3. Biochemical Reactions and Principles of Energetics
Ever wonder how that morning cup of coffee or your breakfast bagel actually translates into the ability to run for the bus or ace a tough physiology exam? It all comes down to the microscopic, highly regulated universe operating inside your cells. As medical and science students, you are often tasked with memorizing overwhelming diagrams of metabolic pathways that look like complicated subway maps. But rote memorization is not the same as understanding.
The core purpose of this chapter is to bridge the gap between abstract thermodynamic physics and the tangible, life-sustaining molecular pathways inside your body. We are going to break down the specific mechanics, structural dynamics, and energetic flows that literally drive life itself. By mastering these principles, those complex pathway maps will finally start to make perfect, intuitive sense. Let’s dive into the fascinating, energetic machinery of the cell.
Slide 1: Introduction to Bioenergetics and Biochemical Reactions
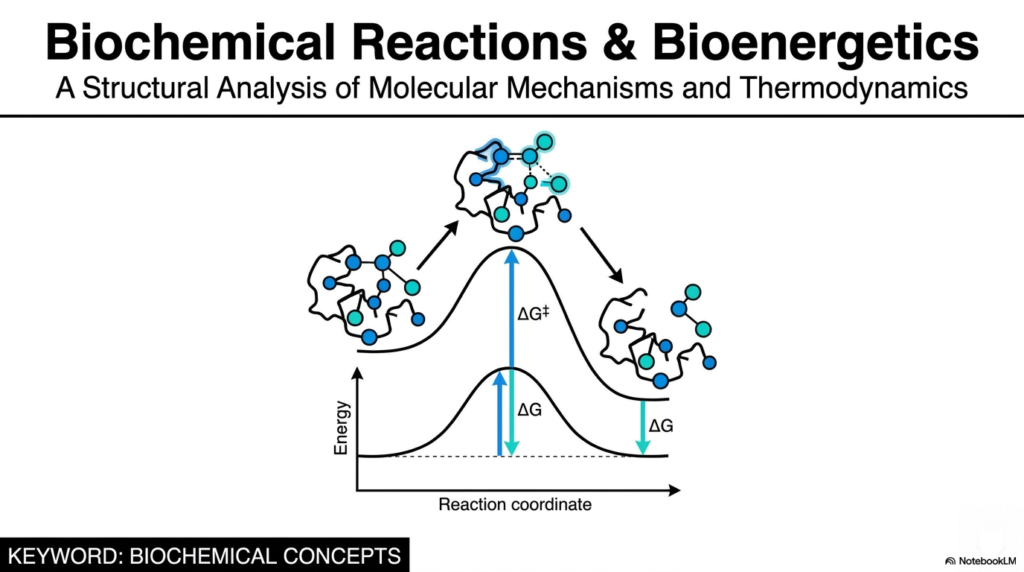
The visual on our first slide serves as the perfect entry point into the fundamental world of cellular thermodynamics and molecular architecture. As college and medical students, you will quickly learn that truly understanding biological systems requires looking beyond surface-level biology and diving deep into the realm of physical chemistry. This slide presents a stark, elegant visualization of a molecular structure perched precariously on a reaction coordinate energy curve. This visual is not just an abstract drawing; it is a structural analysis representing the core mechanisms of life. We are examining the foundational principles that govern how molecules behave during profound transformations.
To grasp how a cell functions, from the firing of a neuron to the contraction of a muscle fiber, you must view the cell as a bustling metropolis of continuous activity. Biochemical Reactions are the absolute currency of this microscopic metropolis. The sloping curve you see beneath the molecule represents the energy barrier—often called the activation energy—that chemical structures must overcome to transform from a starting material into a final product. It illustrates the profound difference between a spontaneous process that happens in a flash and one that requires the careful, specialized hand of a biological catalyst, known as an enzyme.
By studying both the precise physical shapes of these molecules (structural analysis) and the mathematical rules governing their energy states (thermodynamics), we unlock the secrets of metabolism, pharmacology, and gene expression. If you want to understand how a specific pharmaceutical inhibitor halts the rapid division of a cancer cell, or how a genetic defect causes a devastating metabolic storage disease, you must first grasp the fundamental energetics of these transformations. Therefore, this entire presentation is meticulously designed to break down these complex Biochemical Reactions into their most basic, digestible parts, ensuring you have a rock-solid foundation for the rest of your scientific or medical career. The journey begins with understanding how potential energy dictates cellular destiny.
Slide 2: The Power of Electron Transfer in Biochemical Reactions
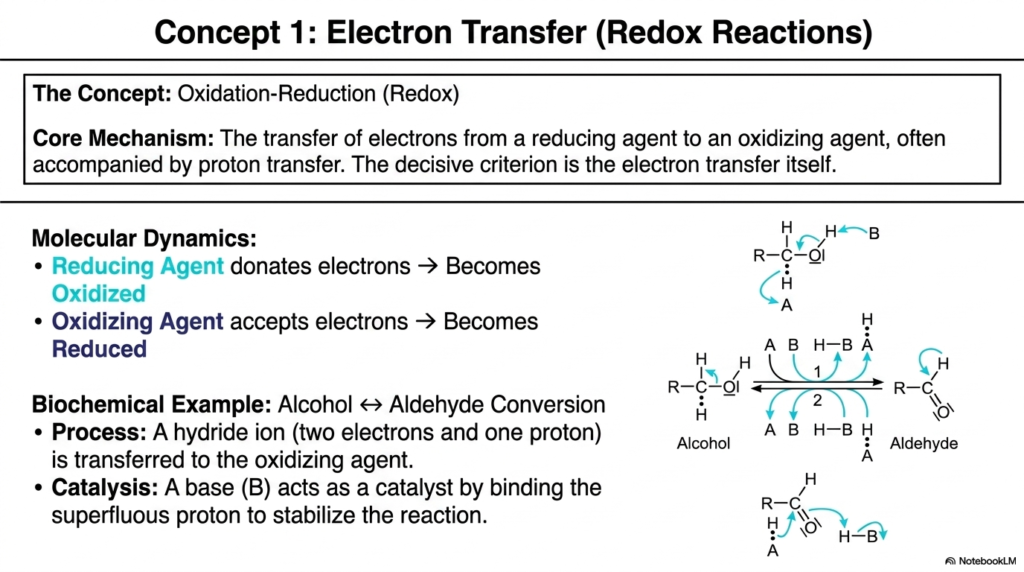
As we move into the specific mechanisms that drive cellular metabolism, we first encounter one of the most vital transformations in biology: oxidation-reduction, or redox for short. The core concept shown on this slide is the precise transfer of electrons from a reducing agent (which donates electrons and becomes oxidized) to an oxidizing agent (which accepts electrons and becomes reduced). If you want to understand how your body generates the vast majority of its ATP through the electron transport chain, you must first master this fundamental mechanism.
The molecular dynamics highlighted here show a classic biochemical example: the conversion between an alcohol and an aldehyde. Think about the clinical relevance of this process. When you consume a beverage containing ethanol, enzymes in your liver called alcohol dehydrogenases rely on these exact Biochemical Reactions to detoxify your blood. The diagram beautifully illustrates the transfer of a hydride ion—which consists of a proton carrying two extra electrons—directly to the oxidizing agent. This isn’t just protons floating around; the decisive criterion for this reaction class is the physical transfer of those negatively charged electrons.
Notice the careful orchestration required for this process to happen efficiently. A base labeled (B) in the diagram acts as a crucial catalyst. It binds to the superfluous proton left behind, stabilizing the entire reaction complex. Without this basic catalyst aiding proton removal, the energy barrier for electron transfer would be significantly higher. These specific electron-swapping Biochemical Reactions are the foundational energetic steps that allow us to harvest chemical energy from the food we eat. Every time a molecule of glucose is broken down in your cells, it undergoes a series of these precise redox steps, stripping high-energy electrons away to eventually power the cellular machinery that keeps you alive.
Slide 3: Mastering Proton Transfer in Acid-Base Biochemical Reactions
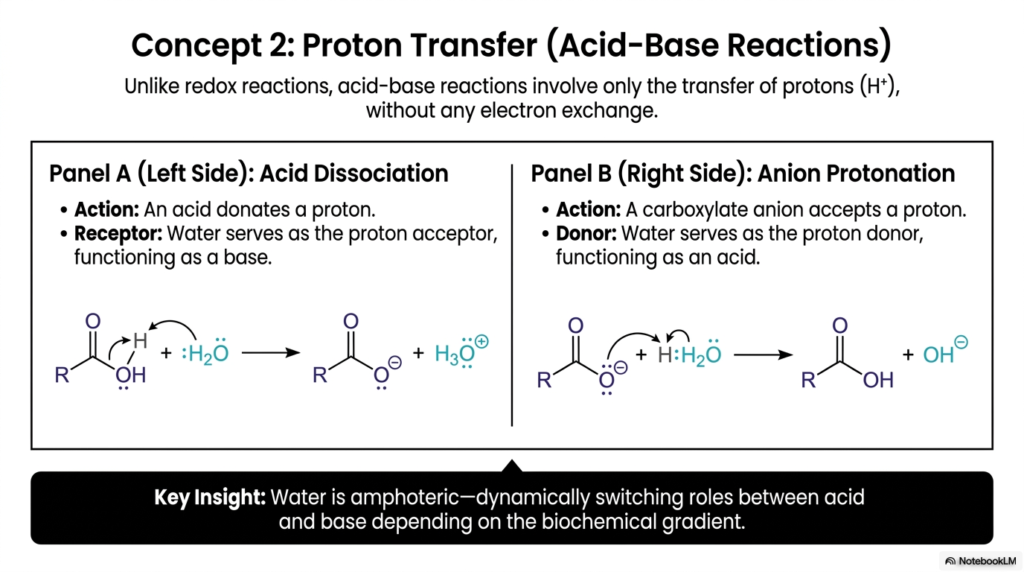
Unlike the electron-swapping dynamics we just explored, this slide introduces a completely different class of transformation: Acid-Base mechanisms. The defining characteristic here is that these processes involve only proton (H+) transfer, with no electron exchange. While redox reactions are about harvesting energy, acid-base processes are often about regulating the environment and preparing molecules for subsequent transformations. For medical students, understanding these dynamics is crucial to grasping concepts such as blood pH buffering, respiratory acidosis, and renal function.
The slide breaks this concept down into two distinct, highly informative panels. Panel A illustrates acid dissociation, where an acidic molecule donates a proton. In this scenario, water acts as the ultimate biochemical chameleon, serving as a proton acceptor and functioning as a base. This creates a hydronium ion. Conversely, Panel B shows the exact opposite: anion protonation. Here, a carboxylate anion accepts a proton, and water switches its role entirely, serving as the proton donor (the acid) to yield a hydroxide ion.
The key insight displayed at the bottom of the slide highlights this remarkable property of water. It is amphoteric, meaning it dynamically switches between acid and base states depending on the local biochemical gradient and the needs of the cellular environment.
This dynamic shifting is not just an interesting trivia fact; it is a matter of life and death. The intricate folded structures of your proteins and the catalytically active sites of your enzymes are held together by delicate electrostatic interactions that are highly sensitive to pH. If these specific proton-transferring Biochemical Reactions become unbalanced, proteins denature, enzymes fail, and cellular death ensues. Your body uses complex buffer systems, like the bicarbonate buffer in your blood, to constantly manage these Chemical Reactions, ensuring that the proton concentration remains within incredibly strict, life-sustaining physiological limits.
Slide 4: Building and Breaking: Multiple Bond Dynamics in Biochemical Reactions
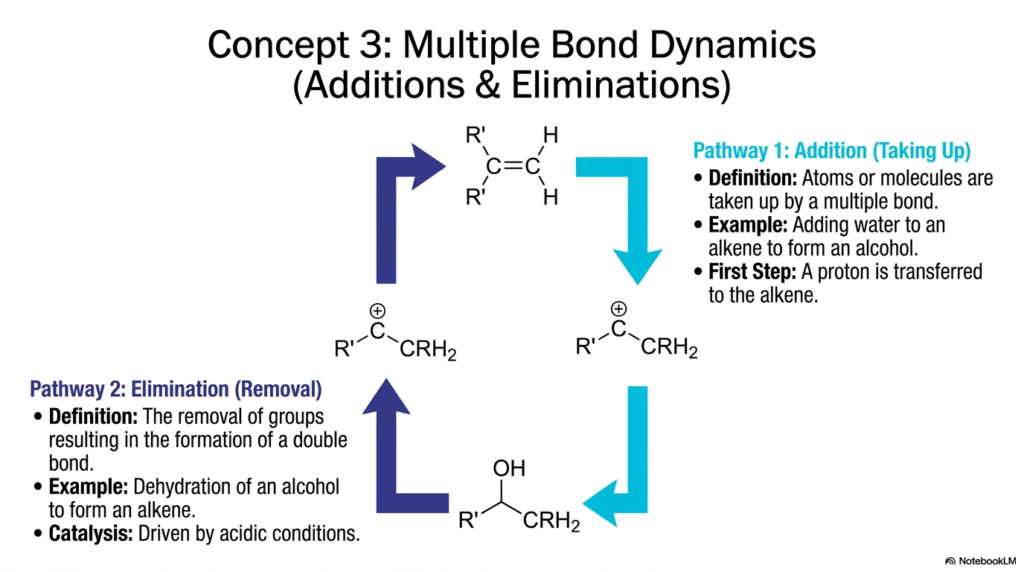
We now move from simple electron and proton transfers to the actual structural restructuring of carbon skeletons. Slide four introduces multiple bond dynamics, specifically focusing on the reciprocal processes of addition and elimination. These are the molecular Lego mechanics of the cell, enabling biological systems to build complex structures or break them down into functional components. Understanding these mechanisms is essential for grasping major metabolic pathways like fatty acid oxidation and the Krebs Cycle.
Let’s look at Pathway 1: Addition. By definition, this occurs when atoms or molecules are “taken up” by a multiple bond, effectively breaking a double bond to add new functional groups. The slide uses the classic example of adding water (hydration) to an alkene to form an alcohol. The first crucial step is the transfer of a proton to the alkene, which “primes” the molecule for the incoming water.
Now look at Pathway 2: Elimination. This is the exact reverse process. It involves the removal of chemical groups from a molecule, which results in the formation of a new double bond. A prime example is the dehydration of an alcohol to form an alkene, a process strongly catalyzed by acids.
These structurally altering Biochemical Reactions are completely reversible, as indicated by the continuous, cyclical arrows in the diagram. This reversibility is a hallmark of biological efficiency. Instead of inventing completely new pathways to break molecules down rather than build them up, cells often run the same machinery in reverse, depending on current metabolic demands. When your body is starved of energy, it uses elimination-style Chemical Reactions
to break down stored fats. When you have an excess of energy, it uses addition-based Chemical Reactions to store it for later use. This elegant, cyclic dynamic is a perfect example of nature’s biochemical economy.Slide 5: The Transient Danger of Carbenium Cations in Biochemical Reactions
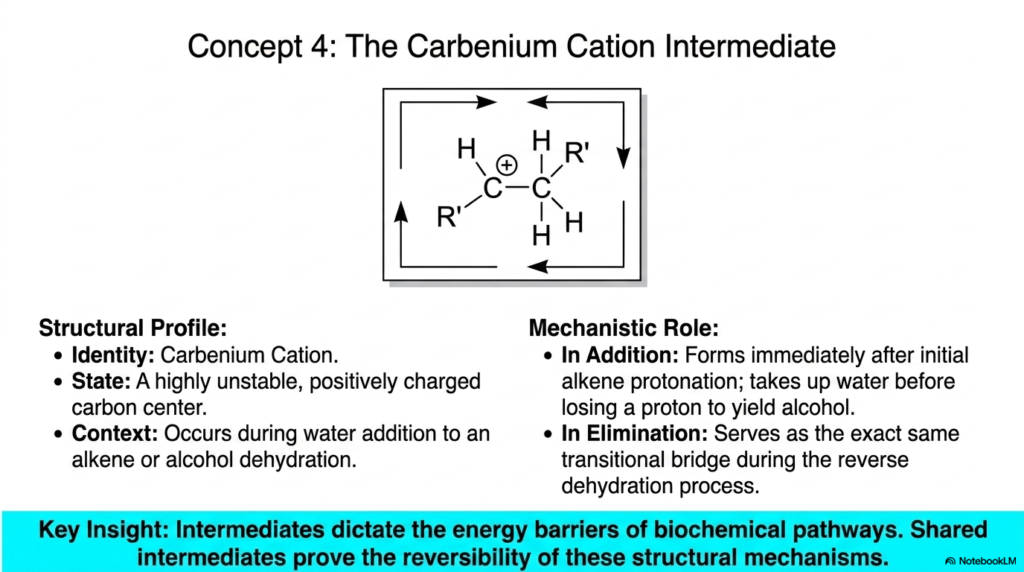
Slide five zooms in on the precarious transitional phases that molecules must endure during transformations. We are introduced to the Carbenium Cation Intermediate. In biochemistry, an intermediate is a fleeting, highly unstable molecular structure that exists momentarily between the starting materials and the final products. The carbenium cation is characterized by a positively charged carbon center, making it an incredibly reactive and dangerous species if left unchecked within the delicate cellular environment.
The structural profile on the slide shows that this intermediate forms precisely during the addition of water to an alkene or during alcohol dehydration—the very processes we discussed on the previous slide. Mechanistically, it forms immediately after the alkene is initially protonated. It must quickly take up water, then lose a proton to yield the stable alcohol product. Interestingly, it serves as the exact same transitional bridge during the reverse elimination process.
The key insight here is profound: shared intermediates prove the fundamental reversibility of these structural mechanisms. Because both the forward and backward paths must climb the same energy mountain and cross the same unstable peak, the pathway’s direction is dictated entirely by cellular concentrations and thermodynamics.
For medical students, understanding intermediates is vital for pharmacology. Many powerful drugs are designed specifically to mimic the shape of these unstable intermediates. By binding tightly to the enzymes that facilitate these specific Biochemical Reactions, the drugs permanently jam the machinery. Furthermore, these unstable intermediates are exactly why enzymes exist in the first place. Without enzymes providing a safe, stabilizing pocket for these highly reactive Chemical Reactions to occur, the energy barriers would simply be too high for life to exist at normal body temperatures. Enzymes are the cellular bodyguards that safely escort molecules through these dangerous transition states.
Slide 6: Swapping Parts via Nucleophilic Substitution Biochemical Reactions
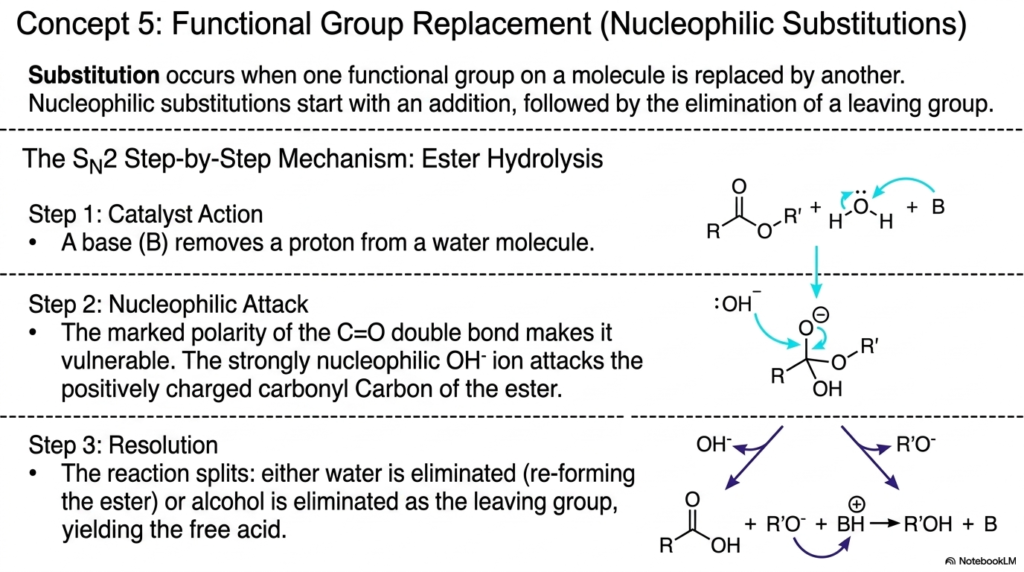
Now we explore one of the most common and versatile ways cells alter molecular structures: functional group replacement, clinically known as nucleophilic substitution. Substitution occurs when one functional group on a molecule is entirely replaced by another. This slide details the step-by-step mechanism of an SN2 (Substitution Nucleophilic Bimolecular) reaction, using ester hydrolysis as the prime example. This specific mechanism is how your digestive system breaks down dietary fats into usable fatty acids.
The slide breaks the process down into three distinct, highly choreographed steps. Step 1 involves the catalyst acting, where a base abstracts a proton from a water molecule. This seemingly simple act creates a highly reactive hydroxide (OH-) ion. Step 2 is the dramatic nucleophilic attack. The diagram highlights the marked polarity of the C=O double bond, which leaves the carbonyl carbon positively charged and highly vulnerable. The strongly nucleophilic, negatively charged OH- ion aggressively attacks this vulnerable carbon center. Step 3 is the resolution. The unstable intermediate splits, eliminating either water (reforming the original ester) or an alcohol as the leaving group, thereby yielding the free acid.
Understanding these specific step-by-step Biochemical Reactions is crucial for mastering biochemistry because they govern how macromolecules are constructed and dismantled. When ribosomes link amino acids together to build proteins, or when lipases cleave triglycerides in your intestines, they are relying on these exact nucleophilic mechanisms. By precisely controlling these part-swapping Chemical Reactions, your cells can continuously remodel their internal structures, repair damage, and manufacture the countless enzymes, hormones, and structural proteins required for healthy human physiology. It is cellular engineering at its finest.
Slide 7: The sp3-Hybridized Crossroads of Substitution Biochemical Reactions
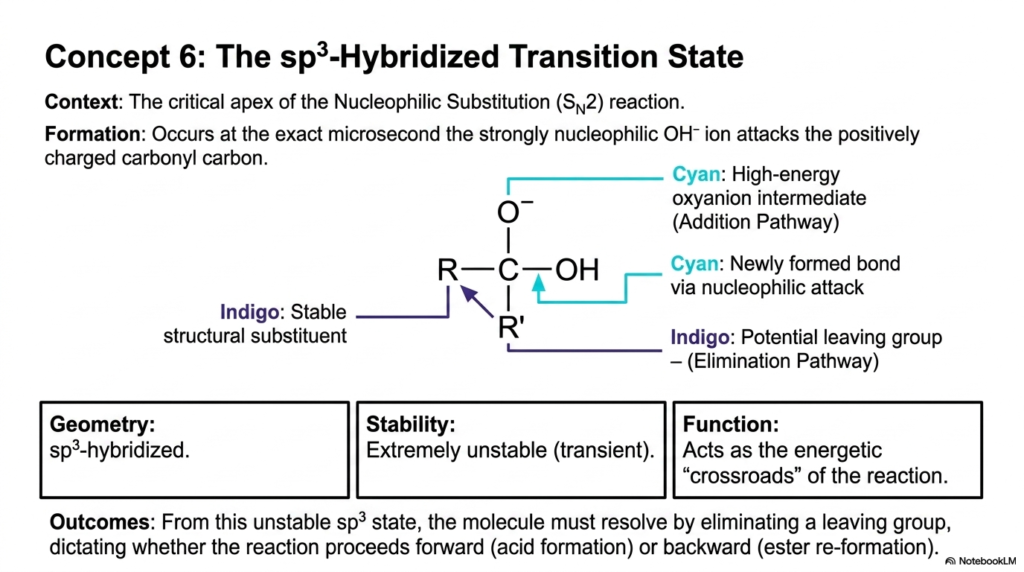
Following the nucleophilic attack discussed in the previous slide, we arrive at the absolute energetic apex of the substitution reaction: the sp3-hybridized transition state. This slide demands your attention because it visualizes the most critical microsecond in the lifespan of a reacting molecule. This is the exact moment when the strongly nucleophilic ion has fully engaged the positively charged carbonyl carbon, forcing the molecule into a highly strained, unnatural geometry.
Let’s break down the complex diagram. You can clearly see that the central carbon atom is forced to interact with five different groups, creating a high-energy oxyanion intermediate. We have the incoming newly formed bond (in cyan) from the nucleophile, the potential leaving group (in indigo), and the stable structural substituent. This state is called sp3-hybridized due to its specific tetrahedral-like geometry. The table clearly states that this structure is extremely unstable and transient. It cannot stay in this configuration. It acts as the ultimate energetic “crossroads” for the transformation.
From this incredibly unstable sp3 state, the molecule must rapidly resolve its geometry by eliminating a leaving group. This single, lightning-fast resolution dictates the entire outcome of these Biochemical Reactions. If it kicks off the newly attached nucleophile, the reaction goes backward, re-forming the starting ester.
If it kicks off the original leaving group, the reaction proceeds forward, creating the new acid product. In the realm of rational drug design, biochemists spend a great deal of time studying these specific transition states. If you can synthesize a drug molecule that permanently mimics this sp3-hybridized crossroads geometry, it will bind to the target enzyme with incredible affinity, effectively shutting down that specific subset of Chemical Reactions and potentially curing a disease or halting an infection.
Slide 8: Shapeshifting Molecules: Intramolecular Shifts in Biochemical Reactions
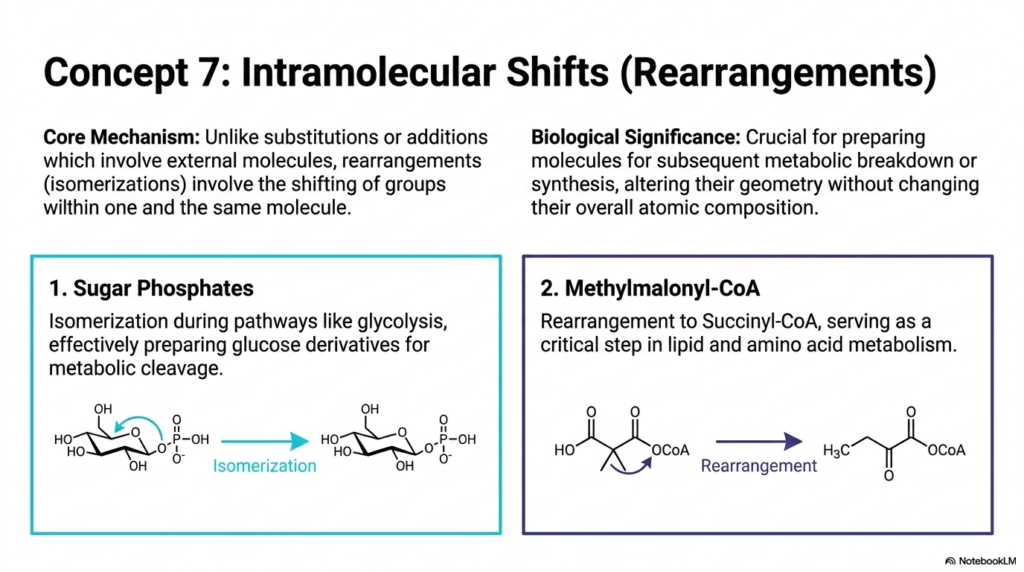
Sometimes, a cell doesn’t need to bring in external molecules or swap out leaving groups; it simply needs to reorganize the furniture within an existing structure. Slide eight introduces intramolecular shifts, also known as rearrangements or isomerizations. Unlike substitutions or additions that require external actors, these core mechanisms involve the shifting of chemical groups entirely within one and the same molecule. This might seem like a trivial change, but it has massive biological significance.
These spatial reorganizations are absolutely crucial for preparing molecules for subsequent metabolic breakdown or synthesis. By altering their geometry without changing their overall atomic composition, molecules are “primed” to fit perfectly into the active sites of the next enzyme in a pathway. The slide provides two classic, high-yield biological examples. First, we see the isomerization of Sugar Phosphates during glycolysis. By converting a six-membered sugar ring into a five-membered ring, the cell is effectively preparing the glucose derivative so it can be symmetrically cleaved in half later in the pathway. Without this specific rearrangement, energy extraction from sugar would grind to a halt.
The second example is the rearrangement of Methylmalonyl-CoA into Succinyl-CoA. This is a critical step in both lipid and amino acid metabolism, and interestingly, it is a pathway that strictly requires Vitamin B12 as a cofactor. If a patient has a B12 deficiency, these specific Biochemical Reactions fail, leading to a dangerous buildup of methylmalonic acid in the blood and severe neurological consequences. Understanding these shapeshifting Chemical Reactions allows medical professionals to connect the dots between microscopic structural isomerism and macro-level clinical symptoms, thereby demonstrating precisely why molecular geometry dictates human health.
Slide 9: The Master Matrix of Biochemical Reactions
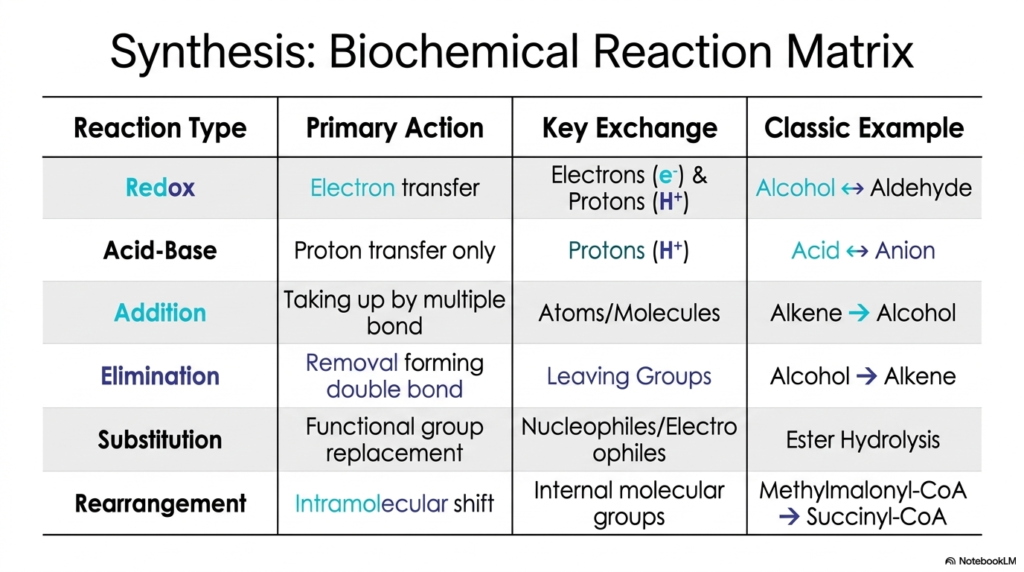
If you are a student overwhelmed by the sheer volume of metabolic pathways you have to memorize, this slide is your new best friend. Slide nine presents the Synthesis Biochemical Reaction Matrix, a phenomenal cheat sheet that categorizes the vast, seemingly chaotic world of cellular metabolism into six clean, logical buckets. Instead of trying to memorize ten thousand distinct transformations, you simply need to understand these six fundamental categories.
The table beautifully summarizes everything we have covered so far. We have Redox reactions that primarily involve electron transfer (like the alcohol-to-aldehyde shift). Acid-Base reactions strictly manage proton transfers. Addition reactions add atoms across multiple bonds, and their counterpart, Elimination reactions, remove groups to form multiple bonds. We covered Substitution, where functional groups are replaced by nucleophiles, and finally Rearrangements, which represent internal intramolecular shifts.
By categorizing metabolism in this way, you can look at any complex pathway chart—whether it’s the urea cycle, purine synthesis, or beta-oxidation—and instantly demystify what is happening at each step. You stop seeing a random jumble of letters and arrows, and start seeing a logical sequence of these six core Biochemical Reactions. This matrix represents the master framework for mastering biochemistry. It proves that despite the staggering complexity of the human body, the underlying biological engineering relies on a beautifully limited, highly efficient set of standardized Chemical Reactions to sustain all of life’s processes.
Slide 10: The Physics of Potential in Biochemical Reactions
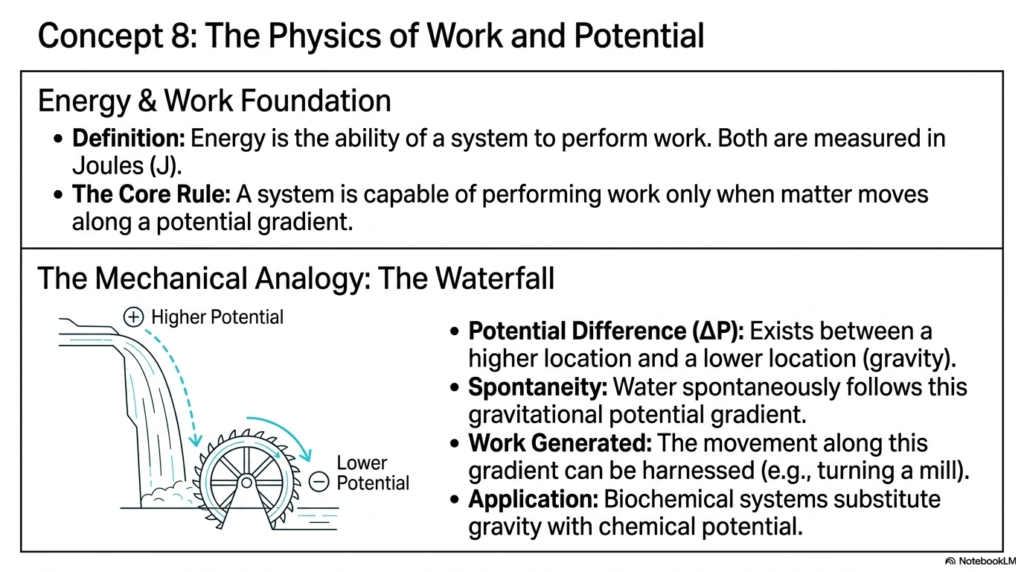
Having thoroughly explored the structural mechanisms, we must now shift our focus to the driving forces that actually drive these transformations. Structure is only half the story; thermodynamics is the other. Slide ten introduces the Physics of Work and Potential. The foundation of this concept is that energy is defined simply as the ability of a system to perform work (both measured in Joules). But the core rule you must remember is this: a system can perform work only when matter moves down a potential gradient.
To make this abstract physics concept intuitive, the slide uses a brilliant mechanical analogy: the waterfall. Imagine water flowing over a cliff to turn a wooden watermill. A potential difference ($\Delta$P) exists between the higher location and the lower location due to gravity. Because of this gradient, the water spontaneously flows downward under the gravitational pull. As it falls, this movement can be harnessed by the wheel to generate usable work. If the water were just sitting flat in a pond, no work could be done.
Biological systems do not rely on gravity to turn tiny molecular wheels; instead, they substitute chemical potential for gravity. Molecules “fall” from states of high chemical potential to states of lower chemical potential. It is this exact downhill movement that the cell harnesses to generate power. Every single time your heart beats or your brain forms a memory, it is being powered by the microscopic work generated by these downhill Biochemical Reactions. Understanding this gradient is the key to understanding how cells harvest the energy required to fight off entropy and maintain the highly organized state of a living organism.
Slide 11: Intensity vs. Capacity Factors in Biochemical Reactions

To deeply understand thermodynamics, you must be able to distinguish between two highly specific dimensions of work: the Intensity factor and the Capacity factor. Slide eleven breaks down this crucial, and often misunderstood, distinction. This concept explains not just if a reaction will happen, but exactly what is driving it and how much stuff is involved.
Let’s look at the definitions. The Intensity factor is the measure of the potential difference; it is the sheer “driving force” of the process. In our waterfall analogy, this is the height difference of the cliff. In electrical systems, it is the voltage. In biological chemistry, the intensity factor is the Chemical potential, specifically measured as Free Enthalpy. Conversely, the Capacity factor measures the amount of the substance being transported or reacted. Mechanically, this is the total weight of the water falling. Electrically, it’s the amount of charge flowing (current). Chemically, it is the actual amount of reacting matter, typically measured in moles.
Why is this vital for medical students? Because it teaches you that sheer quantity does not dictate spontaneity. You could have a massive capacity factor—say, millions of moles of a specific reactant sitting in a cell—but if the intensity factor (the chemical potential gradient) isn’t favorable, absolutely nothing will happen. The molecules will just sit there. To advance biological pathways, the cell must continually adjust the intensity factor. By altering the concentrations of products and reactants, the cell manages the driving forces behind these vital Biochemical Reactions, ensuring that metabolism continues in the correct, life-sustaining direction.
Slide 12: Decoding Chemical Potential and Free Enthalpy in Biochemical Reactions

Building directly on the concept of the intensity factor, slide twelve introduces the most important mathematical variable in all of biochemistry: Gibbs Free Energy, or the Change in Free Enthalpy. This is the mathematical representation of the chemical potential we’ve been discussing. Free Enthalpy (G) represents the absolute chemical potential of a specific molecule or combination of molecules.
The driving force of any biological process is governed by a simple rule: when molecules spontaneously react, the products must end up at a lower overall potential than the reactants. The mathematical difference in chemical potentials between the starting educts (the reactants) and the final products is defined as ΔG. However, there is a major measurement constraint. We cannot directly determine the absolute or total values of G in a lab. There is no biological speedometer for absolute enthalpy. Instead, biochemists must calculate ΔG mathematically using the reaction’s equilibrium constant. By seeing where a reaction naturally settles, we can retroactively calculate the exact intensity of the driving force.
Understanding ΔG is like having a metabolic crystal ball. It allows scientists and doctors to examine any theoretical pathway and immediately predict whether it is physically possible under cellular conditions. It represents the ultimate judge, jury, and executioner for biological pathways. If a drug is designed to force a pathway to run in a direction that violates the rules of Free Enthalpy, that drug will fail. Mastering the concept of $\Delta$G allows you to truly understand the absolute energetic boundaries that govern all Biochemical Reactions inside the human body.
Slide 13: Predicting the Spontaneity of Biochemical Reactions

Slide thirteen cuts to the absolute core of biochemical thermodynamics by defining Reaction Spontaneity based on the principles of Free Enthalpy (ΔG). The slide opens with a perfect, intuitive rule: “Water never flows uphill spontaneously.” In biology, spontaneity depends entirely on the potential difference between the final and initial states (ΔP = P2 – P1). This gives us three possible metabolic scenarios that dictate every aspect of cellular life.
First, we have Exergonic Processes. Here, the final state has less energy than the initial state, meaning ΔP (or ΔG) is negative. The outcome? The process occurs spontaneously and releases energy, performing valuable cellular work. This is the biological equivalent of water rushing down the waterfall. Second, we have Equilibrium, where the final state equals the initial state (ΔP = 0). The system is totally static; there is no net potential difference, and zero work can be done. For a living cell, reaching total equilibrium means death. Life is a constant struggle to stay away from equilibrium!
Finally, we have Endergonic Processes. These are the “uphill” battles where the final state requires more energy than the initial state (ΔP is positive). The outcome is that these processes do not proceed spontaneously; they strictly require a massive influx of external energy. Think about building a complex muscle fiber or copying a massive strand of DNA—these are highly endergonic tasks. Understanding the difference between these states is vital because it explains human nutrition. We must eat food to drive exergonic Chemical Reactions, which in turn give us the power to force those difficult, endergonic Chemical Reactions up the hill.
Slide 14: The Magic of Energetic Coupling in Biochemical Reactions
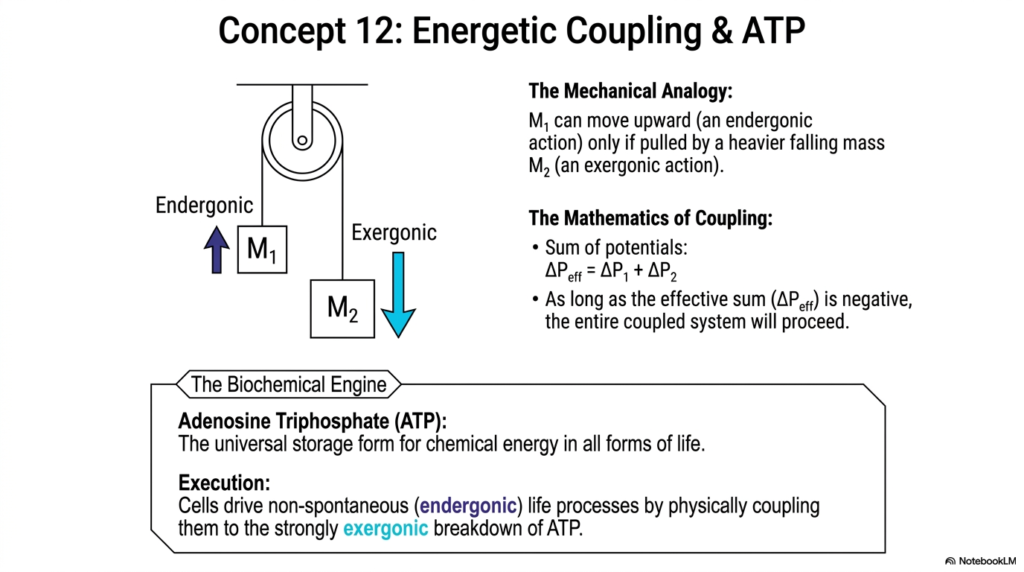
If endergonic (uphill) processes cannot happen spontaneously, how do cells ever manage to build complex structures, divide, or move? Slide fourteen reveals the greatest magic trick in all of biology: Energetic Coupling and ATP. To explain this, the slide uses a brilliant mechanical analogy of a pulley system. Imagine trying to lift a heavy block (Mass 1) upward. This is an endergonic action; it won’t happen on its own. But, if you tie it via a pulley to an even heavier, falling block (Mass 2), the exergonic downward pull of the heavy block easily drags the lighter block upward.
The mathematics of coupling are beautifully simple. You sum the potentials: ΔP effective = ΔP1 + ΔP2. As long as the effective sum remains negative overall, the entire coupled system will proceed smoothly. In the biochemical engine of your body, that heavy, falling block is almost exclusively Adenosine Triphosphate (ATP). ATP is the universal storage form for chemical energy in all forms of cellular life. It is the charged battery of the biological world.
Execution is everything. Cells drive absolutely vital, non-spontaneous life processes by physically and chemically coupling them to the violently, strongly exergonic breakdown of ATP. When ATP is hydrolyzed, it releases a massive burst of energy. Enzymes are designed to trap this burst and use it to force an endergonic reaction up the energy hill. Without this specific trick of coupling these exact Chemical Reactions together, complex multicellular life would be thermodynamically impossible. We literally survive by linking our metabolic machinery to the continuous, exergonic destruction of ATP.
Slide 15: Synthesizing the Bioenergetic Framework of Biochemical Reactions
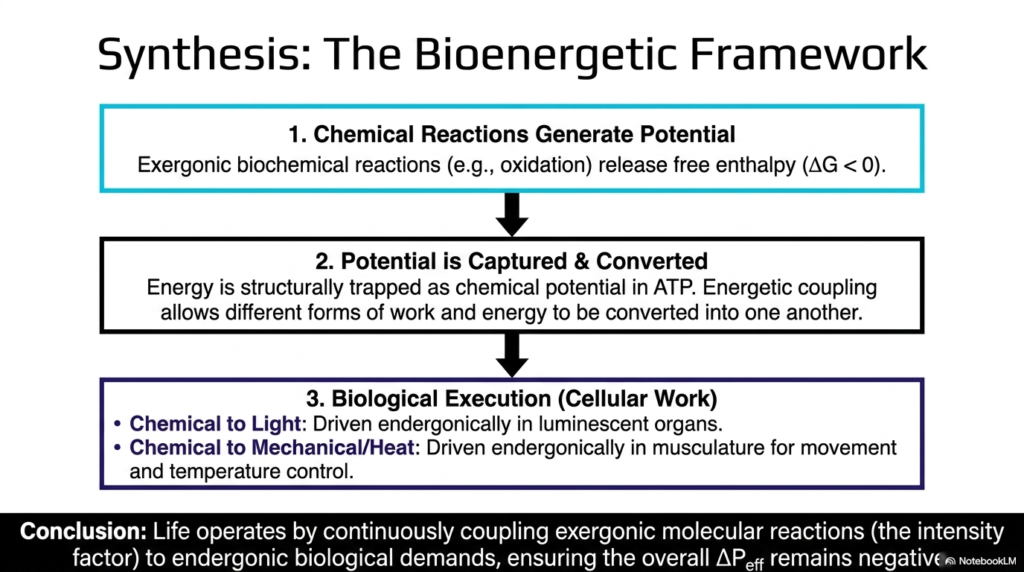
We have reached the grand finale. Slide fifteen brings together every structural mechanism and thermodynamic principle we have discussed to synthesize the complete Bioenergetic Framework of life. This is the overarching cycle that explains how biological entities harvest, store, and utilize energy to survive in a chaotic universe. The framework operates in a continuous, unbroken three-step sequence.
Step 1: Chemical Reactions Generate Potential. We consume fuel (food), and through highly exergonic biochemical mechanisms such as redox reactions, we break that fuel down, releasing large amounts of free energy (ΔG < 0). We are effectively creating a rushing chemical waterfall. Step 2: Potential is Captured and Converted. The cell cannot let that energy just dissipate as heat. Instead, the energy is structurally trapped as chemical potential inside ATP molecules. Through the magic of energetic coupling, different forms of work and energy are converted into this single, universal currency.
Step 3: Biological Execution (Cellular Work). The stored ATP is finally used to meet the deep endergonic demands of life. The slide highlights amazing examples: converting chemical energy into light (like fireflies lighting up the night sky), or converting chemical energy into mechanical force and heat (like your muscles contracting to help you run or shivering to keep you warm in the cold).
The ultimate conclusion is profound: life operates by continuously coupling the exergonic breakdown of molecules to the endergonic demands of survival, ensuring that the overall effective potential difference remains negative. Mastering these foundational Chemical Reactions is not just about passing a biochemistry exam; it is about understanding the very machinery that allows us to exist, adapt, and thrive.
Check out our social media channels:
Please read our Content Disclaimer Statement.