of the Peptide Bond
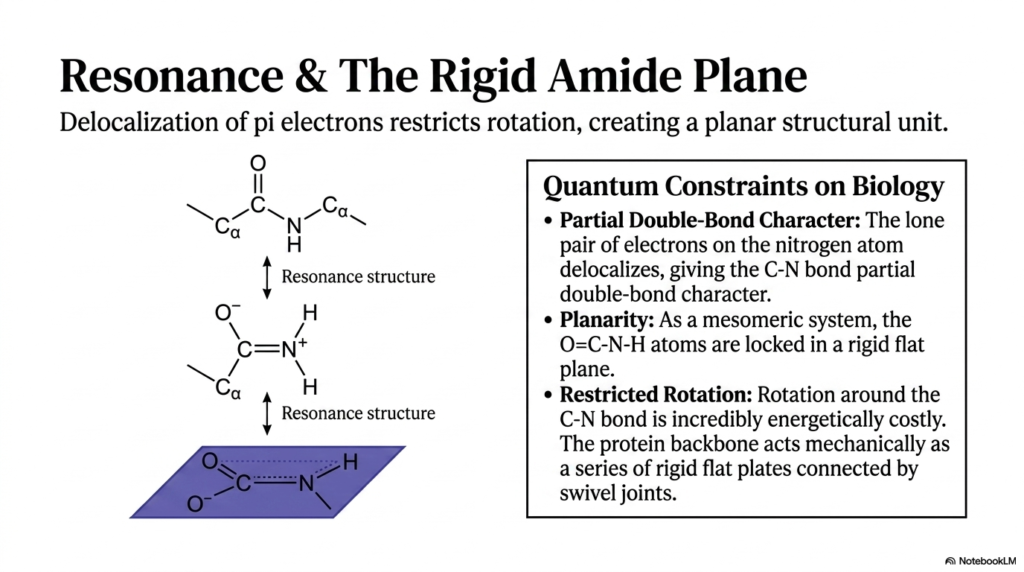
The sixth slide thoroughly explores the profound quantum-mechanical properties that define the peptide bond and severely restrict its mobility. Unlike standard single covalent connections found elsewhere in chemistry, this unique chemical linkage features the vital delocalization of pi electrons across multiple interacting atoms. A specific lone pair of electrons on the nitrogen atom closely interacts with the adjacent carbonyl carbon, creating a highly dynamic resonance structure. This electron delocalization rapidly imparts a highly important partial double-bond character to the carbon-nitrogen bond. Understanding this specific quantum phenomenon is hyper-critical, as it fundamentally dictates the physical behavior and mechanical limitations of the entire protein chain.
Because of this unique partial double-bond character, the peptide bond is fundamentally not freely rotatable like other simple chemical linkages. The electrons actively shared effectively lock the surrounding participating atoms into a rigid, perfectly flat arrangement, known as the amide plane. Specifically, the oxygen, carbon, nitrogen, and hydrogen atoms heavily involved in the linkage operate collectively as a single, inflexible mesomeric system. This molecular planarity is a strict, non-negotiable structural constraint that relentlessly forces the atoms to remain completely flat relative to one another. This geometric locking mechanism is simply the defining mechanical feature of all protein architecture.
Attempting to rotate the molecule around the rigid peptide bond by force is energetically costly and virtually impossible under standard physiological conditions. The ultimate biological consequence of this chemical restriction is massive; it permanently prevents the protein backbone from collapsing into a chaotic, useless, tangled mess. Instead, the protein backbone acts mechanically as a precise series of rigid flat plates connected intimately by specialized swivel joints. The atoms permanently trapped within the amide plane provide a remarkably stable foundation, effectively ensuring that the three-dimensional structure of the folded macromolecule remains consistently reliable over long periods.
By actively visualizing the backbone as a repeating series of rigid planes, biochemistry students can more accurately predict how a protein will behave in an aqueous solution. The detailed resonance structures beautifully shown on the slide demonstrate that the actual physical state of the molecule exists perfectly somewhere between a single and a double bond. This highly delicate quantum balance represents a fundamental, unbreakable constraint on biology. The sheer chemical rigidity of the peptide bond is undeniably the primary reason structural proteins can form highly specific, durable structures like enzymes, receptors, and muscle fibers without randomly unraveling into useless strings.
Slide 7: Cis and Trans Isomerism Around the Peptide Bond
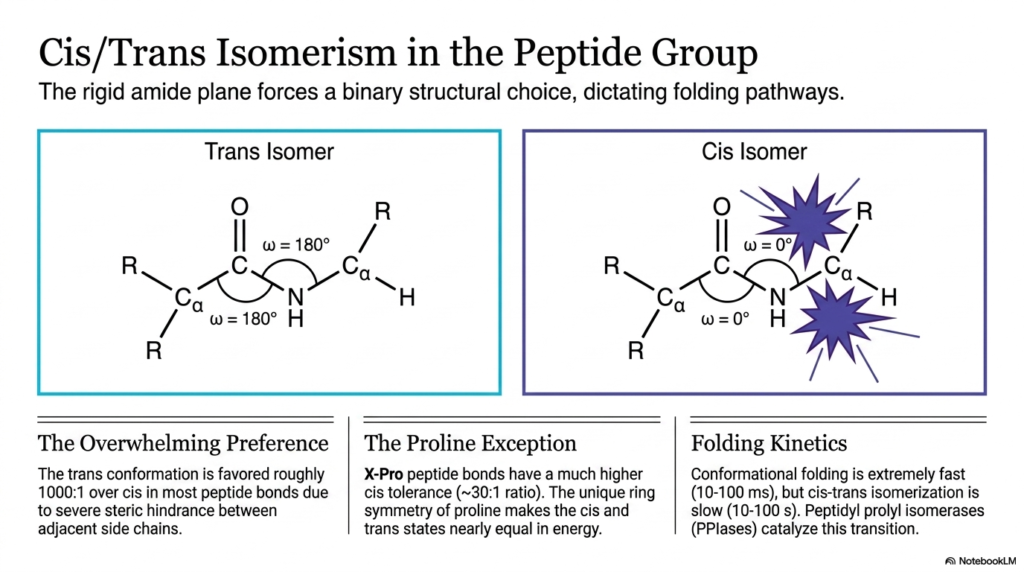
The seventh slide focuses intently on the specific geometric orientations imposed on the molecule by the rigid peptide bond. Because the stable amide plane severely restricts rotation, the adjacent atoms must adopt a binary choice, orienting themselves in either a cis or a trans configuration. This strict geometric locking mechanism fundamentally and permanently dictates the available folding pathways for the entire resulting macromolecule. Students exploring complex protein folding must fully understand how this binary choice profoundly influences the overall three-dimensional shape and absolute stability of the mature protein.
In the vast majority of biological proteins, the trans isomer is overwhelmingly and consistently preferred by the folding molecule. In a trans configuration, the extremely bulky side chains of the adjacent interacting amino acids are heavily positioned on opposite sides of the rigid peptide bond. This specific spatial arrangement drastically minimizes steric hindrance, effectively preventing the large electron clouds of the chemical groups from physically clashing. Due to these massive energetic advantages, the trans conformation is naturally favored roughly one thousand to one over the cis conformation in standard amino acid linkages. This overwhelming mathematical preference powerfully ensures a predictable and highly stable zigzag pattern along the protein backbone.
However, a highly significant biological exception exists exclusively when the specific amino acid proline is directly involved in the linkage. When any standard amino acid directly precedes proline, the resulting newly formed peptide bond exhibits a much higher structural tolerance for the normally forbidden cis conformation. Because proline features a unique cyclic ring structure that directly bonds to its own backbone nitrogen, the cis and trans states mysteriously become nearly equal in overall energy. This unique chemical symmetry shockingly results in a much narrower biological preference ratio of approximately 30:1, making cis-proline linkages a crucial regulatory feature in specific protein folds.
The physical transition occurring between these two distinct isomeric states has a massively profound impact on the kinetics of cellular protein folding. While a polypeptide chain can rapidly navigate its overall conformational folding, physically isomerizing a peptide bond from cis to trans is an incredibly sluggish process. This extremely slow rotation can dangerously trap a folding protein in an entirely inactive state for up to 100 seconds. To forcefully accelerate this transition and ensure rapid biological function, living cells deploy highly specialized enzymes called peptidyl prolyl isomerases to aggressively catalyze the switch and rapidly finalize the protein’s functional architecture.
Slide 8: Conformational Space Beyond the Peptide Bond
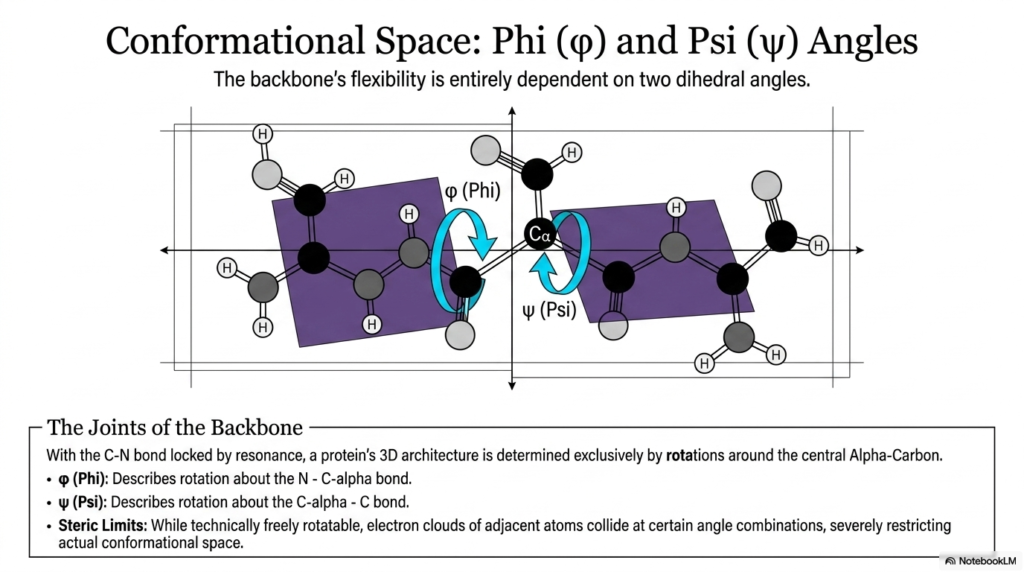
The eighth slide rapidly zooms in on the highly specific molecular joints that enable the necessary biological flexibility in the protein backbone. While the peptide bond itself is completely and rigidly locked by quantum resonance, the adjacent single bonds readily provide the absolutely necessary rotational freedom for the macromolecule to fold into a functional shape. The molecule’s final three-dimensional architecture is ultimately determined entirely by simple rotations occurring around the central alpha-carbon atom. The presentation heavily introduces the phi and psi dihedral angles, which precisely and mathematically describe the specific torsion of these flexible joints. Mastering these angles is essential for accurately visualizing the conformational space available to a twisting protein chain.
The phi angle specifically describes the physical rotation about the exact bond connecting the rigid backbone nitrogen to the central alpha-carbon. Conversely, the psi angle carefully describes the rotation about the specific bond connecting the central alpha-carbon to the highly adjacent carbonyl carbon. These two highly dynamic swiveling joints sit firmly on either side of the inflexible peptide bond, operating much like well-oiled hinges on a solid wooden door. By carefully adjusting these two precise angles in a highly specific sequence along the entire polymer chain, the resulting biological protein can fold into an almost infinite variety of complex, highly functional three-dimensional shapes.
While these unique bonds are technically freely rotatable from a purely chemical standpoint, severe and absolute physical restrictions exist. As the phi and psi angles dynamically twist, the incredibly bulky electron clouds of adjacent atoms can easily and dangerously swing into one another. These harsh physical collisions instantly result in extreme steric hindrance, which severely and permanently restricts the actual geometric conformational space the molecule can ever adopt. Because the peptide bond permanently and rigidly locks six specific atoms into a completely flat plane, the surrounding functional groups ironically have very little room to maneuver without clashing, thereby massively narrowing the possible geometries the backbone can successfully achieve.
This highly delicate interplay occurring between strict rigidity and necessary flexibility represents the absolute core mechanical principle of structural biology. The fixed, unwavering geometry of the peptide bond ensures basic structural stability, while the phi and psi angles happily provide the targeted, precise flexibility necessary to create binding pockets and highly catalytic active sites. Without these specific rotational geometric parameters, biological proteins would either be too rigid to function or too floppy to maintain a stable biological shape. Understanding these specific dihedral angles is undeniably the very first step toward accurately predicting how a linear sequence of molecules will ultimately fold into a flawless functional machine.
Slide 9: The Ramachandran Plot and Peptide Bond Geometry
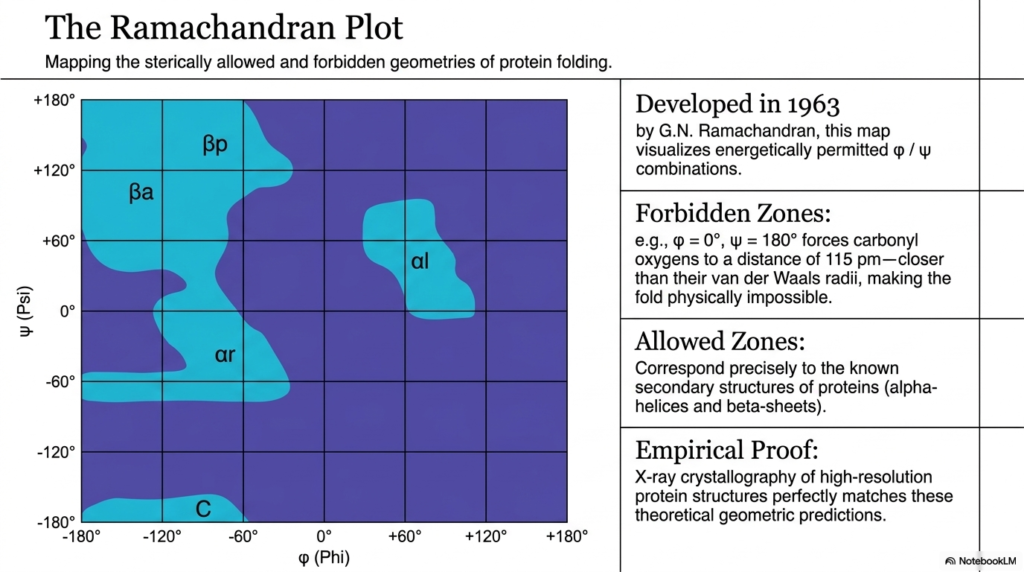
The ninth slide masterfully introduces one of the most vital visualization tools in modern structural biology: the Ramachandran plot. Initially developed in nineteen sixty-three, this revolutionary graphical map brilliantly visualizes the sterically allowed and completely forbidden geometries of protein folding. By successfully plotting the phi angles directly on the x-axis and the psi angles on the y-axis, structural researchers can instantly and reliably identify exactly which rotational combinations are physically possible in nature. The plot effectively translates the highly rigid mechanical restrictions heavily imposed by the stable peptide bond into a wildly clear, easy-to-read two-dimensional diagram that flawlessly dictates the rules of molecular architecture.
The intensely dark blue regions of the scientific graph represent strictly forbidden zones where catastrophic atomic collisions render folding mathematically impossible. For example, if both the phi and psi angles are forcefully pushed to certain extreme orientations, the rigid carbonyl oxygens are violently pushed much closer than their allowed van der Waals radii. The relentlessly inflexible nature of the peptide bond utterly guarantees that these physical, destructive clashes cannot ever be avoided by simply bending the local backbone. As a direct result, the protein chain is strictly prohibited from entering these specific mathematical coordinates, drastically limiting the total number of ways a biological polymer can fold.
Conversely, the brightly shaded areas represent the energetically allowed zones where the distinct atoms fit together seamlessly without ever experiencing negative steric clashes. These highly energetic permitted zones correspond precisely and miraculously to the specific known secondary structures famously found in nature, such as spiraling alpha-helices and flat beta-sheets. The rigid, unyielding geometry of the peptide bond practically forces the entire polypeptide chain to adopt these specific, highly stable motifs. This mathematically means that the beautiful, wildly repeating coils and flat sheets frequently seen in large protein structures are absolutely not random occurrences; they are strict mathematical inevitabilities driven entirely by the spatial limitations of the molecule’s chemical joints.
The brilliant theoretical geometric predictions painstakingly mapped out by this historic plot have been rigorously and completely confirmed through endless empirical evidence. High-resolution x-ray crystallography of massive, highly complex protein structures perfectly and consistently matches the narrow allowed zones accurately predicted decades ago. The structural boundaries established by the peptide bond are remarkably rigid and universally applicable across virtually all known forms of biological life. For dedicated students entering the medical or biochemical fields, the famous Ramachandran plot serves as an indispensable, permanent reference for understanding how atomic-level steric constraints govern the macroscopic shapes and biological functions of all known cellular proteins.
Slide 10: Structural Extremes Affecting the Peptide Bond: Glycine vs. Proline
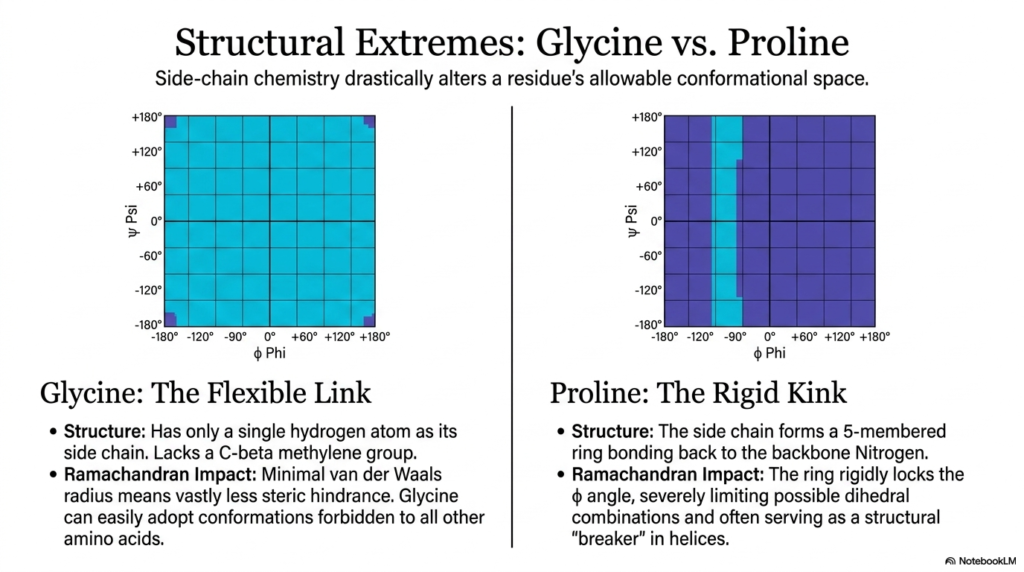
The tenth slide explores how specific amino acid side chains can drastically alter a single residue’s allowable conformational space. While the peptide bond consistently maintains a highly consistent, rigid geometry along the entire backbone, the unique chemical structures of glycine and proline readily create fascinating, biologically crucial structural extremes. These two highly specific residues aggressively bend the standard, universal rules of protein folding, acting as heavily crucial structural modifiers within the massive macromolecule. By actively comparing their respective unique Ramachandran plots, ambitious students can precisely visualize how specific side-chain chemistry significantly influences the rotational freedom of adjacent molecular joints.
Glycine brilliantly acts as the ultimate, highly flexible structural link within a massive protein chain. Unlike literally all other standard amino acids, it completely lacks a bulky, heavy carbon-beta methylene group, possessing only a single, incredibly tiny hydrogen atom as its designated side chain. Because of this incredibly small van der Waals radius, glycine shockingly experiences almost no negative steric hindrance as it dynamically rotates in space. This severe lack of steric hindrance readily allows glycine to adopt highly twisted conformations that are strictly and permanently forbidden to all other residues. Consequently, glycine is frequently found in tight turns and highly flexible loops, where the peptide bond must dynamically navigate sharp, extreme changes in direction.
At the opposite end of the structural spectrum, proline sits, permanently serving as the most rigid residue in biology. Proline’s highly unique side chain aggressively curves backward to form a solid five-membered ring that covalently bonds directly back to the backbone nitrogen atom. This unique cyclic architecture physically and permanently locks the local phi angle in place, severely limiting the possible dihedral combinations the specific residue can adopt. Because its structural rotation is severely restricted, proline forces the adjacent peptide bond into a highly specific orientation, thereby severely limiting the conformational freedom of the surrounding protein chain.
Due to its extreme rigidity, proline frequently serves as a major structural breaker within delicate secondary structures like spiraling alpha-helices. It predictably introduces a harsh, permanent kink into the protein backbone that cannot be straightened. The stark visual contrast between the absolute structural flexibility of glycine and the complete rigidity of proline highlights the highly dynamic nature of overall protein architecture. The fascinating biological interaction between the completely unyielding peptide bond and these highly specialized side chains enables biological organisms to precisely fine-tune the mechanical properties of their cellular machines, creating incredible structures ranging from highly elastic fibers to rock-hard enzymatic cores.
Slide 11: Degradation and Cleavage of the Peptide Bond
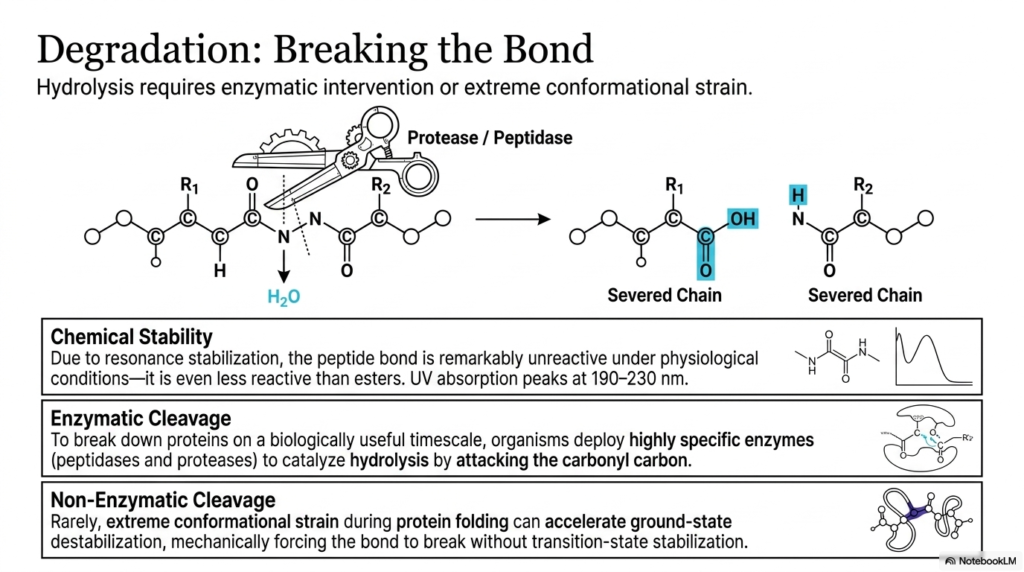
The eleventh slide thoroughly investigates the precise biological and chemical mechanisms absolutely required for the successful degradation and complete cleavage of the backbone. Because the peptide bond is remarkably unreactive and highly stable under normal physiological conditions, simply passively waiting for spontaneous hydrolysis is absolutely not biologically viable. Due to massive, profound resonance stabilization, the specific linkage is incredibly mechanically stable, repeatedly proving to be even less chemically reactive than standard ester bonds. To rapidly recycle amino acids or quickly activate specific, vital signaling cascades, living organisms must intentionally deploy highly targeted strategies to violently overcome this immense chemical stability and successfully sever the chain.
To forcefully break down massive proteins on a biologically useful and rapid timescale, cells actively utilize highly specific, deadly enzymes known universally as proteases and peptidases. These molecular scissors are expertly designed to aggressively catalyze hydrolysis by directly and efficiently attacking the resilient carbonyl carbon in the tight linkage. The slide visually depicts this precise enzymatic intervention, brilliantly demonstrating how the forceful addition of a water molecule ultimately severs the tough backbone, cleanly yielding two separate, much shorter molecular chains. Without these highly targeted biological catalysts, the incredibly stable peptide bond would remain completely intact for centuries, halting the essential biological processes of cellular turnover and rapid digestion.
The detailed slide also brilliantly highlights the highly specific chemical properties that ultimately make this enzymatic cleavage incredibly difficult to perform. The massive resonance stabilization that uniquely creates the rigid amide plane also actively distributes the local electron density in a precise way that highly repels outside chemical attacks. This unique electronic configuration can be readily measured in a laboratory setting, as the peptide bond exhibits distinct ultraviolet absorption peaks comfortably situated between 190 and 230 nanometers. This incredibly robust chemical profile ensures that vital structural proteins remain intact even when subjected to extremely harsh, rapidly fluctuating environments within the human body.
While targeted enzymatic cleavage is undeniably the primary biological mechanism heavily used for degradation, rare non-enzymatic cleavage can surprisingly occur under highly specific, extreme conditions. Very rarely, massive, extreme conformational strain occurring during the folding process can physically and aggressively accelerate dangerous ground-state destabilization. In these incredibly unique cases, the massive protein mechanically twists the peptide bond to such an extreme and violent degree that it forces the linkage to break structurally without traditional, safe transition-state stabilization. Thoroughly understanding both the extreme stability of this crucial bond and the highly specialized mechanisms strictly required to brutally break it is deeply essential for dedicated students rapidly studying pharmacology, digestion, and the targeted drug design of protease inhibitors.
Slide 12: Synthesizing the Big Picture: From the Peptide Bond to 3D Structure
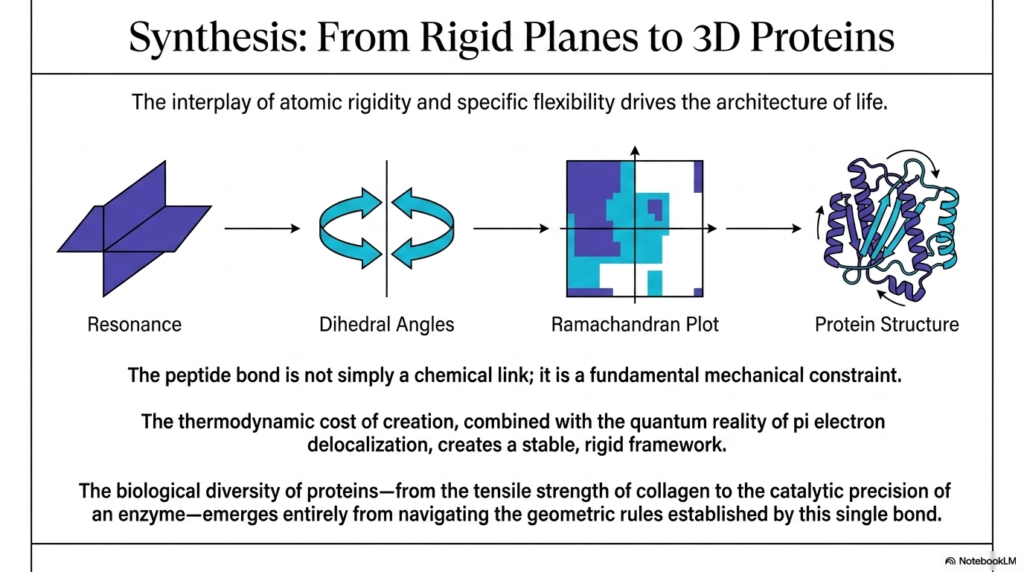
The twelfth and absolutely final slide gracefully serves as a comprehensive, highly detailed summary, brilliantly synthesizing how highly localized atomic properties dictate massive macroscopic biological structures. The educational presentation beautifully illustrates a crystal-clear mechanical pipeline, brilliantly demonstrating exactly how the fundamental structural physics of the peptide bond scales massively to flawlessly create highly complex, three-dimensional proteins. It smoothly begins with the deep quantum reality of pi-electron delocalization, which uniquely generates a highly rigid molecular plane. This initial, unbreakable rigidity directly and strictly governs the physical behavior of the adjacent dynamic dihedral angles, which subsequently permanently define the permitted geometries, as visualized on the Ramachandran plot, ultimately dictating the final folded architecture.
The core scientific message of this presentation is that the peptide bond is not simply a passive, boring chemical link. Instead, it powerfully acts as a massive, fundamental mechanical constraint that aggressively forces the entire molecular chain to rapidly adopt highly specific, highly ordered physical configurations. The dynamic interplay between the rigid, flat atomic planes and the highly specific flexibility of the swiveling dihedral joints drives the incredible architecture of life. Without the incredibly strict geometric rules vigorously enforced by the peptide bond, all biological proteins would tragically lack the absolute structural integrity intensely required to rapidly perform their incredibly diverse range of biochemical functions.
The incredibly high thermodynamic cost of creating this highly specific, rigid linkage is entirely and easily justified by the massive resulting structural payoff. By actively investing precious cellular energy to forcibly forge these highly stable atomic connections, the biological system guarantees that the resulting macromolecule will maintain a highly consistent, durable physical shape. The remarkable mechanical stability of the peptide bond enables delicate proteins to efficiently withstand extreme physical stress and survive incredibly harsh chemical environments. This incredibly robust, reliable framework is exactly what allows structural molecules to firmly weave together into immensely strong fibers and enables highly delicate enzymes to flawlessly maintain the microscopic precision required for advanced chemical catalysis.
Ultimately, the absolutely incredible biological diversity observed in functional proteins emerges almost entirely from seamlessly navigating the strict geometric rules established by this single, foundational connection. From the practically incredible tensile strength found within massive collagen fibers sitting in human tendons to the intense microscopic precision of highly active digestive enzymes, absolutely every functional protein totally relies on this specific molecular stability. By actively and rigorously studying the complex quantum, structural, and thermodynamic realities surrounding the peptide bond, future biochemistry students absolutely gain a deeply profound appreciation for the beautifully elegant, highly physical logic that perfectly underpins all known biological life on this entire planet, literally.
Please read our Content Disclaimer Statement.
Check out our social media channels: