31. Activated Metabolites: The Currency of Cellular Energy
Imagine a sprawling, bustling metropolis trying to function without a standardized currency; complete chaos would inevitably ensue. Similarly, a living biological cell requires a reliable, universal energy currency to fund its continuous, energy-demanding operations and biosynthetic pathways. This presentation explores the elegant biochemical solutions that evolution has crafted to sustain life at the molecular level. The core purpose of this slide deck is to thoroughly demystify the complex structures and thermodynamic pathways of cellular energy carriers.
Slide 1: Introduction to Activated Metabolites
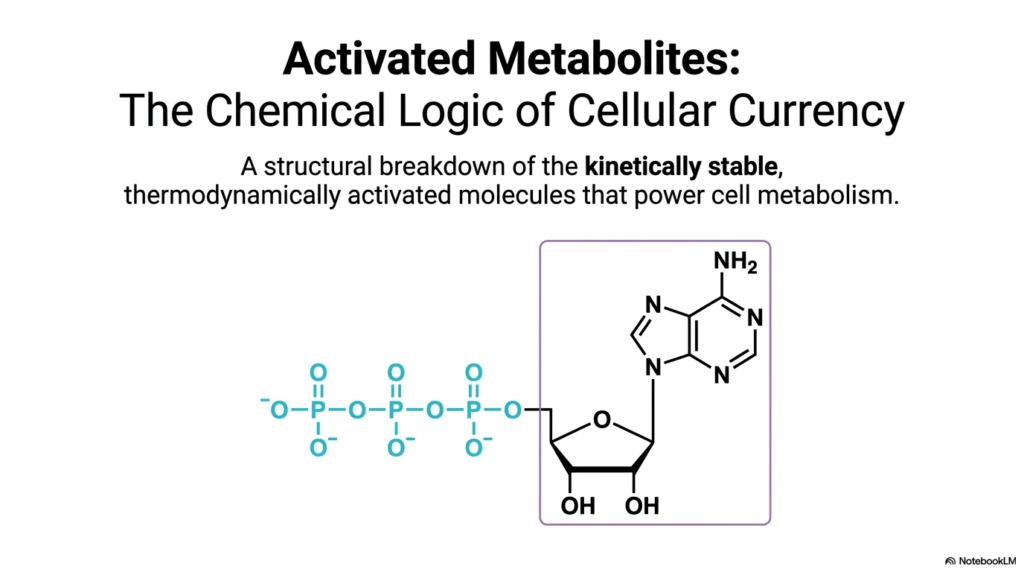
The cellular environment is an intricate ecosystem that requires a constant, regulated influx of energy to maintain order and drive complex, thermodynamically unfavorable biological processes. At the absolute heart of this energetic, bustling area are activated metabolites. These highly specialized molecules serve as the fundamental cellular currency, providing the energy required to fuel essential biosynthetic reactions. Understanding the nature of activated metabolites is absolutely essential for any student seeking to master the profound chemical logic that underpins all known cellular life.
This introductory slide visualizes the classic structure of ATP, the most ubiquitous of these energy carriers, highlighting the unique atomic arrangements that are crucial for energy storage. By examining the exact chemical architecture of these molecules, one can begin to deeply appreciate how their specific functional groups are perfectly adapted for precise biological roles. These biochemical structures are not simply random collections of carbon and phosphorus; they are highly refined, molecular machines sculpted by billions of years of evolutionary pressure.
To fully grasp the biochemical power of activated metabolites, one must first understand their unique and defining dual nature. They act as kinetically stable yet thermodynamically activated molecules that are exclusively responsible for powering cell metabolism. This defining trait means they do not randomly break apart, releasing their energy prematurely in the cell. Instead, they patiently wait to be acted upon by highly specific cellular enzymes, thereby unleashing their significant, life-sustaining power.
Slide 2: The Metabolic Paradox of Activated Metabolites
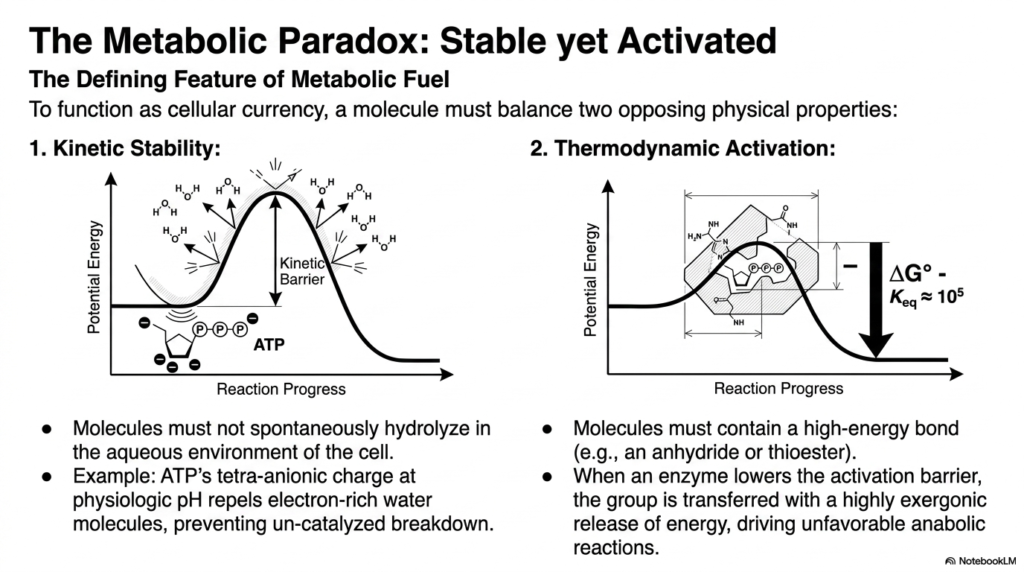
A fundamental feature of metabolic fuel is a fascinating biochemical contradiction, formally known as the metabolic paradox. To function effectively as a reliable cellular currency, activated metabolites must perfectly balance two entirely opposing physical and chemical properties. First, they require high kinetic stability to prevent spontaneous hydrolysis in the cytoplasm’s water-rich environment. If cellular energy carriers simply broke down on their own, biological organisms would quickly waste their energy reserves.
The classic textbook example of this critical kinetic stability is seen in the three-dimensional structure of ATP, the most famous of the activated metabolites. At normal physiological pH levels, the ATP molecule carries a dense tetra-anionic charge. This significant, concentrated negative charge actively repels the electron-rich oxygen atoms of surrounding water molecules, effectively preventing uncatalyzed, spontaneous breakdown. This built-in kinetic barrier ensures the stored chemical energy remains safely locked away.
Conversely, the second required physical property for activated metabolites is massive thermodynamic activation. These specialized molecules must contain a high-energy chemical bond, such as an unstable phosphoric anhydride or a reactive thioester linkage. When a specific cellular enzyme lowers the structural activation barrier, the energetic chemical group is transferred with a highly exergonic, explosive release of energy. This vital thermodynamic release is what actually drives stubbornly unfavorable anabolic biological reactions forward.
Slide 3: The Toolkit of Eight Essential Activated Metabolites
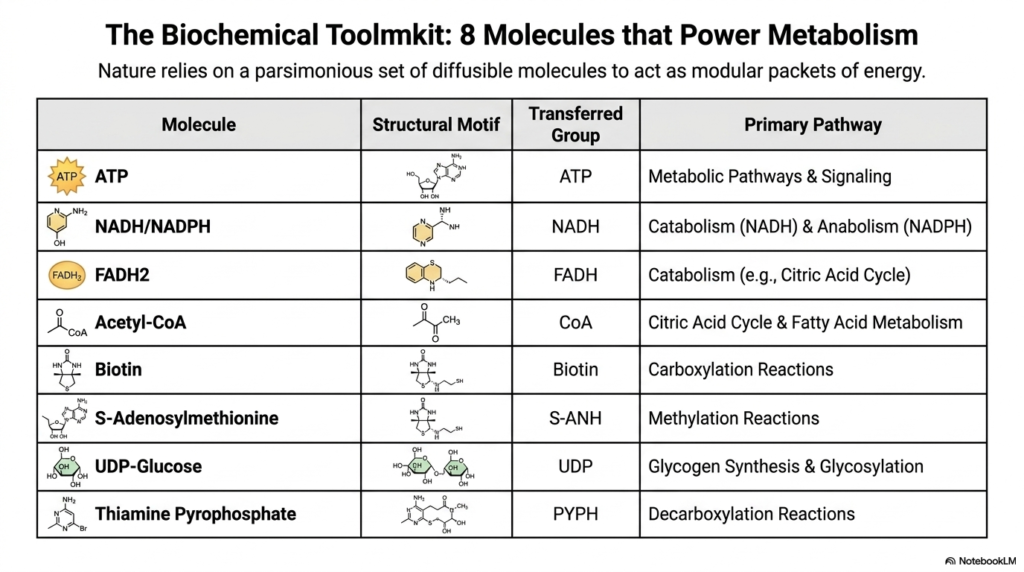
Nature is incredibly efficient and relies heavily on a parsimonious, highly conserved set of diffusible molecules that act as modular packets of energy. This slide introduces the comprehensive biochemical toolkit, detailing the eight primary activated metabolites that are universally required to power cellular metabolism. Rather than creating a unique energy source for every single metabolic reaction, the cell relies on this versatile, standardized group of chemical carriers to sustain its diverse biological operations.
Among these critical activated metabolites, ATP stands out as the primary energy currency for generalized metabolic pathways and crucial cellular signaling. However, the toolkit also includes specialized electron carriers like NADH, NADPH, and FADH2, which are absolutely vital for driving both catabolic breakdown and anabolic biosynthesis. By compartmentalizing different types of energetic transfers into distinct modular molecules, the biological cell maintains tight, sophisticated control over its complex metabolic networks.
The remaining molecules in this group of activated metabolites serve highly specialized chemical roles that are equally vital for survival. Acetyl-CoA acts as the master vehicle for transferring two-carbon fragments during the citric acid cycle. Meanwhile, Biotin handles critical carboxylation reactions, and S-Adenosylmethionine manages precise methylation events. Rounding out the toolkit, UDP-Glucose drives glycogen synthesis, and Thiamine Pyrophosphate powers essential decarboxylation reactions, completing the elegant suite of metabolic tools.
Slide 4: ATP as the Universal Donor Among Activated Metabolites
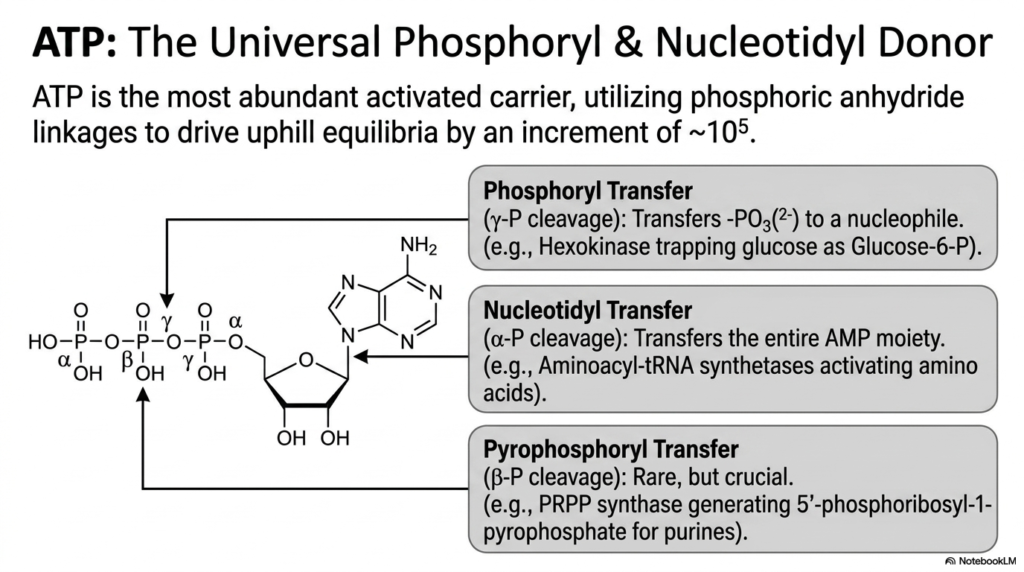
Adenosine triphosphate, universally known as ATP, is by far the most abundant and well-recognized of the activated metabolites within the biological cell. It acts as the universal phosphoryl and nucleotidyl donor, utilizing its famously unstable phosphoric anhydride linkages to drive uphill chemical equilibria by an astonishing increment of ten to the fifth power. This massive, energetic leverage is what enables cells to synthesize complex biomolecules from simple precursors.
The true versatility of ATP, a premier activated metabolite, lies in its ability to undergo three distinct types of chemical cleavage. The most common mechanism is phosphoryl transfer, where the terminal gamma-phosphate is cleaved and transferred to a nucleophile. A classic example of this is the enzyme hexokinase, which efficiently traps glucose inside the cell by rapidly converting it into glucose-6-phosphate during the first step of glycolysis.
Beyond simple phosphoryl transfer, these activated metabolites can also provide more complex energy contributions. Nucleotidyl transfer involves the precise cleavage of the alpha-phosphate, thereby transferring the entire AMP moiety to a target, as seen when aminoacyl-tRNA synthetases activate amino acids for protein synthesis. Although less common, pyrophosphoryl transfer involves beta-phosphate cleavage, which is absolutely crucial for generating the highly reactive foundations necessary for complex purine nucleotide synthesis.
Slide 5: Electron Carriers as Activated Metabolites
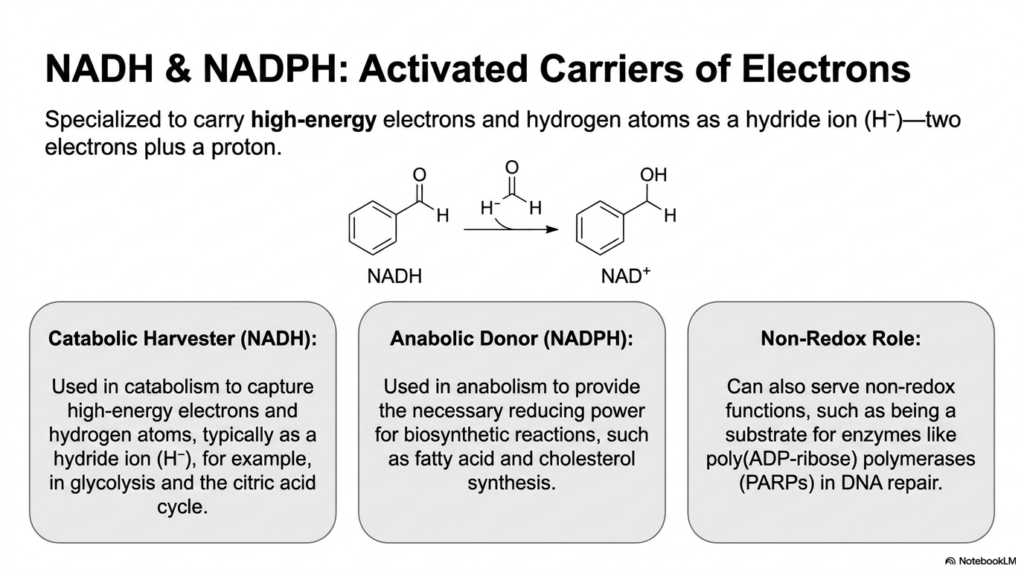
While ATP primarily handles phosphate transfers, NADH and NADPH represent a different, critically important class of activated metabolites perfectly specialized for redox reactions. These molecules are uniquely designed to carry high-energy electrons and hydrogen atoms, specifically packaged as a reactive hydride ion containing two electrons and one proton. This unique chemical configuration allows them to efficiently shuttle reducing power throughout the bustling, highly active cellular environment.
In the realm of these specialized activated metabolites, there is a strict, elegant division of metabolic labor. NADH acts predominantly as the catabolic harvester, explicitly used to capture high-energy electrons stripped away during the breakdown of molecular fuels. This energy harvesting typically occurs during the core reactions of glycolysis and the citric acid cycle, where the resulting NADH is then fed into the electron transport chain to produce large amounts of ATP.
Conversely, its closely related structural cousin, NADPH, functions primarily as the anabolic donor within the cellular pool of activated metabolites. It provides the heavy reducing power absolutely necessary to drive energetically expensive biosynthetic reactions, such as the elaborate construction of fatty acids and cholesterol. Fascinatingly, these molecules also play critical non-redox biological roles, serving as direct chemical substrates for crucial DNA repair enzymes such as poly(ADP-ribose) polymerases.
Slide 6: Acetyl-CoA and the Thioester-Activated Metabolites
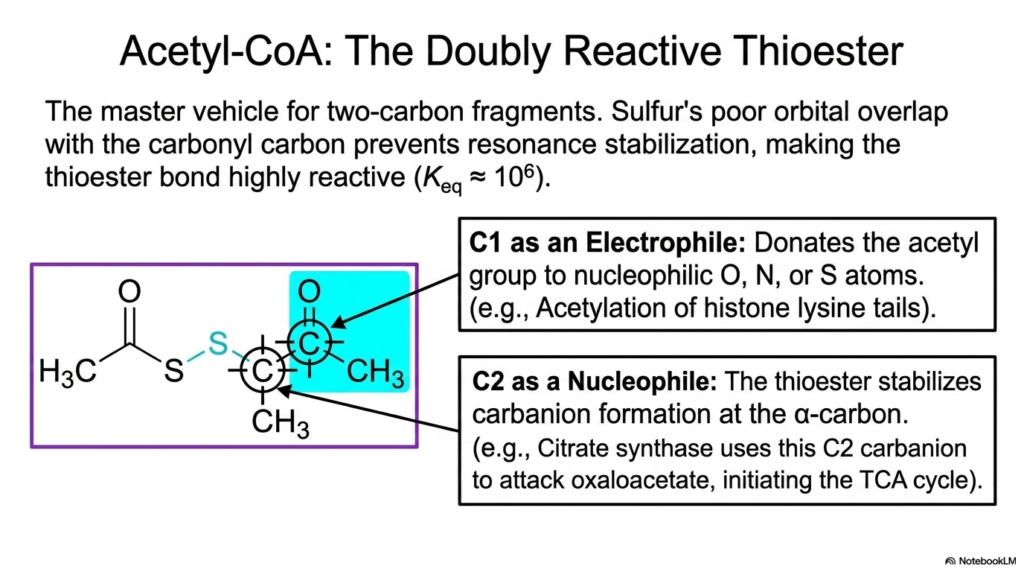
Acetyl-CoA holds a highly prestigious position among activated metabolites as the master molecular vehicle for transporting and reacting two-carbon fragments. The secret to its immense biochemical power lies entirely in its highly reactive thioester bond. Because sulfur atoms exhibit notoriously poor orbital overlap with the adjacent carbonyl carbon, stabilizing resonance cannot occur, making the resulting chemical bond extremely high in energy and exceptionally reactive.
This extreme chemical reactivity gives these specific activated metabolites a unique, biologically vital dual personality. At the C1 position, Acetyl-CoA acts as a powerful electrophile, aggressively donating its acetyl group to various nucleophilic oxygen, nitrogen, or sulfur atoms. A prime biological example of this electrophilic mechanism is the precise acetylation of histone lysine tails, a foundational biochemical process that actively regulates and controls the expression of entire genomic sequences.
Simultaneously, the unique thioester linkage of these activated metabolites allows the C2 position to act as an incredibly effective nucleophile. The sulfur-containing bond chemically stabilizes the formation of a highly reactive carbanion at the alpha-carbon. This specific nucleophilic property is exploited brilliantly by the enzyme citrate synthase, which uses this exact C2 carbanion to aggressively attack oxaloacetate, effectively initiating the vital energy-producing machinery of the central citric acid cycle.
Slide 7: SAM and the Methylating Activated Metabolites
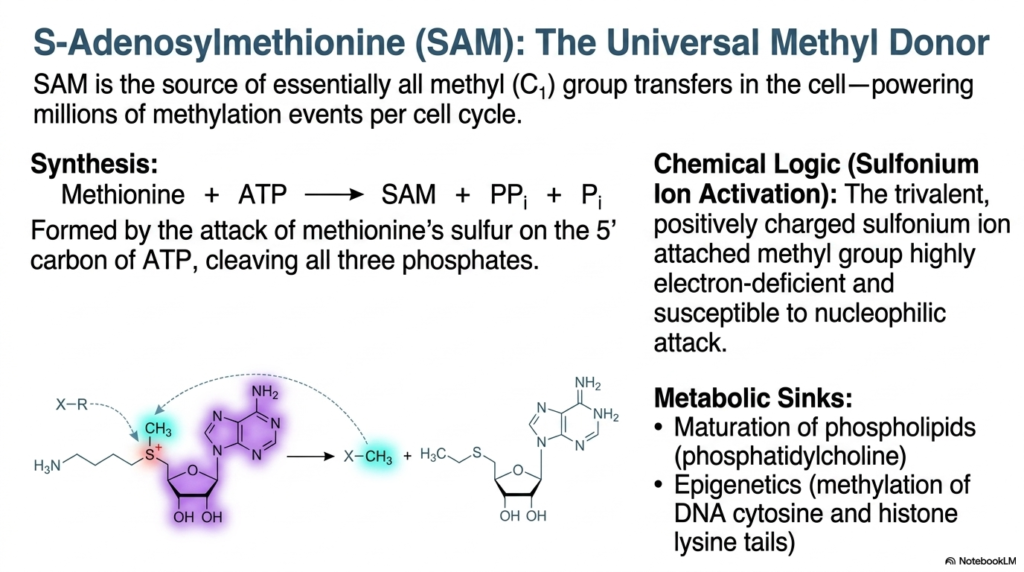
S-Adenosylmethionine, commonly abbreviated as SAM, is universally recognized as the premier methyl donor among all cellular activated metabolites. It serves as the primary chemical source for essentially all one-carbon methyl group transfers occurring within the biological cell. This immense responsibility means that SAM actively powers millions of individual critical methylation events during every successful cellular division cycle, underscoring its absolute necessity for biological life.
The synthesis of these specific activated metabolites requires an immense energetic investment. SAM is formed through a highly unusual biochemical reaction in which the sulfur atom of the amino acid methionine aggressively attacks the 5′-phosphate group of a fresh ATP molecule. This dramatic chemical event results in the complete cleavage and loss of all three ATP phosphates, showcasing just how much thermodynamic energy is required to successfully prime this essential methyl carrier.
The true chemical logic behind these activated metabolites relies on the activation of highly reactive sulfonium ions. The trivalent, positively charged sulfonium ion makes the attached methyl group highly electron-deficient and incredibly susceptible to nucleophilic attack. This rapid transfer mechanism supplies crucial metabolic sinks, actively driving the maturation of membrane phospholipids such as phosphatidylcholine and fundamentally regulating complex epigenetics through the targeted methylation of cytosine bases in DNA and essential histone proteins.
Slide 8: UDP-Glucose and Glycosylation Activated Metabolites
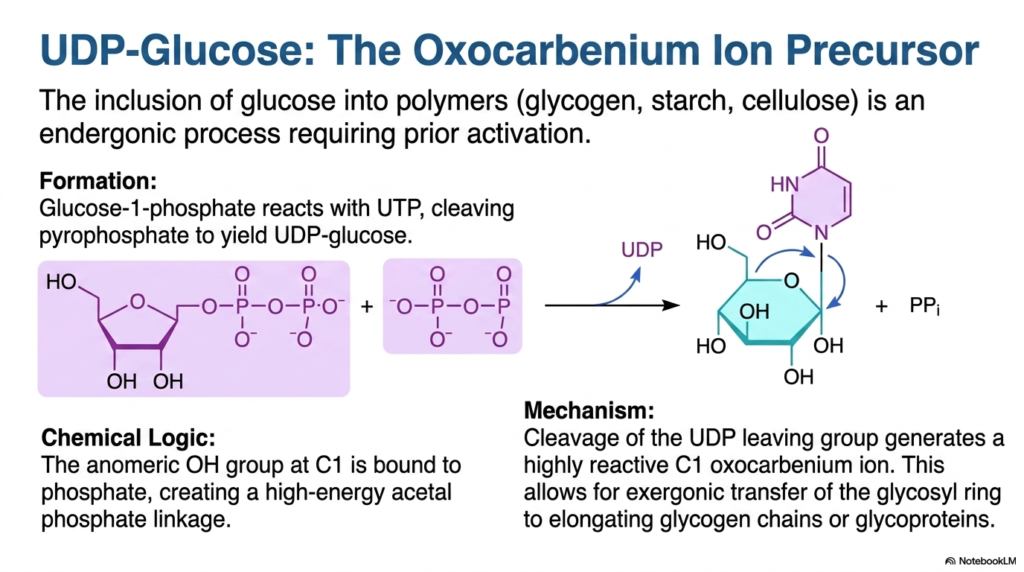
The biological construction of complex carbohydrate polymers such as glycogen, starch, and cellulose is a highly endergonic, thermodynamically unfavorable process that requires prior chemical activation. To overcome this massive energy barrier, the cell relies heavily on UDP-Glucose, a highly specialized activated metabolite. This unique molecule serves as the essential oxocarbenium-ion precursor, providing the necessary energetic drive to weave simple sugars into massive structural and storage polymers.
The formation of these specific activated metabolites begins when standard Glucose-1-phosphate aggressively reacts with UTP. This enzymatic reaction cleaves a leaving pyrophosphate group, yielding the high-energy UDP-glucose molecule. The underlying chemical logic relies on the fact that the anomeric hydroxyl group at the C1 position becomes tightly bound to a phosphate, artificially creating an incredibly unstable, high-energy acetal phosphate linkage that is primed and completely ready for rapid biosynthetic action.
When it is time for these activated metabolites to perform their biological duty, the precise cleavage of the massive UDP leaving group rapidly generates a highly reactive C1 oxocarbenium intermediate ion. This incredibly unstable structural state allows for the highly exergonic, thermodynamically favorable transfer of the entire glycosyl ring. This transfer aggressively elongates growing glycogen chains or actively decorates glycoproteins destined for the outer cellular membrane.
Slide 9: Nitrogen Processing and Activated Metabolites
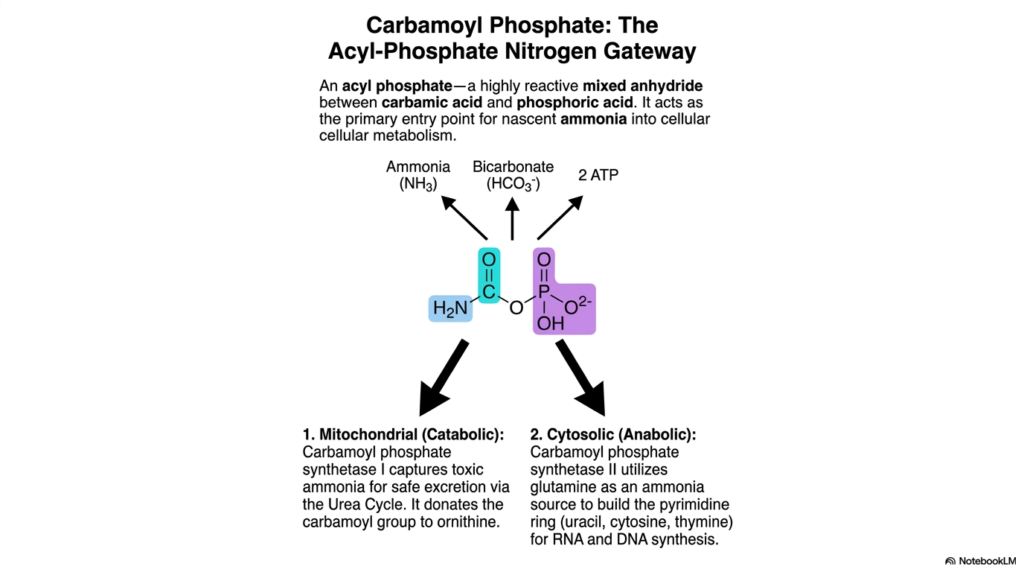
Carbamoyl Phosphate acts as the absolutely essential acyl-phosphate nitrogen gateway for the biological cell. Among the various activated metabolites, this highly reactive molecule represents a fascinating structural hybrid—a mixed chemical anhydride formed specifically between carbamic acid and phosphoric acid. It serves as the absolutely primary biological entry point for capturing nascent, highly toxic ammonia and safely funneling it directly into mainstream cellular nitrogen metabolism.
The biological utility of these specific activated metabolites is strictly divided between two distinct, highly compartmentalized cellular pathways. In the mitochondrial matrix, Carbamoyl phosphate synthetase I aggressively captures toxic ammonia specifically for safe cellular excretion. Acting primarily in a catabolic role, this enzyme complex aggressively donates the highly reactive carbamoyl group directly to ornithine, thereby driving the crucial urea cycle that safely removes dangerous nitrogenous waste from the entire biological organism.
Conversely, in the cellular cytosol, these activated metabolites fulfill an entirely different, heavily anabolic biological destiny. Here, the distinct enzyme Carbamoyl phosphate synthetase II adeptly uses glutamine as a safe ammonia source to build the fundamental pyrimidine ring structures. This specific cytosolic pathway is absolutely critical for synthesizing essential nucleobases such as uracil, cytosine, and thymine, making this activated carrier a fundamental prerequisite for all biological RNA and DNA synthesis.
Slide 10: Isoprenoids and Lipid-Activated Metabolites
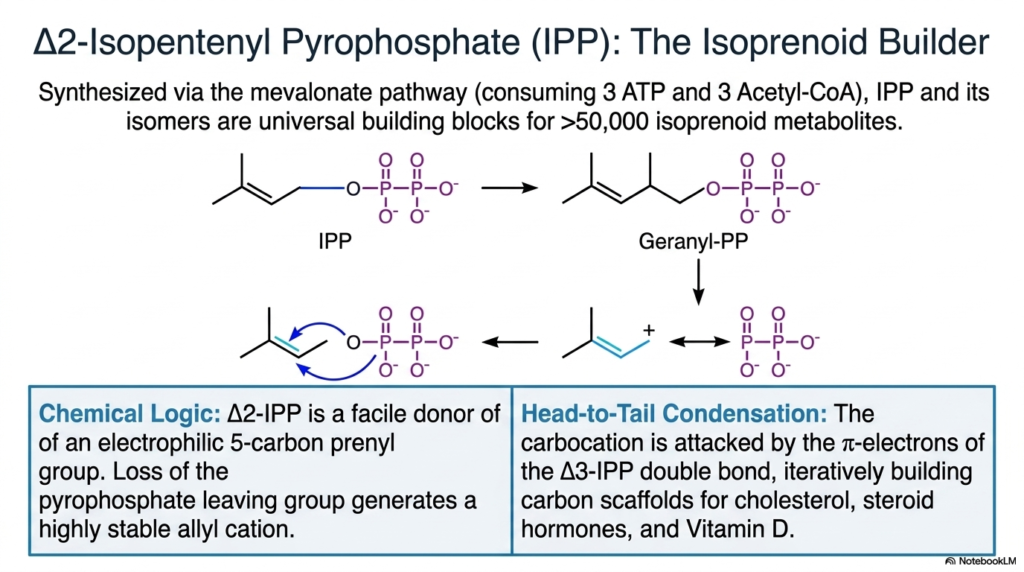
Delta-2-Isopentenyl Pyrophosphate, universally known as IPP, stands out as the ultimate isoprenoid builder among the cellular activated metabolites. Synthesized deep within the complex mevalonate pathway—a process that heavily consumes three molecules of ATP and three molecules of Acetyl-CoA—IPP and its close structural isomers serve as the incredibly versatile, universal molecular building blocks for an astonishing array of more than fifty thousand distinct, biologically essential isoprenoid metabolites.
The underlying chemical logic driving these specific activated metabolites is both elegant and highly aggressive. The Delta-2-IPP molecule acts as a remarkably facile chemical donor of an electrophilic five-carbon prenyl group. When the bulky pyrophosphate leaving group is enzymatically lost, it instantly generates a highly stable, exceptionally reactive allyl cation. This energetic structural transformation perfectly primes the molecule for the rapid, thermodynamically favorable synthesis of complex biological lipids.
The actual construction utilizing these activated metabolites relies on a brilliant mechanism known as head-to-tail condensation. The newly formed, highly reactive carbocation is aggressively attacked by the pi-electrons of a neighboring Delta-3-IPP double bond. This iterative, highly regulated chemical process systematically builds massive, complex carbon scaffolds. These very scaffolds are the absolute structural foundation for incredibly vital biological molecules, including cellular cholesterol, powerful steroid hormones, and essential Vitamin D.
Slide 11: Specialized Activators Among Activated Metabolites
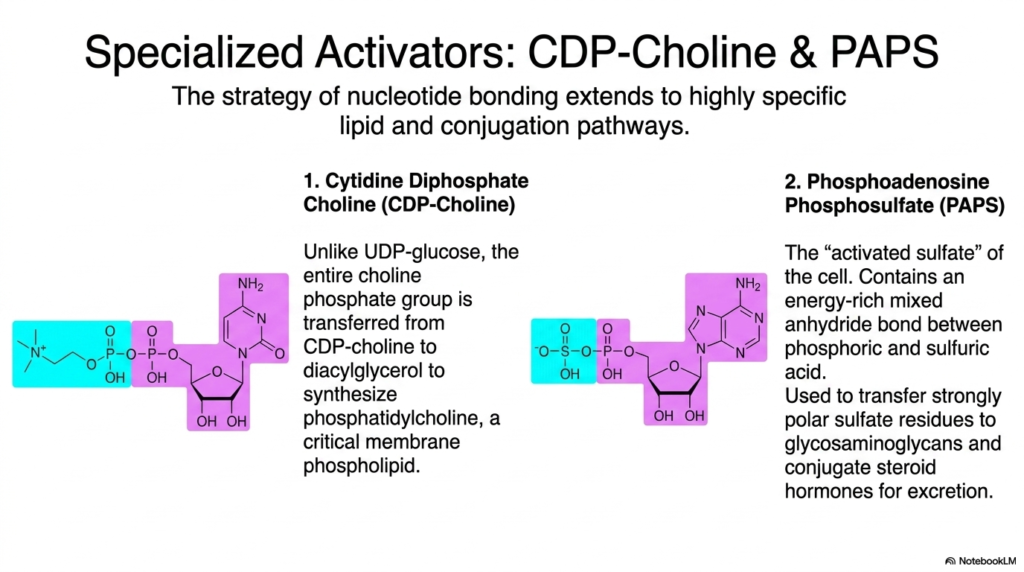
The brilliant biological strategy of nucleotide bonding extends far beyond basic energy transfer to encompass highly specific cellular lipid and conjugation pathways. This slide prominently highlights two highly specialized activated metabolites: CDP-Choline and PAPS. These unique molecules demonstrate how the cell brilliantly repurposes the fundamental chemical logic of massive nucleotide handles to perform highly targeted, incredibly specific biological construction projects that are absolutely vital for maintaining overall cellular homeostasis.
Cytidine Diphosphate Choline, or CDP-Choline, operates uniquely compared to other activated metabolites. Unlike UDP-glucose, which transfers just the sugar, the entire energetic choline phosphate group is aggressively transferred directly from CDP-choline to diacylglycerol. This highly specific enzymatic reaction is absolutely crucial for the rapid biological synthesis of phosphatidylcholine, a fundamental membrane phospholipid that forms the structural boundary of almost every biological cell.
Phosphoadenosine Phosphosulfate, commonly known as PAPS, represents the absolute “activated sulfate” of the biological cell. Containing an incredibly energy-rich mixed anhydride bond between phosphoric and sulfuric acids, it is one of the most highly reactive activated metabolites available. It is widely used to forcibly transfer strongly polar sulfate residues directly onto glycosaminoglycans and to conjugate potent steroid hormones, ensuring their safe, highly efficient urinary excretion.
Slide 12: Molecular Oxygen as the Exceptional Activated Metabolite
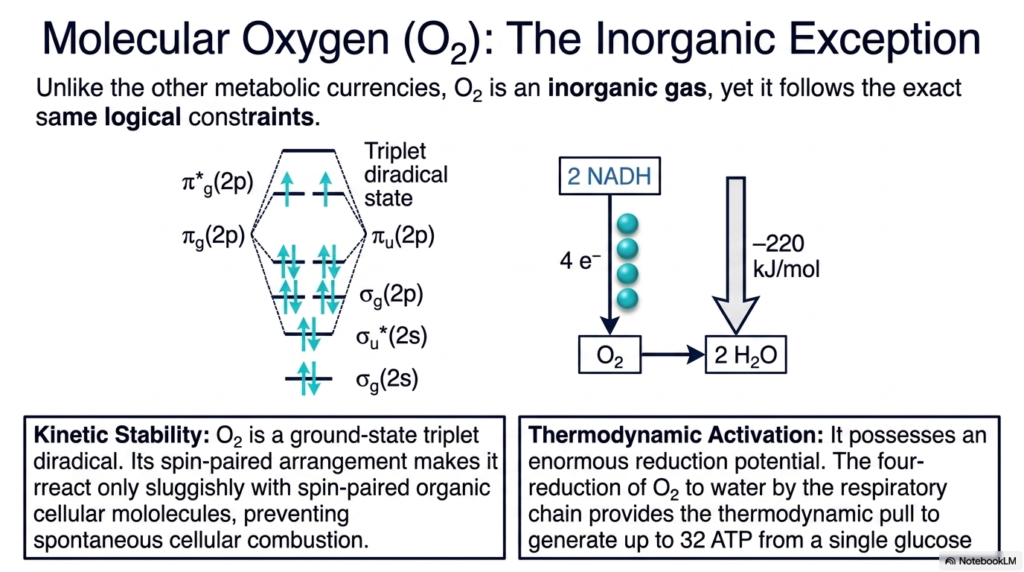
While it may initially seem surprising, molecular oxygen (O2) is a fascinating, notable inorganic exception among the standard activated metabolites. Unlike the massive, complex organic currencies synthesized internally by the cell, oxygen is a simple inorganic gas acquired directly from the surrounding environment. However, when critically analyzed through the strict lens of bioenergetics, it astonishingly follows the exact same logical, thermodynamic constraints as its complex organic counterparts.
The necessary kinetic stability of this unique member of the activated metabolites is firmly rooted in its underlying quantum-chemical principles. Molecular oxygen naturally exists as a ground-state triplet diradical. This highly specific, spin-paired electron arrangement physically forces it to react only with the overwhelmingly spin-paired organic molecules that make up the biological cell, but only incredibly sluggishly. This crucial quantum barrier actively and successfully prevents the horrific, spontaneous chemical combustion of all carbon-based life.
Simultaneously, the extreme thermodynamic activation of these unusual activated metabolites is absolutely staggering. Molecular oxygen possesses an enormous, practically unmatched chemical reduction potential. When specifically handled by the respiratory chain, the complex four-electron reduction of this oxygen to inert water provides the massive, incredibly powerful thermodynamic pull required to successfully generate up to thirty-two fresh molecules of ATP from a single, isolated molecule of oxidized glucose.
Slide 13: The Evolutionary Origins of Activated Metabolites
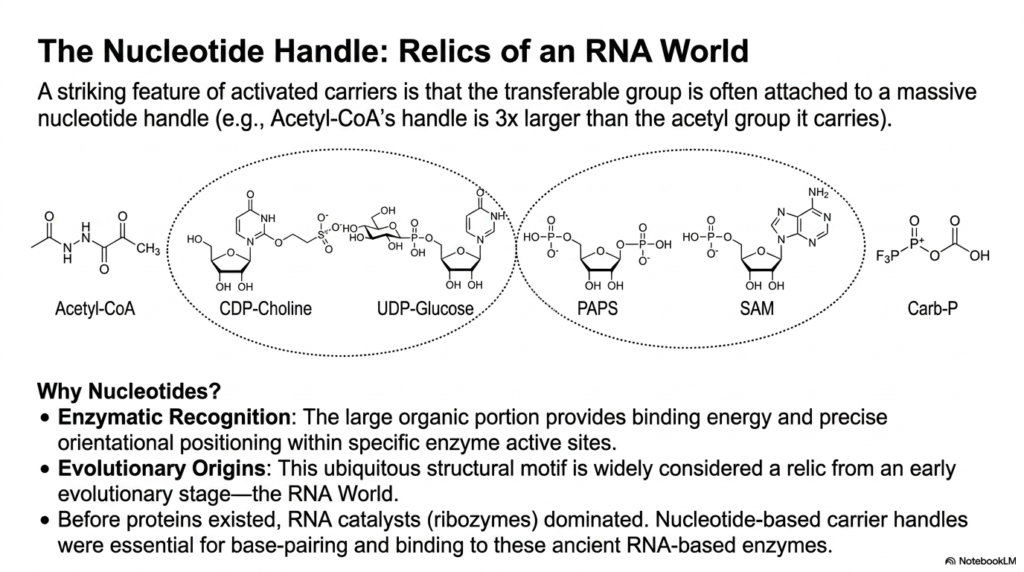
A truly striking, highly consistent structural feature of almost all activated metabolites is that the specific transferable chemical group is very often firmly attached to a surprisingly massive, bulky nucleotide handle. For example, the structurally massive nucleotide handle of Acetyl-CoA is incredibly three times larger than the tiny acetyl group it actually carries. This seemingly inefficient, disproportionate molecular architecture demands a profound, logically sound biochemical and evolutionary explanation.
The immediate biochemical reason for this structural bulk among activated metabolites involves precise enzymatic recognition. The massive, complex organic portion of the nucleotide provides incredibly vital binding energy and absolutely precise orientational positioning deep within highly specific, sophisticated enzyme active sites. Without this large molecular handle, the target enzymes would severely struggle to consistently find, firmly bind, and correctly orient the tiny, highly reactive chemical payloads in the chaotic cellular environment.
However, the profound evolutionary origins of these ubiquitous activated metabolites are even more fascinating. This universal structural motif is widely considered by biochemists to be a direct, ancient relic from an early evolutionary stage known as the RNA World. Long before complex protein enzymes existed, ancient RNA catalysts dominated early life. These large, nucleotide-based carrier handles were absolutely essential for direct base-pairing and physical binding to these ancient, highly primitive RNA-based ribozymes.
Slide 14: System Synthesis and Protein-Modifying Activated Metabolites
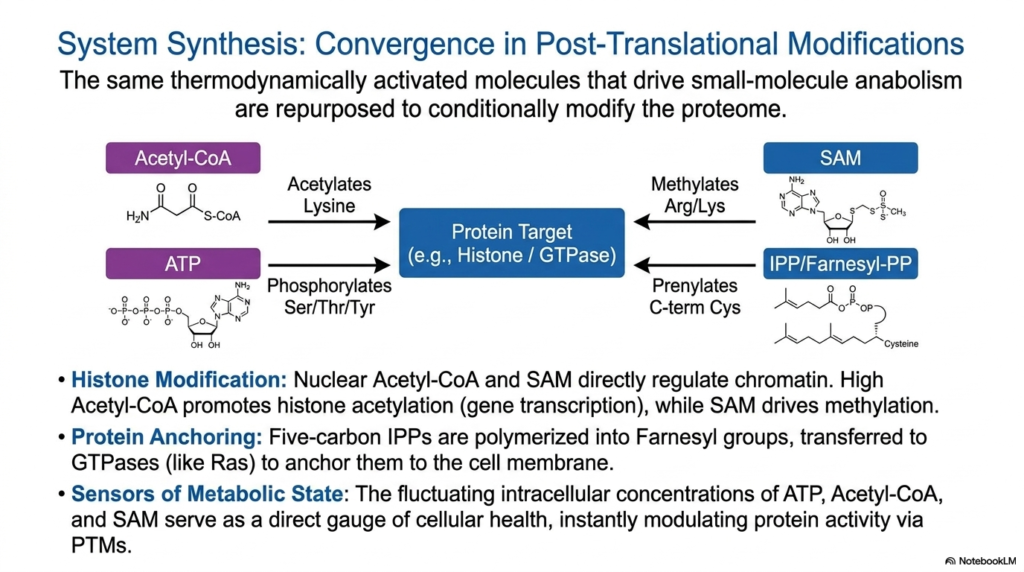
The ultimate chemical brilliance of biological metabolism is profoundly revealed in the way the exact same thermodynamically activated metabolites that normally drive basic small-molecule anabolism are actively repurposed to carefully and conditionally modify the massive cellular proteome. This process relies heavily on post-translational modifications. By carefully recycling these fundamental molecular currencies, the biological cell can instantly and efficiently rewrite the explicit functional instructions of thousands of proteins already manufactured within the cell.
This brilliant system relies heavily on specific activated metabolites, such as nuclear Acetyl-CoA and SAM, to directly and profoundly regulate chromatin structure. High local concentrations of nuclear Acetyl-CoA actively promote crucial histone acetylation, massively opening up DNA for rapid gene transcription. Simultaneously, SAM actively drives the targeted methylation of key arginine and lysine residues. Furthermore, massive five-carbon IPP molecules are actively polymerized to physically anchor essential, highly active GTPases directly to the inner cell membrane.
Crucially, the violently fluctuating intracellular concentrations of these specific activated metabolites serve directly as an incredibly precise, real-time gauge of overall cellular health and immediate energetic status. When cellular resources are incredibly abundant, these energetic currencies physically spill over, instantly and aggressively modulating target protein activity via widespread post-translational modifications. This elegant system ensures that cellular behavior is always perfectly, instantly synchronized with immediate, localized metabolic realities.
Slide 15: The Summary Logic of Activated Metabolites
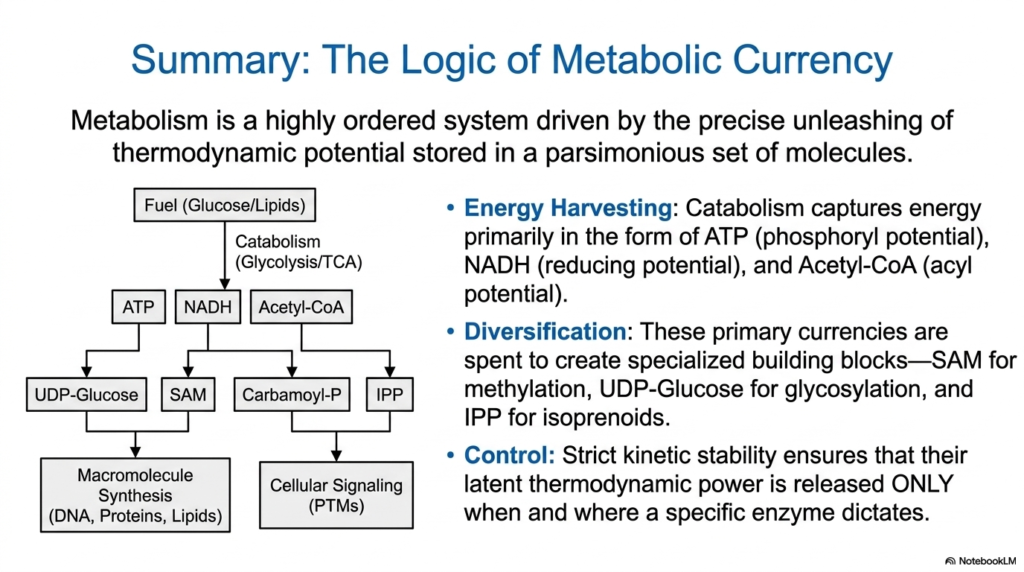
In a final summary, cellular metabolism is not a chaotic mess, but rather a highly ordered, beautifully optimized system driven entirely by the precise, controlled unleashing of massive thermodynamic potential safely stored in a parsimonious set of activated metabolites. This presentation has conclusively demonstrated that the fundamental biological phenomenon of life depends on the strict, highly regulated chemical logic embedded in these incredible, highly specialized energy-carrier molecules.
The fundamental process clearly begins with aggressive energy harvesting. Cellular catabolism efficiently captures large amounts of thermodynamic energy, primarily as the highly useful forms of ATP for phosphoryl potential, NADH for crucial reducing potential, and Acetyl-CoA for essential acyl potential. These three primary activated metabolites form the fundamental energetic foundation of the biological cell, representing the initial, highly critical chemical payoff resulting directly from the complete oxidative breakdown of standard metabolic fuels.
Finally, this energetic system relies on strategic diversification and absolute, uncompromising enzymatic control. These primary activated metabolites are carefully spent to deliberately create highly specialized molecular building blocks, strictly ensuring that their massive latent thermodynamic power is released only exactly when and where a highly specific cellular enzyme rigorously dictates. This incredible kinetic stability ensures that the spectacular power of activated metabolites is never wasted, securing the absolute ongoing survival of the biological organism.
Please read our Content Disclaimer Statement.
Check out our social media channels: