43. Regulation of Carbohydrate Metabolism: A Comprehensive Guide
Imagine driving a car with both the gas pedal and the brakes pressed to the floor; it wastes fuel and destroys the engine. The human body avoids this metabolic disaster through precise cellular controls. This presentation breaks down the core molecular logic, pathways, and physiological integration involved in energy homeostasis. By exploring these slides, medical and university students will discover exactly how cells seamlessly transition between energy storage and mobilization to keep the human machine running efficiently.
Slide 1: Introduction to the Regulation of Carbohydrate Metabolism
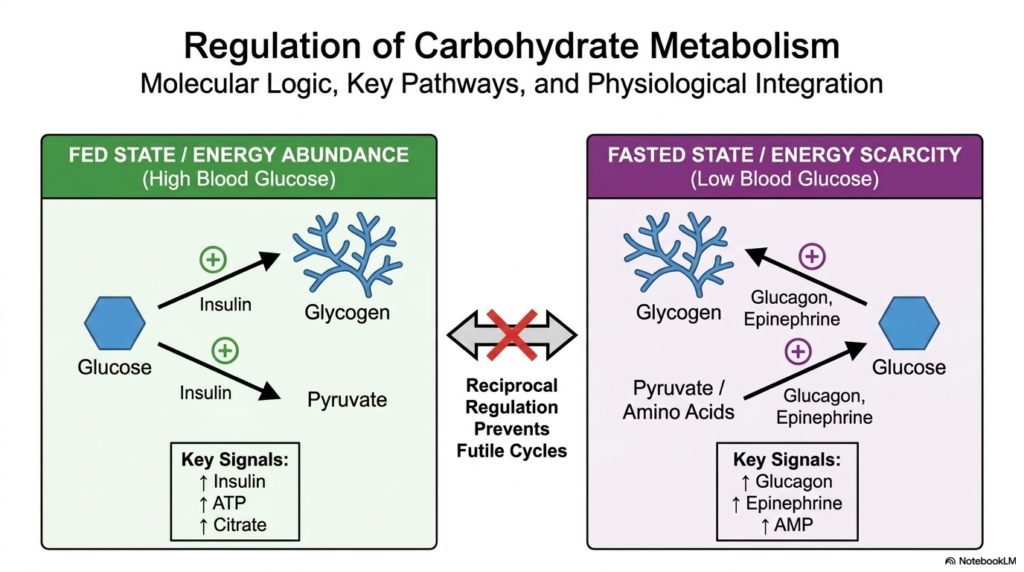
Understanding the core molecular logic of energy states forms the ultimate foundation for the Regulation of Carbohydrate Metabolism. During the fed state, characterized by energy abundance and high blood glucose, the pancreas secretes insulin. This anabolic hormone triggers the liver and muscles to store glucose as highly branched glycogen. Simultaneously, it drives the conversion of glucose into pyruvate for immediate energy utilization. This beautifully coordinated response ensures that excess dietary nutrients are safely sequestered. The Regulation of Carbohydrate Metabolism depends entirely on these rapid, insulin-mediated physiological shifts to maintain postprandial balance and prevent prolonged hyperglycemia.
Conversely, the fasted state plunges the body into energy scarcity, marked by a drop in blood sugar. During this critical window, hormones like glucagon and epinephrine dominate the physiological landscape. These catabolic messengers instruct the liver to rapidly break down its stored glycogen reserves and push glucose back into the bloodstream. Additionally, they stimulate the production of entirely new glucose molecules from precursors such as pyruvate and amino acids mobilized from protein. Proper Regulation of Carbohydrate Metabolism ensures that the brain and erythrocytes are never starved of their primary energetic fuel during fasting.
The absolute masterpiece of this metabolic architecture is reciprocal regulation, a system perfectly engineered to prevent futile cycles. If the body simultaneously synthesized and broke down glycogen at high rates, it would merely burn valuable ATP without doing any useful biological work. The body avoids this by turning one pathway off while turning the opposite pathway on. This flawless reciprocal control acts as an incredibly sensitive metabolic traffic light, safely keeping opposing pathways separated.
This reciprocal control operates at multiple levels of biological organization. From millisecond allosteric interactions within the cytosol to longer-term hormonal modifications via covalent phosphorylation, the cell utilizes diverse mechanisms to ensure unidirectional metabolite flow. Mastering this molecular logic gives students a profound appreciation for how tightly the body guards its energy reserves, seamlessly directing the flow of biochemical energy in response to the organism’s changing nutritional landscape.
Slide 2: The Liver’s Role in the Regulation of Carbohydrate Metabolism
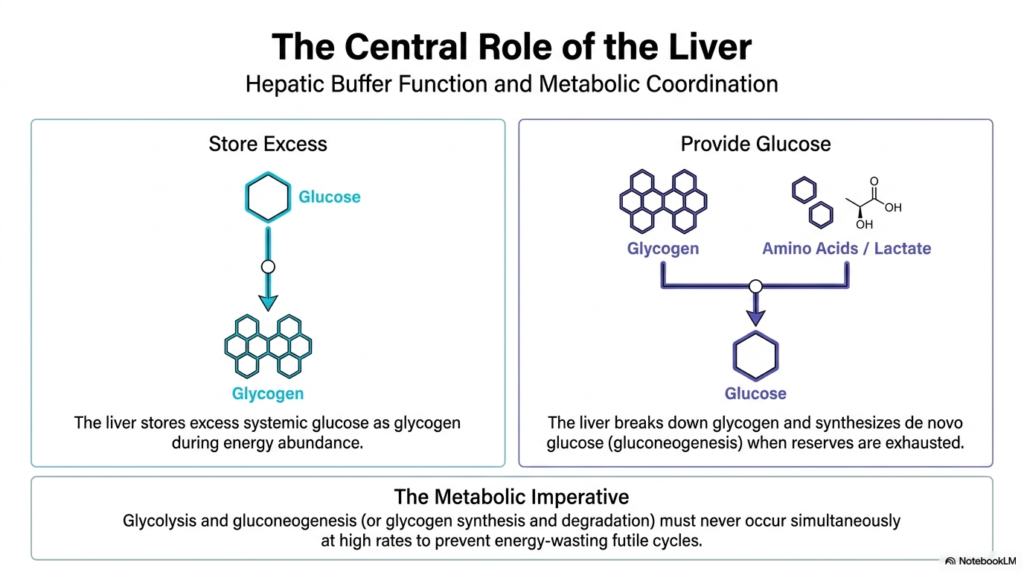
The liver functions as the ultimate metabolic altruist, playing an incredibly central role in the Regulation of Carbohydrate Metabolism. While tissues such as skeletal muscle consume glucose strictly for their own energy needs, the liver acts as a massive systemic buffer. During periods of caloric abundance, hepatic cells aggressively take up excess circulating glucose. This incoming sugar is then rapidly polymerized into complex glycogen granules for efficient storage. By clearing this post-meal glucose spike, the liver prevents dangerous osmotic imbalances. Therefore, the Regulation of Carbohydrate Metabolism is intimately tied to hepatic function and its ability to sense systemic nutritional status.
When energy reserves begin to dwindle between meals, the liver’s altruistic nature truly shines. Sensing the drop in blood sugar, hepatic tissues initiate rapid glycogen breakdown to release free glucose back into circulation. When these short-term glycogen stores are fully exhausted, the liver shifts to its backup protocol. It begins synthesizing de novo glucose from non-carbohydrate precursors, such as amino acids derived from muscle breakdown. This crucial aspect of the Regulation of Carbohydrate Metabolism helps maintain stable systemic glucose levels during prolonged fasting or intense physiological stress.
This complex balancing act highlights a fundamental metabolic imperative: the absolute prevention of simultaneous, opposing pathways. The liver simply cannot afford to perform glycolysis to burn glucose while simultaneously running gluconeogenesis to create it. Doing so would rapidly deplete cellular ATP reserves, resulting in a disastrous and energy-wasting futile cycle. To prevent this, the biochemical pathways of synthesis and degradation are reciprocally controlled. When the liver is instructed to store energy, all degradation machinery is actively silenced.
Students must recognize that this hepatic buffer function relies on highly specific enzyme isoforms found only in liver tissue. Unlike muscle cells, liver cells express specialized glucose transporters and enzymes that allow bidirectional metabolite flow across the cell membrane. This unique biochemical toolkit equips the liver to act as the central clearinghouse for macronutrients, constantly evaluating systemic hormonal signals to dictate whether the body should store fuel or aggressively mobilize its carefully guarded reserves.
Slide 3: Irreversible Bypass Reactions in the Regulation of Carbohydrate Metabolism
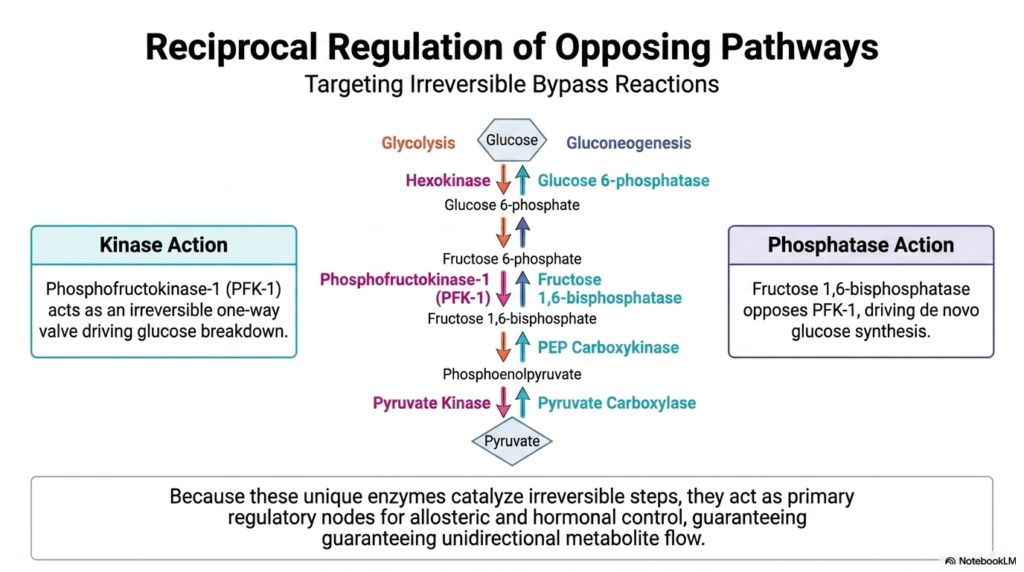
To strictly enforce metabolic directionality, the Regulation of Carbohydrate Metabolism targets key irreversible bypass reactions. In glycolysis, enzymes like hexokinase, phosphofructokinase-1 (PFK-1), and pyruvate kinase act as one-way valves. These kinases catalyze reactions with massive drops in free energy, meaning they physically cannot run backward under normal cellular conditions. This thermodynamic finality is essential because it forcefully pulls metabolites through the catabolic pathway. By serving as unidirectional checkpoints, these kinases provide perfect targets for cellular regulation. The Regulation of Carbohydrate Metabolism relies on regulating these specific enzymatic valves to control the overall rate of glucose breakdown.
When the cell needs to reverse this flow and build glucose during gluconeogenesis, it cannot simply use those same kinase enzymes. Instead, the cell uses specialized phosphatases to bypass these thermodynamic roadblocks. Enzymes such as glucose 6-phosphatase and fructose 1,6-bisphosphatase, along with the combined action of specific carboxylases, provide distinct alternative routes. These gluconeogenic enzymes strip phosphate groups off metabolites, driving the production of new glucose molecules. Effective Regulation of Carbohydrate Metabolism is achieved by ensuring that these bypass phosphatases are active only when their opposing kinases are inactivated.
Because these specific enzymes govern irreversible steps, they function as the primary regulatory nodes for the entire metabolic network. They are exquisitely sensitive to allosteric effectors and hormonal signals, allowing the cell to rapidly adjust enzymatic activity in response to changing energy demands. By having distinct enzymes for the forward and reverse reactions, the cell gains independent control over glycolysis and gluconeogenesis. This physical separation of regulatory targets guarantees unidirectional metabolite flow and prevents chaotic substrate cycling.
For medical students, understanding these bypass reactions is absolutely critical for grasping metabolic pharmacology and pathology. Many genetic metabolic disorders stem from mutations in these specific bypass enzymes. For example, a deficiency in glucose 6-phosphatase traps glucose in the liver, preventing its release and causing severe fasting hypoglycemia. By mastering how these irreversible nodes operate, future physicians can better understand how hormonal imbalances disrupt cellular energy homeostasis at the most fundamental molecular level.
Slide 4: Energy Charge and the Regulation of Carbohydrate Metabolism

At the most fundamental cellular level, the Regulation of Carbohydrate Metabolism is largely dictated by allosteric control via the cellular energy charge. When a cell has abundant fuel, it enters a high-energy state characterized by elevated levels of ATP, citrate, and acetyl-CoA. These energy-rich molecules do not simply sit idle; they act as powerful allosteric feedback inhibitors. By binding to specific regulatory sites on key glycolytic enzymes, they induce conformational changes that dramatically reduce enzymatic activity. This intrinsic Regulation of Carbohydrate Metabolism ensures that a cell does not needlessly break down glucose when its internal batteries are already completely charged.
Conversely, when a cell is forced to do heavy work or is starved of nutrients, its energy charge rapidly plummets. This low-energy state is biochemically signaled by rising concentrations of AMP and ADP, which indicate a severe deficit in cellular ATP. These molecules act as potent allosteric activators. Elevated AMP levels instantly bind to and supercharge the enzymes responsible for glycogen degradation and glycolysis. Simultaneously, AMP acts as a strict inhibitor of gluconeogenesis. The cellular Regulation of Carbohydrate Metabolism uses this AMP signal as an emergency alarm to instantly prioritize immediate energy production.
This allosteric system allows individual cells to make highly localized, split-second metabolic decisions independent of systemic hormones. Long before insulin or glucagon levels change in the bloodstream, a contracting muscle cell can sense its own rising ADP levels and instantly accelerate its glycolytic rate to survive. This localized metabolite feedback is the absolute first line of defense against cellular energy depletion, keeping energy production perfectly matched to energy expenditure at all times.
Understanding the nuances of energy charge is vital for comprehending how different tissues adapt to stress. While the liver manages whole-body glucose levels, the individual cells within muscles or the brain must still protect their internal ATP pools to maintain vital cellular processes. This beautiful interplay between highly sensitive enzymes and fluctuating metabolite pools showcases the remarkable evolutionary refinement of cellular metabolism, prioritizing survival through exquisite, immediate allosteric feedback.
Slide 5: Fructose 2,6-bisphosphate in the Regulation of Carbohydrate Metabolism
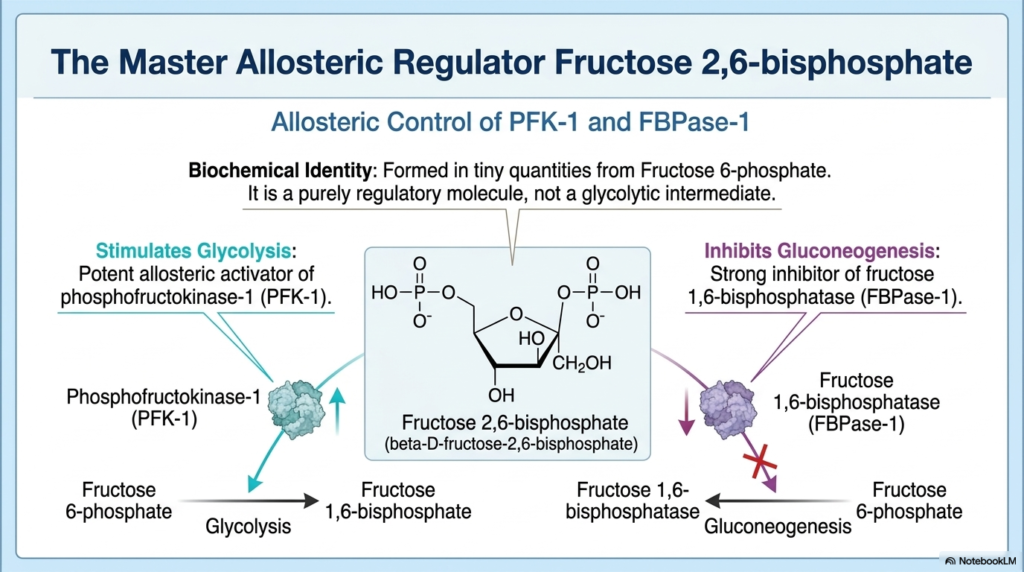
Beyond basic ATP sensing, the Regulation of Carbohydrate Metabolism employs highly specialized molecules like Fructose 2,6-bisphosphate. Unlike standard metabolic intermediates that are consumed to produce energy, Fructose 2,6-bisphosphate is synthesized in tiny, distinct quantities solely to act as a master allosteric regulator. It is structurally derived from Fructose 6-phosphate but does not participate directly in the glycolytic cascade. Instead, it serves as an incredibly potent intracellular signal. The Regulation of Carbohydrate Metabolism relies heavily on this unique molecule to amplify hormonal signals, bridging the gap between endocrine messages and cytosolic enzymatic activity.
The primary biochemical role of Fructose 2,6-bisphosphate is to act as a massive stimulant for glycolysis. It achieves this by binding directly to phosphofructokinase-1 (PFK-1), supercharging the enzyme’s affinity for its substrate while simultaneously shielding it from ATP-induced inhibition. This means that when Fructose 2,6-bisphosphate is present, glycolysis will race forward even if cellular ATP levels are relatively high. This potent activation ensures that the Regulation of Carbohydrate Metabolism can compel the liver to aggressively take up glucose and convert it into storage molecules during periods of extreme caloric abundance.
Simultaneously, Fructose 2,6-bisphosphate exerts massive reciprocal control by acting as a strong inhibitor of gluconeogenesis. It binds tightly to fructose 1,6-bisphosphatase (FBPase-1), drastically reducing its catalytic efficiency. By simultaneously stepping on the gas pedal for glycolysis and slamming the brakes on gluconeogenesis, this master regulator ensures perfect unidirectional flow. It completely prevents the futile cycling of carbon skeletons between these two opposing pathways, carrying out the liver’s systemic mandate to strictly store excess energy.
Students should appreciate the brilliant efficiency of this regulatory system. By relying on a single master regulator to simultaneously control two opposing enzymes, the cell avoids the need for complex, uncoordinated signaling networks. A simple fluctuation in the concentration of Fructose 2,6-bisphosphate cleanly flips the entire metabolic switch of the cell. This elegant molecular mechanism perfectly demonstrates how slight structural modifications to basic sugars transform them into overarching regulators of mammalian energy homeostasis.
Slide 6: Bifunctional Enzymes Governing the Regulation of Carbohydrate Metabolism
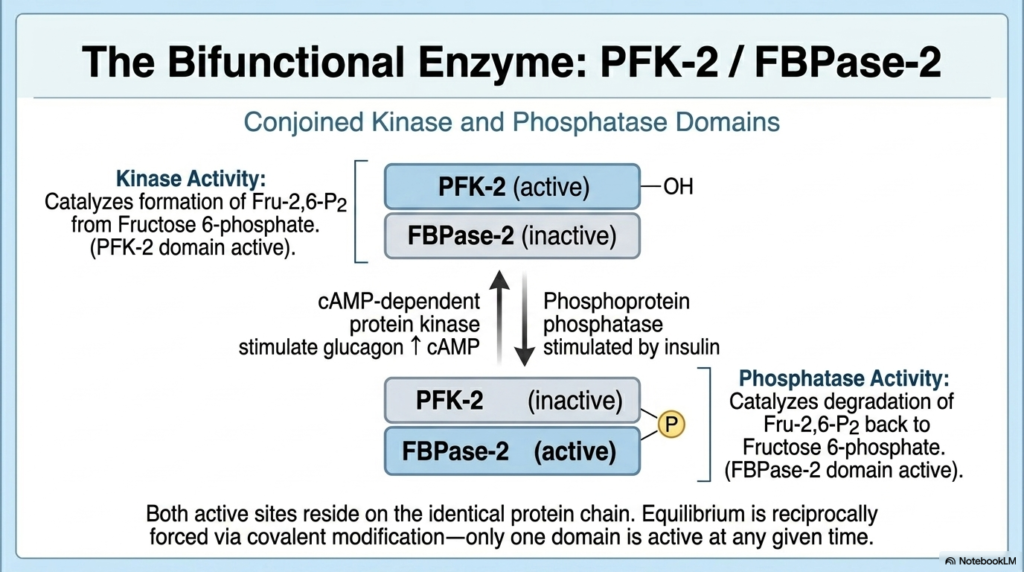
The synthesis and destruction of the master regulator molecule is governed by a remarkable biochemical marvel, central to the Regulation of Carbohydrate Metabolism. This marvel is the bifunctional enzyme known as PFK-2/FBPase-2. Unlike typical enzymes that catalyze a single reaction, this unique protein possesses two distinct, diametrically opposed active sites physically conjoined on the exact same polypeptide chain. One domain, the kinase (PFK-2), is responsible for creating Fructose 2,6-bisphosphate. Proper Regulation of Carbohydrate Metabolism leverages this kinase activity to drive glycolysis forward when fuel is abundant and needs processing.
The opposite domain on this very same protein chain acts as a phosphatase (FBPase-2). When active, this phosphatase domain catalyzes the rapid degradation of Fructose 2,6-bisphosphate, converting it back into standard Fructose 6-phosphate. By destroying the master regulator, this domain effectively shuts down glycolysis and violently removes the inhibition on gluconeogenesis. The Regulation of Carbohydrate Metabolism uses this phosphatase activity to seamlessly switch the liver into a glucose-producing factory during periods of fasting, ensuring that life-saving glucose is mobilized for the brain.
The true genius of this bifunctional enzyme lies in its reciprocal equilibrium, which is tightly controlled via covalent modification. Both active sites reside on identical protein chains, but they are never fully active at the exact same time. The balance between the kinase and phosphatase domains is dictated by phosphorylation. When the protein is physically phosphorylated by external signals, one domain turns off while the other turns on. This physical coupling guarantees that the cell cannot simultaneously build and destroy its master regulatory molecule.
For medical professionals, understanding this bifunctional enzyme provides deep insight into how cellular signaling cascades execute highly specific physical changes in target proteins. It beautifully illustrates the concept of allostery and covalent modification working in tandem. By packing two opposing functions into a single macromolecular machine, the body ensures an instantaneous, coordinated, and biologically flawless transition between metabolic states, preventing catastrophic drops or spikes in systemic blood sugar levels.
Slide 7: Signal Transduction within the Regulation of Carbohydrate Metabolism
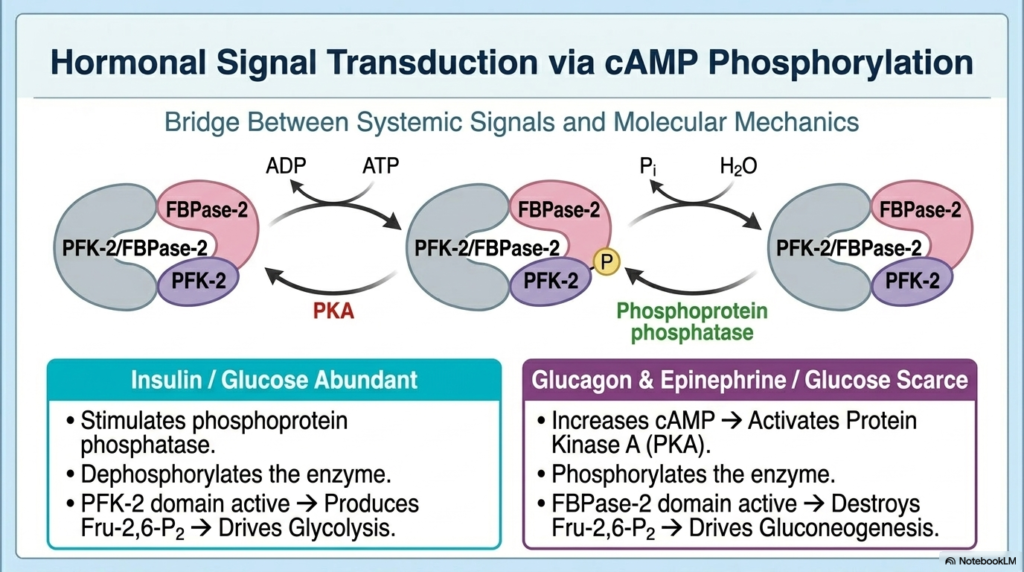
To physically flip the switch on the bifunctional enzyme, the Regulation of Carbohydrate Metabolism utilizes hormonal signal transduction via cAMP phosphorylation. When systemic glucose is scarce, the pancreas releases glucagon, and the adrenals release epinephrine. These survival hormones bind to cellular receptors, triggering a massive intracellular spike in cyclic AMP (cAMP). This secondary messenger rapidly activates Protein Kinase A (PKA). PKA then physically attaches a bulky, negatively charged phosphate group onto the bifunctional enzyme. The Regulation of Carbohydrate Metabolism relies on this precise covalent modification to translate a systemic fasting signal into concrete molecular action.
This phosphorylation profoundly alters the shape and function of the bifunctional enzyme. The addition of the phosphate group completely inactivates the PFK-2 kinase domain while simultaneously supercharging the FBPase-2 phosphatase domain. Consequently, the cell rapidly destroys all its Fructose 2,6-bisphosphate. With the master activator gone, glycolysis grinds to a halt, and gluconeogenesis is fully unleashed. Through this elegant signaling cascade, the Regulation of Carbohydrate Metabolism ensures that the liver rapidly responds to starvation by becoming a powerful engine for de novo glucose synthesis, rescuing systemic blood sugar levels.
Conversely, during periods of energy abundance, insulin dominates the hormonal landscape. Insulin signaling actively degrades cAMP and strongly activates another cellular enzyme called phosphoprotein phosphatase. This enzyme functions as a molecular eraser, selectively clipping the phosphate group right off the bifunctional enzyme. This dephosphorylation event flawlessly reverses the metabolic state. It reactivates the PFK-2 domain to produce Fructose 2,6-bisphosphate while silencing the FBPase-2 domain, thereby forcing glycolysis to reactivate and process incoming postprandial glucose.
This slide beautifully highlights the critical bridge between systemic endocrinology and molecular biochemistry. Students must grasp that hormones do not magically alter metabolism; they initiate highly specific, structurally transformative protein modifications. The addition or removal of a single phosphate group drastically alters the electrostatic interactions within the enzyme’s core. This mechanism demonstrates the incredible speed and precision with which human cells can completely reorganize their internal metabolic machinery in response to a simple meal or a fast.
Slide 8: Hepatic Override and the Regulation of Carbohydrate Metabolism
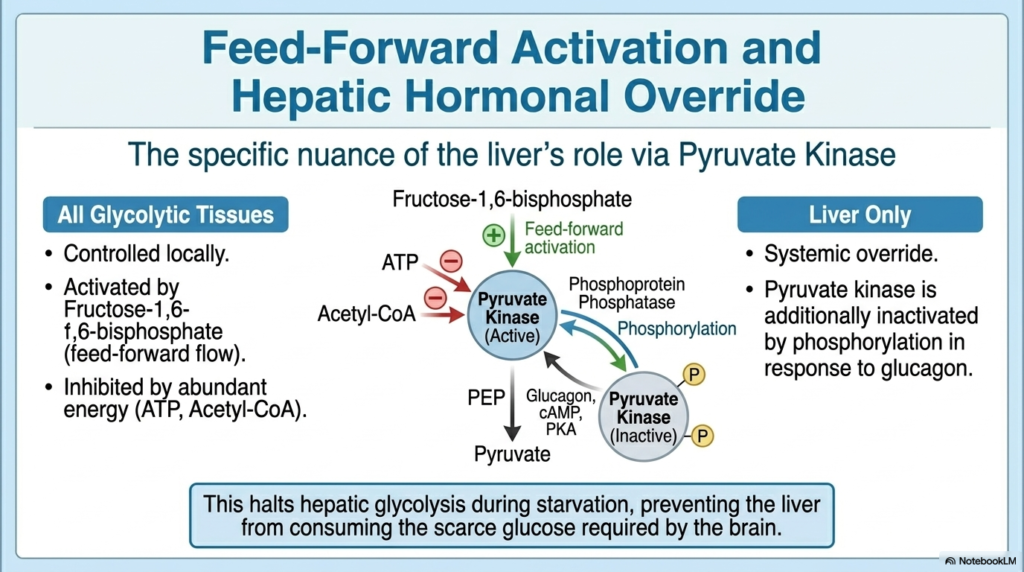
While glycolysis operates in many tissues, the Regulation of Carbohydrate Metabolism introduces specific nuances for hepatic function, particularly concerning feed-forward activation and hormonal override. In all standard glycolytic tissues, the enzyme pyruvate kinase is heavily controlled by local metabolites. It experiences strong feed-forward activation by Fructose-1,6-bisphosphate, meaning an upstream intermediate essentially yells ahead to speed up the final steps of the pathway. Concurrently, it is inhibited by abundant cellular energy markers, such as ATP. This localized Regulation of Carbohydrate Metabolism ensures that general tissues only burn glucose when they specifically require immediate energy.
However, the liver requires a completely different set of rules to fulfill its role as the overarching systemic glucose buffer. In the liver, pyruvate kinase is additionally regulated by a powerful systemic hormonal override. When blood sugar drops and glucagon is released, the resulting cAMP cascade not only shifts the bifunctional enzyme but also directly phosphorylates hepatic pyruvate kinase. This phosphorylation violently inactivates the enzyme, completely shutting off the final step of hepatic glycolysis. This incredibly specific Regulation of Carbohydrate Metabolism occurs exclusively in the liver to prevent the organ from burning the glucose it is trying to synthesize.
This targeted hepatic shutdown is an absolute masterpiece of metabolic engineering. During starvation, the liver works tirelessly to generate glucose via gluconeogenesis to save the brain. If hepatic pyruvate kinase remained active, the newly formed glucose precursors would simply fall back down the glycolytic pathway, creating a disastrous futile cycle within the liver itself. By covalently inactivating this specific enzyme isoform, glucagon ensures that all hepatic carbon flow is strictly directed upward toward glucose release, aggressively halting any downward glycolytic flux.
This mechanism clearly illustrates the profound importance of tissue-specific enzyme isoforms. Muscle tissue completely lacks this specific phosphorylation site on its version of pyruvate kinase. Consequently, during a fight-or-flight response, epinephrine can halt liver glycolysis to mobilize glucose while simultaneously allowing muscle glycolysis to massively accelerate to power sprinting or fighting. Understanding these tissue-specific differences is paramount for students studying the highly specialized nature of human endocrinology.
Slide 9: The Pyruvate Crossroads in the Regulation of Carbohydrate Metabolism
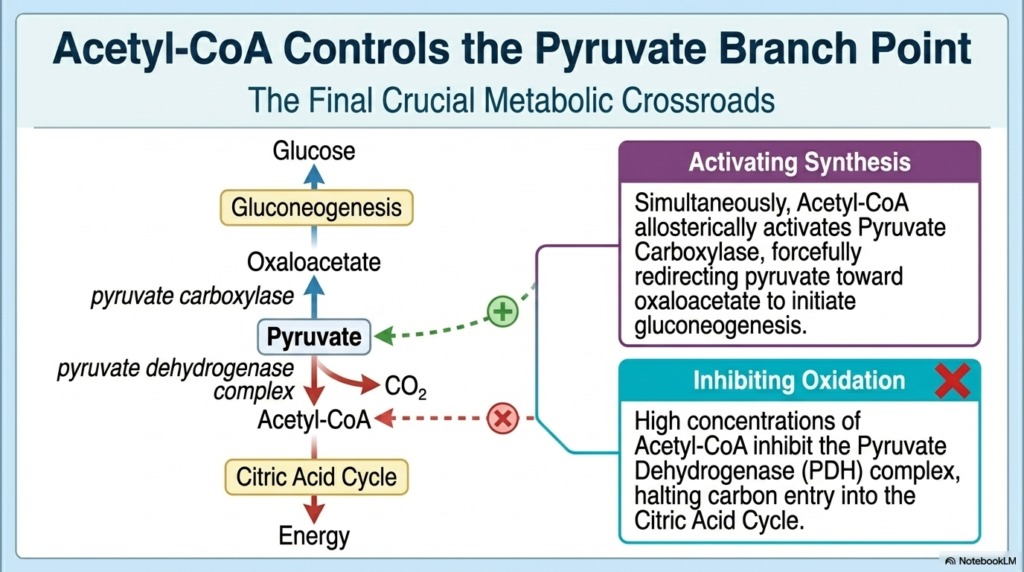
As metabolites flow deeper into the cell, Acetyl-CoA emerges as the final crucial crossroads in the Regulation of Carbohydrate Metabolism. When the cell breaks down abundant fats or carbohydrates, mitochondrial levels of Acetyl-CoA rise dramatically. This two-carbon molecule acts as the ultimate gatekeeper dictating the fate of pyruvate. High concentrations of Acetyl-CoA allosterically bind to and strongly inhibit the Pyruvate Dehydrogenase (PDH) complex. This crucial step in the Regulation of Carbohydrate Metabolism slams the door shut on further carbon entry into the Citric Acid Cycle, signaling that the cell’s energetic needs have been entirely met.
Simultaneously, this exact same pool of Acetyl-CoA acts as an incredibly potent allosteric activator for an entirely different enzyme: Pyruvate Carboxylase. By activating Pyruvate Carboxylase, Acetyl-CoA forcefully redirects the accumulating pyruvate away from oxidation and directs it into the synthesis of oxaloacetate. This represents the very first committed step of massive gluconeogenesis. The cellular Regulation of Carbohydrate Metabolism utilizes Acetyl-CoA not just as fuel, but as a sophisticated metabolic traffic cop, directly forcing carbon skeletons to flow upwards into glucose production.
This elegant divergence is critical during periods of fasting or intense exercise, when the body relies heavily on fat oxidation. As fatty acids are broken down, they flood the liver mitochondria with Acetyl-CoA. This massive influx logically tells the liver that it no longer needs to burn precious glucose for energy. By simultaneously inhibiting PDH and activating Pyruvate Carboxylase, the lipid-derived Acetyl-CoA forcefully preserves existing pyruvate and channels it directly toward brain-saving via systemic glucose generation.
For medical students, this slide bridges the vital gap between lipid and carbohydrate metabolism. It demonstrates that biochemical pathways do not operate in isolated silos; they are deeply physically entwined. A surge in fat breakdown directly causes a mandatory shift in carbohydrate synthesis. Grasping this specific mitochondrial crossroads is essential for understanding complex physiological states such as diabetic ketoacidosis, where massive lipid oxidation overwhelms these precise regulatory checkpoints.
Slide 10: Glycogen Storage and the Regulation of Carbohydrate Metabolism
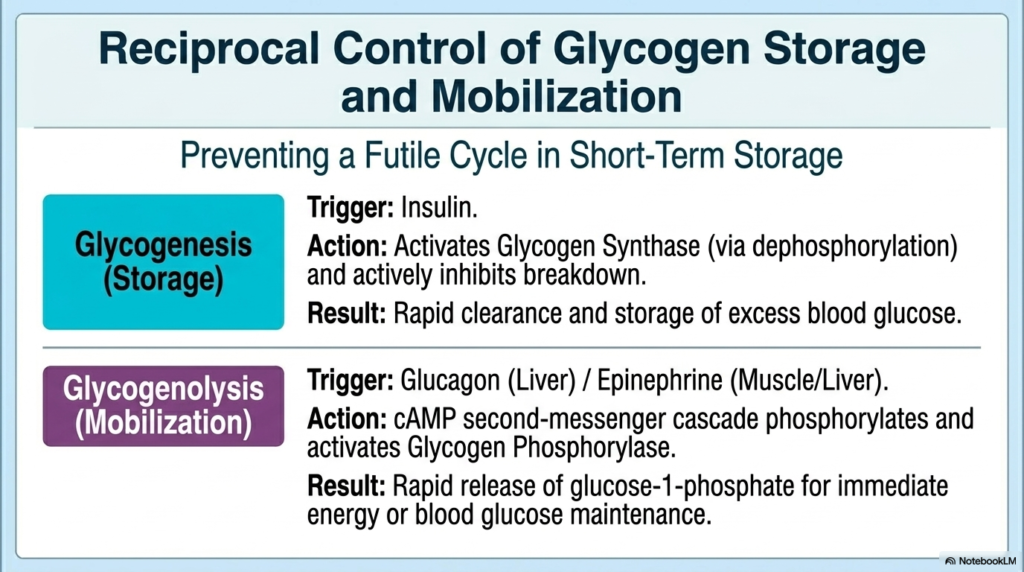
Beyond active energy processing, the Regulation of Carbohydrate Metabolism requires strict, reciprocal control of short-term energy storage as glycogen. To prevent another disastrous, futile cycle, the pathways of glycogenesis and glycogenolysis are never active simultaneously. During the fed state, insulin is the primary trigger of glycogenesis. Insulin initiates a powerful dephosphorylation cascade that chemically activates Glycogen Synthase, the enzyme responsible for building glycogen chains, while aggressively inhibiting the enzymes responsible for its breakdown. This effective Regulation of Carbohydrate Metabolism ensures the rapid clearance and safe storage of excess blood glucose.
Conversely, when blood sugar plummets, the body must tap into these biological reserves immediately. Hormones like glucagon in the liver and epinephrine in both muscle and liver tissues trigger the massive mobilization phase known as glycogenolysis. These hormones initiate a classic cAMP second-messenger cascade. This cascade forcefully phosphorylates and activates Glycogen Phosphorylase, the enzyme that acts like molecular scissors to cleave glucose molecules off the glycogen polymer. This rapid Regulation of Carbohydrate Metabolism floods the cell with glucose-1-phosphate, providing instant energy or systemic blood glucose maintenance.
The genius of this specific system lies in its absolute reciprocity, driven once again by covalent phosphorylation. The exact same hormonal signals that phosphorylate and activate the breakdown enzyme simultaneously phosphorylate and inactivate the synthesis enzyme. It is a brilliant, failsafe molecular switch. The body physically cannot build a glycogen branch and cut it off at the exact same moment. This guarantees that metabolic intent is executed flawlessly without ever wasting a single molecule of precious cellular ATP.
Understanding this strict on/off dynamic is deeply crucial for clinical applications. Many debilitating glycogen storage diseases arise when these regulatory enzymes are mutated or entirely missing. If a patient cannot properly activate glycogen phosphorylase due to a genetic defect, their liver will engorge with inaccessible glycogen while they suffer from severe, life-threatening fasting hypoglycemia. Mastering these reciprocal cascades provides students with the foundational knowledge needed to understand the molecular basis of these complex metabolic disorders.
Slide 11: The Pentose Phosphate Pathway in the Regulation of Carbohydrate Metabolism

While energy generation is vital, the Regulation of Carbohydrate Metabolism also incorporates parallel systems for cellular biosynthesis via the Pentose Phosphate Pathway. This offshoot of standard glycolysis serves two essential biological functions unrelated to ATP production. First, it is the cell’s primary engine for generating reductive power in the form of NADPH. Unlike NADH, which is oxidized for energy, NADPH provides the essential reducing equivalents required to synthesize large biomolecules such as fatty acids and cholesterol. Proper Regulation of Carbohydrate Metabolism ensures the cell has the chemical building blocks necessary for structural growth.
Furthermore, NADPH is absolutely critical for cellular redox balance and for aggressively defending against toxic oxidative stress. It is the core molecule that recycles glutathione, the cell’s premier antioxidant, protecting tissues from catastrophic free radical damage. The second major function of the Pentose Phosphate Pathway is the massive synthesis of Ribose-5-phosphate. This five-carbon sugar is the fundamental structural backbone of DNA, RNA, and vital cellular coenzymes. The integrated Regulation of Carbohydrate Metabolism guarantees that dividing cells always have access to these critical genetic building blocks.
The flux of metabolites into this specialized pathway is tightly controlled and gated by the cytosolic NADP+/NADPH ratio. The primary control enzyme, Glucose-6-phosphate dehydrogenase (G6PD), is incredibly sensitive to this ratio. If a cell has ample reducing power, high NADPH levels strongly inhibit G6PD, forcing glucose-6-phosphate to enter standard glycolysis instead. However, if reductive power drops, the accumulating NADP+ instantly activates the enzyme, actively shunting carbon into the Pentose Phosphate Pathway to replenish the cellular antioxidant shields.
This specific slide highlights a critical clinical correlation for medical students: G6PD deficiency. As the most common human enzyme defect, a lack of G6PD robs red blood cells of their ability to produce NADPH. Without this critical reductive power, erythrocytes cannot neutralize oxidative stress from certain drugs, infections, or dietary triggers, leading to severe hemolytic anemia. Understanding how this specific pathway integrates with overall metabolic regulation is totally essential for treating patients with diverse biochemical vulnerabilities.
Slide 12: The Fed State and the Regulation of Carbohydrate Metabolism
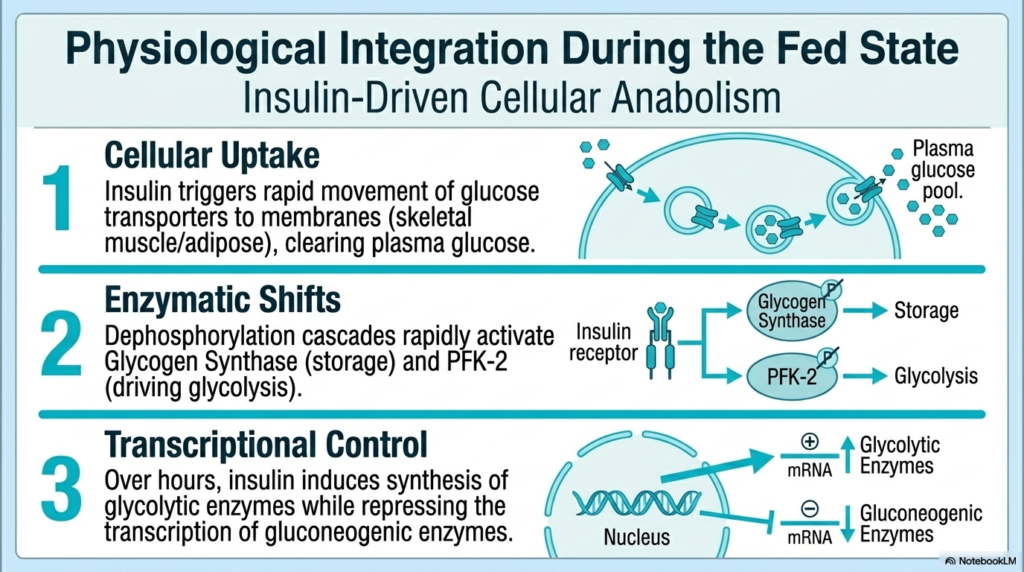
To fully grasp human metabolism, one must examine the physiological integration of the fed state, in which insulin drives overall cellular anabolism. The Regulation of Carbohydrate Metabolism begins immediately after a meal, as insulin acts on multiple distinct physiological levels to clear plasma glucose. First, it triggers rapid cellular uptake. Insulin binds to cell receptors on skeletal muscle and adipose tissue, causing specialized glucose transporters to rapidly fuse with the cell membrane. This instantly sucks massive amounts of glucose out of the blood plasma, actively lowering the postprandial glucose spike. This rapid Regulation of Carbohydrate Metabolism effectively prevents the toxic effects of vascular hyperglycemia.
Simultaneously, insulin drives massive enzymatic shifts within the cell cytosol. Through potent dephosphorylation cascades, insulin strongly activates key storage enzymes such as Glycogen Synthase, rapidly packing incoming glucose into dense glycogen granules. Concurrently, it dephosphorylates and activates the PFK-2 enzyme, ensuring the massive production of Fructose 2,6-bisphosphate to forcefully drive glycolysis. The Regulation of Carbohydrate Metabolism uses insulin not just as a cellular door-opener but as a master cytosolic conductor, ensuring that once glucose enters the cell, it is efficiently processed.
Finally, insulin exerts profound, long-term transcriptional control over the cellular nucleus. While allosteric and covalent modifications occur within seconds to minutes, insulin fundamentally alters the cell’s gene expression over hours to days. It heavily upregulates the transcription of messenger RNA for key glycolytic and lipogenic enzymes, physically building more metabolic machinery. At the exact same time, it fiercely represses the transcription of all gluconeogenic enzymes. This perfectly ensures the cell completely retools its genetic landscape to become a massive energy-storing powerhouse.
This highly structured, multi-tiered approach beautifully demonstrates the sheer power of endocrinology. For future clinicians, this slide perfectly encapsulates the devastating systemic failure that occurs in Type 2 Diabetes. When cells become resistant to this insulin signaling pathway, transporters fail to move, enzymes fail to dephosphorylate, and genes fail to transcribe properly. The resulting metabolic chaos underscores why mastering these highly integrated pathways is the bedrock of modern internal medicine.
Slide 13: Systemic Catabolism in the Regulation of Carbohydrate Metabolism
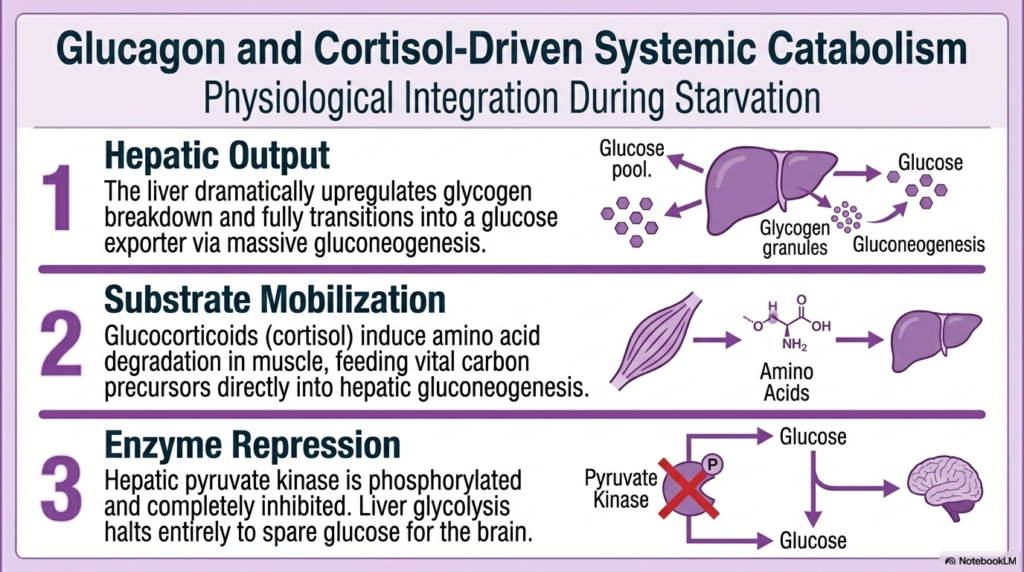
When energy abundance shifts to starvation, the body aggressively orchestrates systemic catabolism driven by glucagon and cortisol. The Regulation of Carbohydrate Metabolism during these fasting periods is fundamentally about survival, specifically prioritizing the preservation of central brain function. The first stage involves massive hepatic output. Glucagon initiates a total reversal of the liver’s metabolic profile, dramatically upregulating glycogen breakdown. When glycogen stores run low, the liver fully transitions into a specialized glucose exporter by driving massive gluconeogenesis. This precise Regulation of Carbohydrate Metabolism ensures the constant, steady release of free glucose into the systemic blood pool.
To sustain this massive hepatic glucose factory, the body requires raw materials, thereby initiating systemic substrate mobilization. This is where glucocorticoids, like cortisol, become critical. Cortisol acts systemically to induce severe amino acid degradation within skeletal muscle. This carefully controlled muscle wasting breaks down structural proteins to feed vital carbon precursors directly into the liver’s gluconeogenic pathway. The Regulation of Carbohydrate Metabolism relies heavily on this brutal but necessary physiological sacrifice to generate the carbon skeletons required to keep the nervous system fueled.
Finally, to maximize absolute efficiency, the liver utilizes strict enzyme repression. As previously discussed, hepatic pyruvate kinase is heavily phosphorylated and completely inhibited by glucagon signaling. This violent shutdown ensures that liver glycolysis halts entirely. The liver absolutely refuses to consume any of the precious glucose it is currently sacrificing muscle tissue to explicitly create. This final, failsafe mechanism safely guarantees that every single molecule of newly synthesized glucose is violently ejected into the bloodstream.
Students must realize that this precise physiological profile is essentially a state of strictly managed physiological stress. It perfectly mimics the metabolic profile of a patient suffering from severe trauma or untreated Type 1 Diabetes, where the lack of insulin artificially mimics a state of profound starvation. The excessive cortisol-driven muscle wasting and uncontrolled hepatic glucose output perfectly explain the severe weight loss and systemic hyperglycemia seen in such dire clinical scenarios.
Slide 14: Comprehensive Mapping of the Regulation of Carbohydrate Metabolism
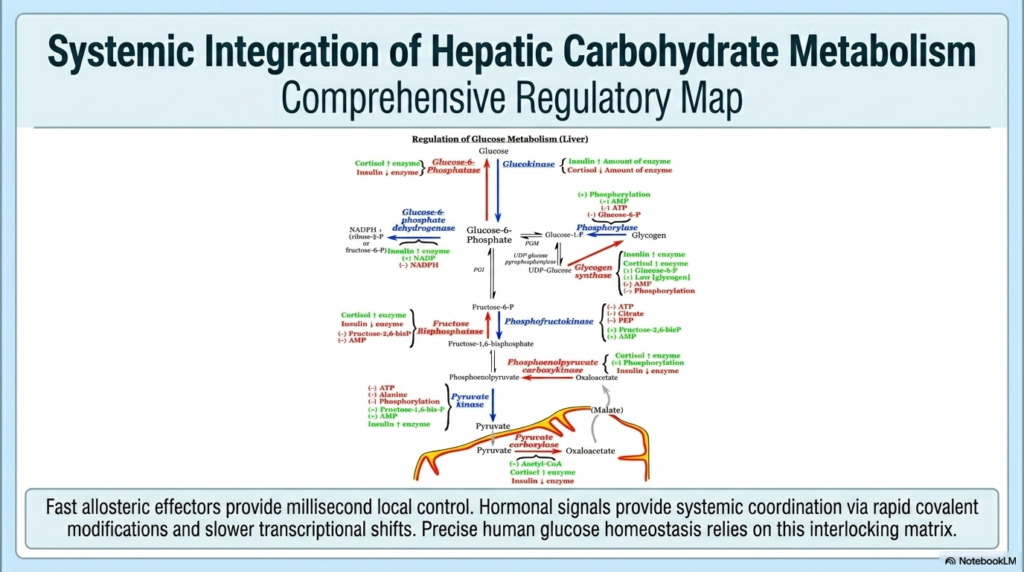
The final conceptual masterpiece is the systemic integration of hepatic carbohydrate metabolism, perfectly represented by a comprehensive regulatory map. The Regulation of Carbohydrate Metabolism is not a simple series of isolated biochemical events; it is a vastly interconnected, multi-layered regulatory matrix. At the very base level, fast allosteric effectors like ATP, AMP, and Fructose 2,6-bisphosphate provide instantaneous, millisecond control over local enzymatic flux. These molecules act as immediate cellular reflex arcs, seamlessly adjusting metabolic speed based on localized energetic demand. This intrinsic Regulation of Carbohydrate Metabolism is the foundational bedrock of all cellular survival.
Layered directly atop this allosteric foundation are the immensely powerful hormonal signals. Molecules like insulin, glucagon, and cortisol provide coordinated systemic signaling across diverse tissue types. They execute their central authority via rapid covalent modifications, such as phosphorylation, which forcefully override local allosteric controls when the entire organism faces an energetic crisis. Furthermore, these overarching hormones orchestrate slower, long-lasting transcriptional shifts within the cellular nucleus. The true Regulation of Carbohydrate Metabolism relies entirely on the flawless synchronization of these rapid reflexes, medium-term overrides, and long-term genetic adaptations.
Precise human glucose homeostasis ultimately relies on this massive interlocking matrix. A change in a single node quickly cascades through the entire network, gracefully shifting the human organism from a post-meal storage state to overnight fasting, and back again. The exquisite beauty of this system lies in its extreme redundancy, its physical precision, and its sheer biochemical complexity. It is an evolutionary marvel perfectly designed to buffer the body against wildly fluctuating environments.
For university and medical school students, this beautifully comprehensive map serves as the ultimate metabolic cipher. Every disease of energy metabolism, every pharmacological intervention in diabetes, and every physiological adaptation to extreme exercise can be explicitly traced back to a specific disruption or enhancement within this very matrix. By internalizing this overarching map, future medical professionals successfully transition from memorizing isolated pathways to truly understanding the magnificent logic of human physiology.
Please read our Content Disclaimer Statement.
Check out our social media channels: