23. Deoxyribonucleic Acid: The Blueprint of Life
The Blueprint of Genetics: How Cells Safely Encode and Store Information
Have you ever wondered how a completely microscopic molecule securely holds the exact instructions to build an entire human being? Every living cell strictly requires a highly stable, meticulously organized chemical database to carefully store its genetic inheritance. This presentation decodes the intricate architecture of these molecular blueprints. By deeply exploring the unique physical properties, strict thermodynamic rules, and incredible structural variations, medical and college students will understand precisely how vital genetic information is safely preserved, rapidly replicated, and flawlessly read.
Slide 1: The Structural Biology of Deoxyribonucleic Acid

To fully understand modern molecular biology and medical genetics, university students must first master the genome’s physical and chemical properties. The structural biology of this complex macromolecule reveals a brilliant, elegant design tailored perfectly for long-term biological data storage. Deoxyribonucleic Acid serves as the foundational master database for all cellular machinery and organismal inheritance. By analyzing Deoxyribonucleic Acid at both the atomic and molecular levels, biochemists can precisely deduce how it interacts with the complex aqueous cellular environment. This foundational knowledge is strictly required to understand cellular replication.
This introductory slide effectively establishes the core themes of the entire presentation: the overarching biochemical structure, the inherent structural polymorphism, and the diverse molecular functions of this essential polymer. Furthermore, Deoxyribonucleic Acid is definitely not a static, rigid entity trapped in a single form. It consistently exhibits remarkable structural polymorphism, meaning the molecule can seamlessly adopt multiple distinct physical conformations depending entirely on its immediate chemical environment and biological state. The upcoming detailed slides will comprehensively explore how fluctuating cellular conditions directly dictate these rapid physical shifts in the double helix.
The dynamic nature of Deoxyribonucleic Acid is absolutely crucial for proper genetic regulation, enabling the massive macromolecule to respond quickly to localized hydration levels and environmental stress. Ultimately, the complex molecular function of this polymer is entirely dictated by its precise three-dimensional physical architecture. From the exact ways biological enzymes selectively access their internal code to the specific methods cells utilize to compact meters of genetic material, every essential biological mechanism is fundamentally driven by structural biochemistry. Mastering this foundational architecture prepares future clinicians for advanced pharmacological analysis.
Recognizing exactly how specialized enzymes safely bind, how dangerous physical damage directly alters the active helix, and how complex epigenetic modifications silently regulate genes allows future medical professionals to understand the true molecular basis of diseases. As students actively progress through this highly detailed biochemical analysis, the profound, inseparable interconnectedness of physical structure and vital biological function will become abundantly clear. This comprehensive examination provides the absolutely essential foundational knowledge rigorously required for advanced studies in both complex human genetics and highly targeted modern molecular therapeutics.
Slide 2: The Monomeric Units Building Deoxyribonucleic Acid
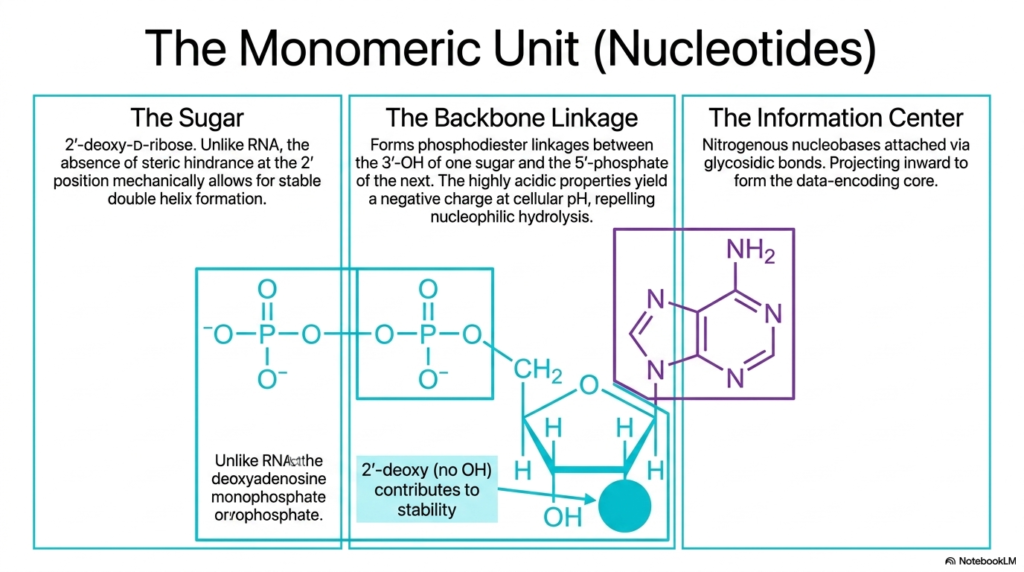
The complex macrostructure of Deoxyribonucleic Acid is fundamentally constructed from much smaller, relatively simple monomeric units called nucleotides. Each individual nucleotide is meticulously composed of three distinct chemical components: a central five-carbon sugar, a highly acidic phosphate backbone linkage, and a nitrogen-rich nucleobase. The central sugar exclusively used in deoxyribonucleic acid is 2′-deoxy-D-ribose. Unlike its close molecular cousin RNA, this highly specific sugar deliberately lacks a reactive hydroxyl group at the 2′ carbon position. This precise evolutionary adaptation mechanically prevents dangerous steric hindrance and vastly increases overall chemical stability.
The structural backbone linkage is robustly formed through highly resilient phosphodiester bonds. These vital chemical bonds specifically connect the 3′-hydroxyl group of one individual sugar molecule directly to the 5′-phosphate group of the immediately adjacent sugar. The highly acidic nature of these phosphate groups reliably yields an overwhelmingly negative electrical charge at standard physiological cellular pH. This pervasive negative charge is biochemically essential because it actively repels nucleophilic attack. Consequently, the protective backbone of Deoxyribonucleic Acid remains exceptionally safeguarded from spontaneous chemical degradation within the water-rich nuclear environment.
The third and final essential component of the nucleotide is the central information center, which explicitly consists of the distinct nitrogenous nucleobases. These distinct molecular rings are covalently attached directly to the central sugar molecules via incredibly strong glycosidic bonds. Structurally, these highly hydrophobic bases project sharply inward from the surrounding sugar-phosphate backbone to forcefully form the shielded data-encoding core of Deoxyribonucleic Acid. By keeping the delicate, vital genetic code completely protected within the helix’s deep interior, the molecule efficiently repels dangerous external chemical threats.
Understanding the precise monomeric structure of these vital nucleotides is absolutely vital for biochemistry and medical students. Every single modern pharmacological agent that specifically targets genetic replication, such as many common systemic chemotherapeutics and targeted antiviral medications, must intimately physically interact with these specific chemical bonds and atomic arrangements. By mastering the precise biochemical properties of the unique central sugar, the acidic protective backbone, and the interior nucleobases, aspiring medical professionals can accurately predict how complex therapeutic molecules will behave at the microscopic level.
Slide 3: Thermodynamics and Base Pairing in Deoxyribonucleic Acid
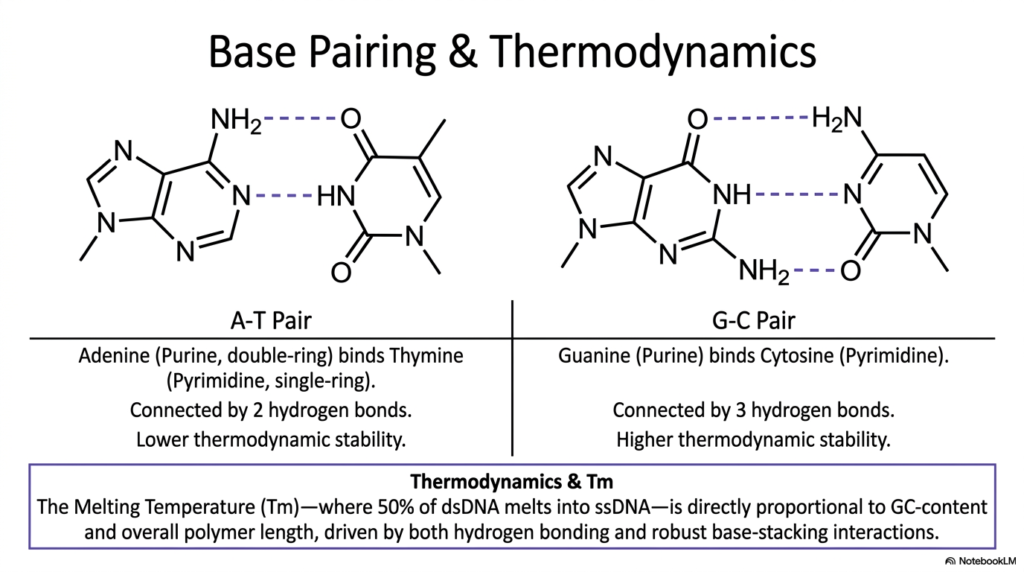
The precise base pairing within Deoxyribonucleic Acid represents the most fundamental biochemical mechanism underlying genetic fidelity and thermodynamic stability. This slide clearly illustrates the highly specific molecular interactions between purines and pyrimidines that hold the double helix together. Adenine, a bulky double-ring purine, exclusively strongly binds to thymine, a smaller single-ring pyrimidine, forming a highly specific A-T pair. This exact interaction is reliably connected by exactly two hydrogen bonds. Because it strictly relies on only two bonds, this pairing possesses inherently lower thermodynamic stability.
Conversely, the vital structural interaction strictly between guanine and cytosine securely forms a much stronger G-C pair. Guanine, another double-ring purine, aligns perfectly with the single-ring pyrimidine cytosine to confidently establish three robust hydrogen bonds. This crucial additional hydrogen bond provides significantly higher thermodynamic stability to the overall architecture of Deoxyribonucleic Acid. Consequently, highly critical regions of the cellular genetic code that strictly require extensive physical durability are typically highly enriched with heavy G-C pairs, making them mechanically much harder to separate during standard biological replication processes.
Understanding these intricate internal chemical bonds introduces the highly critical structural concept of Melting Temperature, commonly denoted as Tm. The Tm is explicitly defined biochemically as the temperature at which exactly 50% of double-stranded Deoxyribonucleic Acid completely melts into exposed, highly vulnerable single-stranded chains. This specific metric is directly proportionally linked to the overall GC-content of the targeted biological sequence, as well as the complete physical length of the polymer. The extra hydrogen bonds found in G-C pairs require significantly more ambient thermal energy to forcefully break.
Furthermore, while highly specific hydrogen bonding is undoubtedly crucial for exact sequence fidelity, the thermodynamic stability of Deoxyribonucleic Acid is equally forcefully driven by robust internal base-stacking interactions. The highly planar, deeply aromatic nature of the central nitrogenous bases allows them to stack tightly on top of one another within the deeply hydrophobic core of the helix. These inherent, powerful van der Waals forces and hydrophobic effects contribute massively to overall structural integrity, actively preventing spontaneous unwinding of the delicate genetic material under standard physiological conditions.
Slide 4: The B-Form Architecture of Deoxyribonucleic Acid
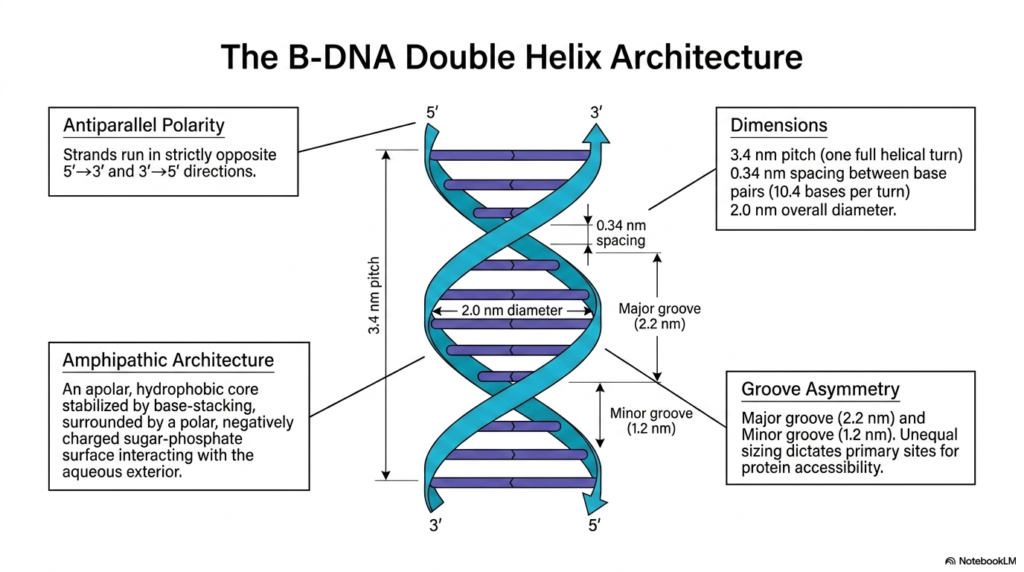
The canonical B-form physical architecture of Deoxyribonucleic Acid is undoubtedly the most incredibly prevalent and biologically significant structural conformation natively found within living eukaryotic cells. A primary defining feature of this magnificent architecture is its strict antiparallel polarity. The two distinct individual strands of the complex double helix run in strictly opposite directions. One continuous strand is oriented strictly in the 5′ to 3′ direction, while the strictly complementary strand is oriented precisely 3′ to 5′. This antiparallel arrangement mathematically aligns internal bases for optimal hydrogen bonding.
The fundamental physical design of Deoxyribonucleic Acid is inherently beautifully amphipathic, meaning it actively possesses both potent water-loving and water-repelling properties. The structural anatomy features a highly apolar, deeply hydrophobic inner core where the flat nitrogenous bases stack securely far away from reactive cellular water. Surrounding this incredibly delicate internal data core is a highly polar, strongly negatively charged sugar-phosphate surface that readily interacts with the bulk aqueous exterior. This brilliant biochemical arrangement successfully intimately shields sensitive genetic data while keeping the massive polymer fully soluble.
Precise mathematical dimensions rigidly govern the classic B-form of Deoxyribonucleic Acid. The biological structure maintains a highly consistent overall physical diameter of exactly 2.0 nanometers. Furthermore, the spiraling helix formally completes one full structural turn, known universally as its pitch, exactly every 3.4 nanometers. Within this exact physical pitch, there is a strict, rigid 0.34-nanometer vertical spacing intimately between each successive flat base pair, perfectly successfully accommodating exactly 10.4 individual bases per complete helical turn. These specific structural metrics elegantly provide the predictable physical geometry absolutely necessary for enzymatic recognition.
Finally, the asymmetric twisting of the rigid structural backbone dynamically generates distinct external spatial features, universally known as the major and minor grooves. The significantly wider major groove measures exactly 2.2 nanometers, while the distinctly minor groove is significantly tighter at exactly 1.2 nanometers. This essential, biologically critical groove asymmetry in Deoxyribonucleic Acid precisely dictates the primary cellular sites for direct sequence-specific protein accessibility. The wider major groove clearly exposes the unique chemical edges of the internal bases to regulatory factors.
Slide 5: The Coding Mechanisms of Deoxyribonucleic Acid
The primary biological utility of Deoxyribonucleic Acid rests entirely in its absolutely unique structural ability to securely encode incredibly complex genetic information. This slide methodically breaks down the precise biochemical mechanisms by which this stored molecular data is physically read and actively interpreted by complex cellular machinery. During the intricate biological process of transcription, the protective double helix is carefully and temporarily unzipped, explicitly revealing two distinct strands with entirely different functional roles. Understanding this division is absolutely crucial for students to analyze modern gene expression comprehensively.
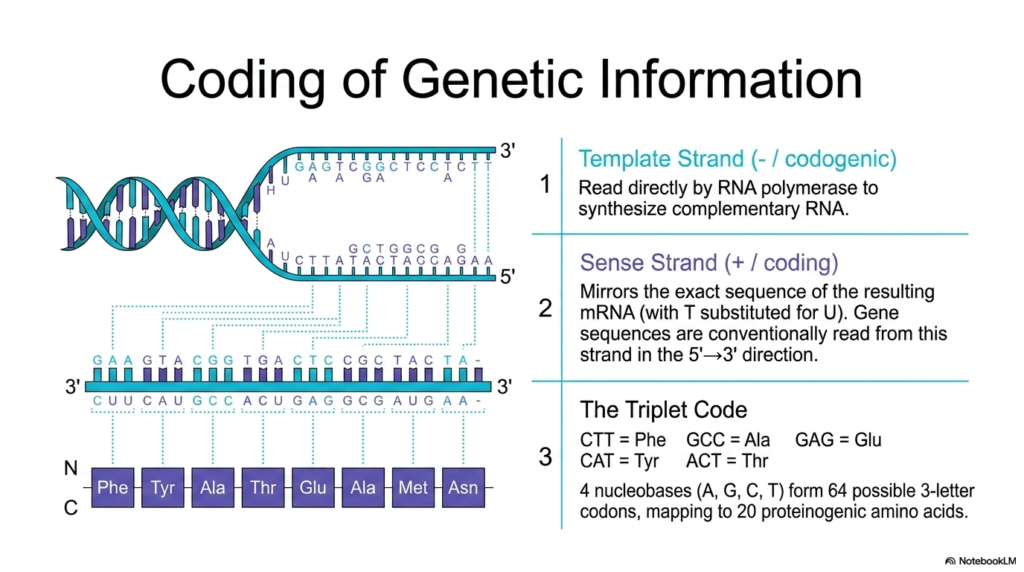
The first vital functional strand is the Template Strand, commonly accurately referred to by biochemists as the negative or codogenic strand. This specific unwound section of Deoxyribonucleic Acid is actively read directly by the massive multi-subunit enzyme RNA polymerase. As the powerful enzyme aggressively traverses the exposed genetic template, it systematically synthesizes a brand-new, exact complementary RNA molecule. The physical sequence of this specific template strand successfully serves as the exact biological mold strictly required to reliably produce highly functional, deeply essential cellular messenger RNA.
Directly opposite the template is the heavily utilized Sense Strand, universally correctly known as the positive or biological coding strand. This structural strand beautifully mirrors the exact sequence of the newly generated mRNA transcript, with the single biological exception that Deoxyribonucleic Acid naturally uses thymine, whereas RNA chemically substitutes uracil for thymine. Conventionally, vitally important gene sequences are consistently always read and accurately reported by molecular biologists directly from this specific sense strand flowing in the strict 5′ to 3′ direction, providing a highly direct, readable biological copy.
The ultimate biological translation of this securely stored genetic information strongly relies on the mathematical Triplet Code. The four basic structural nucleobases are neatly mathematically organized strictly into three-letter informational combinations called discrete codons. This brilliant mathematical arrangement securely and perfectly allows for exactly 64 possible codon combinations, which map to the 20 standard proteinogenic amino acids used by life. For instance, the exact sequence CTT explicitly and reliably codes for phenylalanine. This beautifully universal coding system heavily highlights the elegant efficiency of Deoxyribonucleic Acid in driving massive protein synthesis.
Slide 6: Structural Polymorphism Within Deoxyribonucleic Acid
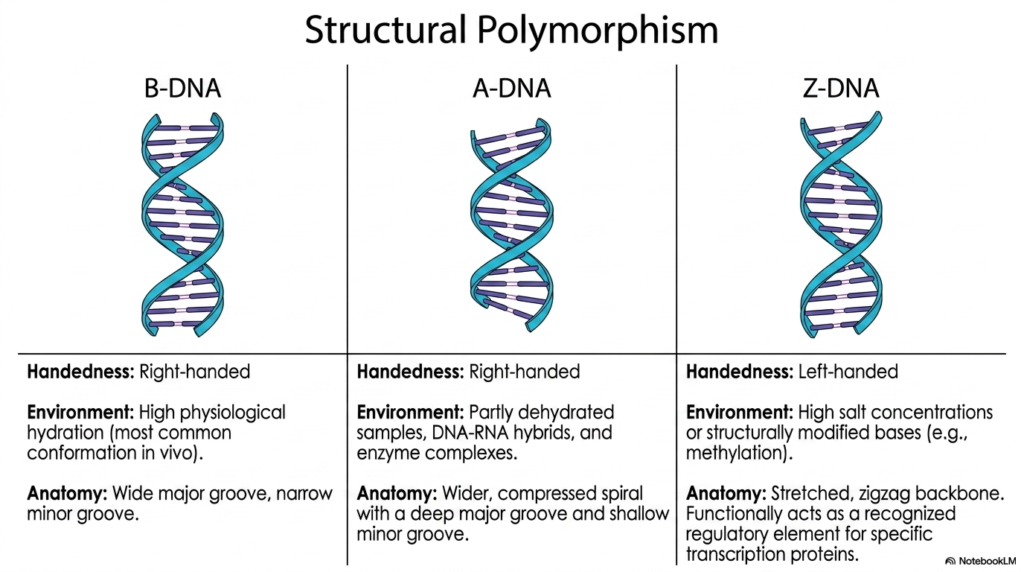
While the classic smooth double helix is widely recognized globally, Deoxyribonucleic Acid actually exists in multiple distinctly different structural conformations. This vital dynamic structural polymorphism wonderfully enables the genetic material to respond successfully to rapidly varying physiological conditions and intense environmental cellular stresses. The most abundant structural conformation natively in vivo is standard B-DNA, biochemically characterized as a beautifully right-handed helix. B-DNA thrives in active cellular environments characterized by extremely high physiological hydration and a recognizable, balanced structural anatomy.
Under highly varying, stressful environmental conditions, Deoxyribonucleic Acid can immediately and dramatically shift directly into the alternative dense A-DNA conformation. Similar to B-DNA, A-DNA is technically a right-handed helix, but it typically forms rapidly in partly physically dehydrated biological samples. It is also the strongly dominant conformation quickly and happily adopted by temporary DNA-RNA hybrid complexes precisely during active genetic transcription. Anatomically, A-DNA presents a much wider, more severely compressed spiral, featuring a notably deep major groove and a remarkably shallow minor groove.
The most dramatic structural variation consistently observed by biochemists is Z-DNA, which successfully adopts a highly exceptional, totally left-handed helical rotation. Z-DNA formation is actively and rapidly triggered by highly specific, extreme cellular environmental cues, such as dangerously high salt concentrations or the physical presence of heavily modified bases. Instead of a beautiful, smooth, continuous spiral, the physical anatomy of this bizarre Deoxyribonucleic Acid conformation explicitly heavily presents a stretched, distinctively jagged zigzag backbone, making it a highly specific biological structural target.
Functionally, this highly bizarre Z-DNA effectively acts as a highly specific, recognized regulatory element reserved strictly for highly specific cellular transcription proteins. By actively, dramatically, and physically shifting directly from a standard right-handed to a left-handed physical conformation, the vital genetic material rapidly and completely alters localized physical tension. Recognizing these exact profound polymorphic states is deeply essential for exceptional medical students, as these dramatic structural transitions actively influence localized complex gene expression, active destructive viral integration, and the overall physical dynamics of Deoxyribonucleic Acid.
Slide 7: Telomeres and G-Quadruplexes in Deoxyribonucleic Acid
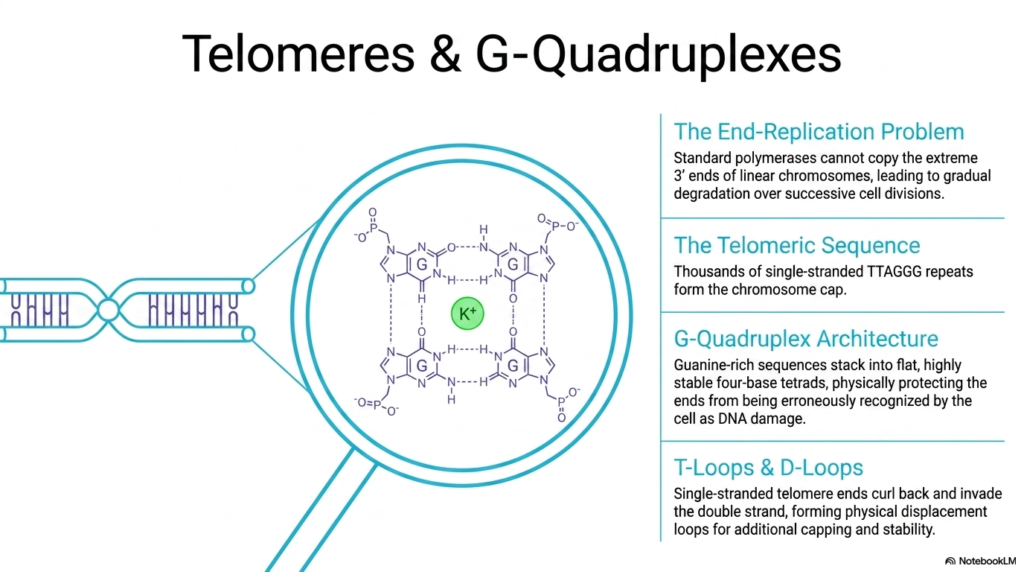
The massive linear biological chromosomes found securely deep within eukaryotic cells constantly face a severe, potentially lethal mechanical challenge, clinically known as the end-replication problem. Because highly standard cellular polymerases absolutely strictly require a pre-existing biological primer to function, they physically utterly cannot reliably copy the extremely vulnerable 3′ ends of linear Deoxyribonucleic Acid strictly during biological replication. Without a highly protective structural defense mechanism, this fundamental chemical limitation would inevitably lead to the gradual, complete, catastrophic degradation of vital, life-sustaining genetic material.
To forcefully and successfully combat this immense cellular vulnerability, eukaryotic organisms consistently and heavily utilize highly specialized, highly robust structural caps uniquely called telomeres. The complex, massive telomeric sequence explicitly and reliably consists of many thousands of highly repetitive, completely single-stranded TTAGGG sequences that actively form a massive physical biological buffer exactly at the chromosome ends. This totally disposable protective stretch of Deoxyribonucleic Acid safely, effectively, and happily absorbs the inevitable, dangerous genetic sequence loss that naturally and painfully occurs during each and every single biological cellular replication cycle, fiercely preventing premature cellular aging.
The uniquely profound, guanine-rich chemical composition of these massive, complex telomeres amazingly and explicitly allows them to rapidly form an incredibly dense, deeply protective alternative structural architecture, universally known as a massive G-Quadruplex. In this exceptionally rare structural conformation, the flat guanine nucleobases stack smoothly into highly stable four-base tetrads, explicitly stabilized by central coordinating potassium ions. This highly complex structural architecture safely and deeply protects the highly exposed, vulnerable ends of the massive, active linear Deoxyribonucleic Acid.
Furthermore, the completely unprotected, single-stranded, vulnerable telomere ends actively and wonderfully curl back directly and intimately upon themselves and physically and aggressively invade the nearby upstream stable double strand. This extraordinarily complex physical and biological maneuver rapidly and beautifully forms intricate topological displacement loops, commonly, and more technically, referred to as protective T-Loops and robust D-Loops. These highly complex, intricate topological knots physically entirely hide the loose, vulnerable genetic ends, explicitly reliably providing immense additional deep biological capping and extreme physical structural stability completely against accidental, dangerous recognition as severely broken cellular Deoxyribonucleic Acid.
Slide 8: Topology and Supercoiling of Deoxyribonucleic Acid
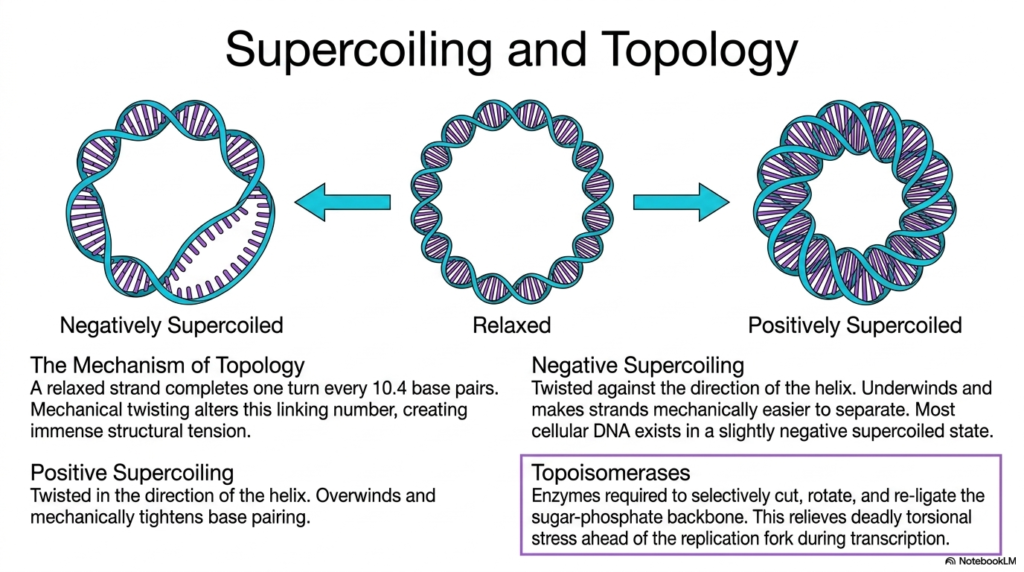
Due to the truly massive, completely immense physical length of highly complex vital cellular genomes, the tiny microscopic physical space Deoxyribonucleic Acid happily occupies must be tightly regulated by a remarkable, amazing property known universally in biochemistry as topology. When a long continuous DNA strand is structurally completely physically relaxed, it beautifully, naturally, perfectly completes exactly one full helical turn precisely every 10.4 base pairs. However, totally natural essential biological processes inevitably introduce intense, violent mechanical twisting that immediately and powerfully alters this highly precise optimal linking number.
When Deoxyribonucleic Acid is forcefully twisted entirely against the natural preferred direction of its biological resting helix, it rapidly enters a highly essential state of strong negative supercoiling. This truly vital, incredibly important biological state fundamentally and actively underwinds the targeted genetic material, successfully and miraculously making the two individual strands mechanically far easier to physically and safely separate. Because active vital biological processes like widespread genetic transcription and entire cellular reproduction strictly require massive, continuous strand separation, most functional native cellular DNA typically exists in a highly underwound state.
Conversely, massive positive supercoiling powerfully forcefully occurs directly entirely when the biological molecule is violently forcefully severely twisted completely in the exact parallel direction of the natural biological resting helix. This intense, violent, massive physical action severely, aggressively, and intensely overwinds the genetic strand, which mechanically, terribly, tightly, and powerfully tightens the internal physical base pairing and strongly, intensely, and dangerously prevents the successful, essential separation of the delicate double helix. Excessive, dangerous, positive supercoiling can forcefully halt cellular processes by actively and deeply rendering the required Deoxyribonucleic Acid practically inaccessible.
To safely, beautifully, and powerfully manage these truly massive, highly destructive torsional forces, all eukaryotic biological systems rely heavily on highly specialized, immensely powerful enzymes known chemically and biologically as complex Topoisomerases. These highly sophisticated, wonderful, complex biological proteins are strictly and continuously required to selectively, precisely, cleanly cut, deliberately, gracefully, smoothly rotate, and precisely, gracefully, perfectly re-ligate the vital, highly stressed sugar-phosphate backbone. By faithfully, beautifully, and safely performing this incredibly complex, intricate biochemical molecular surgery, these crucial topoisomerases effectively, smoothly, and thoroughly relieve the deadly, massive torsional stress, thereby preventing Deoxyribonucleic Acid destruction.
Slide 9: Higher-Order Chromatin Packaging of Deoxyribonucleic Acid
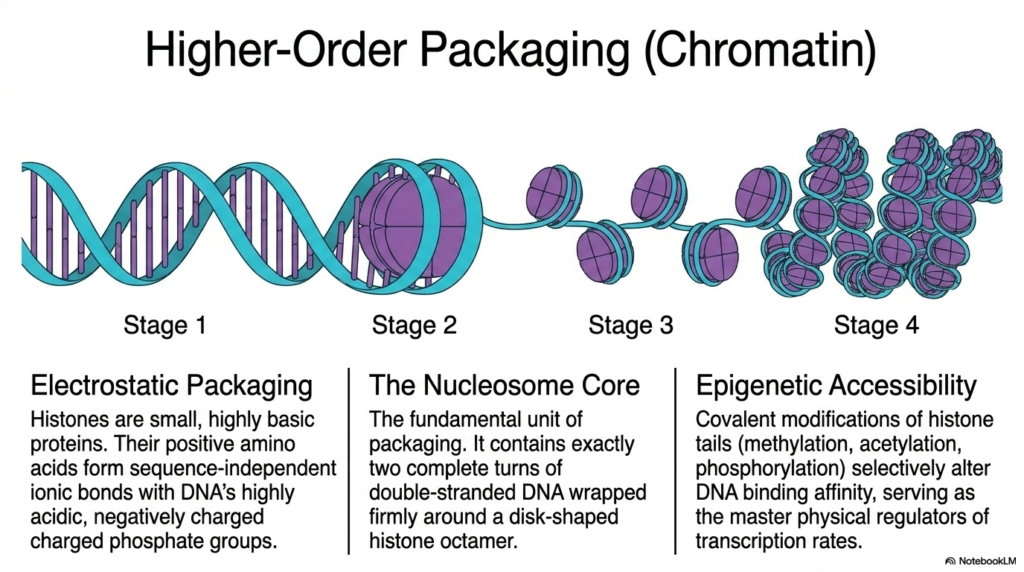
To successfully fit nearly two full meters of vital Deoxyribonucleic Acid securely into a tiny, microscopic cellular nucleus, eukaryotic organisms heavily utilize highly sophisticated, higher-order physical packaging mechanisms. This massive spatial biological challenge is elegantly solved strictly through a highly specific process called dense electrostatic packaging. The biological system fundamentally relies on small, highly basic cellular proteins known as histones. These crucial structural proteins provide the necessary foundational framework to dramatically compress the massive genome while maintaining its overall structural integrity.
Because these vital histones are incredibly rich in positive basic amino acids, they seamlessly form intense, tight ionic bonds with the highly acidic, negatively charged phosphate groups found throughout Deoxyribonucleic Acid. The fundamental primary structural building block of this intense packaging system is universally known as the nucleosome core. This microscopic, highly dense structural unit features exactly two complete geometric turns of double-stranded genetic material wrapped incredibly tightly around a rigid, disk-shaped central histone octamer, thereby condensing vast lengths of genetic code.
This precise structural arrangement creates a highly compact unit that resembles microscopic beads densely strung along a continuous molecular string. This intricate cellular packaging is organized into multiple distinct physical stages, ultimately forming the massively complex macromolecular structure biologically recognized as chromatin. While this dense biological compaction completely solves the initial spatial challenge, it inherently restricts essential molecular access to the underlying Deoxyribonucleic Acid. The genome is effectively physically protected, but the required biological code becomes heavily physically buried and temporarily inaccessible to crucial reading enzymes.
To effectively overcome this immense physical barrier, living cells actively utilize precise epigenetic accessibility mechanisms. Through targeted covalent chemical modifications of completely exposed histone tails, such as direct methylation or rapid phosphorylation, the cell can selectively alter physical binding affinity. These highly regulated chemical changes accurately serve as the master physical regulators of cellular transcription rates, actively loosening or tightly tightening the dense Deoxyribonucleic Acid architecture to dynamically control targeted genetic networks. This elegant regulation is absolutely crucial for proper cellular differentiation and overall biological development.
Slide 10: Chemical Modifications and Epigenetics of Deoxyribonucleic Acid
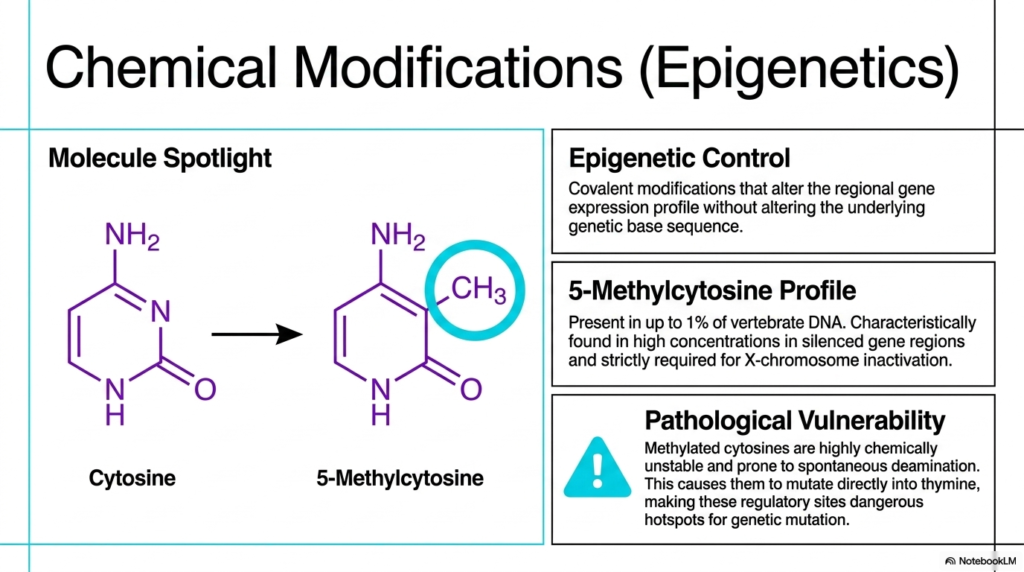
Beyond massive structural physical packaging, the incredibly complex biological function of the cellular genome is strictly, heavily regulated by direct, highly targeted chemical modifications to the Deoxyribonucleic Acid itself. The most incredibly prominent biological example of this intense epigenetic control directly involves the targeted covalent chemical modification of specific internal nitrogenous bases. These delicate, highly precise chemical additions completely alter the regional gene expression profile without fundamentally altering the underlying biological base sequence, producing distinct downstream phenotypic outcomes across entirely different tissue types.
A truly prime biological example of this essential epigenetic regulation is the precise cellular formation of 5-Methylcytosine. Highly specialized cellular biological enzymes strategically and securely attach a very small methyl group directly to standard cellular cytosine molecules heavily embedded within the sequence. This highly specific biological modification is incredibly functionally prevalent, physically present in up to 1% of all vertebrate Deoxyribonucleic Acid. High localized concentrations of these heavily methylated cytosines are consistently found deep within highly silenced regulatory functional gene regions.
The widespread physical presence of 5-Methylcytosine is definitively not merely a general biological regulatory mechanism; it is strictly and functionally required for vital biological processes such as targeted X-chromosome inactivation in human females. By densely methylating an entire targeted chromosome, the cell permanently shuts down its active expression to maintain proper functional genetic dosage. Thus, direct chemical epigenetic modification of Deoxyribonucleic Acid acts precisely as a highly powerful, extremely long-term biological switch that fundamentally, forcefully governs complex cellular lineage and safe, healthy biological differentiation.
However, this powerful biological regulatory mechanism unfortunately introduces a highly dangerous pathological vulnerability directly into the delicate genome. Methylated cytosines embedded within Deoxyribonucleic Acid are notoriously structurally chemically unstable and highly prone to a dangerous biological process chemically called spontaneous deamination. When this dangerous reaction occurs rapidly, 5-methylcytosine immediately chemically mutates into standard thymine. Because the cell’s repair machinery heavily struggles to properly recognize this exact specific mismatch, these exact regulatory sites, unfortunately, reliably become exceedingly dangerous active hotspots for permanent genetic mutation.
Slide 11: Biological Mutagens and Damage to Deoxyribonucleic Acid
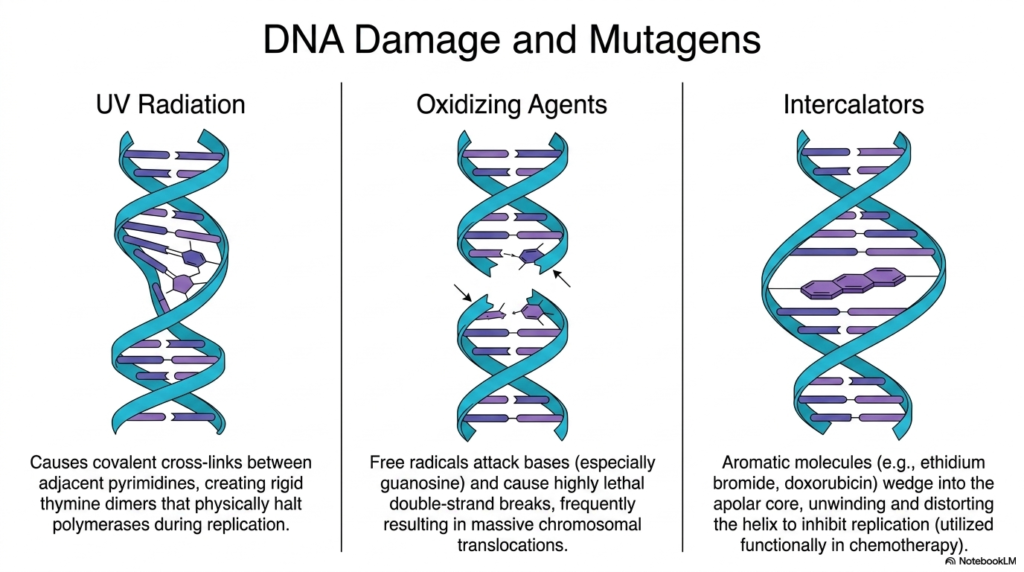
The highly delicate physical structure of the complex cellular genome is constantly under brutal attack from a vast array of highly dangerous external environmental and highly reactive internal chemical mutagens. One of the most prevalent physical threats to Deoxyribonucleic Acid is standard ultraviolet radiation directly originating from the sun. When cellular genetic material rapidly absorbs highly aggressive incoming UV photons, it actively causes highly dangerous covalent cross-links to form directly between adjacent internal pyrimidine bases, instantly creating highly rigid, highly disruptive thymine dimers.
These massive, heavily bulky thymine dimers severely distort the normally perfectly smooth physical architecture of the genome. When highly active cellular biological polymerases suddenly encounter these massive structural roadblocks during standard active replication, the required enzymes physically halt the vital biological cellular reproduction process. If the living cell tragically fails to rapidly and actively excise and fully repair this extremely localized biological damage, the massively stalled active replication forks can directly, easily, and instantly lead to massive, catastrophic cellular death.
Another highly severe, aggressive systemic threat to Deoxyribonucleic Acid rapidly routinely comes from highly aggressive cellular internal oxidizing agents. Normal essential cellular metabolism constantly produces highly dangerous reactive free radicals that aggressively attack the delicate internal genetic nucleobases, particularly targeting the highly vulnerable guanosine molecules. This extremely rapid, brutal oxidative metabolic stress frequently heavily tragically causes completely highly lethal severe double-strand biological breaks precisely in the sensitive structural backbone, severely violently completely physically separating the massive biological chromosome entirely into multiple dangerous unattached fragments.
Finally, complex, highly artificial chemical and biological intercalators constitute a truly major, severe external source of induced, massive genetic damage. Extremely large, highly aromatic, bulky chemical molecules physically rapidly violently wedge themselves entirely directly strongly into the protected, deep, apolar interior core of the double biological helix. By forcefully, violently, physically slipping, strictly, tightly between the beautifully stacked nitrogenous bases, these complex, bulky intercalators aggressively, severely, deeply unwind and massively, structurally, violently distort the overall, completely vital Deoxyribonucleic Acid architecture, a mechanism frequently therapeutically, successfully utilized in targeted oncology.
Slide 12: Protein Interactions and Deoxyribonucleic Acid Regulation
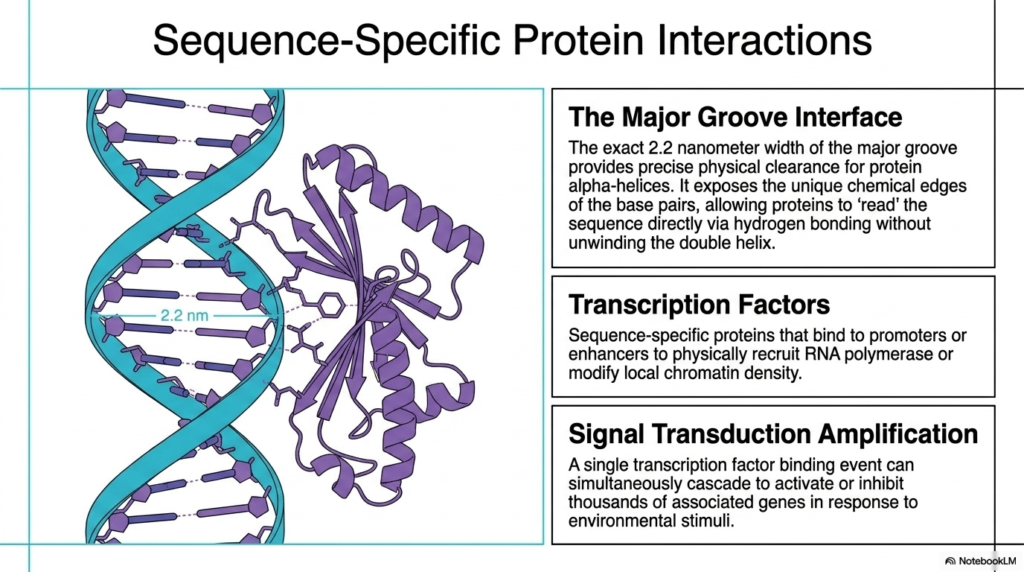
The entirely static, beautifully stored, immense genetic information located deep within the cellular genome is utterly and entirely functionally useless without highly specialized, active cellular proteins capable of reliably and reliably reading the structural code. To successfully, biologically, and correctly access this completely vital data, the highly precise physical architecture of Deoxyribonucleic Acid explicitly and cleverly provides a truly massive, open, structural regulatory gateway, chemically universally known as the major groove interface. The exact 2.2-nanometer external width of this massive major groove securely provides perfect protein access clearance.
Crucially, this exceptionally wide external major groove fully and actively exposes the unique complex chemical edges of the internal functional base pairs to the surrounding aqueous cellular physiological environment. This entirely brilliant structural-physical orientation strictly and perfectly allows specialized biological regulatory sensory proteins to actively and clearly decode the underlying vital sequence directly via complex, intricate, highly specific hydrogen-bonding biological networks. Remarkably, these specialized functional regulatory proteins can accurately and completely decode the underlying Deoxyribonucleic Acid without ever dangerously unwinding the helix.
The massive, complex, specialized proteins that are fully responsible for actively navigating this vital biological interface are widely known as sequence-specific transcription factors. These highly specialized biological molecules directly bind to highly targeted upstream promoters or large, distant regulatory enhancer regions. Once completely securely bound directly to the specific sequence, these vital transcription factors actively act to physically explicitly recruit the required functional RNA polymerase machinery. Ultimately, these incredibly precise, sequence-specific transcription factors serve as the powerful master gatekeepers for all crucial downstream cellular genetic expression.
A single, highly targeted transcription factor binding event, safely anchored to Deoxyribonucleic Acid, can fundamentally and quickly trigger truly massive active signal transduction functional amplification across the interconnected biological cellular network. By successfully quickly dynamically recognizing a highly specific localized structural motif precisely successfully embedded entirely within the genome, one fully activated essential regulatory protein can simultaneously quickly powerfully cascade to completely actively activate or strictly forcefully entirely inhibit thousands of associated complex cellular downstream biological genes in a completely rapid direct biological response to external environmental stimuli.
Slide 13: The Enzymatic Modifiers of Deoxyribonucleic Acid
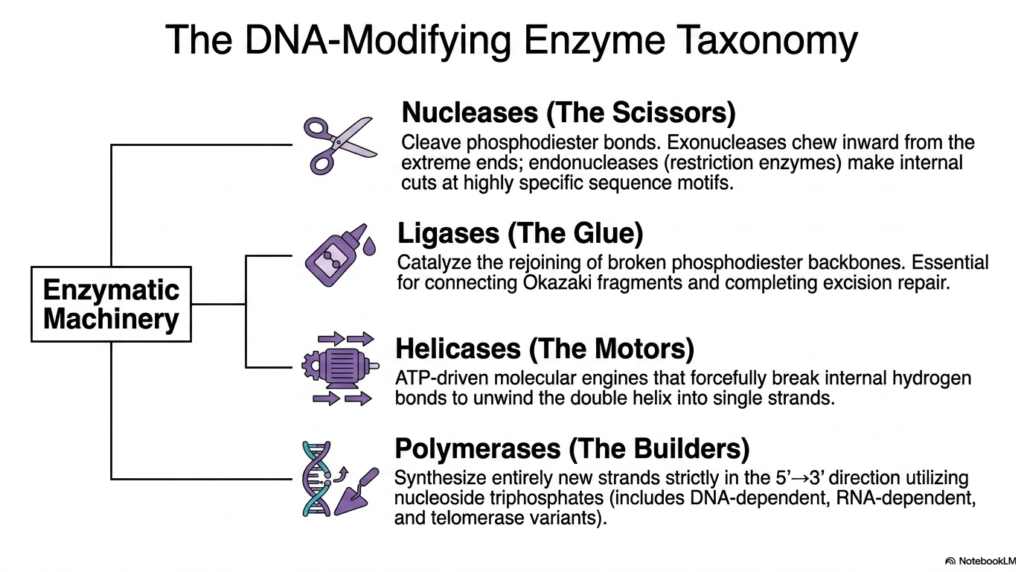
To successfully replicate, structurally repair, and correctly express the complex cellular genome, eukaryotic organisms require a functionally diverse taxonomy of highly specialized deoxyribonucleic acid-modifying enzymes. These mechanically efficient, high-performance molecular machines actively perform the precise physical work required to manipulate the massive double helix. This essential biological toolkit provides the living cell with exactly what it needs to manage the massive, heavy polymer completely reliably. The absolutely first major category of this enzymatic machinery is colloquially known as the cellular scissors, biochemically classified as the vital Nucleases.
Within this vital nuclease family, specialized exonucleases actively and methodically chew inward directly from the exposed, open terminal ends of deoxyribonucleic acid strands. Meanwhile, incredibly powerful functional endonucleases, often called restriction enzymes, make precisely targeted, highly specific internal cuts in DNA at specific sequence motifs. Once these vital enzymatic cuts have successfully served their direct biological purpose, the structural backbone must be carefully and actively repaired. To fully resolve these internal breaks and carefully, beautifully restore genomic stability, the living cell relies entirely on the powerful action of biological Ligases.
Acting strictly as a highly targeted biological glue, vital cellular Ligases catalyze the rapid rejoining of violently broken biological phosphodiester backbones. This critical restorative process safely restores complete structural continuity within Deoxyribonucleic Acid, which is biologically essential for successfully connecting Okazaki fragments during replication and completing vital, precise excision repair. Additionally, Helicases function powerfully as robust, ATP-driven molecular motors. These intense biological engines forcefully, successfully, and gracefully break the internal hydrogen bonds exactly between the complementary nitrogenous bases, actively unwinding the stable double helix entirely into highly exposed single strands.
Finally, the highly complex biological Polymerases serve as the foundational molecular-cellular builders. These massive multi-subunit enzymes reliably and continuously synthesize entirely new biological complementary genetic strands. They operate strictly in the universally required 5′ to 3′ biological direction, diligently ensuring perfect sequence fidelity. By systematically and efficiently matching exact specific complementary nucleoside triphosphates directly to the exposed Deoxyribonucleic Acid cellular template strand, they guarantee accurate genetic survival. This specific enzyme taxonomy—comprising exactly the essential cutters, robust glues, powerful motors, and diligent builders—provides the complete necessary biochemical foundation for all life.
Slide 14: Genetic Recombination of Deoxyribonucleic Acid
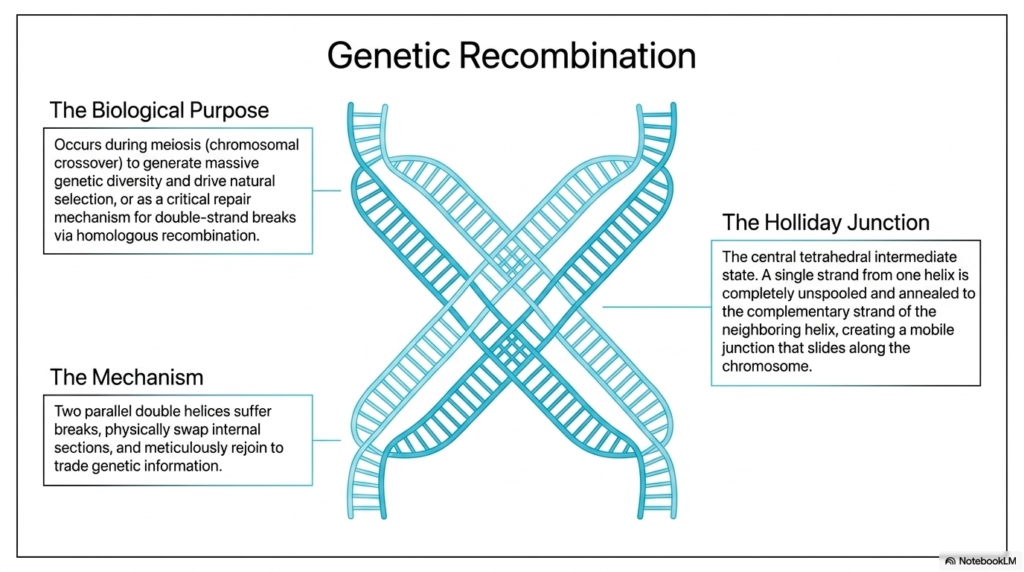
Genetic recombination is an intricate biological mechanism designed to ensure biological diversity and meticulously manage extensive structural genomic repair. The primary biological purpose of this vital function is to occur deeply during cellular meiosis, specifically through highly targeted chromosomal crossover. By actively shuffling the internal genetic deck, living organisms successfully generate the massive vital genetic diversity strictly required to continually drive natural selection. This brilliant dynamic shuffling ensures that the inherited Deoxyribonucleic Acid is consistently and fundamentally functionally distinct from that of the original parental cells.
Beyond merely driving vital reproductive diversity, complex homologous recombination serves as a critical biological repair mechanism specifically targeted to highly lethal double-strand breaks. The underlying physical mechanism strictly and heavily involves two closely and perfectly parallel double helices simultaneously undergoing severe structural breaks. These identical sequences then actively, physically, and seamlessly swap massive internal sections of their genetic code. The broken ends seamlessly rejoin to trade highly specific vital genetic information beautifully, perfectly, faithfully, and carefully, without ever losing any vital sequence data from the overall Deoxyribonucleic Acid structure.
The Holliday Junction is a central, critical intermediate state necessary for biological recombination. During this important biochemical stage, a completely damaged single strand from a broken DNA helix is unspooled. This strand is then carefully and precisely annealed to the complementary strand of a neighboring intact helix. This forms a highly mobile tetrahedral junction that can smoothly slide along the chromosome.
Understanding this specific physical tetrahedral junction is essential for modern genetics students. When the complex cellular enzymes resolve this central mobile junction effectively and efficiently, they cleanly and accurately separate the massive chromosomes into entirely new, distinct biological arrangements.
Slide 15: Synthesis and Determinism of Deoxyribonucleic Acid
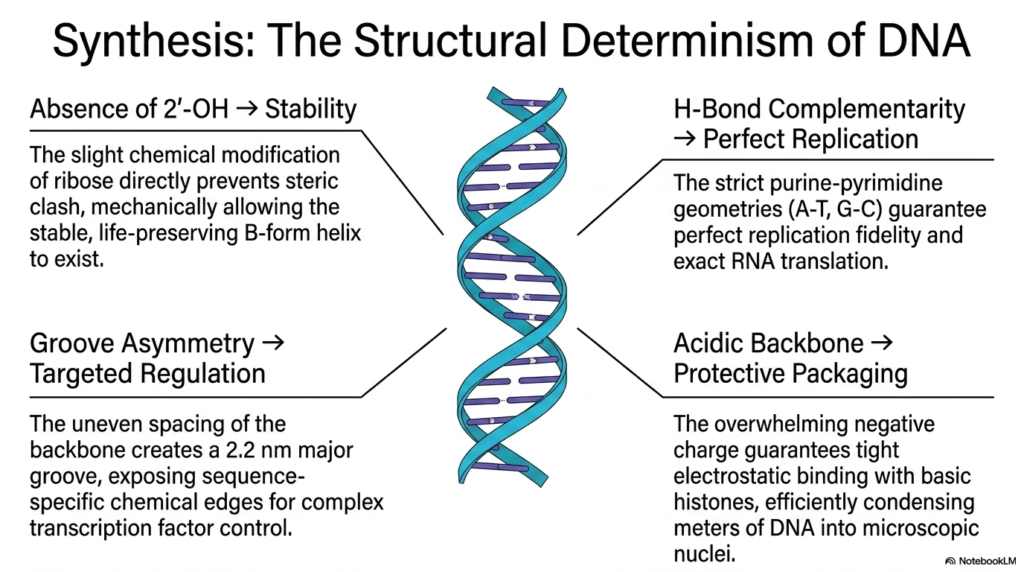
This final comprehensive slide successfully synthesizes the absolutely critical foundational structural relationship, explicitly and beautifully defining the entire presentation: beautiful, complete, structural, physical determinism. Every single unique physical atomic property of Deoxyribonucleic Acid directly, reliably, and strictly dictates an essential corresponding biological survival function. For instance, the complete, strict absence of the 2′-hydroxyl group directly on the internal central ribose sugar immediately and brilliantly prevents a lethal steric clash. This tiny, subtle chemical modification enables the incredibly stable, life-preserving B-form helix to safely exist precisely inside the cell.
Furthermore, the beautiful, exact, strict hydrogen-bond complementarity embedded deeply within the protected internal data core guarantees absolute, perfect replication fidelity. The incredibly strict biological purine-pyrimidine physical structural geometries, specifically the precise A-T and robust G-C pairings, mathematically, completely, explicitly, and gracefully ensure that every single biological cellular replication cycle powerfully, beautifully, smoothly, efficiently, and completely produces an absolutely exact copy. This precise internal mathematical pairing is identically strictly completely cleanly deeply heavily required exactly for completely beautifully safely seamlessly flawlessly exact vital required RNA translation.
The distinct structural features of the complex helix provide highly effective and precise gene regulation. Its unique groove asymmetry plays a crucial role in this process, enabling specific biological functions. The carefully arranged, protective acidic backbone creates a prominent 2.2-nanometer groove that strategically exposes sequence-specific internal chemical edges. This exposure enables the effective binding of specific transcription factors to DNA, facilitating essential cellular processes.
The extremely powerful and continuously reinforced structural foundation of Deoxyribonucleic Acid (DNA) is characterized by its unique and profoundly effective design. This robust structure is highly negatively charged, providing essential stability and a secure environment for DNA packaging and survival. The combination of these features ensures that the DNA is not only protected but also functions effectively in its biological context.
Please read our Content Disclaimer Statement.
Check out our social media channels: