14. Isoprenoids: Metabolism, Biosynthesis, and Pathways
When you look closely at the fundamental chemistry of life, you quickly realize that nature is the ultimate architect, utilizing surprisingly simple building blocks to construct incredibly complex and dynamic structures. This blog post and accompanying slide deck serve as a comprehensive visual and intellectual journey into the architecture of isoprenoids. Our purpose is to rigorously analyze their modular assembly, complex metabolic pathways, and their profound biological functions across different species. For medical and college students embarking on this biochemical journey, mastering these concepts is not just an exercise in memorization; it is the absolute key to unlocking the secrets of cellular energy, human disease, and the future of modern pharmacology.
Slide 1: The Foundation and Architecture of Isoprenoids
Welcome to the foundational blueprint of biochemistry. This initial slide introduces the staggering scale and molecular elegance of isoprenoids. When we examine the fundamental chemistry of living systems, we see that nature acts as a master builder, relying on simple, repeating units to generate massive biological complexity. This slide deck provides a deep, comprehensive visual analysis of the biochemical architecture of isoprenoids, exploring their modular assembly, metabolic synthesis, and critical biological functions. For college and medical students diving into this field, understanding these molecules is absolutely essential; they are the metabolic key to understanding cellular life, cardiovascular disease, and the design of modern therapeutics.
Isoprenoids represent by far the largest and most structurally diverse family of natural products found in living organisms. Despite their incredible variety—ranging from the volatile, fragrant scents of pine trees to the massive, rigid cholesterol molecules that stabilize our cell membranes—they all share a remarkably elegant and singular origin. Every single one of these complex molecules is constructed from a basic five-carbon (C5) precursor unit.
This modular assembly process is a central theme in our biological studies, demonstrating how a singular structural motif can be elongated, folded, cyclized, and chemically modified to serve a near-infinite array of biological needs. By mastering how cells manipulate isoprenoids, students can appreciate the precise enzymatic control required to sustain life.
The primary purpose of this presentation is to meticulously map out this architecture.
We will traverse the core biochemical pathways that generate these C5 units, specifically analyzing the vastly different evolutionary routes taken by mammals, plants, and infectious pathogens. We will then examine the structural elongation processes that transform simple precursors into massive lipid polymers. Finally, we will connect these atomic mechanisms to real-world clinical applications, exploring how isoprenoid metabolism governs everything from hormonal signaling and cellular respiration to cancer progression. Let us dive into the chemical logic of life.
Slide 2: Divergent Fates of Activated Acetic Acid and Isoprenoids
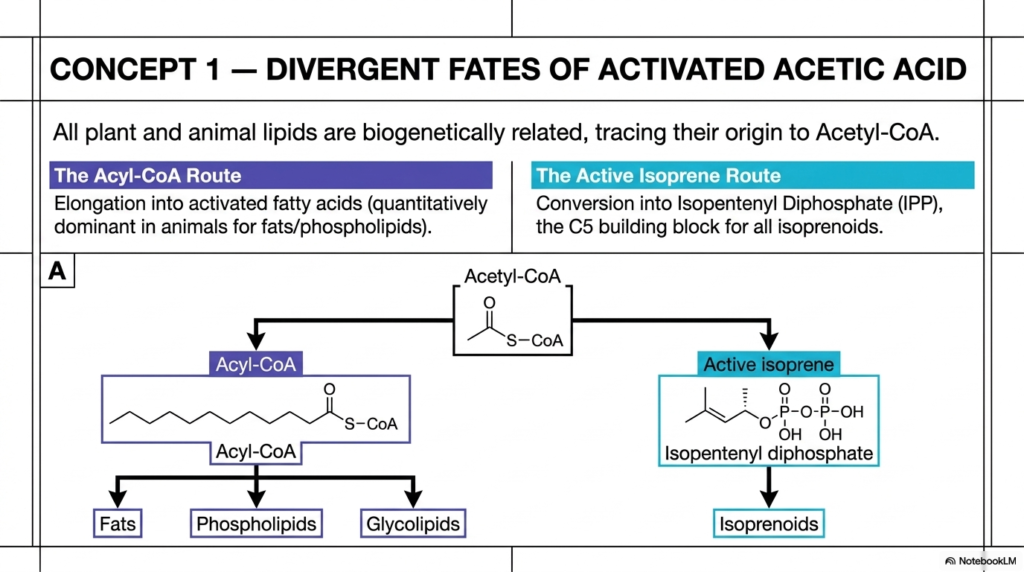
To truly comprehend the biochemical origins of isoprenoids, we must first examine a fundamental metabolic crossroads within the cell: the fate of activated acetic acid, universally known as Acetyl-CoA. Acetyl-CoA is arguably one of the most critical and central hub molecules in all of biology. Derived from the catabolic breakdown of the carbohydrates, fats, and proteins in our diet, it serves as the ultimate starting material for synthesizing a massive variety of structural and energetic lipids.
However, as this slide brilliantly illustrates, Acetyl-CoA has two distinct biochemical fates that depend entirely on the enzymatic machinery it encounters. One path leads to the flexible, linear chains of standard dietary fats, while the other leads to the incredible structural diversity of isoprenoids.
The first major fate is the Acyl-CoA Route.
In this highly active pathway, Acetyl-CoA units are sequentially strung together to form long, straight-chain activated fatty acids. This particular pathway is quantitatively dominant in human and animal systems for producing energy-storing fats, as well as the phospholipids and glycolipids that physically make up the bulk of all cellular membranes. It relies on a relatively straightforward, repeating cycle of carbon elongation. In stark chemical contrast stands the Active Isoprene Route. Instead of forming long linear chains, Acetyl-CoA is shuttled through a significantly more complex series of energetic reactions to form Isopentenyl Diphosphate (IPP). This specific molecule is the essential, universal C5 building block for all downstream isoprenoids.
Why is this enzymatic branching point so profoundly important for future clinicians and scientists? The metabolic partitioning of Acetyl-CoA dictates overall cellular health and underlies numerous metabolic disorders. When a cell commits its Acetyl-CoA to the Active Isoprene Route, it is investing its carbon and energy resources in synthesizing essential isoprenoids such as membrane cholesterol, steroid hormones, and the vital electron carriers required for mitochondrial respiration. Recognizing this divergence is the critical first step in understanding how our cells balance the basic need for bulk membrane materials against the complex need for highly specialized regulatory lipids.
Slide 3: Compartmentation of Biosynthesis for Isoprenoids
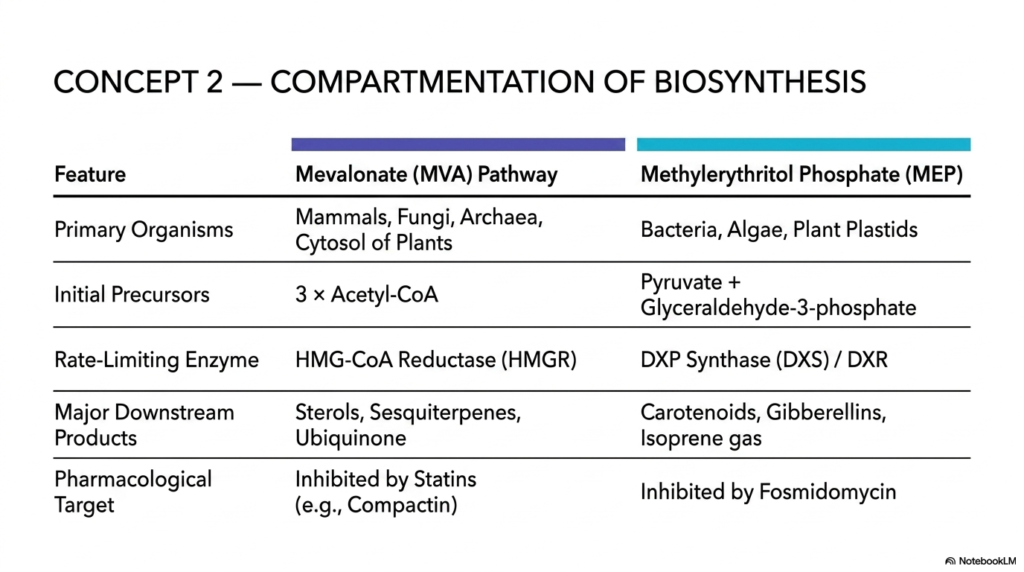
One of the most profound and clinically relevant biological concepts regarding isoprenoids is how their biosynthesis is strictly compartmentalized across different domains of life, and even within the individual compartments of a single cell. This slide introduces Concept 2: the distinct evolutionary division between the Mevalonate (MVA) Pathway and the Methylerythritol Phosphate (MEP) Pathway. For decades, the scientific community believed there was only one possible way to build these molecules. The discovery of a completely alternative, non-mevalonate route fundamentally revolutionized our modern understanding of plant biochemistry, evolutionary biology, and targeted antimicrobial drug design.
The Mevalonate Pathway represents the classic, textbook route. It operates primarily in the cytosol of mammals, fungi, and archaea, and extensively in the cytoplasm of plants. It utilizes three molecules of Acetyl-CoA to eventually form the required C5 building blocks. The ultimate gatekeeper of this pathway is the highly regulated, rate-limiting enzyme HMG-CoA Reductase (HMGR), which directs the massive downstream synthesis of sterols (like human cholesterol), specific sesquiterpenes, and ubiquinone.
On the other side of this biological divide is the MEP pathway. Found exclusively in bacteria, algae, and the specialized plastids (such as chloroplasts) of plants, this route entirely bypasses Acetyl-CoA. Instead, it utilizes pyruvate and glyceraldehyde-3-phosphate, relying on entirely distinct enzymes like DXP Synthase (DXS) and DXR to produce massive quantities of major downstream products such as plant carotenoids and volatile isoprene gas.
The clinical implications of this distinct compartmentalization of isoprenoids cannot be overstated in modern medicine. Because humans rely exclusively on the MVA pathway to synthesize our vital lipids, our cells completely lack the enzymes of the MEP pathway. This presents a massive, incredibly safe therapeutic window! Clinicians can use statins to directly inhibit HMGR and safely lower human cholesterol levels. Conversely, the MEP pathway is a prime biochemical target for potent antimicrobials such as Fosmidomycin. By biochemically attacking a route that is essential for a deadly pathogen’s survival but entirely absent in human biology, we can achieve high therapeutic efficacy with minimal toxic side effects.
Slide 4: The Mevalonate Cascade in the Synthesis of Isoprenoids
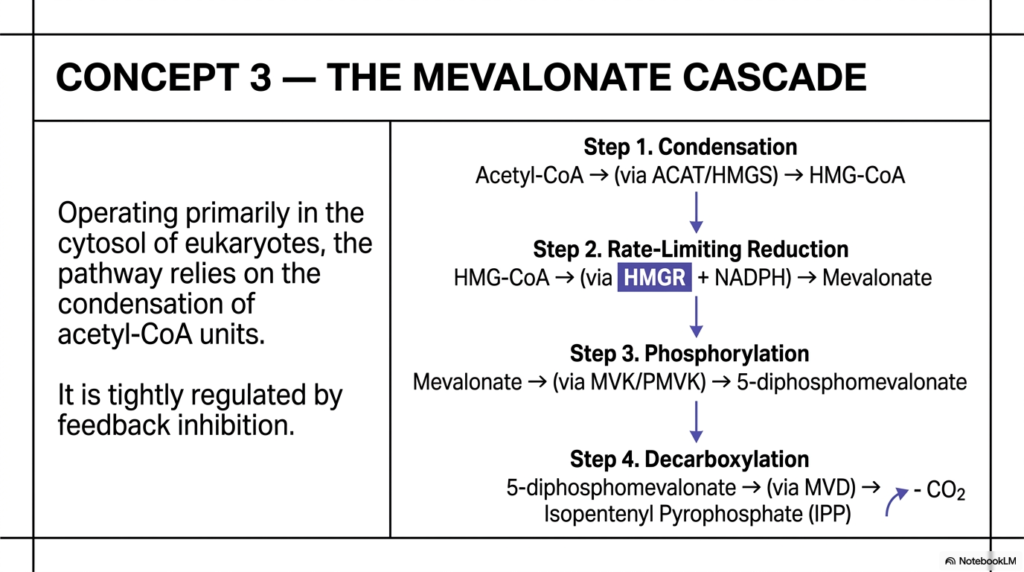
This detailed slide takes us deep into the busy cytosolic factories of eukaryotic cells to rigorously examine Concept 3: The Mevalonate Cascade. For medical students and future physicians, mastering this highly specific sequence of events is an absolute necessity, as it lies at the very core of cardiovascular pharmacology and global lipid metabolism. The synthesis of isoprenoids via this exact pathway is a true marvel of enzymatic precision, relying on the tight, instantaneous regulation of carbon condensation and sequential phosphorylation to successfully build the active C5 precursor, Isopentenyl Pyrophosphate (IPP).
The molecular cascade begins with a crucial condensation phase, in which simple two-carbon Acetyl-CoA units are carefully stitched together by specific enzymes (ACAT and HMGS) to form a branched six-carbon intermediate, universally known as HMG-CoA. This step leads directly into Step 2, the absolutely critical bottleneck and regulatory juncture of the entire biochemical pathway: the rate-limiting reduction of HMG-CoA directly into Mevalonate. This highly energetic step is catalyzed by the legendary cellular enzyme HMG-CoA Reductase (HMGR) and requires the immense reducing power of NADPH. Because this single step dictates the total downstream production of all cellular isoprenoids, HMGR is placed under intense biological surveillance, subject to profound genetic feedback inhibition by its own downstream lipid products, most notably cholesterol.
Once the intermediate mevalonate is formed, the cell must chemically ‘activate’ it so it can participate in subsequent condensation reactions. Through Step 3, mevalonate undergoes sequential, energy-intensive phosphorylation by specific kinases (MVK and PMVK), ultimately yielding 5-diphosphomevalonate. This strategic addition of bulky, highly energized phosphate groups perfectly primes the molecule for the final enzymatic act. In Step 4, a highly specialized enzyme called mevalonate diphosphate decarboxylase (MVD) forcibly removes a carbon dioxide molecule in a critical decarboxylation event. This final, irreversible structural adjustment yields the required IPP. Understanding this step-by-step cascade reveals exactly how much energy cells must invest to create the highly reactive building blocks required to sustain the complex architecture of human isoprenoids.
Slide 5: The Non-Mevalonate Cascade Producing Isoprenoids

As we shift our biochemical focus to Concept 4, we must deeply explore the evolutionary marvel known as the Non-Mevalonate, or MEP pathway. While human and fungal biochemistry rely exclusively on the classic MVA pathway to build their vital isoprenoids, many of the world’s most dangerous pathogens, along with all botanical life, utilize this entirely distinct biochemical cascade. For college students studying microbiology, agriculture, and infectious disease, thoroughly understanding this specific pathway is just as critical as understanding human metabolism. The enzymes in this sequence hold the elusive keys to developing next-generation agricultural herbicides, potent antibiotics, and novel antimalarial drugs.
The MEP cascade operates through a fundamentally different and fascinating chemical logic. In Step 1 (the Initiation phase), the cellular machinery completely ignores Acetyl-CoA. Instead, the specialized enzyme DXS merges pyruvate and glyceraldehyde-3-phosphate (G3P)—both of which are core, highly abundant metabolites of normal glycolysis—to successfully form 1-deoxy-D-xylulose 5-phosphate (DXP). Step 2 features the absolute critical reduction of DXP via the heavily studied enzyme DXR to ultimately yield methylerythritol phosphate (MEP).
Just as the famous HMGR controls the mammalian lipid pathway, DXR acts as the heavily scrutinized, rate-limiting checkpoint in this bacterial and plastidial route. Following this reduction, the biochemical cascade moves into Step 3, featuring an intricate, multi-enzyme cyclization process managed precisely by the IspD/E/F enzyme complex, which transforms the linear MEP directly into a ringed 2,4-cyclodiphosphate. This complex pathway finally culminates in Step 4, a brilliant terminal reduction carefully catalyzed by the enzymes IspG and IspH.
This final chemical manipulation breaks open the cyclized intermediate, producing a crucial, highly reactive mixture of the universal C5 building blocks: Isopentenyl Diphosphate (IPP) and its indispensable isomer, Dimethylallyl Diphosphate (DMAPP). The overarching importance of these pathogen-specific isoprenoids absolutely cannot be overstated in a clinical setting. Bacteria desperately rely on these specific lipids to build their essential, protective cell walls and their respiratory quinones. Because this entire sequence of highly specialized enzymes is completely absent in humans, the MEP cascade represents an incredibly lucrative, safe frontier for drug discovery, allowing us to selectively starve pathogens of their essential isoprenoids.
Slide 6: Modular Elongation and the Assembly of Isoprenoids
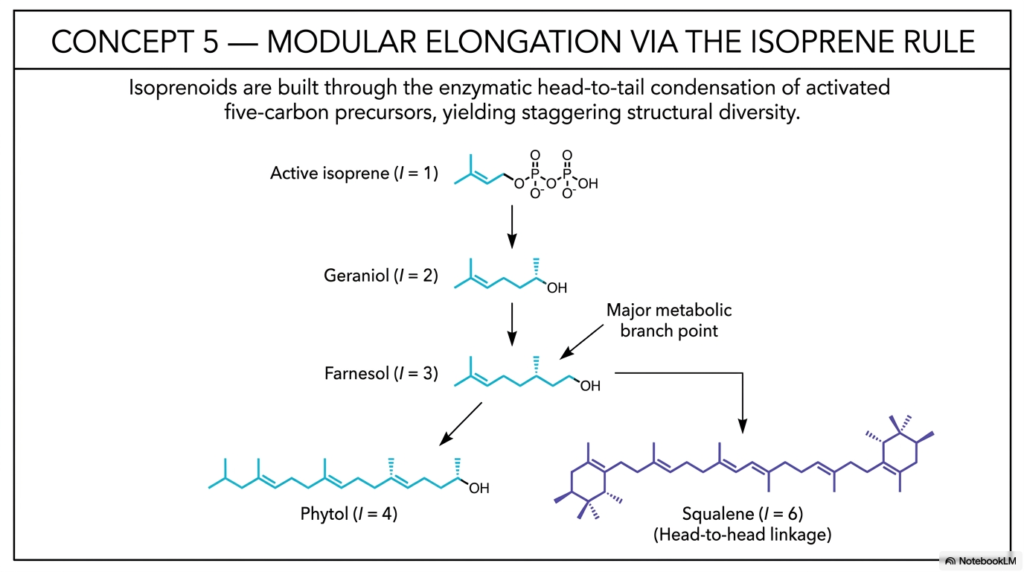
With our highly reactive C5 building blocks firmly in hand, Concept 5 expertly illustrates the true architectural genius of cellular biochemistry: modular elongation. This critical slide formally introduces the “Isoprene Rule,” a unifying, universal chemical principle that strictly dictates how nature builds almost unimaginable structural diversity from a single, simple, repeating motif. Just as complex, functional proteins are built from a highly finite alphabet of amino acids, the vast, expansive universe of complex isoprenoids is constructed entirely through the careful enzymatic head-to-tail condensation of these activated five-carbon precursors.
The complex assembly process begins rapidly with active isoprene (where the basic isoprene unit count, I = 1). Through the tireless work of highly specialized cellular enzymes called prenyltransferases, these individual C5 units are violently snapped together. When the cellular machinery combines two distinct C5 units, you generate the heavily studied 10-carbon molecule known as Geraniol (I = 2).
Adding yet another C5 unit systematically yields Farnesol (I = 3), a crucial 15-carbon intermediate. Farnesyl pyrophosphate is arguably one of the most vitally important metabolic branch points in all of eukaryotic biology. From this single, highly trafficked 15-carbon junction, the cell can deliberately continue adding C5 units to form the elongated 20-carbon Phytol (I = 4), which is absolutely essential for synthesizing vital human vitamins and plant chlorophyll.
Alternatively, the cell can dramatically and rapidly alter its synthetic strategy at this exact junction. Instead of continuing the standard sequential head-to-tail additions, the cellular enzymes can take two distinct 15-carbon Farnesyl molecules and forcefully fuse them together in a unique, highly specialized head-to-head linkage to form Squalene (I = 6). This massive, symmetrical 30-carbon molecule is the absolute biological prerequisite for human cholesterol and all subsequent steroid hormones. By clearly visualizing these exact pathways, college and medical students can truly appreciate how simple, regulated variations in enzymatic linkage allow our cells to generate a highly diverse class of life-sustaining isoprenoids that dictate human health.
Slide 7: Monoterpenes as Volatile Signals among Isoprenoids
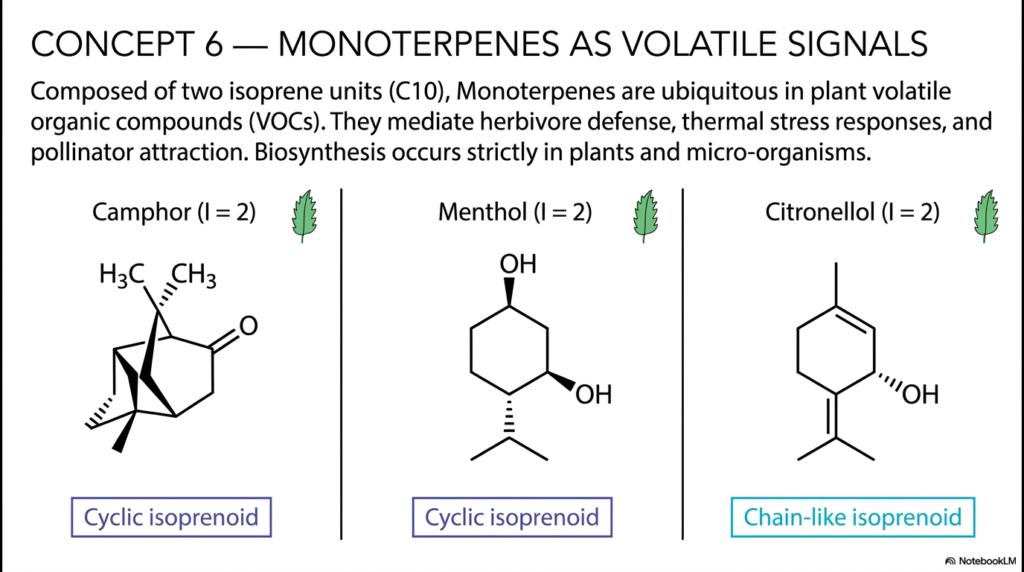
Moving steadily up the biochemical ladder of molecular complexity, Concept 6 dives deeply into the fascinating botanical world of Monoterpenes, the smallest and most aromatic class of elongated isoprenoids. Composed of exactly two linked isoprene units (yielding a highly specific 10-carbon, or C10, skeleton), these tiny molecules are globally recognized for their intensely aromatic and highly potent biological activities. For students rigorously studying botany, environmental ecology, or pharmacognosy, monoterpenes represent the very literal chemical language of the plant kingdom. While their unique biosynthesis occurs exclusively in plants and certain microorganisms, their ecological impact resonates throughout the biosphere, profoundly influencing global agriculture, perfumery, and modern medicine.
As highly volatile organic compounds (VOCs), monoterpenes readily evaporate into ambient air at normal temperatures. This specific physical property is perfectly and evolutionarily suited to their primary biological functions. Plants intelligently utilize these specific, lightweight isoprenoids to mediate absolutely critical ecological interactions in their environment.
When a vulnerable plant is aggressively attacked by hungry herbivores, it can rapidly synthesize and release a massive defensive cloud of toxic or severely foul-tasting monoterpenes as an immediate chemical deterrent. Conversely, the incredibly sweet, floral scents of specific monoterpenes are utilized to attract highly specific pollinators from miles away. Furthermore, these unique molecules actively help plants survive severe thermal stress by safely stabilizing their delicate cell membranes during intense summer heat waves.
The underlying structural diversity within this single, small class of isoprenoids is genuinely astounding. The slide vividly contrasts the unique cyclic monoterpenes, such as the famous cooling compound Menthol and the rigid, strongly aromatic Camphor, directly against simple chain-like structures like Citronellol. Even highly minor chemical changes in the stereochemistry or the internal cyclization of that basic 10-carbon backbone dramatically alter how these molecules interact with human olfactory receptors and pharmacological targets. Today, we globally harvest these exact botanical compounds for massive use in essential oils, powerful antimicrobial topical treatments, and highly effective mosquito repellents, proving the massive real-world value of isoprenoids.
Slide 8: Sesquiterpenes as Hormones and Therapeutics in Isoprenoids
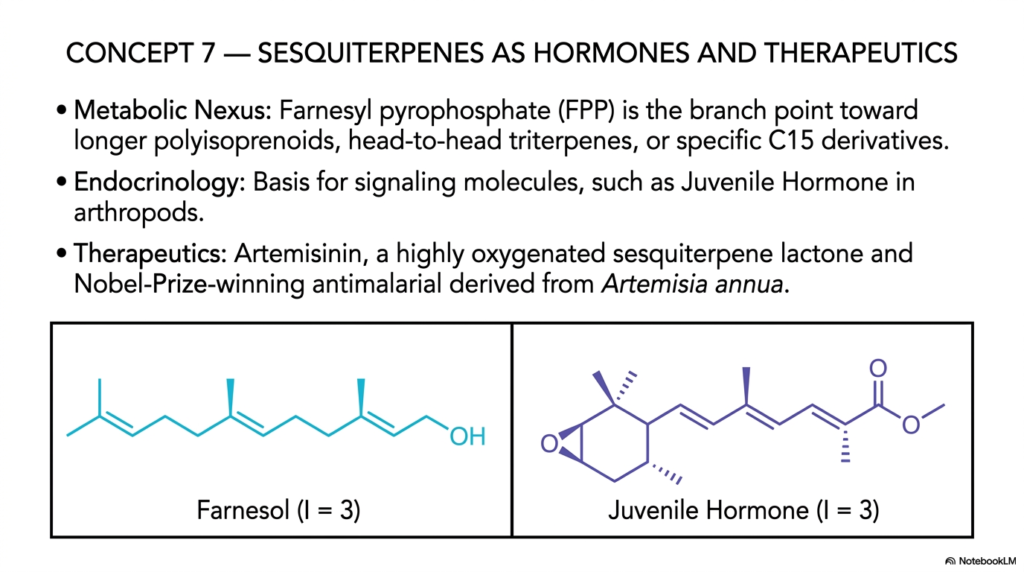
Concept 7 systematically introduces us to the incredibly dynamic and highly versatile world of 15-carbon (C15) sesquiterpenes. Meticulously built from exactly three linked C5 units, this intermediate class of isoprenoids represents a profound and highly significant evolutionary leap in biological utility. If the smaller monoterpenes are considered the volatile perfumes of the dense forest, sesquiterpenes are the highly complex biological regulators and potent medicinal agents of the biochemical world. For medical students and structural biochemists, the careful synthesis of these C15 molecules highlights a major, critical metabolic nexus that flawlessly bridges simple lipid elongation with highly complex animal endocrinology and life-saving human pharmacology.
At the absolute, undeniable center of this metabolic nexus is the molecule Farnesyl pyrophosphate (FPP). As the slide so clearly illustrates with the molecule Farnesol (I=3), this specific 15-carbon intermediate is the grand, bustling intersection of all cellular isoprenoid metabolism. It serves as the direct, primary branch point toward the much longer polyisoprenoids, the massive head-to-head triterpenes like squalene, and an incredibly vast array of specific C15 regulatory derivatives. In the fascinating realm of animal endocrinology, these specific C15 isoprenoids act as exceptionally powerful, highly targeted signaling molecules. A classic, textbook example is Juvenile Hormone in arthropods, a vital sesquiterpene that meticulously and flawlessly dictates the exact timing of insect molting and physical metamorphosis.
However, the absolute true clinical marvel of studying sesquiterpenes lies entirely in their modern application as profound human therapeutics. The slide heavily highlights the molecule Artemisinin, a highly oxygenated, deeply complex sesquiterpene lactone naturally and beautifully derived from the Artemisia annua plant. This single, remarkable molecule is a Nobel-Prize-winning antimalarial compound that has literally saved millions of human lives across the globe. Its highly unique endoperoxide bridge—a remarkably rare structural chemical feature in naturally occurring isoprenoids—reacts violently and deliberately with the heavy iron located inside the malaria parasite, generating massive oxidative stress that selectively and totally destroys the deadly pathogen.
Slide 9: Diterpenes as Essential Lipid Anchors and Vitamin Isoprenoids
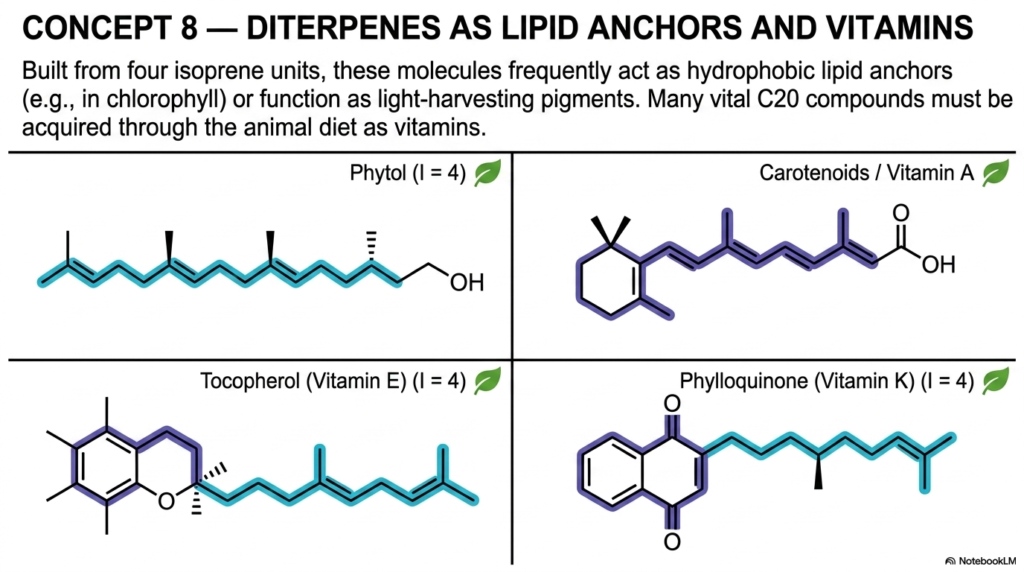
As we reach the critical 20-carbon threshold (I=4), we fully enter the vital biological domain of Diterpenes, a massive and highly significant structural class of isoprenoids, as detailed in Concept 8. These large C20 molecules elegantly demonstrate how biochemical structure strictly dictates cellular function. Due to their incredibly long, highly hydrophobic, and greasy hydrocarbon chains, diterpenes frequently and heavily serve as deep-set lipid anchors, plunging deeply into cellular membranes to hold absolutely critical, life-sustaining protein complexes firmly in place.
For students heavily studying human nutrition and medical physiology, diterpenes are of paramount importance because several of them cannot be synthesized by human cells and must be acquired entirely through our daily diet as vital, life-sustaining vitamins.
The detailed slide beautifully showcases Phytol, a classic, perfectly linear 20-carbon chain. In the vast plant kingdom, phytol acts as the highly essential hydrophobic tail that permanently anchors the vital light-harvesting pigment chlorophyll directly into the busy photosynthetic membranes of green chloroplasts.
Without these specific C20 isoprenoids acting as permanent biological tethers, the complex molecular machinery of global photosynthesis would literally and immediately fall apart. But the true biological utility of diterpenes extends far, far beyond mere structural anchoring in plants. The deeply, beautifully pigmented Carotenoids, evolutionarily derived from this exact same metabolic lineage, are fundamentally and absolutely required for healthy human vision. When we consume these botanical pigments, our highly adapted bodies chemically cleave them into Vitamin A, a powerful diterpene derivative essential for maintaining our retinas.
Furthermore, this highly specific class of isoprenoids includes incredibly potent, life-saving biological protective agents. Tocopherol, widely known in clinical practice as Vitamin E, features a long diterpene tail permanently attached to a highly reactive chemical ring, allowing it to embed deeply in our cell membranes and rapidly intercept damaging free radicals before they can destroy our lipids. Similarly, the molecule Phylloquinone (Vitamin K) utilizes a massive C20 tail to deeply embed itself in human tissues, where it is absolutely and undeniably required for healthy blood coagulation and maintaining strong bone metabolism, proving that our survival depends on botanical isoprenoids.
Slide 10: Triterpenes and the Membrane Integrity of Isoprenoids
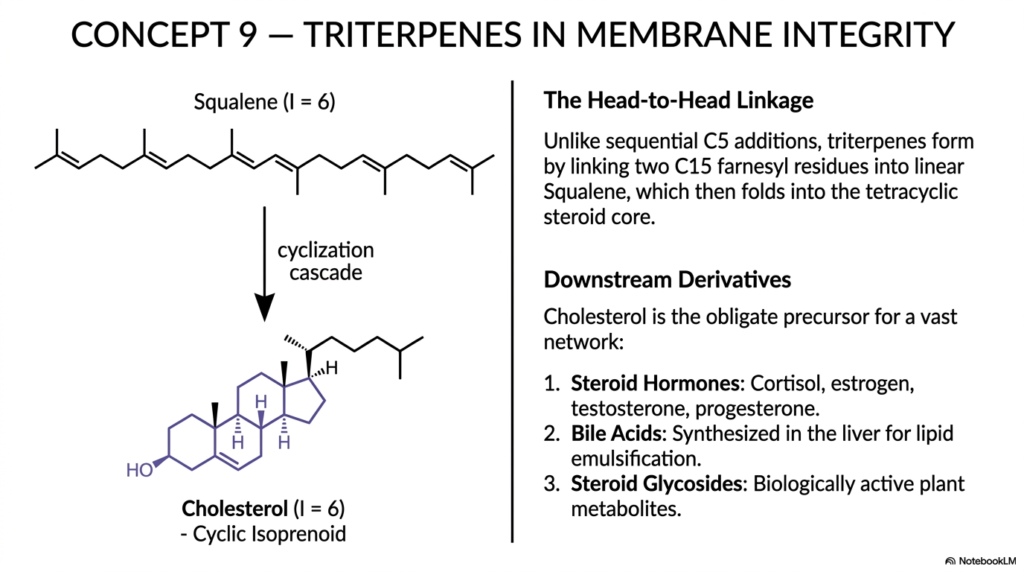
Concept 9 rapidly brings us to one of the most clinically relevant and medically significant slides in the entire educational deck: the complex formation of Triterpenes (C30) and their vital role in maintaining membrane integrity. Carefully formed from exactly six isoprene units (I=6), triterpenes represent a highly dramatic, fascinating shift in exactly how the eukaryotic cell chooses to build its massive isoprenoids. Instead of continuously and predictably adding simple C5 blocks sequentially to the end of a growing lipid chain, the cell intelligently takes two massive 15-carbon Farnesyl residues and violently smashes them together in a highly unique “head-to-head” enzymatic linkage. This remarkable, highly energetic enzymatic feat perfectly produces Squalene, a long, symmetrical, deeply linear 30-carbon molecule.
Through an absolutely dazzling, highly complex, multi-step cyclization cascade, this linear squalene folds entirely and perfectly upon itself, chemically zipping up tightly to form the highly rigid, deeply hydrophobic four-ringed tetracyclic core of mammalian Cholesterol. While it is often highly villainized in popular media strictly for its role in causing human cardiovascular disease, cholesterol is a cyclical isoprenoid that is absolutely, undeniably obligate for normal human survival.
By carefully interspersing itself directly among the highly flexible phospholipids of our cellular membranes, cholesterol acts as a vital structural buffer, preventing our delicate cells from becoming far too fluid in the heat or far too brittle and rigid in the cold. Without this specific class of isoprenoids, mammalian cells would physically and rapidly collapse.
Beyond vital membrane mechanics, cholesterol is the primary obligate precursor for an incredibly vast downstream human endocrine network. Our bodies meticulously modify this rigid triterpene core to rapidly synthesize highly potent Steroid Hormones, including human cortisol, estrogen, testosterone, and progesterone. These highly specific molecules tightly regulate nearly every facet of human physiology, from our daily stress responses to successful reproduction. Additionally, the human liver rapidly oxidizes excess cholesterol to create highly functional Bile Acids, which are heavily secreted into the gut to successfully emulsify our dietary fats, showcasing the profound power of isoprenoids.
Slide 11: Polyisoprenoids in the Cellular Respiration of Isoprenoids
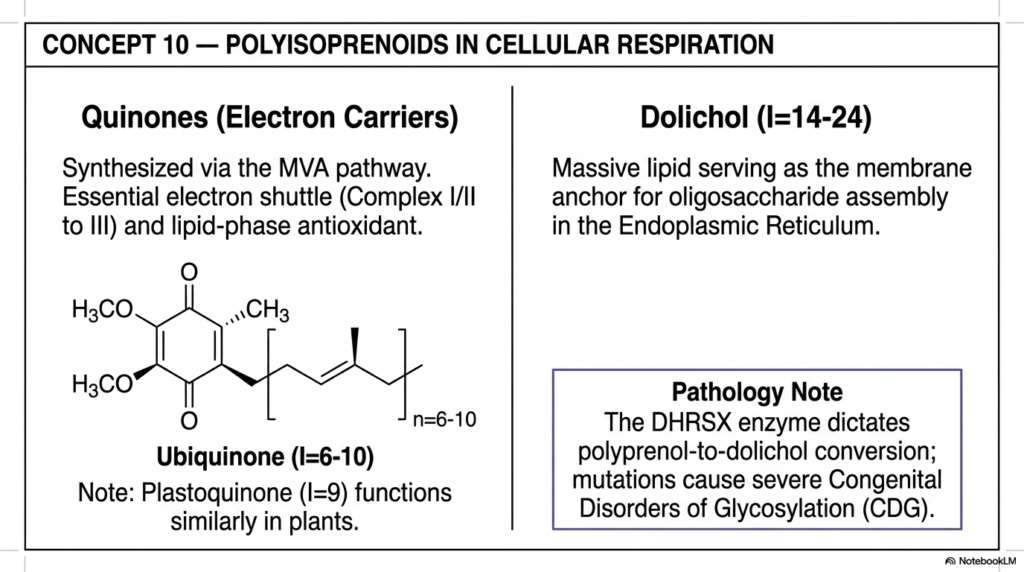
Moving massively beyond the standard C30 sterols, Concept 10 formally introduces the truly, unbelievably massive Polyisoprenoids. These are gigantic, extremely long molecules consisting of anywhere from 6 to well over 24 distinct isoprene units, and they flawlessly carry out vital biological functions essential for driving cellular energy production and complex protein folding. For advanced students deeply exploring bioenergetics and complex molecular genetics, thoroughly understanding these massive isoprenoids is highly crucial, as they form the literal, undeniable life-support energetic systems of both our human mitochondria and our busy endoplasmic reticulum.
The very first major functional class discussed here is the highly reactive Quinones, perfectly synthesized entirely via the human Mevalonate pathway. The absolute star metabolic player here is Ubiquinone, universally recognized in the modern medical field as the vital supplement Coenzyme Q10 (CoQ10). This fascinating molecule features a massive, highly hydrophobic isoprenoid tail (containing precisely 6 to 10 repeating C5 units depending heavily on the specific animal species) permanently attached to a highly reactive quinone head group.
Buried deeply and safely within the inner mitochondrial membrane, CoQ10 functions as an absolutely essential, highly mobile biological electron shuttle. It rapidly and constantly ferries highly energetic electrons between Complex I/II and Complex III of the human respiratory chain, directly and powerfully driving the massive daily production of cellular ATP.
The detailed slide also heavily highlights Dolichol, an absolute biochemical behemoth containing a staggering 14 to 24 linked isoprene units. Dolichol serves as a massive, highly reliable lipid anchor for complex oligosaccharide assembly directly in the Endoplasmic Reticulum, effectively functioning as the biological conveyor belt for critical N-linked protein glycosylation. Proteins absolutely must be properly and flawlessly glycosylated to fold and function correctly in the human body. The clinical pathology note on this slide is profoundly vital: dangerous mutations in the specific DHRSX enzyme, which carefully dictates the final conversion of polyprenol directly to dolichol, completely and tragically disrupt this process. This leads directly to devastating Congenital Disorders of Glycosylation (CDG), proving our reliance on huge isoprenoids.
Slide 12: Protein Lipidation and Signal Transduction via Isoprenoids
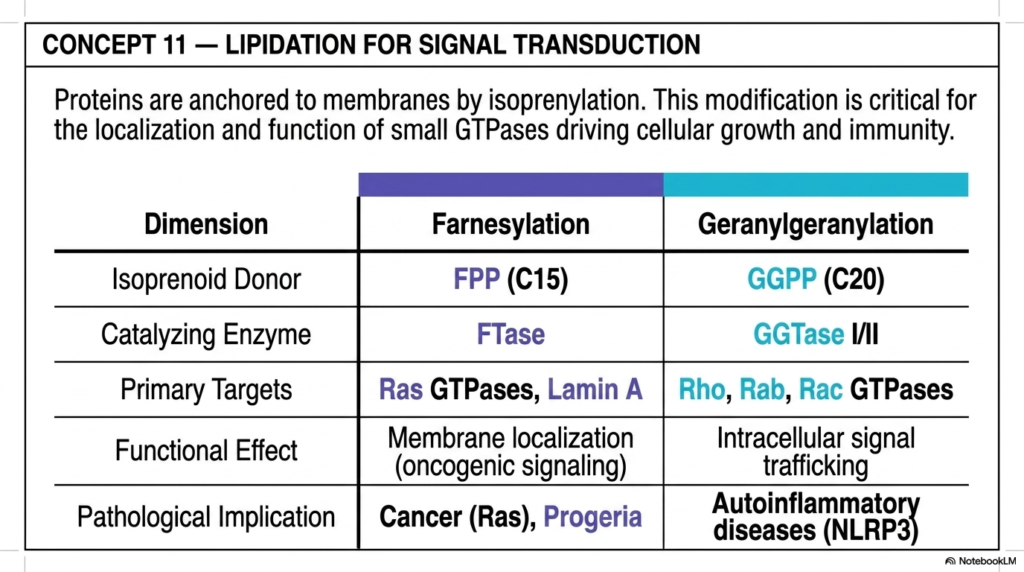
Concept 11 systematically introduces a completely different, highly targeted, and chemically fascinating biological function for these vital molecules: complex protein lipidation. Up to this point in the education, we have largely viewed functional isoprenoids as highly independent, free-floating lipids in biological membranes or as massive hormone precursors. However, this critical slide brilliantly reveals that human cells use highly specific 15- and 20-carbon isoprenoids as permanent chemical “staples” to anchor critical, otherwise water-soluble signaling proteins directly to internal cellular membranes. This specific, highly regulated process, clinically known as isoprenylation or prenylation, is an absolute, undeniable physiological requirement for the proper cellular localization of small GTPases.
The highly detailed table vividly and clearly contrasts the two distinct, critical dimensions of this vital post-translational protein modification. The process of Farnesylation strictly uses the highly abundant C15 Farnesyl pyrophosphate (FPP) as the primary lipid donor, and is catalyzed by the specialized cellular enzyme Farnesyltransferase (FTase). The primary, highly scrutinized biological targets for this highly specific lipidation are the famous Ras family of cellular GTPases and critical structural nuclear proteins like Lamin A.
When the Ras protein is properly and securely farnesylated, it embeds permanently in the plasma membrane, acting as a pivotal, tightly controlled biochemical switch for safe cell division. Conversely, Geranylgeranylation carefully uses the larger C20 GGPP donor to securely anchor a completely different suite of signaling molecules, notably the vital Rho, Rab, and Rac GTPases.
For dedicated medical students and aspiring medical oncologists, the severe pathological implications of these specific isoprenoids are genuinely and truly staggering. When these specific Ras genes genetically mutate and become highly and constantly hyperactive, they directly drive massive, entirely uncontrollable oncogenic signaling, rapidly leading to numerous deadly human cancers. Because the mutated Ras protein absolutely must be anchored by its highly greasy C15 isoprenoid tail to actually cause cancer, the prenylation pathway is a massive, highly researched target for modern chemotherapy. Furthermore, genetic defects in Lamin A farnesylation directly and tragically cause Progeria, a devastating, rapid-aging disease, proving how isoprenoids dictate human life.
Slide 13: Exploiting Pathway Distinctions for Therapeutics Using Isoprenoids

Concept 12 brilliantly and successfully bridges the complex educational gap between highly basic biochemistry and the modern pharmacy shelf. Having thoroughly and meticulously mapped the intricate synthesis pathways of these complex molecules, this slide clearly details how modern human medicine deliberately and carefully exploits evolutionary pathway distinctions to successfully create some of our most powerful, life-saving therapeutics. For practicing clinicians, the highly targeted pharmacological manipulation of cellular isoprenoids is definitely not a tiny niche sub-specialty; it is absolutely one of the most widely practiced medical interventions globally, used daily to safely prevent heart attacks and cure devastating parasitic infections.
The detailed top panel highlights vital therapeutics that safely target the human Mevalonate (MVA) Pathway. The absolute undisputed, massively prescribed heavyweights here are modern Statins. By effectively acting as highly competitive, direct inhibitors of the rate-limiting enzyme HMGR, statins dramatically and safely reduce the human liver’s daily production of downstream cholesterol. However, astute biochemistry students must recognize the associated metabolic cascade effects: statins also indirectly and substantially deplete the critical cellular pools of both FPP and GGPP. This severely alters Ras protein prenylation (offering potential, highly studied anti-cancer benefits) but also drastically reduces cellular CoQ10 levels, which directly causes the notoriously common side effect of statin-induced muscle pain. Another major MVA target is the enzyme Farnesyltransferase.
The lower educational panel shifts deeply toward powerful anti-infectives that safely target the pathogenic MEP pathway. As previously and firmly established, human beings entirely and completely lack this specific non-mevalonate route, making it an incredibly attractive, highly safe, high-specificity antibiotic drug target. The potent drug Fosmidomycin aggressively and permanently inhibits the vital bacterial enzyme DXR. Because the dangerous pathogen absolutely cannot produce its highly essential, life-sustaining isoprenoids, it rapidly perishes. Fosmidomycin brilliantly demonstrates highly potent, life-saving efficacy against both dangerous Gram-negative bacteria and Plasmodium falciparum, the incredibly deadly parasite wholly responsible for human Malaria, perfectly highlighting the clinical power of targeting isoprenoids.
Slide 14: Analytical Quantification and Measurement of Isoprenoids

Concept 13 rapidly transitions from clinical pharmacology to the highly complex realm of advanced analytical laboratory chemistry. To truly and deeply understand, accurately diagnose, and therapeutically and safely manipulate these highly complex metabolic lipid pathways, modern scientists absolutely must be able to measure them precisely in a lab setting. However, due to the staggering, massive structural diversity and wildly varying chemical properties of isoprenoids, this remains a tremendous, highly difficult analytical laboratory challenge. This highly technical slide carefully details the highly sophisticated instrumentation absolutely required to successfully quantify everything from highly volatile, lightweight plant scents to massively hydrophobic, heavy membrane sterols.
The absolute primary, highly reliable workhorse of modern clinical lipidomics is Targeted LC-MS/MS (Liquid Chromatography coupled heavily with tandem Mass Spectrometry). This highly sensitive, deeply complex analytical technique is widely used daily to accurately quantify both hydrophobic neutral lipids (such as the vital Coenzyme Q) and highly polar, highly water-soluble chemical intermediates (such as the reactive prenyl diphosphates FPP and GGPP). When carefully analyzing the critical cellular redox ratios of ubiquinone to ubiquinol, dedicated laboratory scientists must strictly employ rapid, immediate cold quenching and incredibly strict metal chelation. Because these highly specific isoprenoids are highly reactive, unstable electron carriers, they will spontaneously and rapidly oxidize upon basic air exposure, severely ruining the data.
For the much lighter, highly aromatic side of the massive molecular family, Gas Chromatography-Mass Spectrometry (GC-MS) is the ideal diagnostic tool. GC-MS is highly tailored for tiny, volatile isoprenoids, such as highly fragrant plant monoterpenes and other lightweight volatile organic compounds (VOCs). Finally, the slide highly highlights the brilliant technique of Stable Isotope Tracing. By safely feeding live cells heavy-labeled precursors like 13C-acetate (to probe the cytosolic MVA activity) or 13C-glycerol (to clearly probe plastidial MEP flux), dedicated researchers can mathematically and perfectly track exactly how carbon flows. This reveals the highly hidden metabolic engines generating our vital isoprenoids.
Slide 15: Mapping C5 Assembly to the Biological Function of Isoprenoids
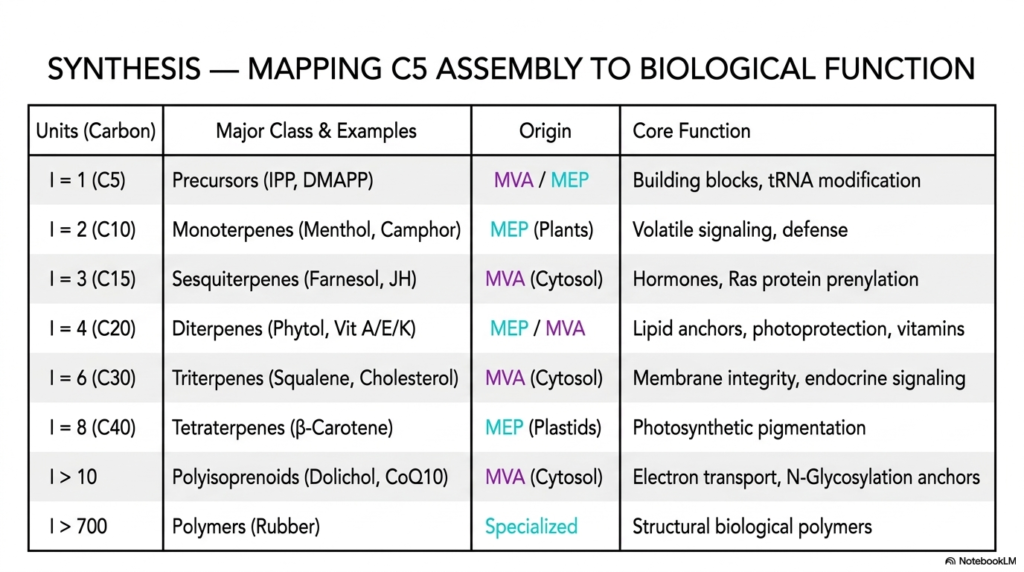
The final, highly comprehensive slide in this vast educational deck serves as the absolute ultimate biochemical synthesis, bringing together all the massive structural, deeply metabolic, and highly clinical data we have rigorously discussed into one perfectly unified, highly readable map. This massive synthesis table meticulously and flawlessly maps the basic C5 modular assembly directly and clearly to complex biological cellular function, successfully providing medical and college students with the absolute ultimate master key for understanding the vast, deeply interconnected universe of complex isoprenoids. By carefully tracking the exact number of isoprene units (I), the total carbon count, and their specific cellular biological origins, we can deeply appreciate exactly how nature scales molecular complexity.
At the absolute foundational base of the detailed table are the highly universal C5 Precursors (IPP and DMAPP). Whether generated actively by the human cytosolic MVA pathway or the deeply botanical plastidial MEP pathway, these are absolutely the indispensable, highly reactive building blocks for all subsequent complex molecules. As we carefully scale up to the C10 Monoterpenes and the C15 Sesquiterpenes, we immediately see the fascinating biological emergence of volatile signaling, highly toxic botanical defense mechanisms, and deeply potent, highly complex hormonal regulators.
By the exact time the enzymatic cellular machinery successfully stitches together massive C20 Diterpenes and highly rigid C30 Triterpenes, the overall biological utility of these highly complex isoprenoids has dramatically and functionally shifted towards maintaining structural membrane integrity.
The absolute extreme upper limits of this highly comprehensive synthesis table reveal the true, deeply astounding extreme boundaries of biological and biochemical cellular engineering. Massive Polyisoprenoids (containing significantly more than 10 units), such as the vital CoQ10 and Dolichol, flawlessly provide the massive, highly stable tethers absolutely necessary for driving cellular respiration and highly precise human N-glycosylation. Finally, at the absolute, undisputed apex of the table, we clearly see highly specialized natural massive polymers like pure Rubber, which amazingly consist of well over 700 perfectly and flawlessly linked isoprene units. The sheer, staggering scale and deep functional diversity of these specific isoprenoids are totally unmatched in biology.
Please read our Content Disclaimer Statement.
Check out our social media channels: