42. Glycogen Metabolism: Structural Principles, Pathways, and Clinical Significance
Ever wonder how a sprinter bursts off the starting line or how the human brain survives the long fast between dinner and breakfast? It all comes down to cellular sugar stores. This comprehensive slide deck explores the dynamic biochemical pathways of energy storage and retrieval. By examining structural properties, enzymatic mechanisms, and intricate regulatory networks, students will uncover the profound physiological importance of these carbohydrate reserves. Furthermore, this presentation highlights exactly what happens when these vital metabolic systems fail in human disease, bridging the gap between basic biochemistry and clinical pathology.
Slide 1: Introduction to Glycogen Metabolism
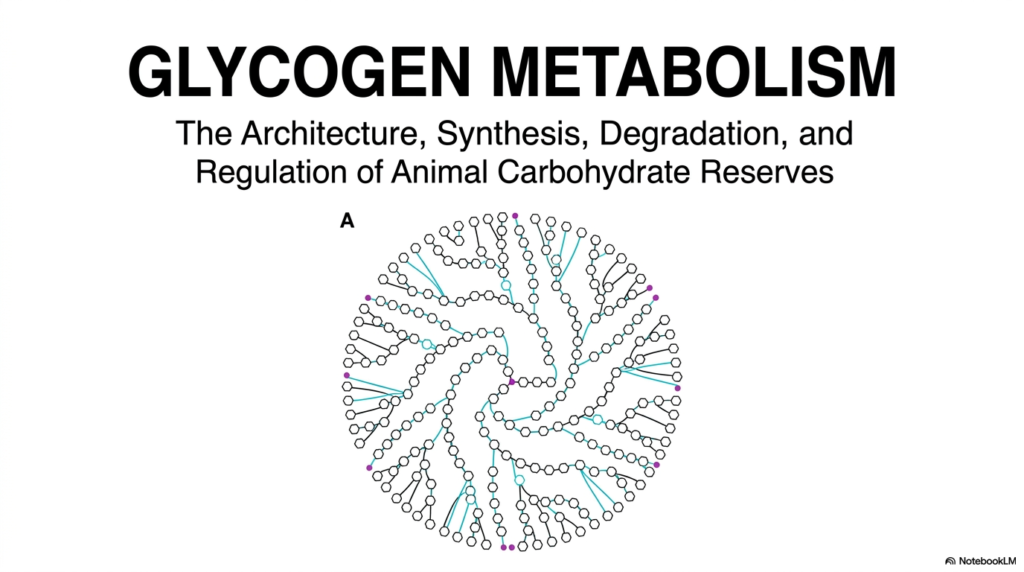
Welcome to the foundational overview of Glycogen Metabolism. This initial slide introduces the massive, beautifully complex architecture of animal carbohydrate reserves. As depicted in the central illustration, this molecule is not a random clump of sugars, but a highly organized, spherical polymer. The intricate branching pattern allows mammalian cells to store immense amounts of energy in a remarkably compact, rapidly accessible form. Understanding Glycogen Metabolism begins with appreciating this structural elegance, which perfectly dictates its biochemical function. This biological architecture represents a pinnacle of evolutionary engineering, specifically designed to sustain life during routine fasting.
The title slide explicitly breaks down the core pillars of Glycogen Metabolism: architecture, synthesis, degradation, and regulation. Each of these foundational pillars represents a meticulously controlled biochemical phase within the cell. Synthesis involves building this complex structural tree during times of nutritional abundance. Conversely, degradation is the rapid mobilization of these crucial sugar reserves when cellular energy drops. The interplay between these opposing pathways is tightly governed by systemic hormones and local metabolites. This dynamic balance ensures that glucose remains continually available for all vital physiological functions.
Finally, the central image of the slide hints at the profound molecular symmetry inherent to Glycogen Metabolism. Notice the dense central core and the sprawling branches radiating outward. This specific spherical geometry is biologically intentional, maximizing the number of non-reducing ends at the surface. By maximizing these exposed sugar ends, enzymes can rapidly and simultaneously cleave glucose molecules when systemic energy demand peaks. This architectural marvel is the absolute cornerstone of how mammalian systems manage their carbohydrate reserves, setting the stage for the detailed enzymatic mechanisms explored next.
Slide 2: Structural Principles of Glycogen Metabolism
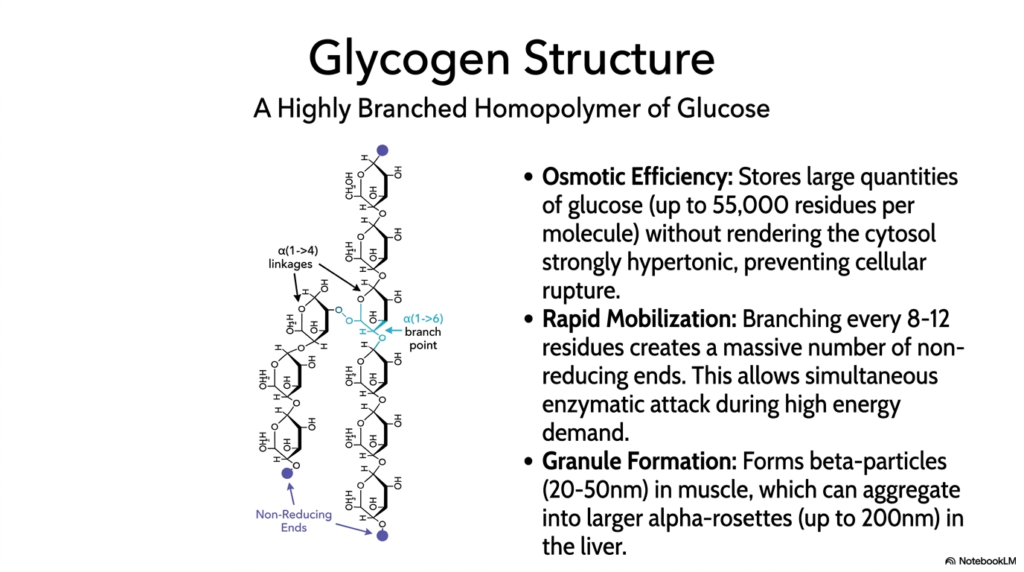
The molecular structure highlighted in this slide is fundamental to the efficiency of Glycogen Metabolism. The molecule is a highly branched homopolymer composed entirely of glucose residues. The linear chains are linked by alpha(1->4) glycosidic bonds, while the crucial branch points are formed by alpha(1->6) linkages. This specific biochemical arrangement creates a massive macromolecule capable of holding up to 55,000 glucose residues. Storing sugar in this polymerized form is a brilliant evolutionary adaptation, solving a major thermodynamic problem for the cell while maintaining a ready supply of fuel.
Osmotic efficiency is a central theme in Glycogen Metabolism. If a cell attempted to store 55,000 individual glucose molecules freely in the cytosol, the resulting osmotic pressure would render the intracellular environment strongly hypertonic, causing water to rush in and rupture the cell. Polymerizing glucose into a single large granule drastically reduces this osmotic threat. Furthermore, these molecules assemble into distinct granules: smaller beta-particles in skeletal muscle and larger alpha-rosettes in the liver. This packaging ensures that massive energy reserves can sit safely within the cytoplasm without disrupting cellular integrity.
The branching structure also directly facilitates the rapid mobilization required during active Glycogen Metabolism. Because a branch occurs every 8 to 12 residues, the molecule possesses a massive number of terminal, non-reducing ends. During periods of high energy demand, degrading enzymes simultaneously attack these numerous ends, releasing thousands of glucose molecules in milliseconds. Without this heavy branching, enzymes could only chew away at a single end, and the release of energy would be far too slow to sustain intense muscular contraction or maintain blood sugar levels during a fast.
Slide 3: Glycogenin and the Core of Glycogen Metabolism
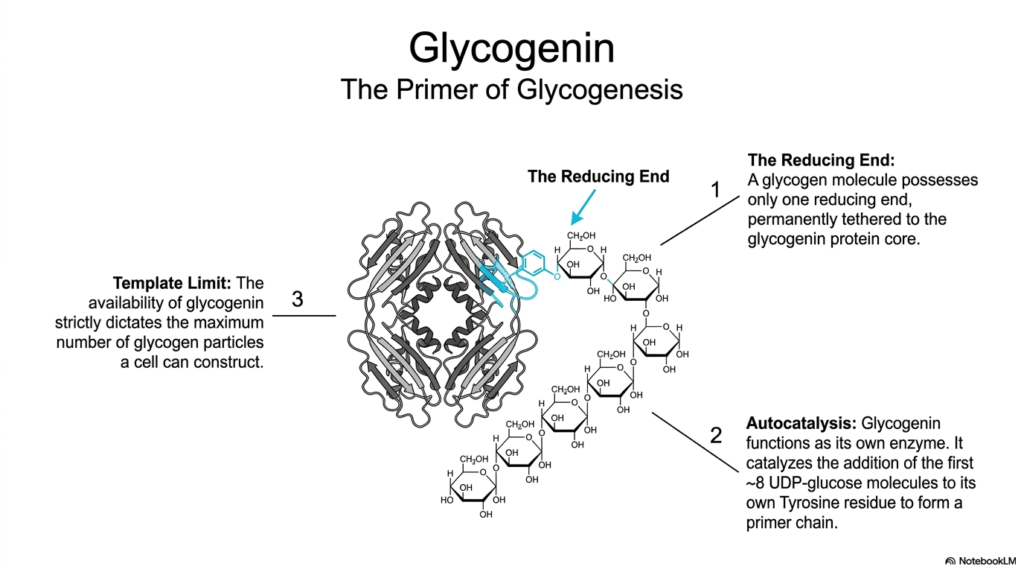
Every monumental structure needs a solid foundation, and in Glycogen Metabolism, that foundation is a protein called glycogenin. The slide illustrates this crucial homodimeric protein sitting at the absolute center of the carbohydrate molecule. Glycogenin acts as the essential primer for glycogenesis, providing the starting point for the massive glucose polymer. Interestingly, a fully formed glycogen molecule has only one reducing end, and this end remains permanently covalently tethered to the glycogenin protein core throughout the molecule’s lifespan in the cell.
A fascinating aspect of Glycogen Metabolism is the autocatalytic nature of glycogenin. It functions as its own enzyme, actively catalyzing the addition of the first few glucose molecules to itself. Specifically, glycogenin uses UDP-glucose to add approximately 8 glucose residues to a specific tyrosine residue on its own surface. This short, self-assembled oligosaccharide chain then serves as the necessary primer for other elongating enzymes to take over. Without this initial self-priming step by glycogenin, the cell is unable to initiate the synthesis of new carbohydrate stores.
This initial priming step establishes a strict template limit for Glycogen Metabolism within any given cell. Because every single carbohydrate granule requires exactly one glycogenin core to begin synthesis, the total availability of glycogenin proteins strictly dictates the maximum number of individual granules a cell can ever construct. This limitation represents a brilliant form of cellular regulation, ensuring that the cell does not over-invest its resources into creating too many storage particles, thereby maintaining an optimal balance between free energy and stored reserves.
Slide 4: Tissue-Specific Roles in Glycogen Metabolism
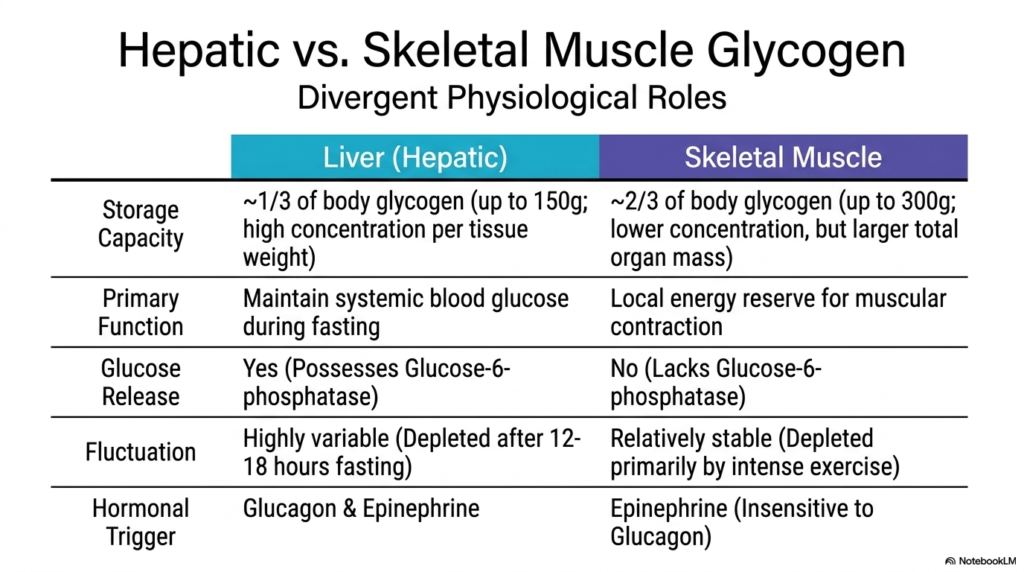
The physiological application of Glycogen Metabolism varies drastically depending on the specific tissue in question. This slide contrasts the two primary storage sites in the human body: the liver and skeletal muscle. While the fundamental biochemistry remains similar, their overarching physiological roles are completely divergent. The liver holds roughly one-third of the body’s total carbohydrate reserves, maintaining a very high concentration of the polymer per gram of tissue. In contrast, skeletal muscle contains the remaining two-thirds, dispersed across a much larger total organ mass throughout the human body.
In hepatic Glycogen Metabolism, the primary biological directive is purely altruistic: maintaining systemic blood glucose levels during periods of fasting. The liver possesses a unique enzyme called glucose-6-phosphatase, which allows it to strip the phosphate group off glucose and release the free sugar directly into the bloodstream for use by other organs, particularly the brain. Consequently, hepatic stores are highly variable, aggressively building up after a carbohydrate-rich meal and becoming completely depleted after just 12 to 18 hours of typical fasting.
Conversely, skeletal muscle Glycogen Metabolism is entirely selfish, serving as a dedicated, local energy reserve strictly for muscular contraction. Skeletal muscle fundamentally lacks the glucose-6-phosphatase enzyme. Therefore, once glucose enters a muscle cell and is stored, it can never leave; it is completely trapped for local use. Because of this, muscle carbohydrate reserves are relatively stable compared to those in the liver, remaining largely intact during normal fasting and only becoming significantly depleted during intense, prolonged physical exercise, when local ATP demands skyrocket.
Slide 5: Glucose Activation in Glycogen Metabolism
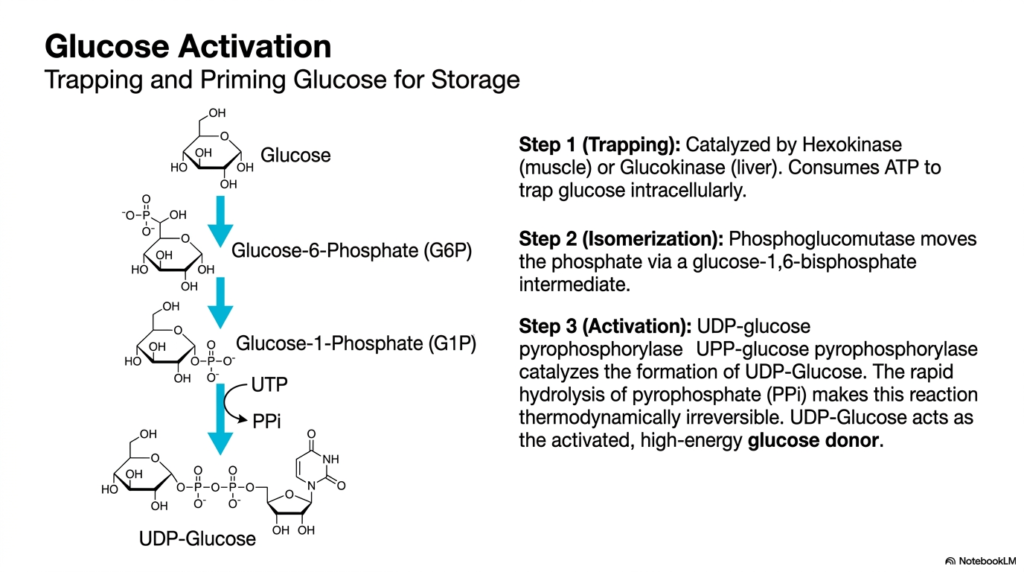
Before synthesis can begin, free glucose must be properly prepared, a critical preparatory phase in Glycogen Metabolism. This slide details the process of glucose activation, which involves trapping the sugar inside the cell and priming it for polymerization. The first step is catalyzed by hexokinase in muscle or by glucokinase in the liver, using cellular ATP to phosphorylate free glucose to Glucose-6-Phosphate. This added phosphate group gives the molecule a negative charge, effectively trapping it inside the cellular membrane so it cannot escape back into the bloodstream.
Following this initial trapping, Glycogen Metabolism requires an isomerization step. The enzyme phosphoglucomutase shifts the phosphate group from the sixth carbon to the first carbon, converting Glucose-6-Phosphate into Glucose-1-Phosphate. This reaction proceeds through a temporary glucose-1,6-bisphosphate intermediate. Moving the phosphate to the first carbon positions it in the precise chemical geometry required for the final activation step. Without this subtle yet vital spatial rearrangement, the sugar molecule would remain incompatible with the subsequent enzymes that build the larger polymer chain.
The crowning step of activation in Glycogen Metabolism is the creation of UDP-glucose, the high-energy donor molecule. The enzyme UDP-glucose pyrophosphorylase catalyzes the reaction between Glucose-1-Phosphate and UTP. A critical thermodynamic feature of this step is the release and subsequent rapid hydrolysis of pyrophosphate. This immediate breakdown of pyrophosphate releases significant energy, making the entire activation reaction thermodynamically irreversible. By creating UDP-glucose, the cell successfully generates an activated, high-energy building block perfectly primed for transfer onto a growing carbohydrate chain.
Slide 6: Elongation and Branching in Glycogen Metabolism
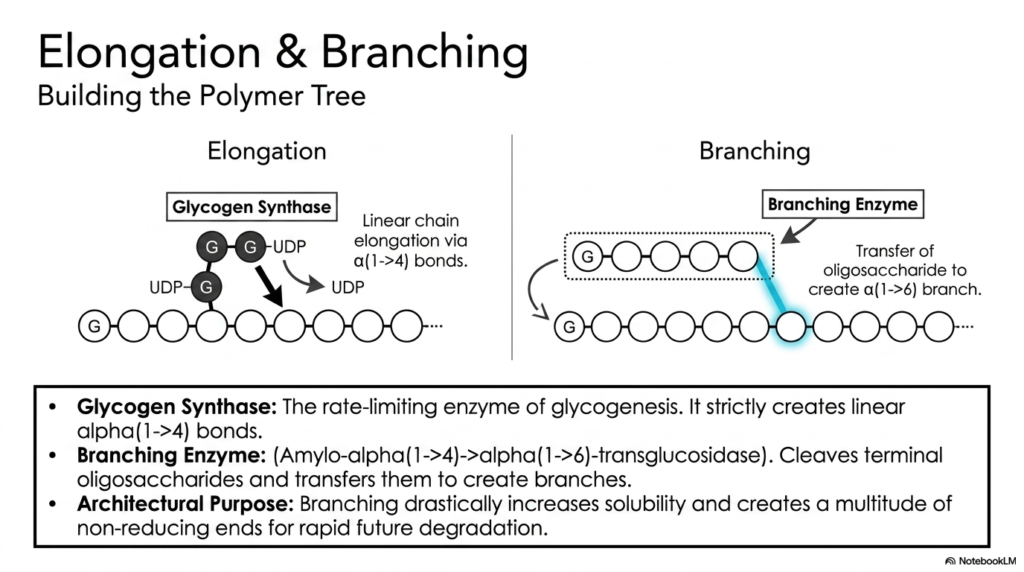
With activated UDP-glucose ready, the active construction phase of Glycogen Metabolism commences. This slide illustrates the dual processes of elongation and branching, which build the massive polymer tree. The primary engine of elongation is Glycogen Synthase, the crucial rate-limiting enzyme in the entire biosynthetic pathway. This enzyme continuously transfers the activated glucose from UDP-glucose to the non-reducing end of an existing chain, creating strictly linear alpha(1->4) glycosidic bonds. However, Glycogen Synthase is entirely incapable of creating branches on its own.
To construct a structurally sound molecule, Glycogen Metabolism relies heavily on the Branching Enzyme. Also known by its complex biochemical name, amylo-alpha(1->4)->alpha(1->6)-transglucosidase, this enzyme performs a remarkably specialized task. Once a linear chain grows sufficiently long, the branching enzyme cleaves a terminal oligosaccharide block of roughly seven residues. It then physically transfers this entire block to an internal location on the same or a neighboring chain, reattaching it via a new alpha(1->6) glycosidic bond to establish a distinct branch point.
The architectural purpose of this branching process is absolutely vital to functional Glycogen Metabolism. Creating these complex branches dramatically increases the overall solubility of the massive macromolecule in the cellular cytoplasm. More importantly, every new branch created adds another terminal, non-reducing end to the molecule. As previously noted, maximizing these non-reducing ends is biologically critical because it ensures that degrading enzymes have multiple target sites for rapid, simultaneous glucose mobilization when energy is urgently required.
Slide 7: Phosphorolytic Cleavage in Glycogen Metabolism
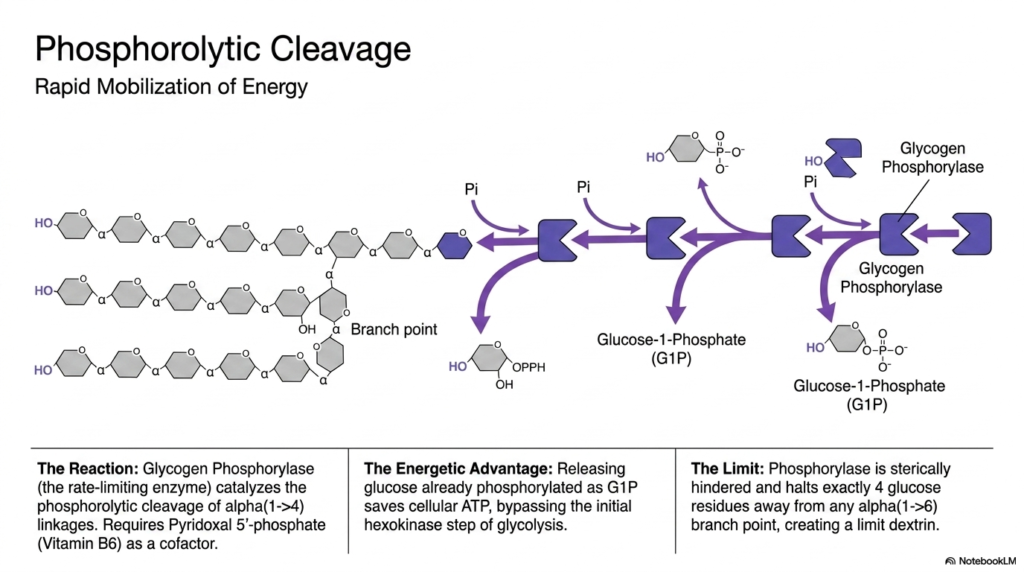
When energy demands surge, the degradative phase of Glycogen Metabolism initiates with phosphorolytic cleavage. This rapid mobilization is driven by Glycogen Phosphorylase, the rate-limiting enzyme of the breakdown pathway. Instead of simply using water to hydrolyze the bonds, this enzyme uses inorganic phosphate to cleave the linear alpha(1->4) linkages occurring at the non-reducing ends of the branches. This specialized phosphorolysis reaction specifically yields Glucose-1-Phosphate. To function correctly, the phosphorylase enzyme absolutely requires the presence of Pyridoxal 5′-phosphate, a derivative of Vitamin B6, as an essential catalytic cofactor.
The energetic advantage of utilizing phosphorolysis in Glycogen Metabolism is remarkably elegant. By cleaving the polymer with an inorganic phosphate rather than water, the enzyme releases the sugar already phosphorylated as Glucose-1-Phosphate. This clever biochemical maneuver entirely bypasses the initial, ATP-consuming hexokinase step of standard glycolysis. Consequently, glucose derived directly from stored carbohydrate reserves yields a higher net ATP yield during cellular respiration than free glucose entering from the bloodstream, representing a highly optimized evolutionary strategy for rapid energy generation.
However, a significant structural limitation exists within this phase of Glycogen Metabolism. Glycogen Phosphorylase is a large, bulky enzyme that experiences severe steric hindrance as it chews down a branch and approaches an alpha(1->6) branch point. Because of its physical size, the enzyme automatically halts its cleavage activity exactly four glucose residues away from any branching linkage. This enzymatic roadblock leaves behind a heavily trimmed, stubby structure biochemically referred to as a “limit dextrin,” which requires a completely different set of enzymatic tools to resolve.
Slide 8: The Debranching Process in Glycogen Metabolism
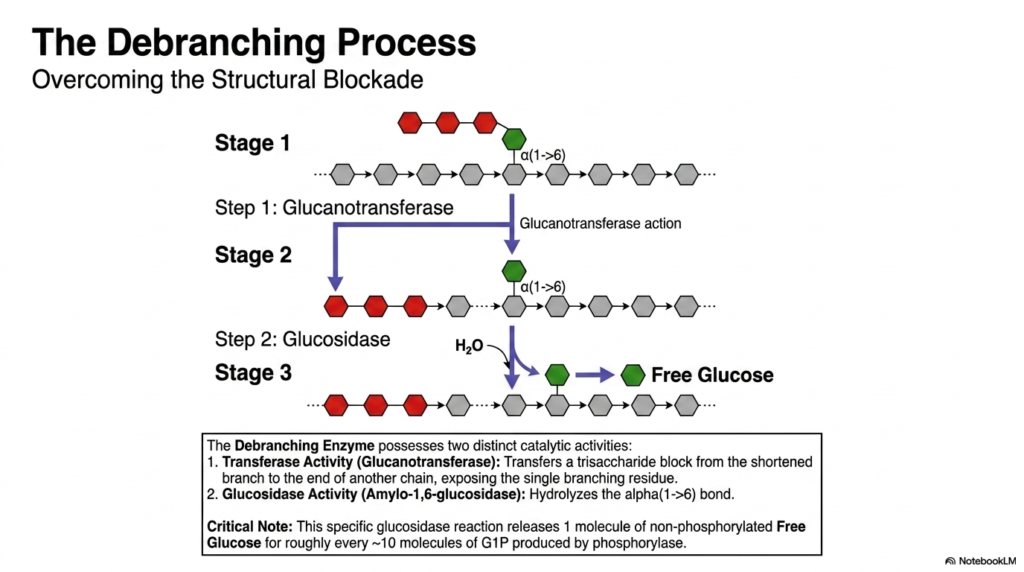
To overcome the structural blockade of the limit dextrin, Glycogen Metabolism employs a specialized Debranching Enzyme. This remarkable bifunctional protein physically resolves the stubborn branch points left behind by phosphorylase. The debranching process occurs in three distinct stages, utilizing two distinct catalytic activities housed within a single enzyme structure. Stage one represents the stalled limit dextrin. In stage two, the enzyme’s glucanotransferase activity forcefully transfers a trisaccharide block from the shortened branch to the end of a longer, neighboring linear chain.
This transferase activity is a brilliant maneuver within Glycogen Metabolism, as it exposes the single remaining glucose residue still attached by the stubborn alpha(1->6) branch point. During stage three, the enzyme shifts to its second catalytic function: the glucosidase activity. Also known as amylo-1,6-glucosidase, this specific active site uses simple hydrolysis (water) to finally sever the alpha(1->6) bond. This specific reaction clears the branch point, allowing the standard phosphorylase enzyme to resume its rapid breakdown of the newly elongated linear chain.
A critical, often-tested nuance of Glycogen Metabolism lies in the specific products of this debranching step. While phosphorylase produces activated Glucose-1-Phosphate, the hydrolytic cleavage performed by the debranching enzyme’s glucosidase activity releases one molecule of pure, non-phosphorylated free glucose. Because branches occur roughly every ten residues, the overall degradative pathway naturally yields about ten molecules of phosphorylated glucose for every single molecule of free glucose released. This precise biochemical ratio highlights the intricate chemical realities of breaking down such a complex polymer.
Slide 9: Fates of Glucose in Glycogen Metabolism
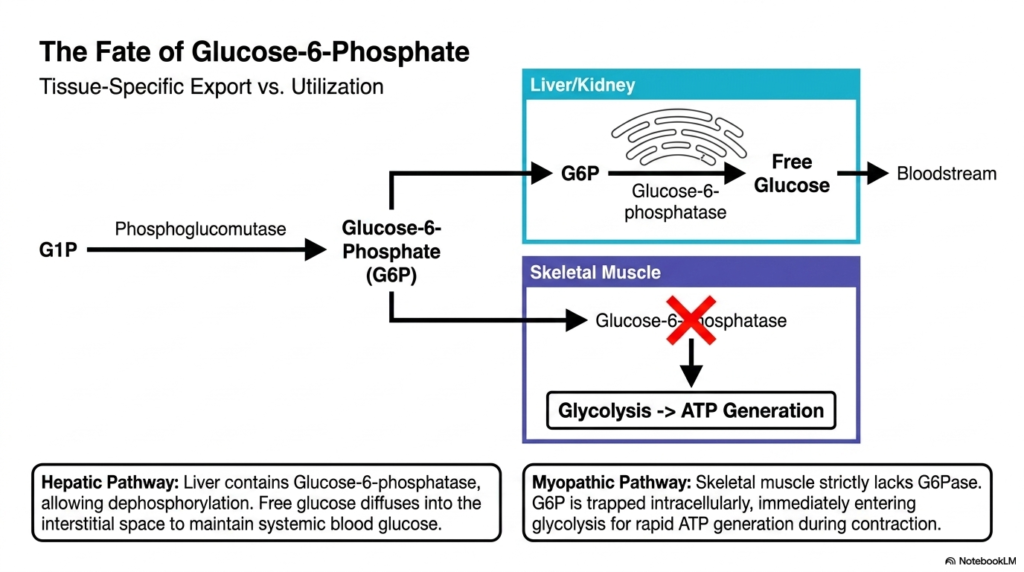
Once degradation yields Glucose-1-Phosphate, the molecule is quickly isomerized back to Glucose-6-Phosphate, a pivotal crossroads in Glycogen Metabolism. This slide highlights how the ultimate fate of this molecule is entirely tissue-dependent. In the liver and kidneys, the overarching goal is supporting systemic biology. These organs possess the specialized enzyme Glucose-6-phosphatase, located strictly within the endoplasmic reticulum. This enzyme diligently strips the phosphate group off the sugar, generating free glucose that can seamlessly diffuse out of the cell and into the bloodstream to nourish other tissues.
This hepatic pathway represents the altruistic side of Glycogen Metabolism. By releasing free glucose into the systemic circulation, the liver effectively buffers against dangerous drops in blood sugar during fasting or intense starvation. The brain, which relies almost exclusively on continuous glucose delivery, heavily depends on the liver’s specific enzymatic capability. Without the presence of Glucose-6-phosphatase, the liver would be utterly incapable of exporting its massive stores of energy, rendering it useless for systemic glucose homeostasis.
In stark contrast, the myopathic pathway highlights the localized nature of skeletal muscle glycogen metabolism. Skeletal muscle strictly lacks the Glucose-6-phosphatase enzyme entirely. Because the charged phosphate group cannot be removed, the Glucose-6-Phosphate remains permanently trapped intracellularly. This trapped molecule is immediately shuttled directly into the glycolysis pathway for rapid ATP generation. This biological design ensures that the muscle always retains its fuel specifically for localized muscular contraction, rather than losing its precious energy reserves to the bloodstream at large.
Slide 10: Lysosomal Degradation and Glycogen Metabolism
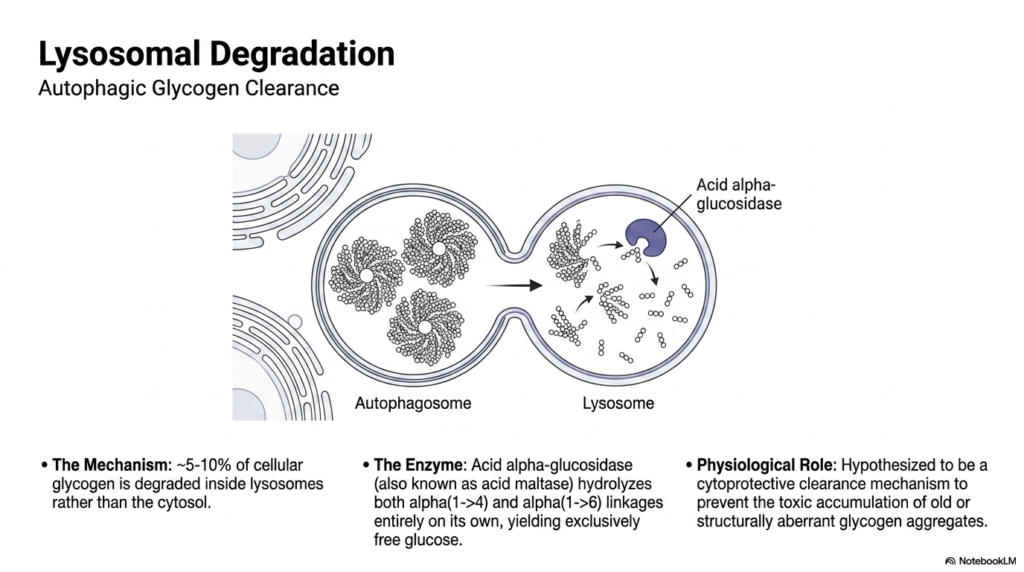
While the primary cytosolic enzymes handle the vast majority of carbohydrate breakdown, a small but vital secondary pathway exists within Glycogen Metabolism. This slide introduces the concept of autophagic clearance and lysosomal degradation. Remarkably, roughly five to ten percent of all cellular glycogen is purposefully sequestered into autophagosomes and subsequently delivered into the highly acidic environment of the lysosomes. Inside the lysosome, a specialized enzyme known as acid alpha-glucosidase, or acid maltase, carries out the degradative process independently of cytosolic signals.
The mechanism of this specialized glycogen metabolism is unique because acid alpha-glucosidase is a biochemical powerhouse. Unlike the cytosolic enzymes, which require a tag-team approach, this single lysosomal enzyme is fully capable of hydrolyzing both the linear alpha(1->4) linkages and the complex alpha(1->6) branch points all on its own. Furthermore, this acidic hydrolysis bypasses phosphorylation entirely, breaking the large polymer down exclusively into molecules of unphosphorylated free glucose, which are then transported out of the lysosome back into the cell’s cytoplasm.
The exact physiological role of this lysosomal branch of Glycogen Metabolism remains a subject of intense scientific study. It is widely hypothesized to be a vital cytoprotective clearance mechanism. By constantly sweeping up and degrading a small percentage of the reserves, the cell actively prevents the toxic accumulation of old, structurally aberrant, or physically damaged glycogen aggregates that could otherwise disrupt normal cellular function. When this specialized autophagic clearance pathway fails, the resulting microscopic cellular damage leads to devastating clinical consequences.
Slide 11: Allosteric Regulation of Glycogen Metabolism
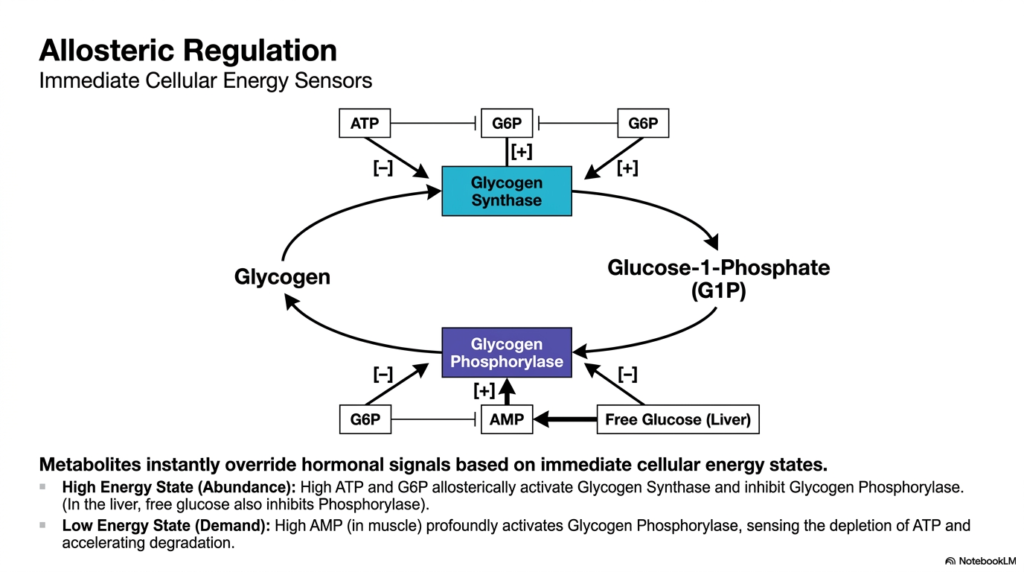
To maintain cellular harmony, Glycogen Metabolism must be exquisitely sensitive to the cell’s immediate energy demands. This slide details the allosteric regulation of the pathway, which utilizes small metabolites as immediate cellular energy sensors. These localized signals are incredibly powerful, capable of instantly overriding broader hormonal commands in response to the local tissue’s immediate energetic reality. This rapid, localized control system ensures that synthesis and degradation enzymes respond instantaneously to fluctuations in internal resources before systemic hormones even have a chance to arrive.
During a high-energy state, indicating cellular abundance, Glycogen Metabolism heavily favors synthesis. Elevated levels of intracellular ATP and Glucose-6-Phosphate act as potent allosteric activators of Glycogen Synthase, forcefully promoting the storage of excess fuel. Simultaneously, these exact same abundant metabolites act as powerful allosteric inhibitors of Glycogen Phosphorylase, aggressively shutting down unnecessary degradation. In the liver, high concentrations of free intracellular glucose also act as a strong inhibitory signal to phosphorylase, indicating that the systemic blood sugar buffers are currently full.
Conversely, during a low-energy state, indicating high cellular demand, the allosteric dynamics of Glycogen Metabolism violently shift. In actively contracting skeletal muscle, rapid ATP consumption generates a massive spike in localized AMP levels. This high concentration of AMP is a potent, immediate allosteric activator of Glycogen Phosphorylase. By directly sensing the dangerous depletion of cellular ATP, the AMP molecule accelerates the massive degradation of carbohydrates, ensuring that the contracting muscle fiber receives an immediate influx of fresh fuel exactly when it needs it most.
Slide 12: Hormonal Regulation of Glycogen Metabolism
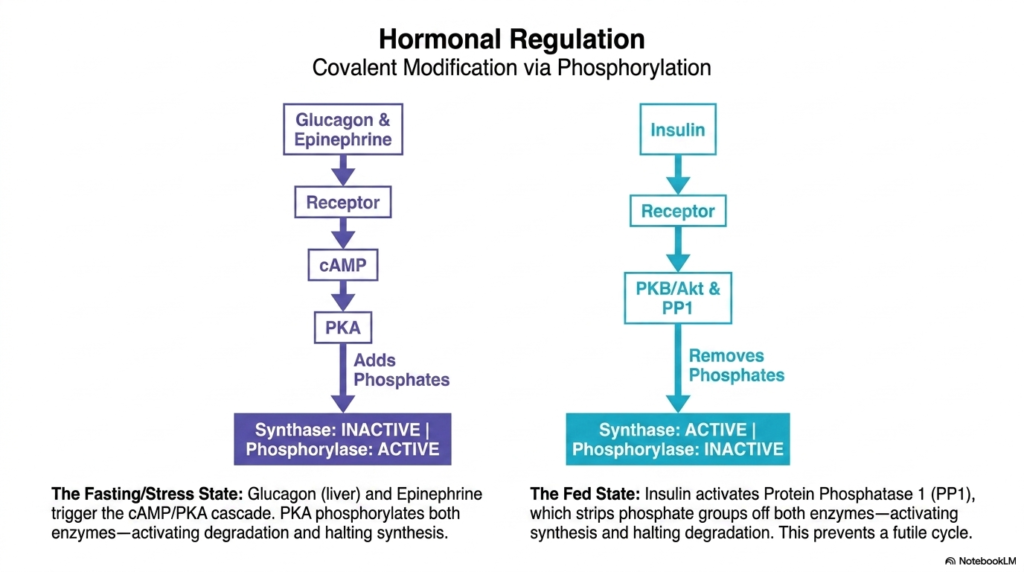
Beyond local metabolic control, Glycogen Metabolism is heavily directed by broad, systemic hormonal regulation via covalent modification. This slide maps out the opposing forces of fasting and feeding. During fasting or stress, the liver responds to glucagon, while the muscle responds to epinephrine. These hormones bind to surface receptors, triggering a powerful signaling cascade that elevates intracellular cAMP levels. This secondary messenger subsequently activates Protein Kinase A (PKA), an enzyme specifically designed to aggressively add phosphate groups to various downstream target proteins.
This covalent phosphorylation event is the master switch of fasting glycogen metabolism. When PKA adds phosphates to the primary enzymes, the results are reciprocally brilliant: Glycogen Synthase becomes physically inactive, while Glycogen Phosphorylase becomes highly active. This elegant, single-step phosphorylation ensures that the cell immediately halts all energy storage and rapidly accelerates energy degradation to meet systemic demands. This mechanism ensures that the liver releases glucose into the bloodstream during a fast and that the muscles mobilize fuel during a stressful fight-or-flight scenario.
In total opposition, the fed state reverses the entire dynamic of Glycogen Metabolism. Following a meal, elevated insulin levels trigger a completely different receptor cascade, ultimately activating Protein Phosphatase 1 (PP1). This phosphatase systematically strips away the phosphate groups previously added by PKA. Dephosphorylation actively restores Glycogen Synthase to its working state and simultaneously shuts down Glycogen Phosphorylase. This effectively prevents a futile metabolic cycle, ensuring that the cell focuses solely on storing incoming nutrients without accidentally degrading them.
Slide 13: Muscular Override in Glycogen Metabolism
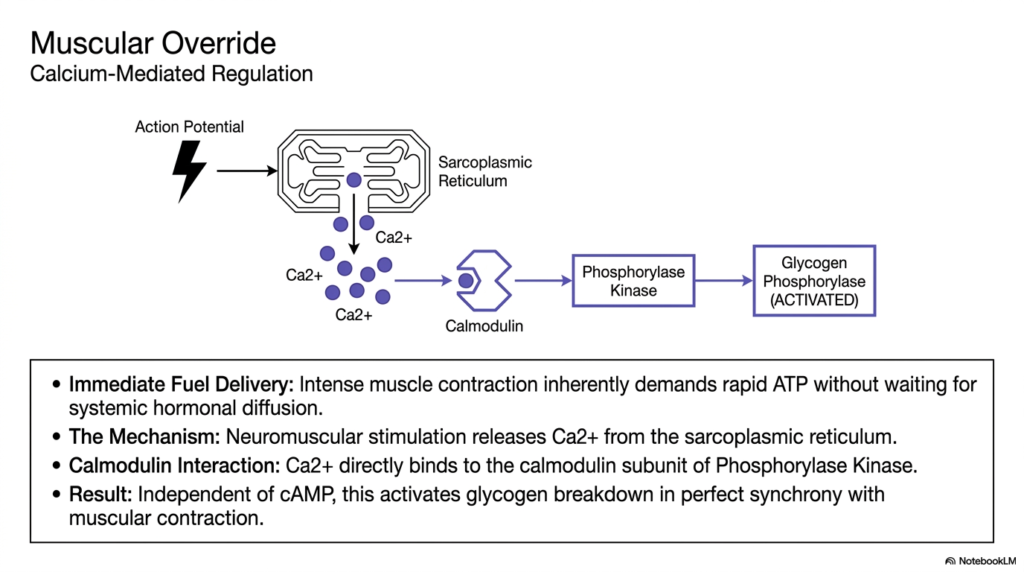
Skeletal muscle requires an incredibly specialized regulatory trick to survive intense exertion, representing a unique override system within Glycogen Metabolism. During a sudden sprint, intense muscle contraction inherently demands a massive, instantaneous influx of ATP. The muscle simply cannot afford to wait for systemic hormones like epinephrine to travel through the bloodstream, bind to receptors, and initiate a lengthy cAMP cascade. Therefore, muscle tissue evolved a brilliant, localized mechanism to bypass standard hormonal pathways and force immediate glucose mobilization.
The mechanism of this override in Glycogen Metabolism is entirely calcium-mediated. When a motor neuron delivers an action potential to the muscle fiber, it triggers a massive, instantaneous release of calcium ions directly from the sarcoplasmic reticulum into the cytoplasm. This flooding of calcium not only drives the physical protein sliding required for the muscle contraction itself but also acts as a potent, immediate biochemical signaling molecule. Calcium directly binds to the specific calmodulin subunit permanently attached to the Phosphorylase Kinase enzyme.
This specific calcium-calmodulin interaction is a masterclass in synchronous Glycogen Metabolism. Upon binding calcium, the kinase enzyme is instantly activated, independent of any cAMP or hormonal signals. Once activated, the kinase rapidly phosphorylates and activates Glycogen Phosphorylase. The spectacular biological result is that the exact same calcium spark that commands the muscle fiber to violently contract simultaneously ignites the biochemical breakdown of stored carbohydrates. This guarantees that intense energy demand and rapid fuel delivery remain in absolute perfect synchrony.
Slide 14: Pathology of Glycogen Metabolism

When the complex enzymes in these pathways fail genetically, the resulting pathologies are known as Glycogen Storage Diseases (GSDs). This slide highlights the severe clinical consequences of disrupted Glycogen Metabolism. Type I, Von Gierke disease, is caused by a catastrophic defect in Glucose-6-Phosphatase. Because the liver cannot export glucose, patients present with severe, life-threatening fasting hypoglycemia, massive hepatomegaly from trapped carbohydrates, and complex downstream issues like lactic acidosis. This highlights the absolute necessity of the hepatic export pathway for human survival.
Type II Pompe disease illustrates a vastly different failure within Glycogen Metabolism. This disease stems from a specific defect in lysosomal acid alpha-glucosidase. Because the cytosolic enzymes function normally, blood sugar is maintained. However, the inability to clear the small percentage of autophagic glycogen leads to massive, toxic lysosomal accumulation. This causes severe cellular damage, particularly presenting as lethal cardiomyopathy and severe muscular hypotonia, tragically resulting in early infant death and highlighting the vital cytoprotective role of the secondary lysosomal pathway.
Types III and V further demonstrate the diverse clinical manifestations of disrupted Glycogen Metabolism. Type III, Cori disease, involves a defective debranching enzyme, leading to milder hypoglycemia and the abnormal accumulation of short, stubby limit dextrins in tissues. Conversely, Type V, McArdle disease, is highly localized, featuring a specific defect in skeletal muscle Glycogen Phosphorylase. Since the liver is unaffected, fasting blood sugar remains completely normal. However, patients suffer from profound exercise intolerance, painful muscle cramps, and myoglobinuria due to their inability to locally mobilize energy.
Slide 15: The Economy of Glycogen Metabolism
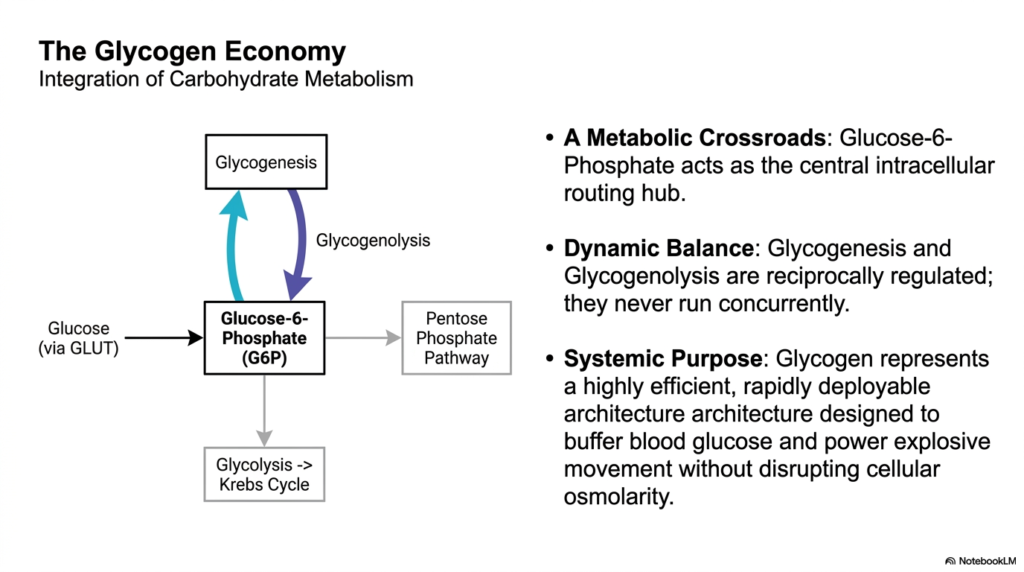
The final slide synthesizes the overarching biological integration of these processes, referring to it as the larger economy of Glycogen Metabolism. At the absolute center of this intracellular routing hub sits Glucose-6-Phosphate. Depending on cellular needs, this single molecule can be polymerized into storage, stripped of its phosphate for systemic export, or directly funneled downwards into glycolysis and the Krebs cycle for immediate ATP generation. It also serves as the critical entry point into the Pentose Phosphate Pathway, proving its role as a master metabolic crossroads.
A core tenet of this economic model is the principle of dynamic balance within Glycogen Metabolism. The pathways of synthesis (glycogenesis) and degradation (glycogenolysis) are exquisitely reciprocally regulated. Through the combined forces of allosteric metabolite sensing and systemic covalent phosphorylation, these opposing pathways are guaranteed to never run concurrently. If both ran simultaneously, the cell would violently burn ATP in a futile cycle of building and breaking the same molecule. Evolution has strictly optimized this system for maximum energetic efficiency.
Ultimately, the systemic purpose of Glycogen Metabolism is undeniable. It provides a highly efficient, rapidly deployable carbohydrate architecture explicitly designed to buffer systemic blood glucose during fasting and to power explosive muscular movement during stress. It achieves all this without disrupting delicate cellular osmolarity. Understanding this complex biochemical economy provides profound insights into fundamental human physiology, daily energy management, and the intricate pathological mechanisms underlying severe metabolic diseases.
Please read our Content Disclaimer Statement.
Check out our social media channels: