34. Adenosine Triphosphate (ATP): The Cellular Energy Currency
Imagine a bustling modern metropolis attempting to run without electricity—everything would instantly grind to a disastrous halt. In the microscopic world of human biology, living cells face the exact same necessity for a reliable, uninterrupted power grid. This comprehensive slide deck explores the extraordinary bioenergetic mechanisms that enable life to continuously power itself. By breaking down complex biochemical structures and intricate metabolic pathways, this presentation illuminates the vital thermodynamic processes that sustain living organisms and offers an accessible guide to molecular energy transfer.
Slide 1: Introduction to Adenosine Triphosphate as the Universal Currency
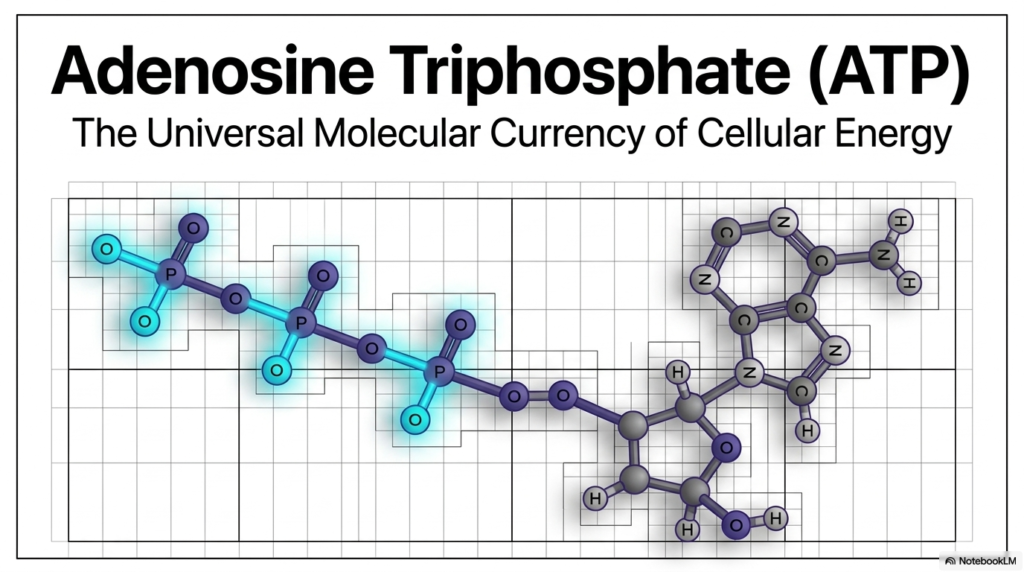
Every biological process, from muscle contraction to complex neural thought, requires energy, and nature has selected a single specialized molecule to act as the primary energetic medium. The first slide introduces Adenosine Triphosphate, beautifully illustrating its complex yet highly efficient molecular structure. Just as human economies rely on a standardized currency to facilitate widespread trade, microscopic cellular ecosystems depend entirely on this extraordinary molecule to support their diverse biological functions. Without a reliable universal energy carrier, the cell’s chaotic thermodynamic environment could never support the highly organized phenomena recognized as complex life.
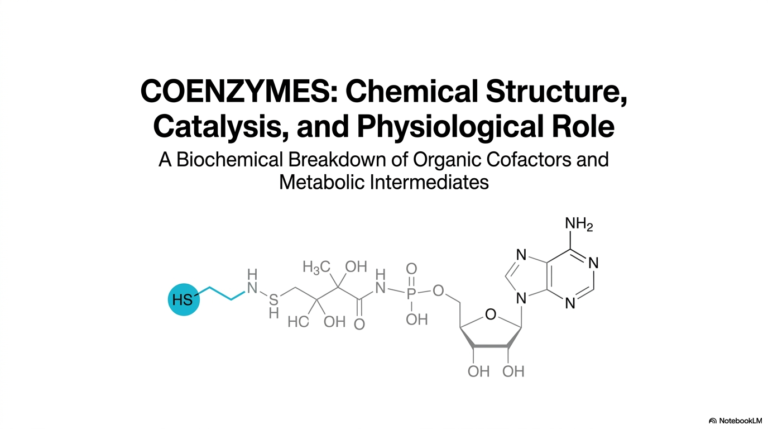
When thoroughly examining the structural representation of Adenosine Triphosphate on this opening slide, one immediately notices the distinct functional regions comprising the molecule. It features a sophisticated nitrogenous base, a central five-carbon ribose sugar, and a highly reactive tail consisting of three phosphate groups. This specific biochemical arrangement represents a finely tuned evolutionary adaptation optimized specifically for rapid biological work. The molecule easily navigates the crowded aqueous environment of the cytoplasm while seamlessly maintaining the intense energetic potential required to drive thermodynamically unfavorable chemical reactions forward across various intracellular compartments.
Grasping the fundamental architecture of Adenosine Triphosphate is an essential first step for any student delving into advanced cellular biochemistry. The carefully balanced covalent bonds within this molecule serve as a reliable microscopic battery, holding tremendous potential energy in a relatively stable form until a cellular mechanism actively demands it. As this presentation progresses, this structural foundation will demystify exactly how breaking and forming these atomic connections translate into the vital work required to keep organisms alive. The universal nature of this energy carrier perfectly highlights the shared evolutionary heritage of cellular biology.
Slide 2: The Transient Storage and Massive Turnover of Adenosine Triphosphate
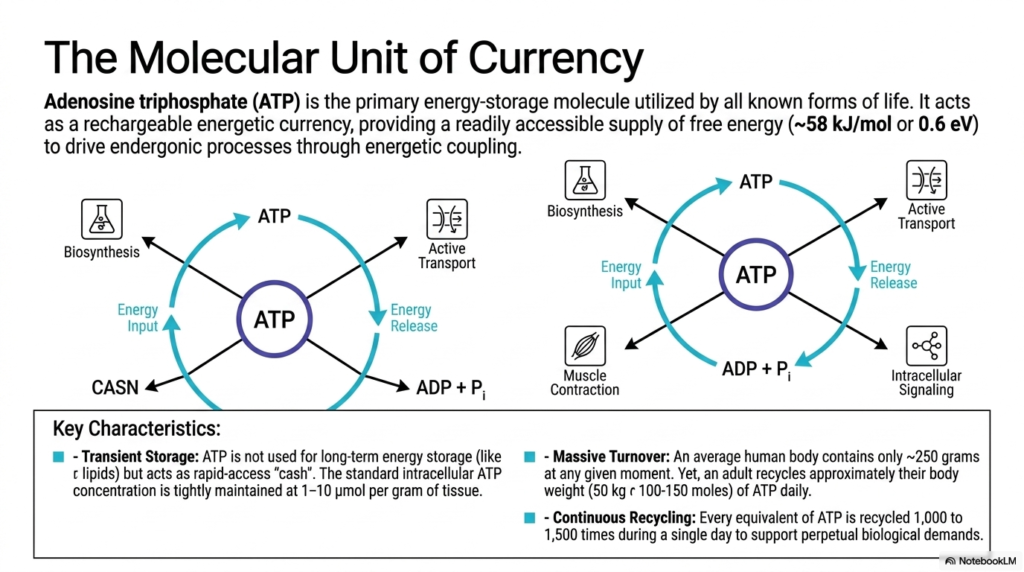
The concept of a rapidly circulating, energetic currency is perfectly encapsulated in the highly dynamic cellular lifecycle of Adenosine Triphosphate. This slide emphasizes that the molecule serves as a readily available source of free energy, capable of delivering approximately 58 kilojoules per mole to drive essential endergonic processes. In complex biological systems, energy must be coupled directly to thermodynamically unfavorable reactions to drive them forward. This molecule powerfully accomplishes exactly that by strategically linking energy-releasing catabolic pathways to essential energy-consuming physiological tasks such as cellular biosynthesis and active membrane transport.
A critical metabolic distinction highlighted in this detailed slide is that Adenosine Triphosphate is never actually utilized for long-term physiological energy storage. Unlike dense lipids or branched glycogen, which serve as long-term metabolic vaults, this highly energetic molecule acts strictly as rapid-access metabolic “cash” for the cell. The standard intracellular concentration is tightly and precisely maintained at roughly one to ten micromoles per gram of living tissue. Because the cell simply cannot stockpile this energetic currency, it must constantly and flawlessly balance ongoing metabolic supply with immediate physiological demand to survive.
Perhaps the most astonishing biochemical fact presented here involves the massive and perpetual turnover rate of Adenosine Triphosphate. An average adult human body remarkably contains only about 250 grams of this molecule at any given isolated moment. Yet, to adequately support the vast biological demands of life, an adult recycles approximately their entire body weight of this molecule every single day. This astonishing volume of continuous chemical recycling vividly illustrates the relentless, unending metabolic engine required to maintain strict biological homeostasis and underscores the truly frantic pace of microscopic life.
Slide 3: Biochemical Structure and the Reactive Bonds of Adenosine Triphosphate
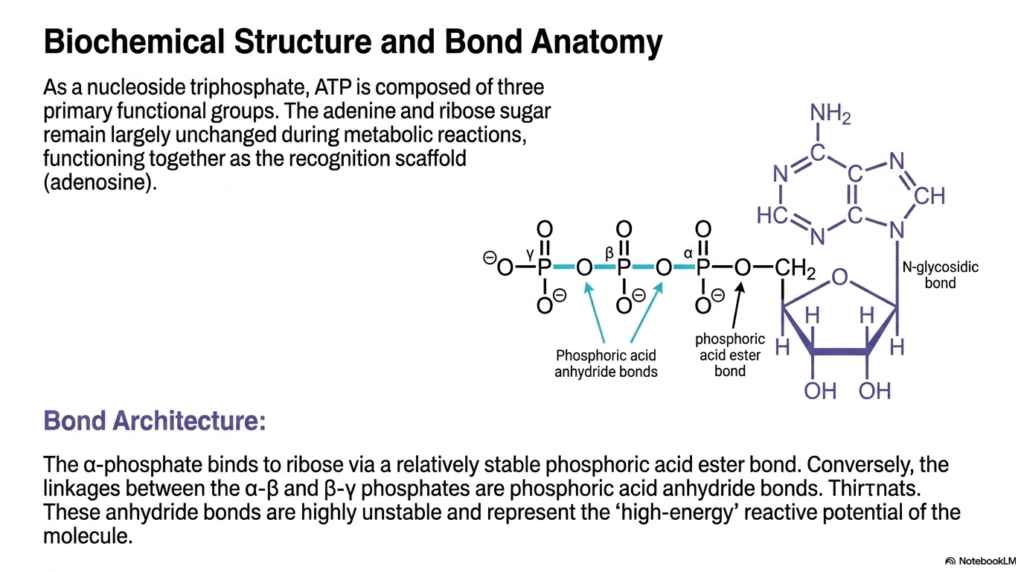
Diving deeper into its molecular anatomy, this slide breaks down the specific chemical components that allow Adenosine Triphosphate to function so effectively as a biological battery. As a nucleoside triphosphate, the molecule features an adenine ring and a ribose sugar that function together securely as the stable recognition scaffold, formally known as adenosine. This stable foundational structure is incredibly important because it allows countless diverse cellular enzymes to physically recognize and safely bind the molecule without accidentally triggering an unwanted, premature release of its highly reactive stored energetic potential.
The critical distinction in bond architecture is fundamental to understanding how Adenosine Triphosphate actually stores energy. The innermost alpha-phosphate securely binds to the central ribose sugar via a relatively stable phosphoric acid ester bond. However, the linkages that structurally connect the alpha-beta and beta-gamma phosphate groups are notably different; they are classified as phosphoric acid anhydride bonds. Due to stringent physical chemistry constraints, these specific anhydride bonds are highly unstable under cellular conditions, reflecting the true ‘high-energy’ reactive potential that defines the molecule’s overall energetic capability.
When an enzyme requires a burst of thermodynamic power, it specifically targets the terminal anhydride bonds in Adenosine Triphosphate for cleavage. The visual diagram clearly maps out where these distinct bonds are located, providing a crucial spatial map for biochemistry students. By isolating the stable ester bond from the highly volatile anhydride bonds, the biological design ensures that only the terminal phosphate groups are rapidly exchanged during standard metabolic reactions. This elegant structural segregation preserves the vital nucleoside scaffold, allowing it to be continuously recycled and re-phosphorylated thousands of times without degrading.
Slide 4: Magnesium and the Active Coenzyme State of Adenosine Triphosphate
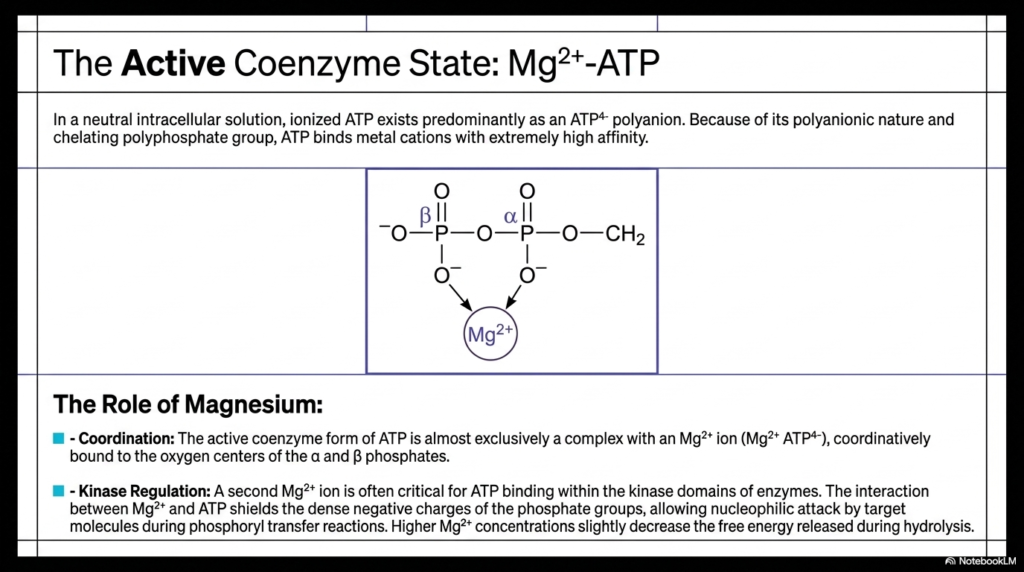
A frequently overlooked but absolutely critical aspect of cellular bioenergetics is the ionic environment required for Adenosine Triphosphate to function properly. In a neutral intracellular solution, this highly charged molecule exists predominantly as a polyanion carrying four negative charges. Because of its intense polyanionic nature and chelating polyphosphate tail, it binds nearby metal cations with extremely high affinity. Consequently, the biologically active coenzyme form of this molecule is almost exclusively a complex tightly bound to a magnesium ion, with the oxygen centers of the innermost phosphate groups coordinated directly to it.
Understanding the direct role of magnesium is essential for grasping how Adenosine Triphosphate interacts with cellular machinery. A second magnesium ion is often absolutely critical for the molecule to bind correctly within the specific kinase domains of various metabolic enzymes. The physical interaction between the magnesium ion and the phosphate groups successfully shields the incredibly dense negative charges radiating from the oxygen atoms. Without this critical ionic shielding effect, the massive electrostatic repulsion would completely prevent approaching target molecules from successfully executing the nucleophilic attacks required for vital phosphoryl transfer reactions.
While the binding of magnesium slightly decreases the total free energy technically released during hydrolysis, it is an unavoidable necessity for practical biochemical utility. By strategically utilizing magnesium to stabilize the explosive negative charges of Adenosine Triphosphate, the cell effectively tames a highly volatile molecular battery, transforming it into a controllable coenzyme. This regulatory mechanism elegantly ensures that energy is only released precisely when an activated kinase enzyme securely locks the shielded molecule into its targeted active site, preventing wasteful spontaneous hydrolysis in the surrounding aqueous cytoplasm.
Slide 5: Thermodynamics and the Hydrolysis of Adenosine Triphosphate
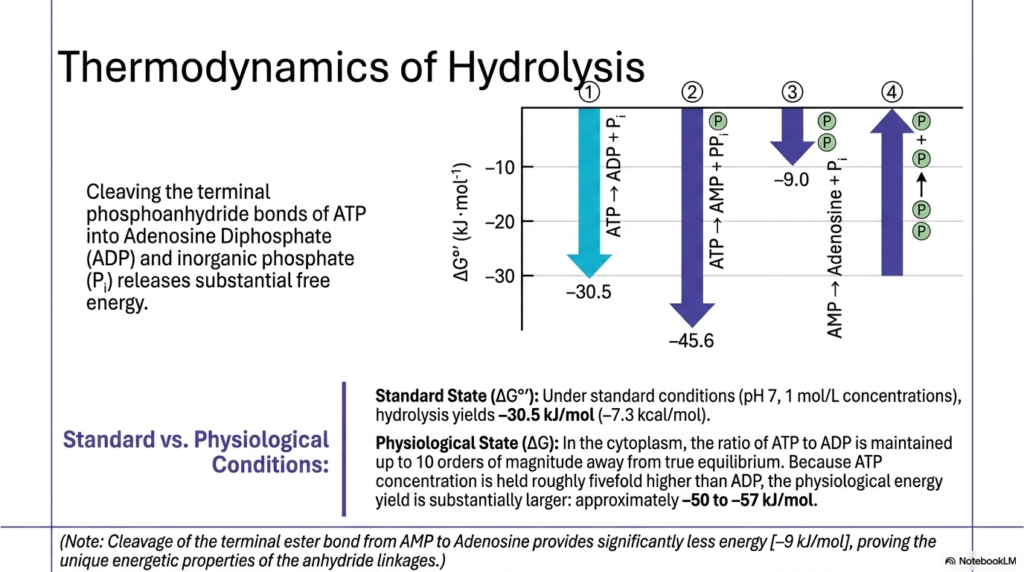
To truly appreciate cellular bioenergetics, students must analyze the raw thermodynamic numbers associated with the cleavage of Adenosine Triphosphate. This slide clearly contrasts abstract standard-state measurements with actual physiological conditions. Under perfect standard laboratory conditions, the hydrolysis of the terminal phosphoanhydride bond yields a substantial release of negative 30.5 kilojoules per mole. However, cells are not static laboratory beakers; they are highly dynamic, carefully controlled systems that actively manipulate chemical concentrations to maximize the thermodynamic driving forces available to them.
The crucial concept here is that the cellular concentration of Adenosine Triphosphate is deliberately maintained roughly fivefold higher than in its completely depleted state, essentially freezing the reaction at a point up to ten orders of magnitude away from true thermodynamic equilibrium. Because the biological system is held so far from the balanced equilibrium, the actual physiological energy yield is substantially greater than that at the standard state. In a living cell, the functional energy extracted from this molecule skyrockets to approximately 50-57 kilojoules per mole, providing a massively enhanced energetic punch for biological work.
To further demonstrate the unique energetic properties of these molecular linkages, the slide thoughtfully compares the cleavage of terminal anhydride bonds with that of standard ester bonds. Removing the final phosphate group directly from the nucleoside scaffold provides vastly significantly less usable energy, yielding only about negative 9 kilojoules per mole. This dramatic thermodynamic contrast irrefutably demonstrates that the true energetic power of Adenosine Triphosphate does not simply lie in the presence of phosphorus, but specifically in the strained, highly reactive geometry of the sequential anhydride linkages.
Slide 6: Physical Chemistry Behind Adenosine Triphosphate Energy Release
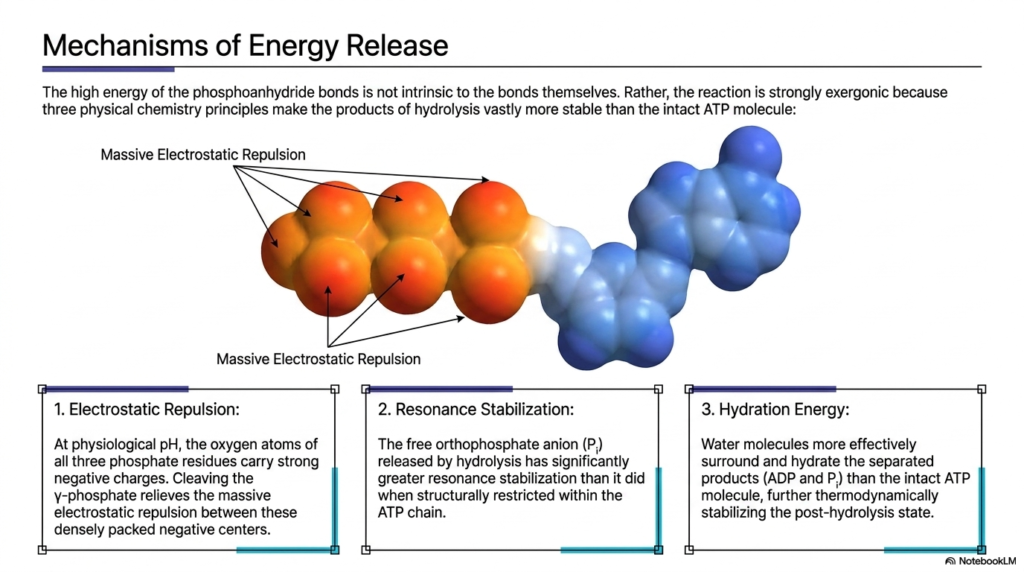
It is a common misconception that the high energy of Adenosine Triphosphate is somehow intrinsically locked entirely within the chemical bonds themselves. This slide fundamentally corrects that oversimplification by explaining that the hydrolysis reaction is strongly exergonic because the resulting post-cleavage products are vastly more thermodynamically stable than the intact molecule. Three major physical chemistry principles collaboratively drive this intense energetic release. The primary driving force is massive electrostatic repulsion; at physiological pH, the densely packed oxygen atoms of the three phosphate residues carry remarkably strong, aggressively competing negative charges.
Cleaving the terminal gamma-phosphate from Adenosine Triphosphate instantly acts like releasing a tightly compressed chemical spring, brilliantly relieving the massive electrostatic repulsion between these densely packed negative atomic centers. Following this explosive separation, the newly freed orthophosphate anion immediately experiences significantly greater resonance stabilization. When physically restricted within the linear triphosphate chain, the electrons had limited mobility. Once liberated into the cellular fluid, the free phosphate can effectively distribute its unstable negative charge symmetrically across multiple oxygen atoms, settling into a much lower and favorable energetic baseline state.
The final stabilizing physical factor contributing to the energetic output of Adenosine Triphosphate is robust hydration energy. Once the molecule is successfully cleaved, numerous surrounding water molecules can much more effectively surround, interact with, and tightly hydrate the newly separated energetic products. This complex aqueous shielding further thermodynamically stabilizes the post-hydrolysis state. By combining intense electrostatic relief, enhanced molecular resonance, and highly favorable water hydration, the complex physical chemistry ensures that the biological breakdown of this universal cellular currency reliably and violently drives vital metabolic reactions forward.
Slide 7: Primary Cellular Mechanisms for Adenosine Triphosphate Production
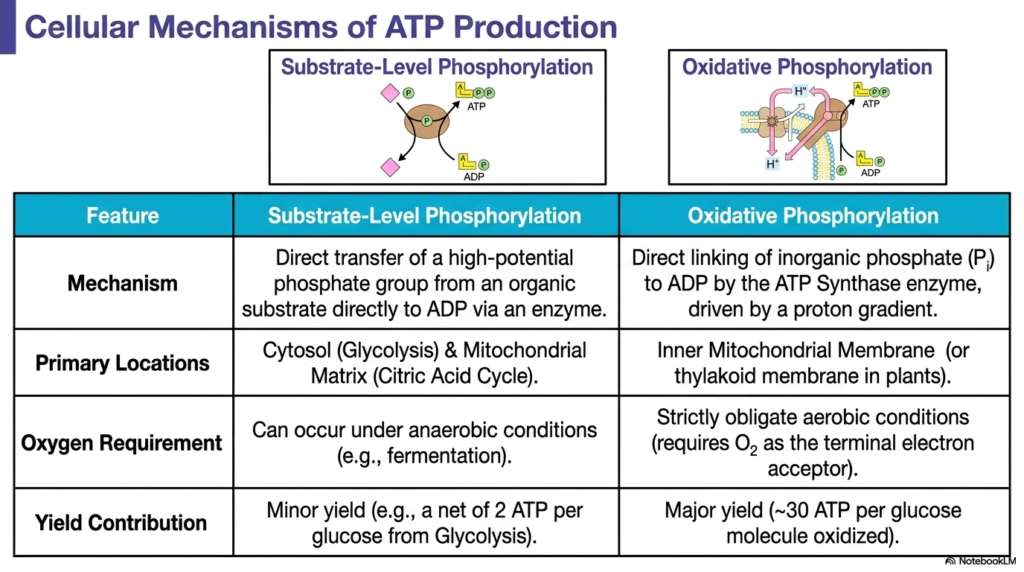
Understanding how a cell spends energy is only half the narrative; students must also comprehend the dual metabolic pathways that produce Adenosine Triphosphate. This slide clearly divides cellular energy production into two incredibly distinct biochemical processes: substrate-level phosphorylation and oxidative phosphorylation. Substrate-level phosphorylation represents the simpler, more direct method. It involves the direct enzymatic transfer of a high-potential phosphate group from an actively reacting organic substrate directly onto a waiting depleted molecular carrier, entirely bypassing the need for complex membrane structures or ionic gradients.
While substrate-level phosphorylation is relatively fast and can seamlessly occur under restrictive anaerobic conditions, it fundamentally provides only a minor contribution to the massive Adenosine Triphosphate demands of a complex living organism. It occurs primarily within the fluid cytosol during glycolysis and yields only a tiny net gain of energy-rich molecules per glucose consumed. In stark contrast, oxidative phosphorylation is an incredibly sophisticated, obligate aerobic process that requires molecular oxygen to function. This complex pathway directly links free inorganic phosphate to the depleted carrier molecule by exploiting a powerful intermembrane proton gradient.
Operating exclusively across the highly folded inner mitochondrial membrane in animal cells, oxidative phosphorylation represents the ultimate evolutionary achievement in Adenosine Triphosphate production. Driven by the remarkable mechanical rotation of a specialized membrane-bound synthase enzyme, this aerobic pathway provides the vast majority of cellular power. Generating roughly thirty high-energy molecules per oxidized glucose, it completely dwarfs the minor energetic yield of primitive substrate-level reactions. This massive production discrepancy beautifully highlights exactly why complex multicellular organisms are entirely dependent on continuous oxygen respiration to survive.
Slide 8: Glycolysis, the Citric Acid Cycle, and Adenosine Triphosphate

Before oxidative mechanisms can generate massive amounts of energy, the cell must meticulously prepare the raw biochemical fuels. This slide outlines how the complete cellular oxidation of standard glucose ultimately provides the vital reducing equivalents strictly necessary to power the bulk of Adenosine Triphosphate synthesis. The preparatory metabolic journey begins in the cytosol with glycolysis, a rapid ten-step enzymatic pathway that systematically breaks down a six-carbon glucose molecule into two molecules of pyruvate. While glycolysis directly yields a small net gain of energetic molecules via substrate-level phosphorylation, its true value lies in chemical preparation.
Following glycolysis, the resulting pyruvate molecules are deeply shuttled into the heavily protected mitochondrial matrix, structurally oxidized into Acetyl-CoA, and continuously fed into the sweeping Citric Acid Cycle. While each turn of this famous biochemical cycle produces a single equivalent of Adenosine Triphosphate through direct substrate-level phosphorylation, its primary physiological purpose is drastically different. The cycle is specifically designed to systematically strip highly reactive, high-energy electrons from the degrading carbon skeleton, aggressively capturing them within specialized, temporary biological transport vehicles known as reduced electron carriers.
These vital chemical carriers, specifically identified as NADH and FADH2, act as highly mobile molecular delivery trucks. They safely transport the highly reactive electrons harvested during glucose breakdown directly to the inner mitochondrial membrane. By temporarily storing the chemical energy originally locked within the covalent bonds of food, these carriers set the ultimate biochemical stage. Their successful generation ensures that the complex membrane machinery responsible for the massive, continuous synthesis of Adenosine Triphosphate has a steady, reliable supply of essential high-energy electrons to power the next critical phase of cellular respiration.
Slide 9: Oxidative Phosphorylation and Adenosine Triphosphate Synthase
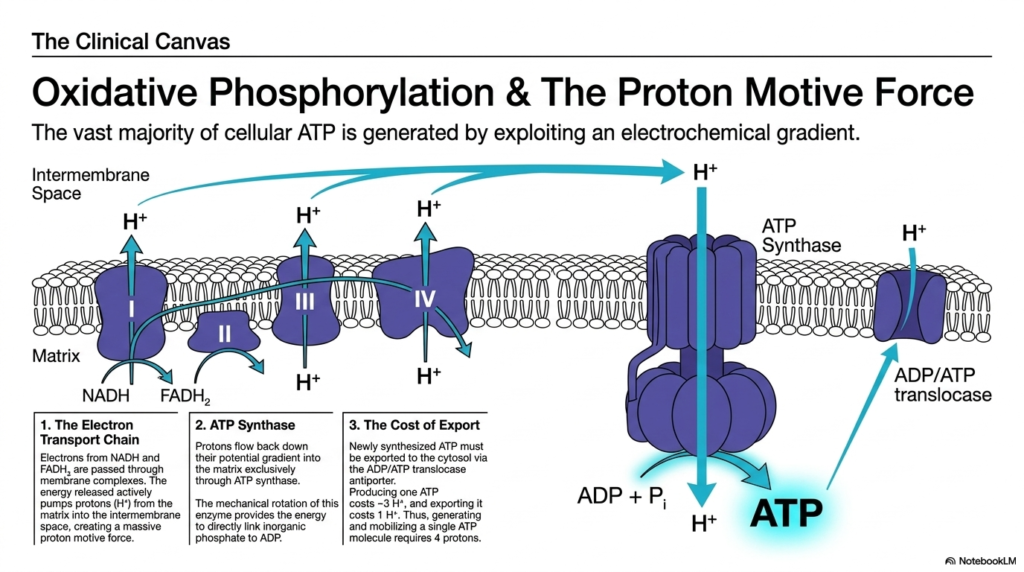
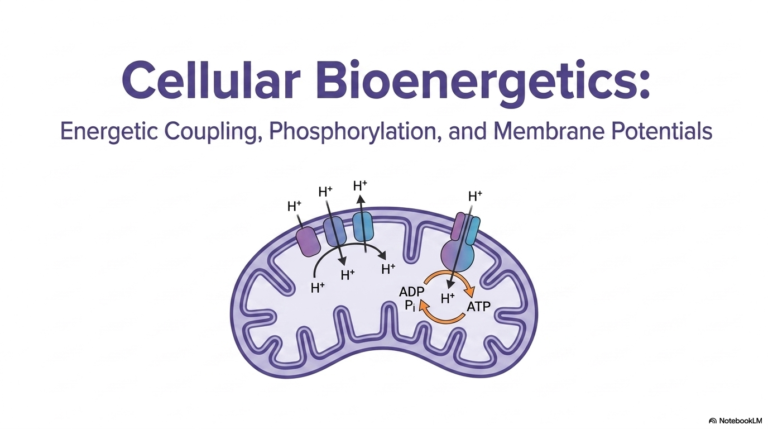
This visually striking slide illustrates the pinnacle of cellular bioenergetics: the intricate electron transport chain and the subsequent generation of the proton-motive force. The highly mobile electron carriers generated in previous metabolic cycles ultimately deliver their volatile payloads to massive protein complexes embedded deep within the mitochondrial membrane. As these reactive electrons are rapidly passed along this complex biological wire, the massive energy released is harnessed to violently pump protons out of the internal matrix. This creates a remarkably dense, highly strained electrochemical gradient across the physical lipid bilayer.
The vast overwhelming majority of cellular Adenosine Triphosphate is synthesized by cleverly exploiting this massive, strained electrochemical gradient. The intensely concentrated protons desperately seek to flow back down their thermodynamic potential gradient into the mitochondrial matrix, but they are physically blocked by the impermeable membrane. Their only available escape route is exclusively through a remarkable, highly specialized biological motor enzyme called ATP Synthase. As energetic protons violently rush through this specific channel, they force the internal protein subunits to rotate at astonishing cellular speeds.
This rapid mechanical rotation brilliantly provides the exact physical activation energy required to directly force a free inorganic phosphate onto a waiting depleted carrier, successfully synthesizing new Adenosine Triphosphate. However, generating the energy is only part of the complex logistical challenge; the newly generated energy must be exported immediately to the wider cell. Mobilizing a single synthesized molecule out of the mitochondria via a specialized antiporter effectively costs the cell an additional proton. Thus, fully generating and successfully mobilizing one usable energetic molecule ultimately requires the thermodynamic power of four highly concentrated protons.
Slide 10: Generating Adenosine Triphosphate Beyond Glucose Catabolism
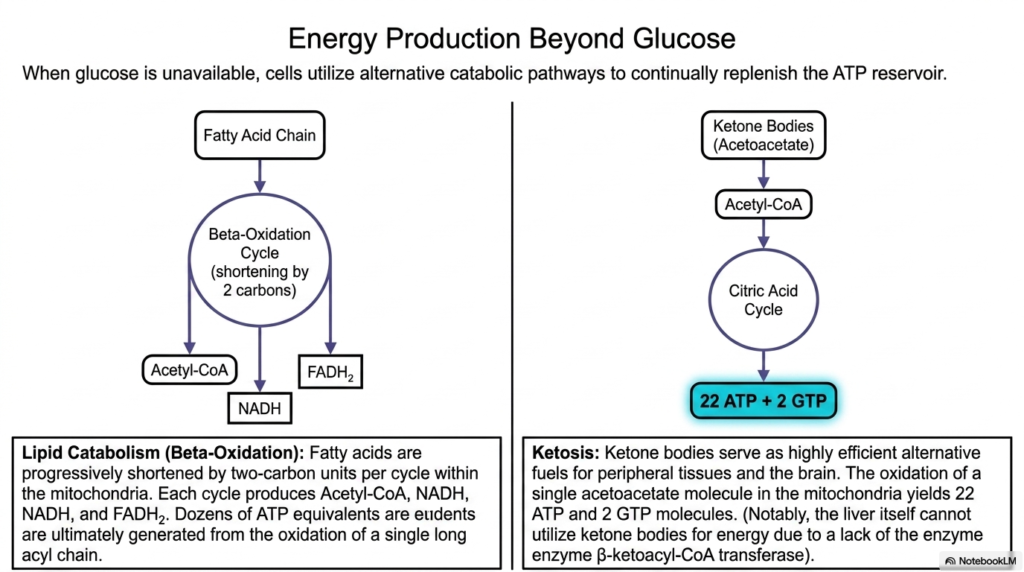
While glucose is famously considered the primary metabolic fuel for human physiology, biological systems are highly adaptable and incredibly resilient. This slide explores the vital alternative catabolic pathways used when readily available circulating carbohydrates are entirely unavailable, ensuring the cellular reservoir of Adenosine Triphosphate is continually replenished during fasting or physiological stress. The most prominent alternative pathway is lipid catabolism, fundamentally driven by the highly efficient beta-oxidation cycle. By systematically shortening dense, heavily reduced fatty acid chains by two-carbon units per cycle within the mitochondria, the cell can harvest massive amounts of chemical energy.
The sequential breakdown of a single long-chain fatty acid generates dozens of essential reducing equivalents and massive quantities of Acetyl-CoA, resulting in the eventual downstream synthesis of staggering amounts of Adenosine Triphosphate. This remarkably high energy yield elegantly explains why biological life evolved to rely so heavily on compact lipid tissues for secure, long-term survival and storage. However, some specialized organs, particularly the heavily protected neural tissues of the brain, cannot effectively utilize circulating lipid chains directly, requiring the stressed liver to rapidly produce highly efficient, water-soluble alternative fuels known as ketone bodies.
The specialized physiological state of ketosis brilliantly demonstrates profound metabolic flexibility. Circulating ketone bodies serve as incredibly highly efficient alternative fuels for both demanding peripheral tissues and the isolated brain. The complete mitochondrial oxidation of a single mobilized ketone molecule smoothly yields an impressive twenty-two molecules of Adenosine Triphosphate. Interestingly, the producing liver cannot, by design, utilize the very ketone bodies it generously synthesizes due to a deliberate, hardwired enzymatic deficiency. This altruistic biochemical safeguard beautifully ensures that vital alternative energy reaches the nervous system smoothly during severe glucose deprivation, without hepatic consumption.
Slide 11: Adenosine Triphosphate in Intracellular Signaling Pathways
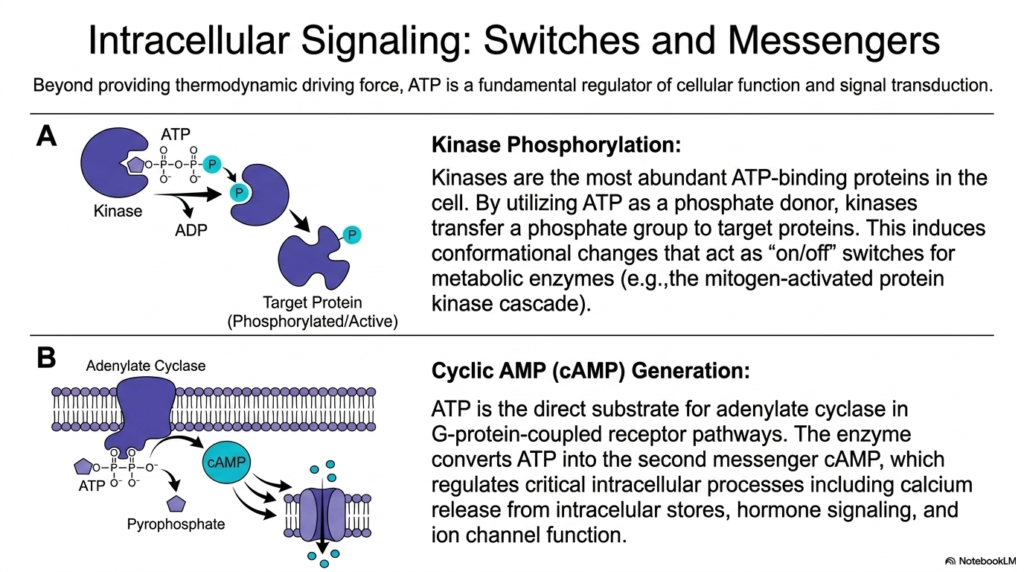
Moving far beyond its famous primary role as a thermodynamic driving force, this important slide reveals how the cell uses its energy currency as a fundamental biochemical regulator. The presence or precise cleavage of Adenosine Triphosphate is deeply integrated into critical intracellular signaling pathways, acting as the ultimate molecular switch for widespread cellular function. The most abundant energy-binding proteins in the living cell are complex kinases. By cleverly using the highly energetic molecule as a direct phosphate donor, these sophisticated enzymes can transfer a bulky, highly charged phosphate group onto specific target proteins.
This targeted, high-energy phosphorylation induces immediate, massive conformational changes in the target protein, effectively acting as an instantaneous “on/off” switch for vital metabolic enzymes and complex signaling cascades. By directly linking protein activation to the immediate availability of Adenosine Triphosphate, the cell intelligently ensures that energetically expensive physiological actions are initiated only when cellular energy levels are fully secure and abundant. This incredibly tight, logical regulatory coupling successfully prevents a stressed cell from accidentally executing biologically costly programs during dangerous periods of severe starvation.
Additionally, the universal energy-carrying molecule serves as the direct physical substrate for membrane-bound adenylate cyclase enzymes in crucial G protein-coupled receptor pathways. When specifically triggered by external hormones, this remarkable enzyme rapidly converts intact Adenosine Triphosphate directly into cyclic AMP, a profoundly important internal second messenger. This rapidly generated signaling molecule quickly diffuses throughout the chaotic cytoplasm, seamlessly regulating deeply critical physiological responses, including massive calcium release and synchronized ion channel function. This beautifully demonstrates how the core energetic medium simultaneously functions as the primary vocabulary for complex internal biological communication.
Slide 12: Adenosine Triphosphate and Building the Machinery of Life
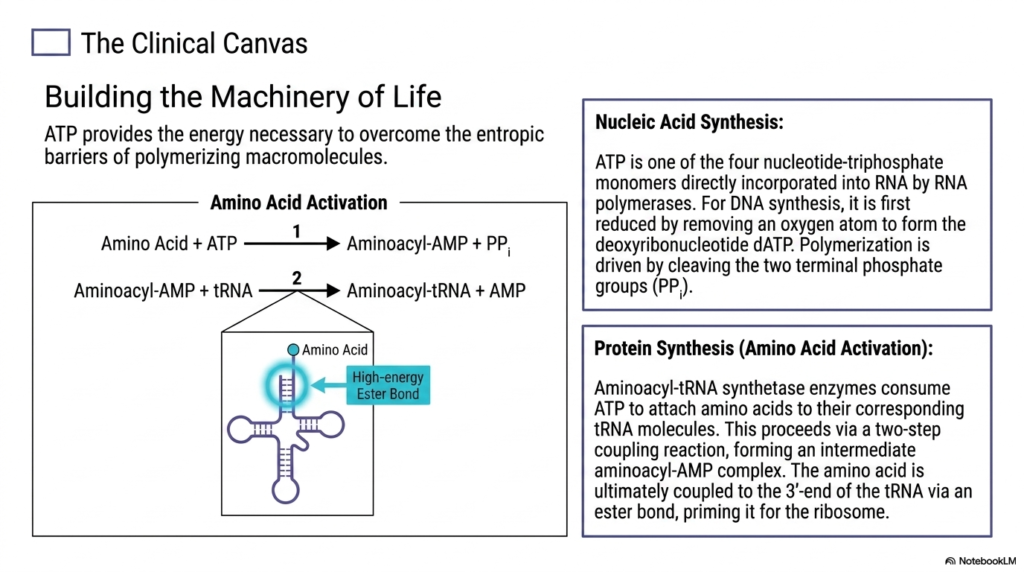
Life fundamentally requires the continuous, highly organized construction of massive macromolecules, a complex process that inherently fights against the universal thermodynamic laws of entropy. This detailed slide explicitly illustrates exactly how Adenosine Triphosphate actively provides the intense, focused chemical energy strictly necessary to overcome these massive entropic barriers. During highly complex nucleic acid synthesis, the universal energy carrier serves a dual function. It is actually one of the four required core nucleotide-triphosphate monomers that are directly incorporated into the growing physical backbone of all active cellular RNA strands by polymerases.
For stable DNA synthesis, the molecule is first carefully and enzymatically reduced by selectively removing a specific oxygen atom, thereby forming the required deoxyribonucleotide variant. In both critical genetic processes, the thermodynamically unfavorable, highly organized polymerization of the growing biological chain is powerfully driven completely forward strictly by violently cleaving the two terminal phosphate groups from the incoming Adenosine Triphosphate. This intense, targeted release of pure chemical energy flawlessly fuels the physical formation of the critical phosphodiester bonds that structurally hold our fundamental genetic code completely intact against chaotic environmental degradation.
The massive energetic requirements of biology are arguably most visible during complex protein synthesis. Specialized synthetase enzymes strictly consume Adenosine Triphosphate to meticulously attach free amino acids directly to their corresponding biological tRNA transport molecules. This strictly proceeds via a brilliant two-step energetic coupling reaction, forming a highly reactive intermediate complex. The activated amino acid is ultimately forcefully coupled to the waiting tRNA molecule via a distinctly high-energy ester bond. This specific energetic priming effectively prepares the amino acid for subsequent rapid, seamless integration into a growing functional protein sequence at the ribosome.
Slide 13: Fueling Mechanical Movement with Adenosine Triphosphate
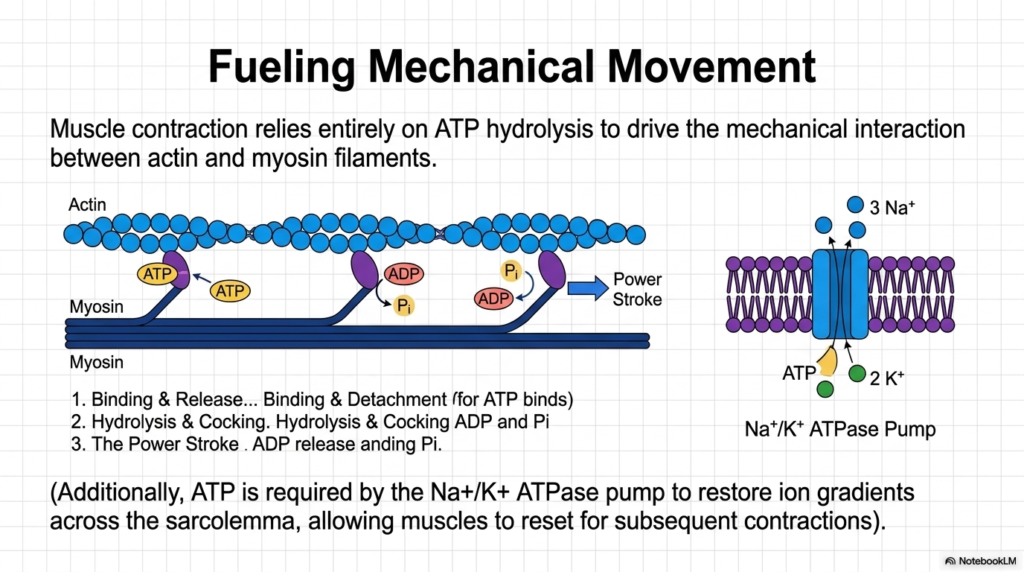
Cellular biochemistry is not solely restricted to microscopic chemical modifications; it frequently translates directly into highly visible, macroscopic physical movement. This engaging slide highlights how large-scale muscle contraction relies entirely on the continuous, rapid hydrolysis of Adenosine Triphosphate to drive the intense mechanical interactions that bind actin and myosin protein filaments. In a relaxed state, the biological myosin head securely holds a partially cleaved energetic complex. When fully activated by calcium, it tightly binds the structural actin cable and forcefully executes a power stroke that shortens the contracting muscle fiber.
Immediately following the violent mechanical power stroke, the completely depleted energetic byproducts are rapidly released. Crucially, the incredibly tight biological bond between the locked muscle proteins can only be successfully broken when a completely fresh, fully intact molecule of Adenosine Triphosphate successfully binds the exhausted myosin head. The subsequent rapid hydrolysis of this newly arrived energetic molecule effectively “cocks” the microscopic protein spring back into its highly strained, high-energy starting position. This astonishingly fast, continuous cycle of binding, cleaving, and releasing literally provides the massive kinetic force that powers every single heartbeat and voluntary human movement.
Furthermore, intense mechanical movement rapidly depletes vital cellular ion gradients that must be strictly maintained for subsequent repeated muscular contractions to occur safely. To successfully reset the heavily disrupted local membrane, the biological cell relies heavily on the highly specialized, energy-dependent sodium-potassium ATPase pump. By directly consuming massive quantities of Adenosine Triphosphate, this incredible membrane-bound machine violently forces displaced ions completely against their natural thermodynamic gradients across the cellular sarcolemma. This immense energetic investment perfectly primes the stressed muscle tissue, allowing it to instantly reset and prepare for the very next critical physical contraction.
Slide 14: Extracellular Purinergic Signaling Using Adenosine Triphosphate
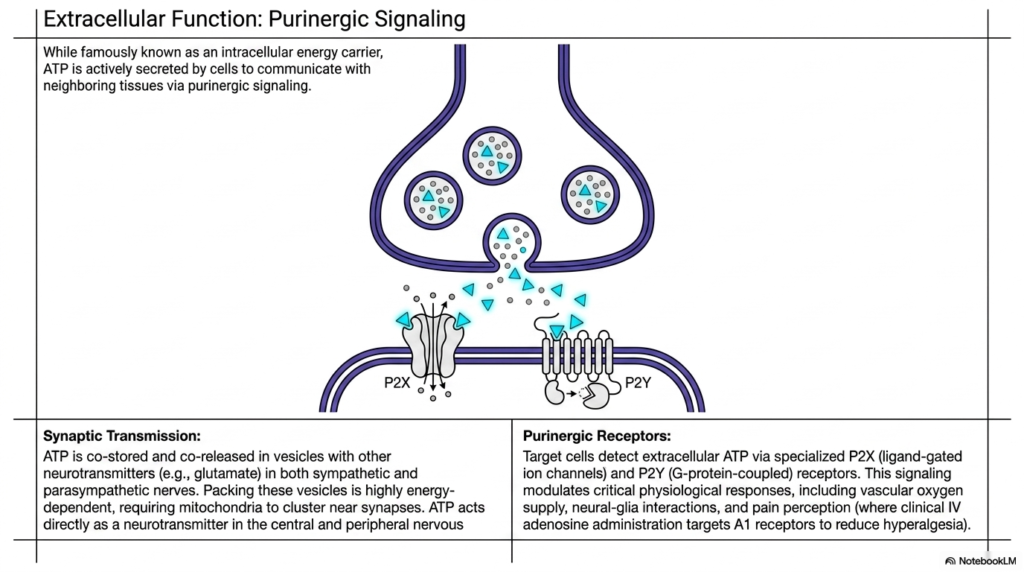
While universally famous as the highly guarded, heavily protected intracellular metabolic energy carrier, this fascinating slide reveals that cells actively and deliberately secrete this highly valuable molecule into the hostile extracellular space to communicate with neighboring tissues. This advanced biological phenomenon is formally known as purinergic signaling. In highly specialized biological synaptic transmission, the energy molecule is frequently continuously co-stored in deep, protective microscopic vesicles alongside traditional, well-known chemical neurotransmitters, such as the highly reactive glutamate, strictly within the complex terminal branches of both the major sympathetic and parasympathetic nerves.
Packing these dense, microscopic biological vesicles with highly charged molecules is an incredibly physically demanding, highly energy-dependent process that directly requires numerous massive mitochondria to cluster extremely close to the active biological synapses. Upon neural activation, the cell deliberately releases intact Adenosine Triphosphate directly into the dangerous extracellular synaptic cleft. Here, it acts entirely directly as a highly potent, functional neurotransmitter within both the central and widespread peripheral nervous systems. Target recipient cells quickly detect this surprising extracellular energy burst via highly specialized, membrane-bound purinergic receptors designed specifically for this task.
These highly specialized external protein receptors fundamentally modulate deeply critical physiological responses completely outside the standard metabolic paradigm. By securely detecting extracellular Adenosine Triphosphate, the complex surrounding biological tissue can seamlessly regulate local vascular oxygen supply, coordinate complex neural-glia interactions, and deeply modulate conscious systemic pain perception. In advanced clinical medical practice, intravenous administration of closely related nucleoside compounds specifically targets the exact same purinergic receptor pathways to successfully reduce dangerous physiological hyperalgesia, beautifully demonstrating the profound medical relevance of extracellular energetic cellular signaling mechanisms.
Slide 15: The Perpetual Cycle and Homeostasis of Adenosine Triphosphate
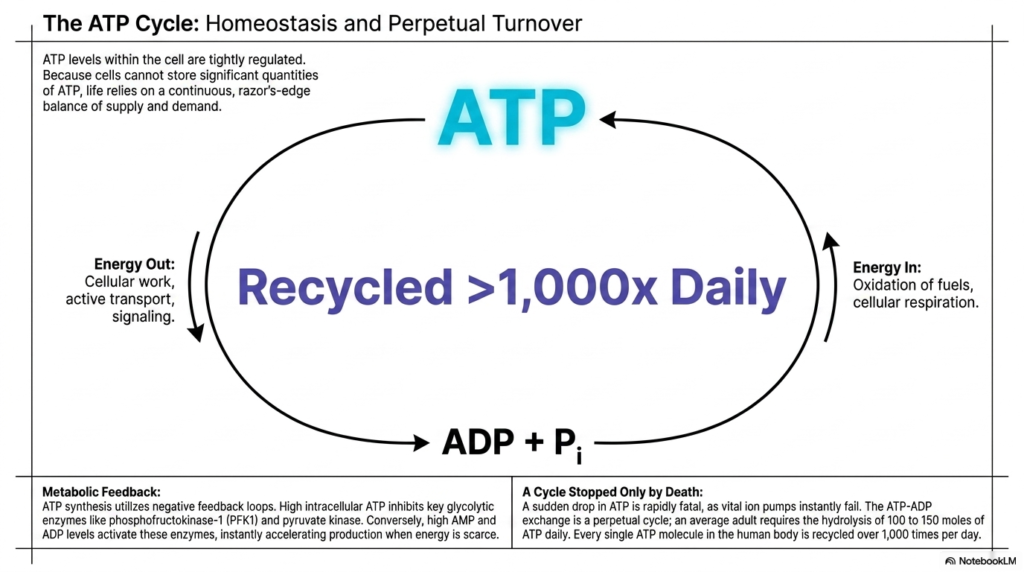
The final concluding slide synthesizes all previous biochemical concepts into the ultimate overarching reality of life: the razor’s-edge balance of biological energy homeostasis. The strict intracellular levels of Adenosine Triphosphate are incredibly tightly regulated at all times. Because complex mammalian cells fundamentally cannot securely store biologically significant quantities of this intensely reactive currency, active living organisms fundamentally rely on a continuous, flawlessly balanced biological loop that safely matches raw energy supply directly to massive, unending physiological demand. Any microscopic deviation in this delicate internal cycle rapidly triggers massive, highly cascading biological consequences.
To successfully maintain this critical homeostasis, the cell brilliantly utilizes powerful, deeply ingrained metabolic feedback loops. Dangerously high intracellular concentrations of synthesized Adenosine Triphosphate chemically act to naturally inhibit highly key, rate-limiting glycolytic biological enzymes, efficiently preventing massively wasteful cellular overproduction. Conversely, when internal energy inevitably drops and cellular byproducts are depleted, these exact same critical enzymes are violently and rapidly activated. This brilliant evolutionary, deeply self-regulating chemical cycle instantly accelerates complex internal metabolic production explicitly only when biologically usable cellular energy is severely, legitimately physically scarce.
This frantic, unending metabolic loop represents a microscopic cellular cycle that is completely stopped only by organismal death. If a massive, sudden, systemic drop in cellular Adenosine Triphosphate occurs, vital, deeply critical transmembrane ion pumps instantly and catastrophically fail, rendering the biological cell chemically dead within minutes. Because an average adult successfully requires the intense daily hydrolysis of roughly one hundred and fifty entire biological moles of this incredibly specific molecule to survive, every single individual energetic molecule in the human body is violently, continuously recycled over one thousand times per day, flawlessly sustaining life.
Please read our Content Disclaimer Statement.
Check out our social media channels: