39. The Electron Transport Chain: Mechanisms of Mitochondrial Bioenergetics
Imagine a microscopic power plant running ceaselessly to keep a bustling city alive. That is exactly what happens inside cellular mitochondria every second of the day. This slide deck breaks down the complex mechanics of oxidative phosphorylation. By exploring these structures, one uncovers the profound biochemical engines that sustain biological life and drive cellular metabolism.
Slide 1: Introduction to the Electron Transport Chain
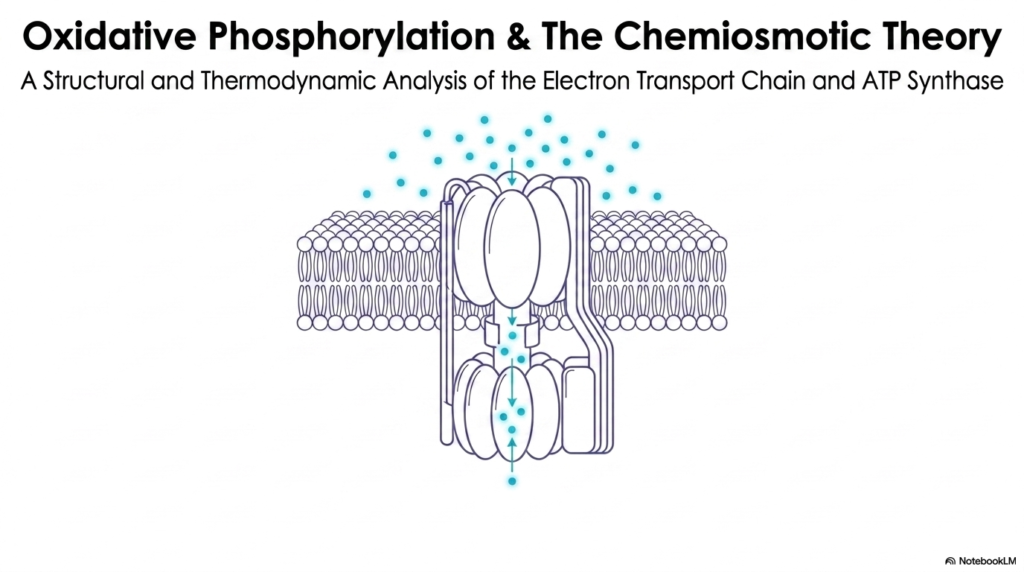
The study of bioenergetics begins with the foundation of oxidative phosphorylation, a process heavily reliant on the Electron Transport Chain. This initial slide introduces the structural and thermodynamic analysis necessary to comprehend how biological systems generate usable energy. At the heart of this process lies a remarkable molecular machine, which operates in tandem with the Electron Transport Chain to drive the continuous synthesis of ATP. This intricate relationship forms the basis of the chemiosmotic theory, fundamentally altering the way biochemists view energy transduction.
To fully appreciate this concept, one must recognize that the Electron Transport Chain is not merely a series of static proteins. Instead, it is a highly dynamic and exquisitely coordinated sequence of oxidation-reduction reactions. As electrons flow through these protein complexes, they release energy that is harnessed to perform mechanical and chemical work. This slide provides a high-level visual representation of ATP synthase embedded within a lipid bilayer, illustrating the ultimate destination for the energy harvested by the chain.
The visual representation of protons flowing through the ATP synthase complex emphasizes the coupling of chemical gradients to mechanical rotation. This structural marvel depends entirely on the gradients established by the Electron Transport Chain. Without this preceding sequence of electron transfers, the turbine-like motion of ATP synthase would grind to a halt. Thus, understanding this first slide is crucial for students, as it sets the stage for the detailed thermodynamic principles that govern cellular respiration.
The thermodynamic analysis mentioned on the slide highlights that energy transformations in biological systems are strictly governed by the laws of physics. The energy extracted from nutrients must be converted efficiently. By coupling exergonic electron transfers within the Electron Transport Chain to the endergonic pumping of protons, cells avoid dissipating all metabolic energy as useless heat, thereby establishing the elegant chemiosmotic paradigm.
Slide 2: The Chemiosmotic Theory and the Electron Transport Chain
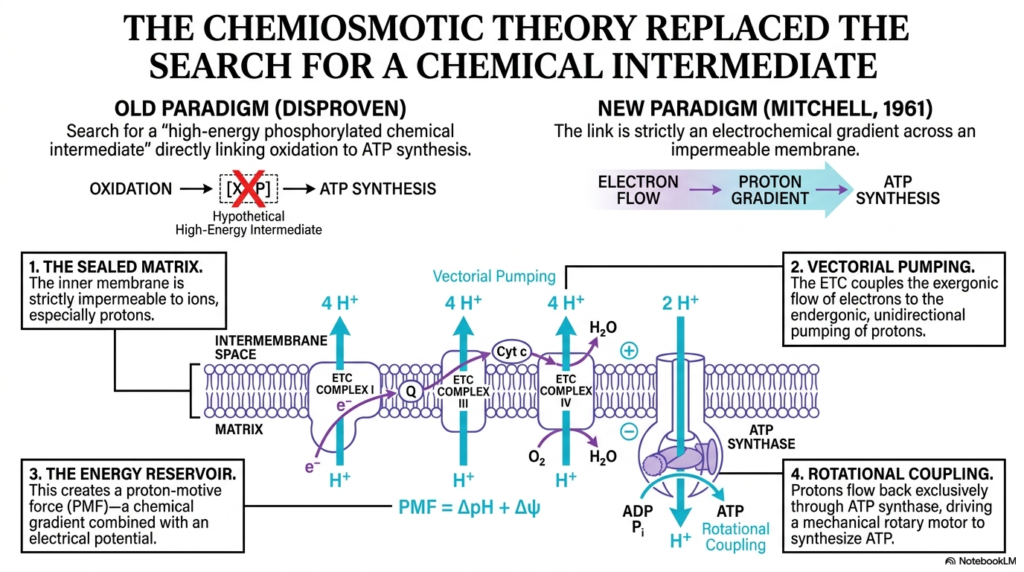
The historical transition from the search for a chemical intermediate to the acceptance of the chemiosmotic theory represents a monumental shift in biochemistry. For decades, scientists fruitlessly hunted for a high-energy phosphorylated molecule directly linking oxidation to ATP synthesis. It was Peter Mitchell in 1961 who proposed that the Electron Transport Chain does not synthesize a chemical intermediate. Instead, it generates an electrochemical gradient across an impermeable membrane.
This slide breaks down the new paradigm into four critical components, starting with the sealed matrix. The inner mitochondrial membrane must remain strictly impermeable to ions, especially protons. Without this sealed barrier, the work performed by the Electron Transport Chain would be instantly undone, as protons would simply diffuse back to equilibrium. This impermeability ensures that the energy reservoir is properly maintained for subsequent cellular work.
The second and third components involve vectorial pumping and the creation of an energy reservoir. The Electron Transport Chain couples the exergonic flow of electrons to the endergonic, unidirectional pumping of protons. This action creates the proton-motive force, a powerful combination of a chemical gradient and an electrical potential. This force represents stored potential energy, completely replacing the disproven old paradigm of a chemical intermediate.
Finally, the fourth concept is rotational coupling. Protons are allowed to flow back down their gradient exclusively through ATP synthase. This flow serves as a mechanical rotary motor for synthesizing ATP. The Electron Transport Chain is thus fundamentally an electrical circuit driving a biochemical pump, providing the proton-motive force required to turn the ATP synthase turbine and sustain biological energy needs.
Slide 3: Spatial Topography of the Electron Transport Chain
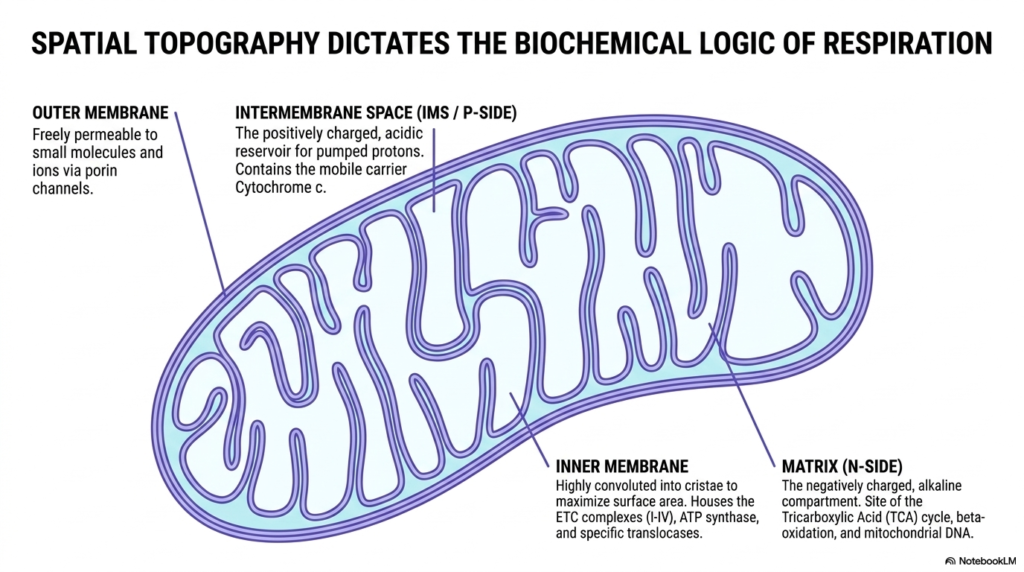
The biochemical logic of cellular respiration is deeply intertwined with mitochondrial architecture. This slide illustrates how spatial topography dictates function, highlighting the distinct compartments required for the Electron Transport Chain to operate. The outer membrane serves as a highly permeable barrier, allowing small molecules and ions to freely pass through porin channels, while the internal structures maintain strict compartmentalization for energy production.
Moving inward, the intermembrane space acts as a positively charged, acidic reservoir for the protons pumped by the Electron Transport Chain. This compartment is essential for housing mobile carriers such as cytochrome c. Without this confined space, the localized concentration of protons required to build the proton-motive force would dissipate into the broader cellular environment, rendering oxidative phosphorylation completely ineffective.
The inner membrane is the true operational hub, highly convoluted into cristae to maximize its surface area. This vast surface houses the massive protein complexes of the Electron Transport Chain, along with ATP synthase and the necessary translocases. The invaginations of the cristae ensure that thousands of these respiratory chains can operate simultaneously within a single mitochondrion, meeting the cell’s immense energetic demands.
Enclosed by this membrane is the matrix, a negatively charged and alkaline compartment. It is the site of the Tricarboxylic Acid cycle, beta-oxidation, and mitochondrial DNA. The matrix provides the crucial electron donors, such as NADH, that feed directly into the Electron Transport Chain. This spatial separation of the alkaline matrix and the acidic intermembrane space is the physical embodiment of the chemiosmotic gradient.
Slide 4: Mobile Carriers in the Electron Transport Chain
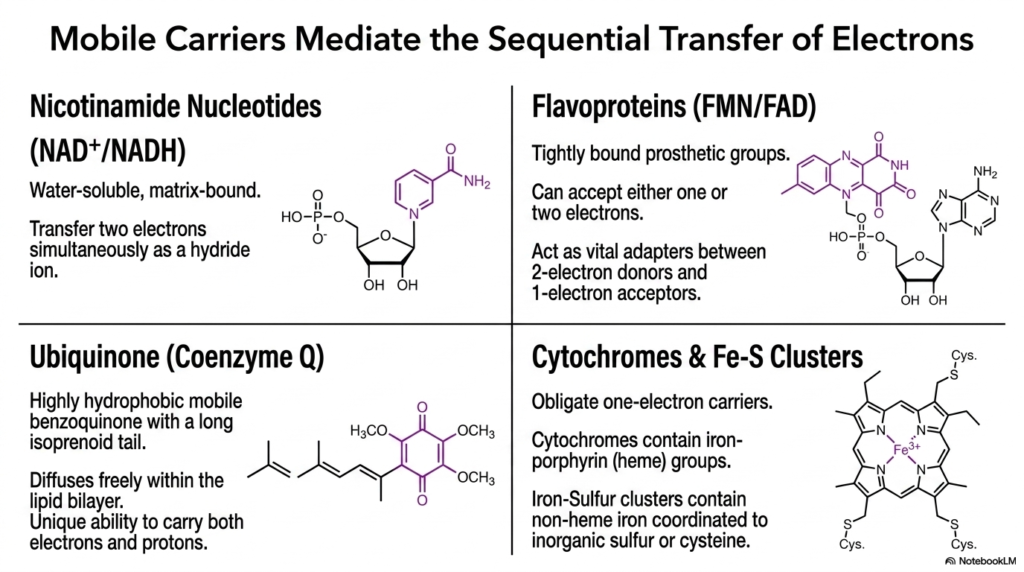
A seamless flow of electrons requires specialized molecules capable of navigating the mitochondrial environment. This slide details the mobile carriers that mediate the sequential transfer of electrons throughout the Electron Transport Chain. These molecules act as biological wires, shuttling energetic electrons between the massive, stationary protein complexes embedded in the inner mitochondrial membrane.
Nicotinamide nucleotides, specifically NAD+ and NADH, serve as water-soluble, matrix-bound carriers. They transfer two electrons simultaneously as a hydride ion. In contrast, flavoproteins containing FMN or FAD possess tightly bound prosthetic groups that can accept either one or two electrons. This flexibility makes them vital adapters within the Electron Transport Chain, perfectly bridging the gap between obligate two-electron donors and one-electron acceptors.
Ubiquinone, or Coenzyme Q, represents another critical component. It is a highly hydrophobic, mobile benzoquinone featuring a long isoprenoid tail. This structure allows it to diffuse freely within the lipid bilayer. Ubiquinone has the unique ability to carry both electrons and protons, serving as an indispensable shuttle that links the initial complexes of the Electron Transport Chain to subsequent stages of respiration.
Finally, cytochromes and iron-sulfur clusters function as obligate one-electron carriers. Cytochromes utilize iron-porphyrin heme groups, while iron-sulfur clusters contain non-heme iron coordinated to inorganic sulfur or cysteine residues. Together, these diverse carriers ensure that electrons move efficiently and safely down the thermodynamic gradient of the Electron Transport Chain, preventing the accidental release of dangerous free radicals.
Slide 5: Complex I of the Electron Transport Chain
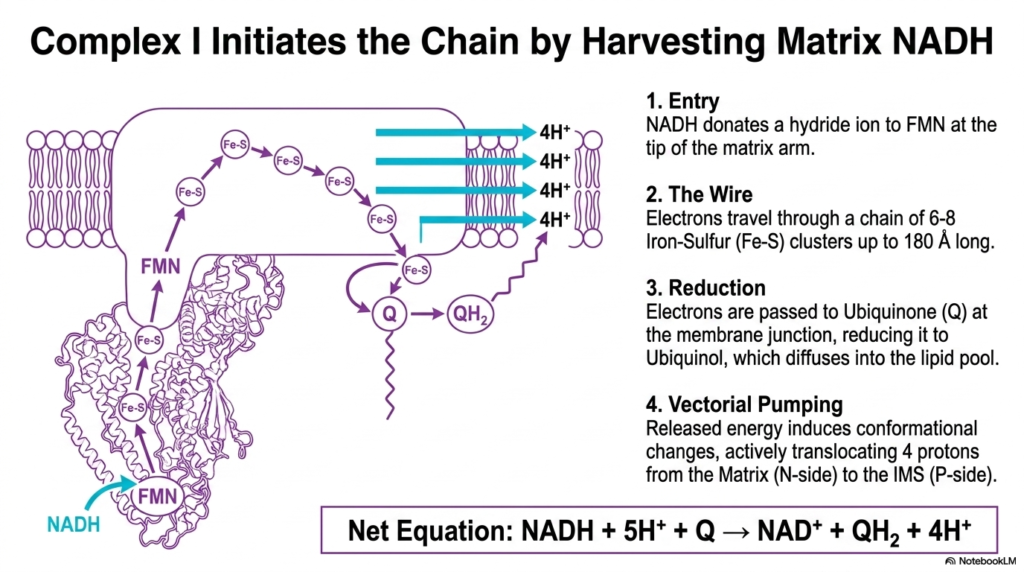
Complex I serves as the primary entry point for most electrons entering the respiratory pathway. This slide explores how Complex I initiates the Electron Transport Chain by harvesting energetic electrons from matrix NADH. The process begins when NADH donates a hydride ion to the FMN prosthetic group at the very tip of the matrix arm, thereby initiating the flow of biological current.
Once inside the complex, electrons travel through a highly organized internal wire consisting of six to eight Iron-Sulfur clusters. This conductive pathway stretches up to 180 angstroms long, rapidly moving electrons away from the initial binding site. This internal electron transfer is a defining feature of the Electron Transport Chain, ensuring that energetic electrons are safely directed to their next destination without causing cellular damage.
At the membrane junction, these electrons are finally passed to Ubiquinone. The addition of electrons reduces Ubiquinone to Ubiquinol, which then detaches and diffuses into the hydrophobic lipid pool. This reduction is a critical step in the Electron Transport Chain, as it shifts electron flow from the hydrophilic matrix arm to the lipid-soluble environment of the inner mitochondrial membrane.
Crucially, the energy released during these electron transfers induces profound conformational changes across the transmembrane domain of Complex I. These mechanical shifts actively translocate four protons from the negative matrix to the positive intermembrane space. By moving these protons, Complex I directly contributes to the electrochemical gradient, proving its vital role as a primary proton pump in the Electron Transport Chain.
Slide 6: Alternative Entry Points to the Electron Transport Chain
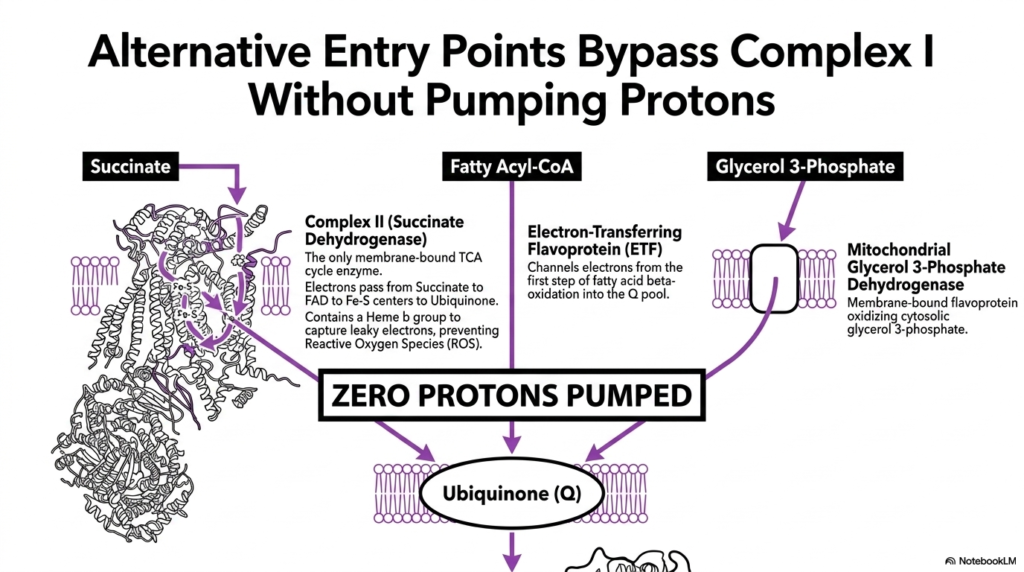
While Complex I is the primary entry point, cellular metabolism requires flexible pathways for different substrates. This slide details alternative entry points that bypass Complex I, thereby feeding electrons directly into the Electron Transport Chain. These pathways rely on distinct enzymes that transfer electrons to the Ubiquinone pool without contributing to the initial proton pumping step.
Complex II, or Succinate Dehydrogenase, is the only membrane-bound enzyme of the TCA cycle. It oxidizes succinate, passing electrons through FAD to Iron-Sulfur centers, and ultimately to Ubiquinone. Notably, it contains a Heme b group designed specifically to capture leaky electrons and prevent the formation of reactive oxygen species, ensuring the safe operation of the Electron Transport Chain during this process.
Fatty acid metabolism also feeds into this system via the Electron-Transferring Flavoprotein. This protein channels electrons generated from the first step of fatty acid beta-oxidation directly into the Ubiquinone pool. This flexibility allows the Electron Transport Chain to harness energy from lipid stores, ensuring that the mitochondria can maintain ATP production even when carbohydrate sources are scarce or depleted.
Lastly, the Mitochondrial Glycerol 3-Phosphate Dehydrogenase provides a pathway for cytosolic reducing equivalents. This membrane-bound flavoprotein oxidizes cytosolic glycerol 3-phosphate, sending electrons to Ubiquinone. A critical concept here is that all these alternative pathways pump zero protons directly. They simply reduce the mobile carrier pool, bypassing the first pumping site of the Electron Transport Chain entirely.
Slide 7: The Q-Cycle within the Electron Transport Chain
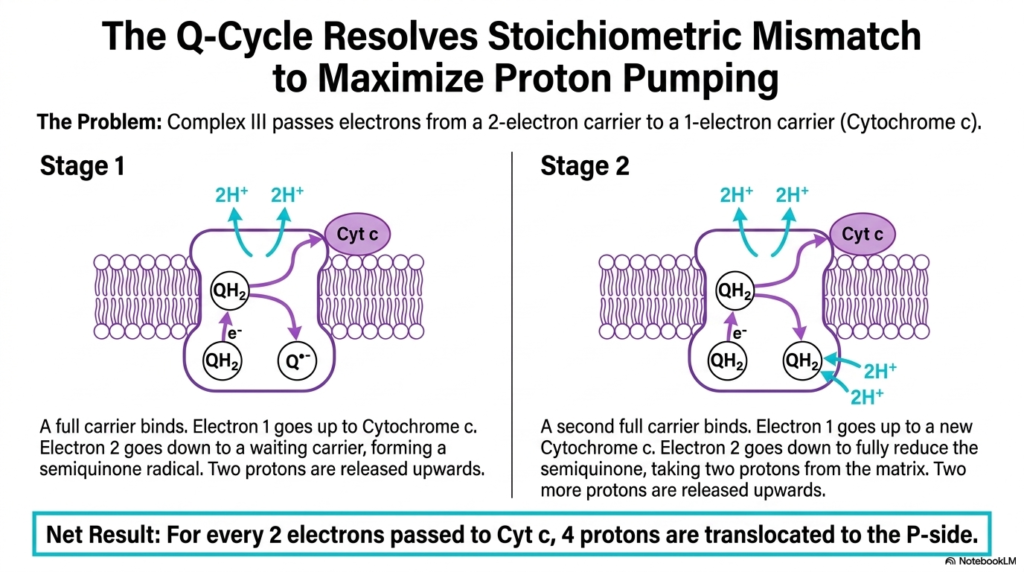
One of the most elegant biochemical solutions in nature occurs within Complex III. This slide examines the Q-Cycle, a complex mechanism that resolves a significant stoichiometric mismatch within the Electron Transport Chain. Complex III faces a distinct problem: it must pass electrons from a two-electron carrier, Ubiquinol, to a strict one-electron carrier, Cytochrome c.
The Q-Cycle operates in two distinct stages to maximize proton pumping efficiency. In the first stage, a fully reduced carrier binds to the complex. One electron travels upward to reduce Cytochrome c, while the second electron travels downward to a waiting oxidized carrier, forming a semiquinone radical. This bifurcated electron flow is a hallmark of this segment of the Electron Transport Chain.
During this first stage, two protons are also released upward into the intermembrane space. In the second stage, a second fully reduced carrier binds. Again, one electron moves up to a new Cytochrome c, while the second electron moves down to fully reduce the waiting semiquinone radical. This step takes two protons from the matrix, demonstrating the intricate proton handling of the Electron Transport Chain.
The net result of the Q-Cycle is highly efficient. For every two electrons passed to Cytochrome c, four protons are translocated to the positive side of the membrane. This mechanism allows the Electron Transport Chain to double its proton-pumping capacity at Complex III, beautifully resolving the stoichiometric mismatch while maximizing the energy stored in the chemiosmotic gradient.
Slide 8: Complex IV of the Electron Transport Chain
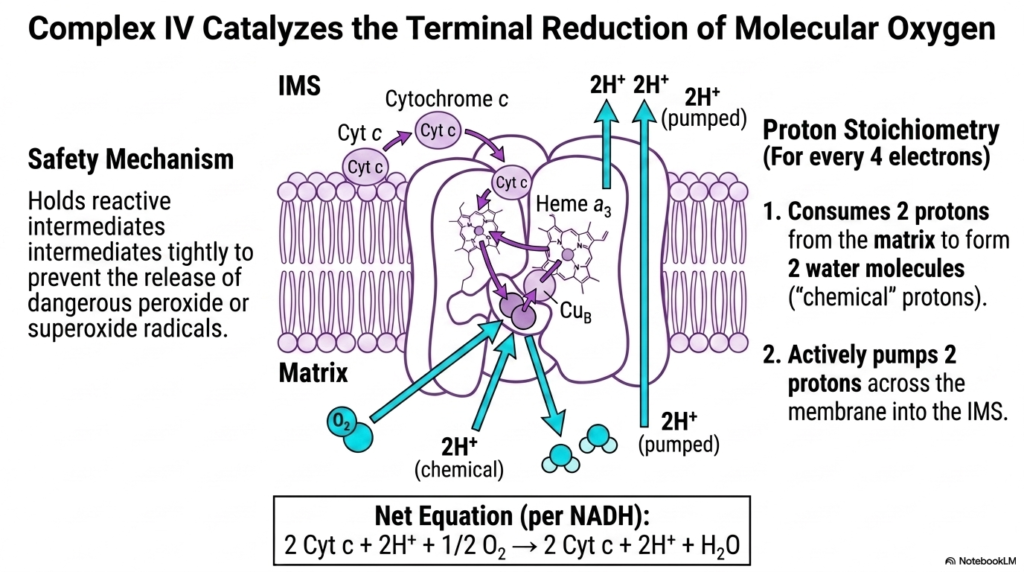
The final step of electron transfer involves the careful handling of molecular oxygen. This slide breaks down how Complex IV catalyzes the terminal reduction of molecular oxygen, serving as the ultimate electron sink for the Electron Transport Chain. Four electrons, delivered sequentially by Cytochrome c, are utilized to reduce a single molecule of oxygen into two harmless molecules of water.
This process requires a sophisticated safety mechanism. Complex IV tightly holds reactive oxygen intermediates between its specialized metallic centers, specifically Heme a3 and Copper B. This grip prevents the disastrous release of peroxide or superoxide radicals into the cell. This safety feature is arguably one of the most critical evolutionary adaptations of the Electron Transport Chain.
The proton stoichiometry of Complex IV is fascinating. For every four electrons processed, the complex consumes four “chemical” protons directly from the matrix to form the two water molecules. This consumption alone contributes to the depletion of matrix protons, thereby enhancing the overall electrochemical gradient generated by the Electron Transport Chain.
Furthermore, the energy released from these final electron transfers is harnessed to actively pump an additional two protons across the membrane into the intermembrane space. Therefore, the net action of Complex IV per NADH oxidized is the transfer of two electrons, the consumption of two chemical protons, and the active pumping of two protons, bringing the primary reactions of the Electron Transport Chain to a close.
Slide 9: Thermodynamics of the Electron Transport Chain

The biological purpose of pumping protons is to store thermodynamic potential. This slide defines the Proton-Motive Force and illustrates how it combines chemical and electrical gradients generated by the Electron Transport Chain. The energy stored is represented by an equation that incorporates the pH difference and the transmembrane electrical potential.
In actively respiring mitochondria, the inner membrane maintains an electrical potential of approximately 0.15-0.20 volts, with the matrix being highly negative. Simultaneously, there is a pH difference of about 0.75 units, making the matrix highly alkaline. This combined electrochemical gradient represents the massive energy reservoir meticulously built by the continuous operation of the Electron Transport Chain.
An energetic accounting reveals the incredible efficiency of this biological system. Pumping a single proton outward costs approximately 20 kilojoules per mole. In contrast, the complete oxidation of one NADH molecule releases roughly 220 kilojoules per mole of energy. This energy release demonstrates that the oxidation reactions of the Electron Transport Chain have sufficient power to drive the pumping mechanisms.
The conclusion drawn from this thermodynamic math is that the energy from NADH safely covers the cost of pumping ten protons, which requires about 200 kilojoules per mole. The minimal energy lost as heat ensures that the Electron Transport Chain operates near thermodynamic equilibrium, maximizing the energy available to build the ATP-synthesis reservoir.
Slide 10: ATP Synthase and the Electron Transport Chain
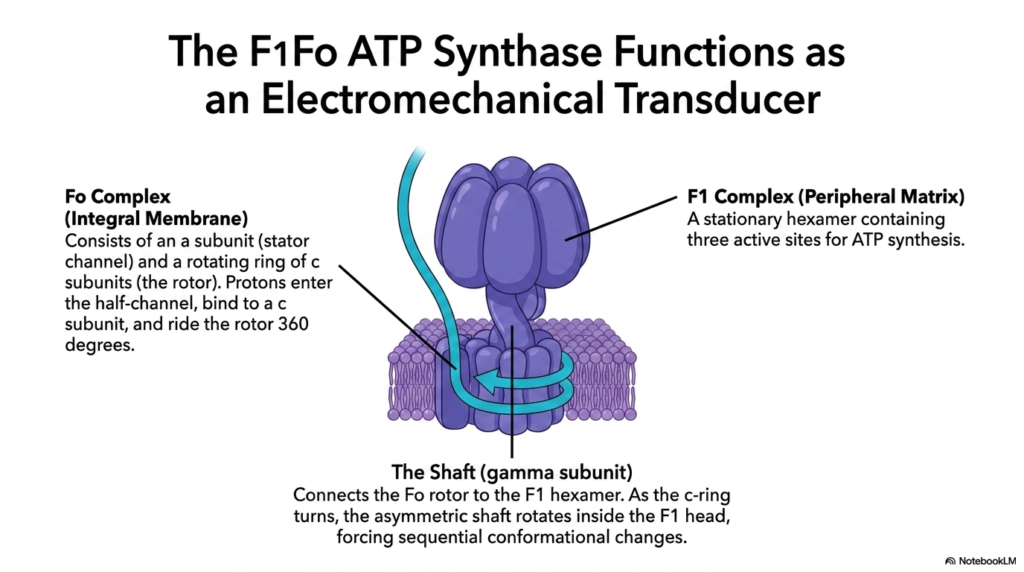
With the proton gradient fully established, the cell must now harvest this potential energy. This slide introduces the F1Fo ATP Synthase, detailing its function as an electromechanical transducer that relies entirely on the Electron Transport Chain. This massive protein complex spans the inner mitochondrial membrane and extends into the matrix, acting as a molecular turbine.
The Fo complex, an integral membrane component, consists of a stator channel and a rotating ring of c subunits known as the rotor. Protons sourced from the gradient built by the Electron Transport Chain enter a half-channel in the stator, bind to a specific c subunit, and ride the rotor for a complete 360-degree turn before exiting into the matrix.
The F1 complex is a stationary peripheral matrix structure containing a hexamer with three active sites dedicated to ATP synthesis. Connecting the Fo rotor to the F1 head is the gamma subunit, which functions as an asymmetrical shaft. The physical link between these components highlights how the electrochemical work of the Electron Transport Chain is converted directly into mechanical torque.
As protons drive the c-ring to turn, the asymmetric gamma shaft rotates violently inside the stationary F1 head. This rotation forces sequential, sweeping conformational changes in the active sites. Therefore, the direct product of the Electron Transport Chain—the proton gradient—is transformed into the mechanical kinetic energy required to manufacture cellular energy.
Slide 11: Mechanical Rotation Driven by the Electron Transport Chain
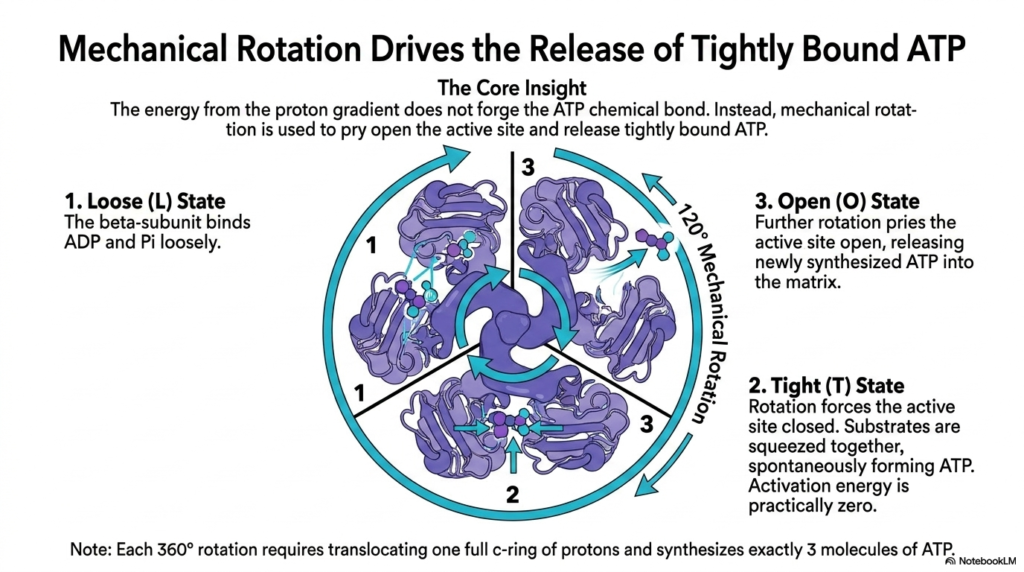
The precise mechanism of ATP creation defies classical chemical intuition. This slide explains how mechanical rotation drives the release of tightly bound ATP, utilizing the gradient built by the Electron Transport Chain. The core biochemical insight here is that the energy from the proton gradient is not actually used to forge the chemical bond between ADP and inorganic phosphate.
Instead, the active sites of the F1 hexamer spontaneously form ATP with practically zero activation energy. However, this newly formed ATP binds so tightly to the enzyme that it cannot leave. The mechanical rotation, powered by the protons pumped from the Electron Transport Chain, provides the sheer physical force required to pry the active site open and release the product.
This process follows three distinct conformational states: Loose, Tight, and Open. In the Loose state, the beta-subunit binds ADP and phosphate loosely. Rotation then forces the site into the Tight state, squeezing the substrates together and allowing ATP to form spontaneously. This squeezing action is indirectly powered by the continuous effort of the Electron Transport Chain.
Finally, further rotation forces the unit into the Open state, releasing the synthesized ATP into the mitochondrial matrix. A full 360-degree rotation, requiring the translocation of one complete c-ring of protons, synthesizes exactly three molecules of ATP. This mechanical marvel represents the ultimate culmination of the energy harvested by the Electron Transport Chain.
Slide 12: Yield Math of the Electron Transport Chain
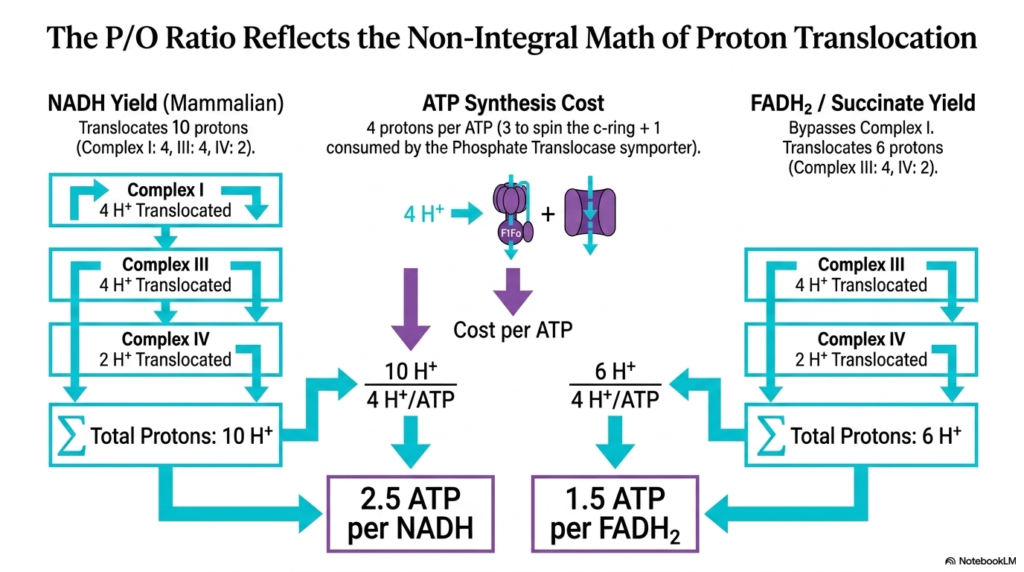
Calculating the exact energy yield of cellular respiration reveals fascinating non-integral values. This slide explains the P/O ratio, reflecting the complex math of proton translocation driven by the Electron Transport Chain. The P/O ratio represents the number of ATP molecules synthesized per oxygen atom reduced, highlighting the stoichiometric efficiency of mitochondrial bioenergetics.
Mammalian NADH oxidation translocates exactly ten protons across the membrane. Complex I pumps four, Complex III pumps four, and Complex IV pumps two. Meanwhile, synthesizing one ATP costs four protons—three to physically spin the c-ring and one consumed by the phosphate translocase symporter. This balance sheet relies completely on the proton inventory generated by the Electron Transport Chain.
Dividing the ten protons pumped by the four protons required yields 2.5 ATP per NADH molecule. In contrast, FADH2 bypasses the first pumping site of the Electron Transport Chain. Its oxidation via Complex II translocates only six protons in total, whereas oxidation via Complex III and Complex IV translocates significantly more, thereby reducing its energetic potential compared to NADH.
Dividing the six protons pumped by FADH2 by the four protons required yields 1.5 ATP per FADH2 molecule. These fractional P/O ratios definitively prove that oxidative phosphorylation is not a simple stoichiometric chemical reaction. Instead, it is a complex, indirectly coupled system driven entirely by the electrochemical gradients of the Electron Transport Chain.
Slide 13: Cytosolic Shuttles to the Electron Transport Chain
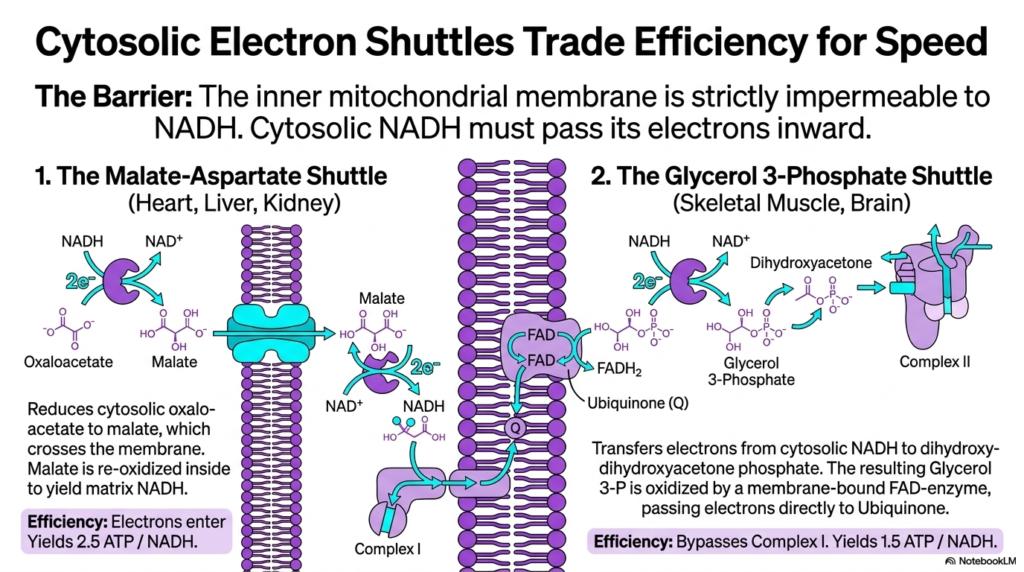
Mitochondria face a logistical challenge regarding cytosolic energy sources. This slide outlines how cytosolic electron shuttles trade efficiency for speed to supply the Electron Transport Chain. The primary biological barrier is that the inner mitochondrial membrane is strictly impermeable to NADH. Therefore, glycolysis-derived cytosolic NADH must pass its electrons inward without crossing the membrane itself.
The Malate-Aspartate shuttle, prominent in the heart, liver, and kidneys, solves this by reducing oxaloacetate to malate. Malate freely crosses the membrane and is re-oxidized inside the matrix to yield fresh NADH. This matrix NADH can then enter the Electron Transport Chain at Complex I. This highly efficient shuttle maintains the maximum theoretical yield of 2.5 ATP per cytosolic NADH.
Conversely, skeletal muscle and brain tissues utilize the Glycerol 3-Phosphate shuttle. This pathway transfers electrons from cytosolic NADH to dihydroxyacetone phosphate. The resulting molecule is oxidized by a membrane-bound FAD-enzyme, which directly passes electrons to the Ubiquinone pool of the Electron Transport Chain. This process completely bypasses the proton-pumping action of Complex I.
While the Glycerol 3-Phosphate shuttle is incredibly fast, it sacrifices efficiency, yielding only 1.5 ATP per NADH. This trade-off ensures rapid energy production during sudden metabolic demands. Both shuttles demonstrate the versatile mechanisms cells use to ensure continuous electron delivery to the Electron Transport Chain across an impermeable membrane.
Slide 14: Disruptions of the Electron Transport Chain

Analyzing metabolic poisons provides deep insights into mitochondrial function. This slide differentiates between the pathological inhibition and physiological uncoupling of the Electron Transport Chain. Inhibitors and uncouplers target distinct components of oxidative phosphorylation, leading to vastly different biochemical consequences for the cell and the organism.
True inhibitors, such as cyanide, carbon monoxide, and rotenone, physically block electron flow through specific complexes. This instantly prevents proton pumping, causing the proton gradient to collapse and halting ATP synthesis completely. Conversely, ATP synthase inhibitors such as oligomycin block the Fo proton channel. In this scenario, the Electron Transport Chain halts due to immense backpressure from a maximally steep proton gradient.
Chemical uncouplers, like 2,4-Dinitrophenol (DNP), operate differently. These hydrophobic weak acids ferry protons back across the membrane, destroying the gradient. Stripped of back-pressure, the Electron Transport Chain runs in extreme overdrive, hopelessly trying to rebuild the gradient. The metabolic energy is entirely dissipated as lethal levels of heat.
However, uncoupling is not always pathological. Physiological uncoupling utilizes regulated protein pores, such as Thermogenin in brown adipose tissue, to deliberately uncouple the gradient. This controlled short-circuiting of the Electron Transport Chain safely generates non-shivering thermogenesis, a vital adaptation for survival in cold conditions in mammals and hibernating animals.
Slide 15: Universality of the Electron Transport Chain
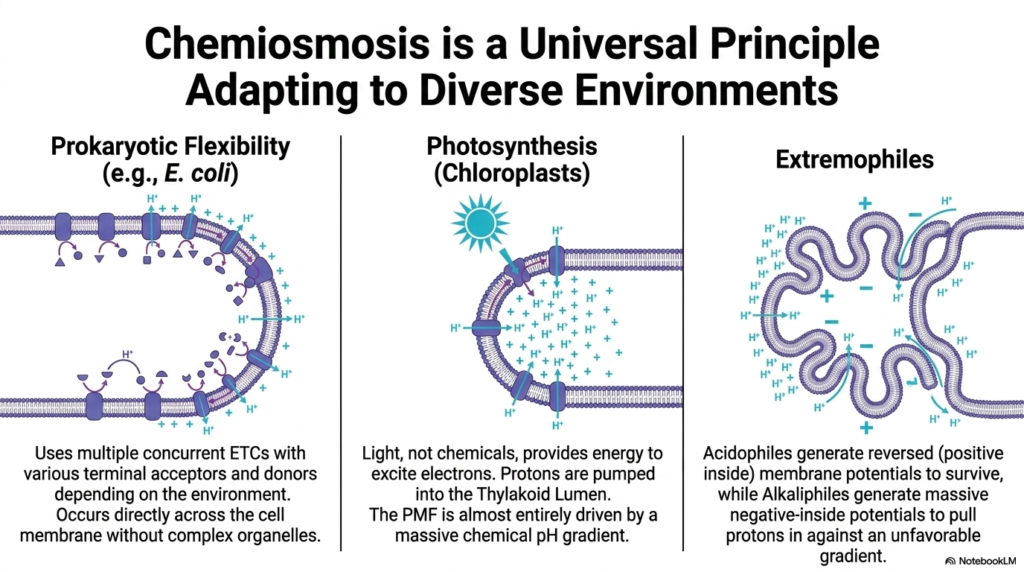
The principles of bioenergetics extend far beyond mammalian mitochondria. This final slide illustrates how chemiosmosis is a universal principle that adapts to diverse environments, driven by variations in the Electron Transport Chain. Across all domains of life, biological entities utilize electrochemical gradients to perform cellular work and synthesize ATP.
Prokaryotes, like E. coli, display immense flexibility. Lacking complex organelles, they have their Electron Transport Chain occur directly across the cellular membrane. They deploy multiple concurrent respiratory chains, using various terminal electron acceptors and donors depending on the availability of nutrients in the environment. This adaptability allows them to thrive in rapidly changing ecological niches.
In photosynthesis, chloroplasts use light, rather than chemicals, to excite electrons. Protons are pumped into the thylakoid lumen by a specialized photosynthetic Electron Transport Chain. Interestingly, this proton-motive force is almost entirely driven by a massive chemical pH gradient, demonstrating a variation on the standard chemiosmotic theme.
Finally, extremophiles showcase the extreme resilience of these systems. Acidophiles generate reversed membrane potentials to survive, while alkaliphiles generate massive negative-inside potentials to pull protons in against highly unfavorable gradients. These incredible adaptations highlight that the fundamental mechanism of the Electron Transport Chain is a central pillar of life on Earth.
Please read our Content Disclaimer Statement.
Check out our social media channels: