46. Fatty Acid Oxidation: Mechanisms of Lipid Metabolism
How do migrating birds fly thousands of miles without eating, and how do humans survive weeks of fasting? The secret lies in the body’s most concentrated energy reserve: lipids. This slide deck provides a comprehensive breakdown of the molecular blueprint of lipid metabolism. It explores everything from physiological mobilization and cellular transport to the intricate enzymatic cycles that power cells. Dive into this academic guide to understand the essential biochemical pathways that fuel complex life.
Slide 1: Introduction to Fatty Acid Oxidation and Lipid Metabolism
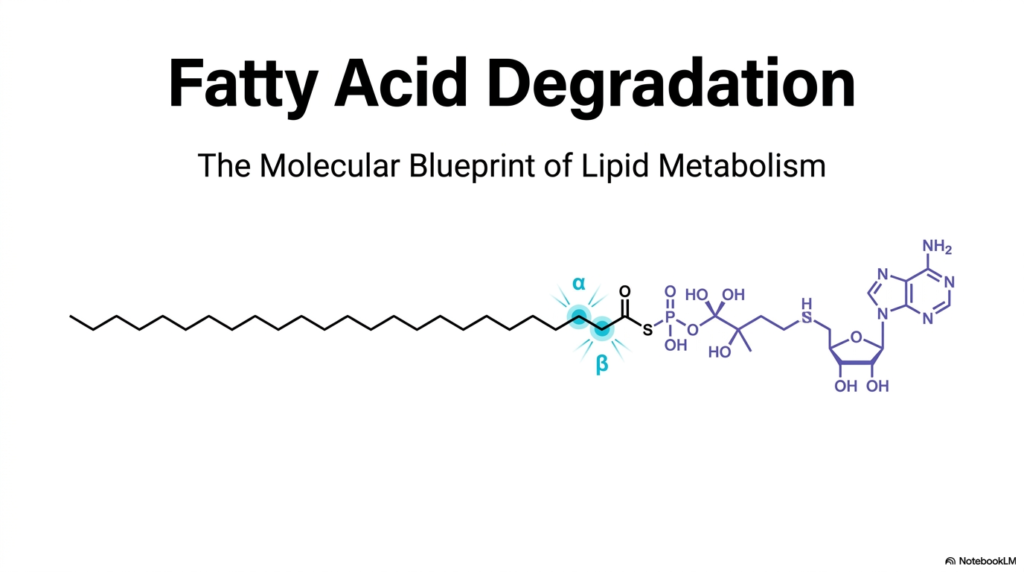
The journey into cellular bioenergetics begins with understanding the structural foundation of lipids. Fatty Acid Oxidation is a central metabolic pathway responsible for breaking down long hydrocarbon chains to generate massive amounts of energy. The title slide illustrates a long-chain fatty acid linked to Coenzyme A, specifically highlighting the alpha and beta carbons. These precise carbon atoms are the structural focal point for the entire degradative process. By targeting the bonds between these carbons, cells can systematically dismantle the lipid molecule.
To fully appreciate the mechanics of Fatty Acid Oxidation, one must examine the molecular blueprint of these energy-dense molecules. The long, unbranched hydrocarbon tail is highly reduced, meaning it holds a wealth of harvestable electrons. When this tail is attached to Coenzyme A, it forms a fatty acyl-CoA thioester. This activation step is crucial because the thioester bond primes the molecule for subsequent enzymatic reactions. The blue highlights on the alpha and beta carbons in the illustration signify where the metabolic machinery will strike first.
As students of biochemistry, visualizing these molecules helps in mastering the complex pathways of Fatty Acid Oxidation. The precise cleavage at the beta-carbon gives the primary degradative pathway its name. This continuous snipping releases two-carbon fragments that will ultimately feed into the citric acid cycle. Understanding this structural orientation is the absolute first step toward grasping how mammalian bodies convert stored fat into usable chemical currency. The overarching goal of this process is to sustain systemic energy during periods of extreme exertion.
The intricate dance of enzymes that facilitates these reactions highlights the elegance of cellular evolution. By mastering the structural blueprint shown here, students can accurately predict how alterations in these molecules might disrupt normal metabolism. This foundational knowledge paves the way for understanding clinical applications, metabolic disorders, and the life-saving nature of lipid breakdown. As the deck progresses, keep this foundational molecular structure at the forefront of your mind.
Slide 2: Evolutionary Advantages Powering Fatty Acid Oxidation
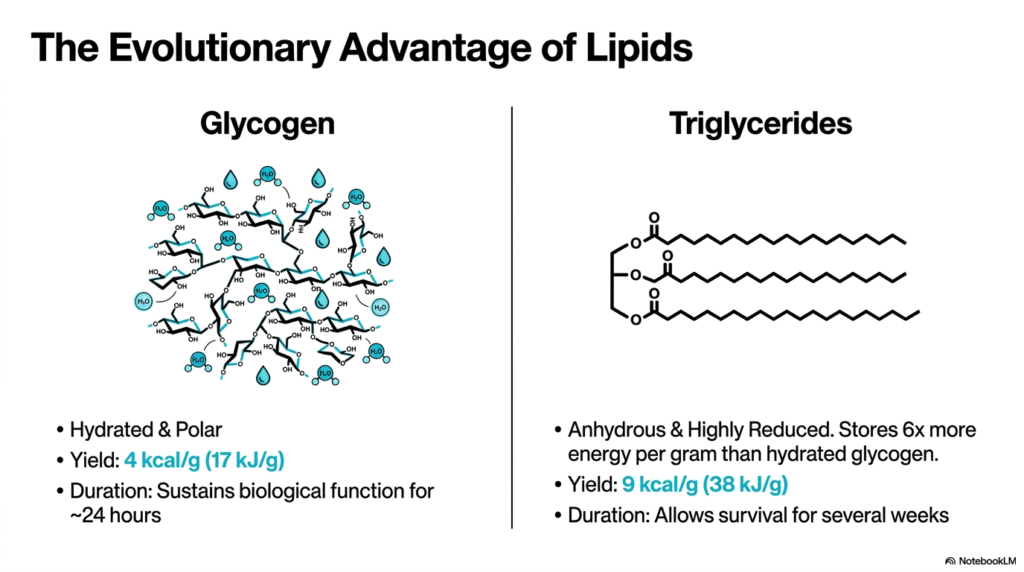
When comparing energy storage molecules, the physiological superiority of lipids becomes immediately apparent. Fatty Acid Oxidation is the preferred method for sustained energy generation due to the unique chemical properties of triglycerides. As the slide demonstrates, glycogen is highly hydrated and polar, meaning it binds a significant amount of cellular water. This extra water weight limits the amount of energy that can be stored efficiently. Consequently, glycogen yields only about 4 kilocalories per gram, sustaining biological function for only 24 hours.
In stark contrast, triglycerides are anhydrous and highly reduced molecules. Because they actively repel water, they can be packed tightly into specialized cells called adipocytes. This dense packing allows lipids to store six times as much energy per gram as hydrated glycogen. Through the biochemical pathway of Fatty Acid Oxidation, these molecules yield an impressive 9 kilocalories per gram. This high energy density is precisely why animals rely on fat stores to survive for several weeks without external sustenance.
From an evolutionary standpoint, the development of Fatty Acid Oxidation provided a monumental survival advantage. Organisms could travel longer distances, survive harsh winters, and endure prolonged starvation. The highly reduced carbon chains of triglycerides provide an abundance of electrons that are eventually transferred to the electron transport chain. Without this efficient, water-free storage system, humans would need to weigh significantly more just to carry enough glycogen for basic survival. This makes lipid metabolism fundamentally essential for complex animal life.
For medical and college students, grasping this thermodynamic difference is absolutely vital. The contrast between 4 kcal/g and 9 kcal/g is not just a statistical fact; it dictates clinical nutrition, human physiology, and metabolic disease management. The mobilization of these dense energy stores ensures that tissues like the heart and skeletal muscle have a constant supply of fuel. This evolutionary masterpiece of energy conservation is the starting point for understanding all downstream metabolic pathways.
Slide 3: Mobilization and Transport in Fatty Acid Oxidation
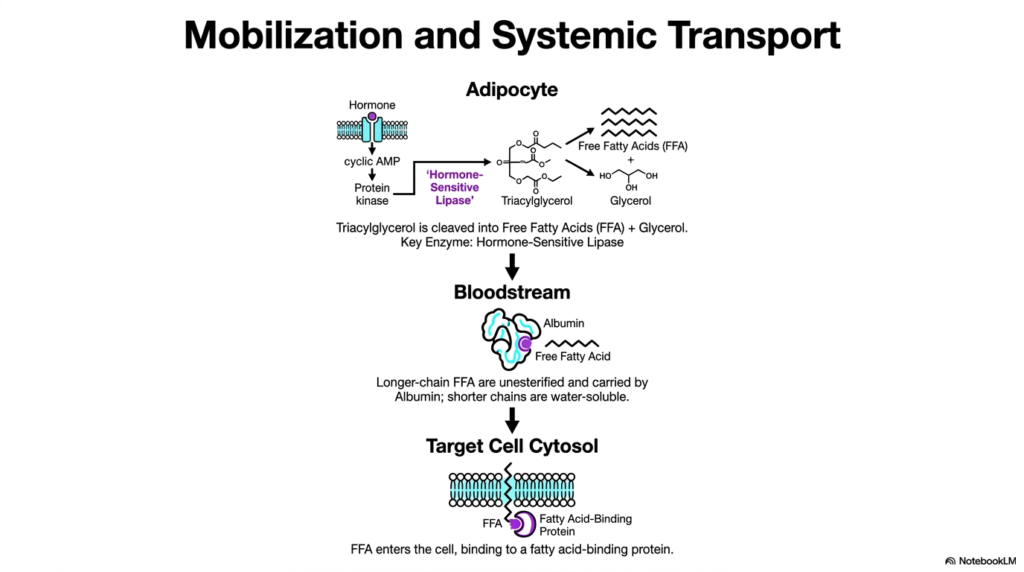
Before cellular energy can be harvested, stored fats must first be mobilized from adipose tissue. The systemic transport system for Fatty Acid Oxidation begins when a hormone, such as glucagon or epinephrine, binds to an adipocyte receptor. This binding triggers an intracellular signaling cascade, elevating cyclic AMP levels and activating Protein Kinase A. This kinase then phosphorylates and activates Hormone-Sensitive Lipase, the key regulatory enzyme that cleaves triacylglycerol into free fatty acids and a glycerol backbone.
Once liberated, these highly hydrophobic free fatty acids cannot travel freely through the aqueous environment of the bloodstream. To facilitate systemic transport for Fatty Acid Oxidation, these molecules bind non-covalently to a specialized transport protein called serum albumin. Albumin acts as a molecular taxi, safely transporting long-chain, unesterified fatty acids through the bloodstream to various target tissues. Without this critical protein binding, the hydrophobic tails would aggregate, causing severe vascular blockages. Shorter lipid chains, however, remain water-soluble enough to travel unassisted.
The biological journey concludes when free fatty acids reach the target cell membrane. They must traverse the lipid bilayer and enter the cytosol, a process that requires further assistance. Upon entry, they immediately bind to a cytosolic Fatty Acid-Binding Protein. This internal chaperone prevents the highly reactive lipids from disrupting cellular membranes or acting as unregulated chemical detergents. This careful cellular handling guarantees that the substrates are safely delivered to the metabolic machinery required for Fatty Acid Oxidation.
Understanding this mobilization cascade is vital for grasping the systemic regulation of human bioenergetics. The activation of Hormone-Sensitive Lipase is a crucial control point at which the body shifts from energy storage to energy consumption. Medical students must recognize that defects in any of these transport mechanisms can lead to profound metabolic disturbances. The seamless handoff from adipocytes to serum albumin to cytosolic binding proteins beautifully illustrates the highly coordinated nature of mammalian metabolism.
Slide 4: Cytosolic Activation Priming Fatty Acid Oxidation
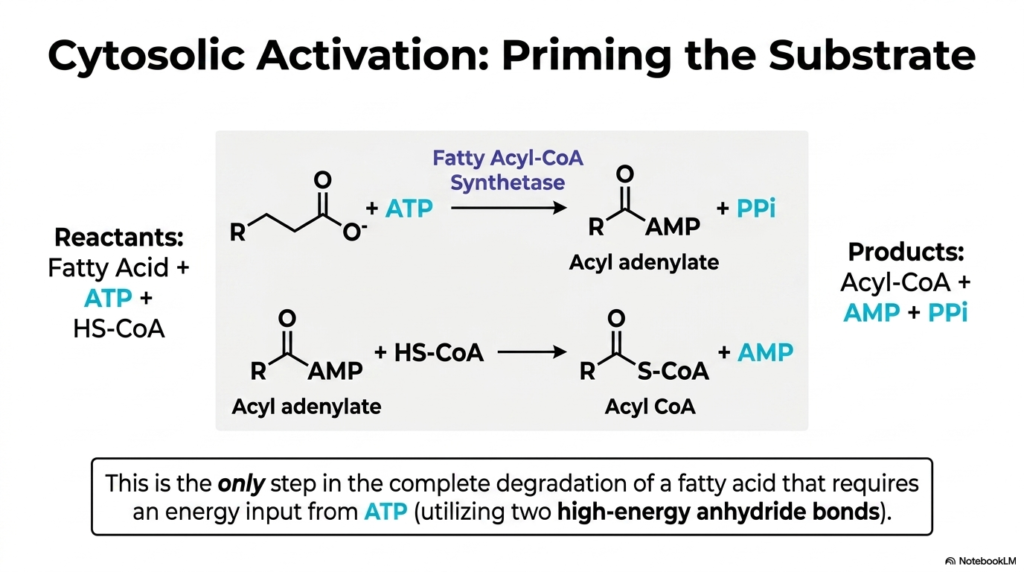
Once inside the target cell, the inert free fatty acid must be chemically primed before it can undergo any degradation. This crucial preliminary step of Fatty Acid Oxidation occurs directly in the cytosol and is catalyzed by the enzyme Fatty Acyl-CoA Synthetase. The reaction functionally links the fatty acid to Coenzyme A, using energy from Adenosine Triphosphate. This specific chemical union creates a high-energy thioester bond, forming Fatty Acyl-CoA, the activated substrate now ready for the mitochondrial machinery.
The biochemistry of this activation is unique and thermodynamically fascinating. The reaction proceeds rapidly via an intermediate known as acyl adenylate, releasing inorganic pyrophosphate into the cytosol. The subsequent hydrolysis of this pyrophosphate strongly drives the reaction forward, making it practically irreversible. This is the only step in the entire process of Fatty Acid Oxidation that directly consumes energy in the form of ATP. Interestingly, it utilizes two high-energy anhydride bonds, costing the cell the equivalent of two ATP molecules.
By investing this initial energetic capital, the cell functionally commits the lipid molecule to its metabolic fate. The addition of the bulky, polar Coenzyme A group prevents the activated molecule from passively diffusing back across the cellular membrane. This biochemical trapping mechanism is essential for ensuring a steady, unidirectional flow of substrates into Fatty Acid Oxidation. The newly formed Fatty Acyl-CoA is now poised to serve as the ultimate fuel source, but it still faces a significant geographical barrier within the cell.
Recognizing the exact energetic cost of this activation is essential for accurate metabolic bookkeeping. When calculating the total cellular ATP yield from lipid degradation, students must remember to subtract the two high-energy bonds invested from the final total. Furthermore, this activation step is a prime example of biological investment: spending a small amount of cellular currency upfront to unlock a massive thermodynamic payload later. This chemically activated molecule is now completely ready to face the mitochondrial transport system.
Slide 5: The Carnitine Shuttle in Fatty Acid Oxidation
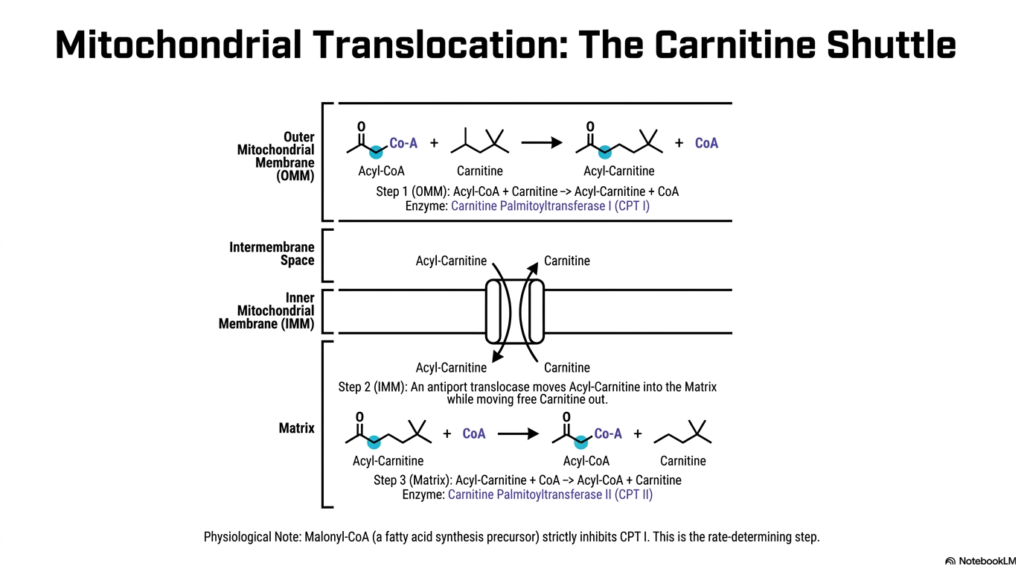
The mitochondrial inner membrane is famously impermeable to large molecules, including the newly activated Fatty Acyl-CoA. To carefully overcome this barrier, the cell employs a specialized transport mechanism known as the Carnitine Shuttle, an absolute prerequisite for mitochondrial Fatty Acid Oxidation. At the outer mitochondrial membrane, the enzyme Carnitine Palmitoyltransferase I exchanges the Coenzyme A group for a molecule of carnitine. This specific reaction forms Acyl-Carnitine, a compound permitted to traverse the mitochondrial barriers.
Once the Acyl-Carnitine is successfully formed, it moves into the intermembrane space and approaches the inner mitochondrial membrane. Here, a dedicated antiport translocase protein moves the Acyl-Carnitine into the mitochondrial matrix while simultaneously moving a free carnitine molecule back out into the intermembrane space. This continuous swapping mechanism ensures an uninterrupted supply of carnitine for ongoing Fatty Acid Oxidation. Once safely deposited within the matrix, the shuttle system’s third and final step can proceed.
Inside the deep mitochondrial matrix, Carnitine Palmitoyltransferase II catalyzes the exact reverse reaction of the outer membrane enzyme. It transfers the acyl group back onto a fresh mitochondrial Coenzyme A molecule, flawlessly recreating the active Fatty Acyl-CoA. The newly freed carnitine is then recycled back through the translocase. This intricate shuttling system ensures that cytosolic and mitochondrial Coenzyme A pools remain strictly separated, a vital regulatory mechanism for Fatty Acid Oxidation and cellular homeostasis.
Clinically, the Carnitine Shuttle represents the crucial, rate-determining step for lipid degradation. The outer membrane enzyme, Carnitine Palmitoyltransferase I, is strictly inhibited by malonyl-CoA, an intermediate of lipid synthesis. This highly logical feedback loop ensures that the cell does not simultaneously build and destroy lipids. Medical students should note that genetic deficiencies in carnitine or these specific transferase enzymes can lead to severe metabolic crises, widely characterized by profound muscle weakness and dangerous drops in blood sugar during fasting.
Slide 6: The Four-Step Engine of Fatty Acid Oxidation
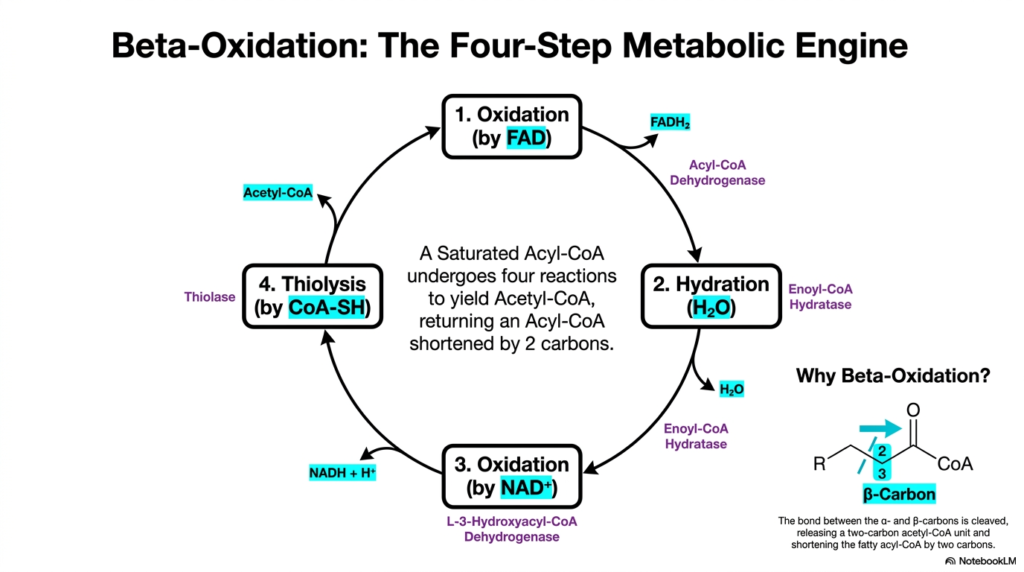
Upon seamlessly entering the mitochondrial matrix, the activated lipid substrate is immediately subjected to a cyclic enzymatic cascade. This core biochemical pathway, Fatty Acid Oxidation, is often referred to as a four-step metabolic engine. The cycle consists of four distinct sequential reactions: a first oxidation, a hydration, a second oxidation, and finally, a thiolysis. By continuously cycling through these four reactions, the cellular machinery can systematically break down a long, saturated hydrocarbon chain into smaller, more manageable chemical fragments.
The entire biological purpose of this enzymatic engine is to cleave specific carbon-carbon bonds. During Fatty Acid Oxidation, the metabolic machinery specifically targets the bond between the alpha-carbon and the beta-carbon of the acyl chain. This precise molecular targeting is exactly why the process is classically termed beta-oxidation. Through the progressive action of dehydrogenases and hydratases, the beta-carbon is chemically oxidized, transforming it from a simple hydrocarbon into a highly reactive carbonyl group.
Each complete turn of this four-step cycle yields impressive biochemical dividends for the cell. The thiolytic cleavage releases a two-carbon unit in the form of Acetyl-CoA, while simultaneously returning an Acyl-CoA molecule that has been shortened by exactly two carbon atoms. In addition to the Acetyl-CoA, this specific phase of Fatty Acid Oxidation produces vital high-energy electron carriers, specifically NADH and FADH2. This newly shortened lipid substrate is then immediately fed back into the cycle for another round of degradation.
For the serious student of biochemistry, viewing this process as a repeating metabolic loop radically simplifies complex cellular accounting. Instead of following a massive linear pathway, one simply repeats the four reactions until the entire lipid chain is completely consumed. The continuous generation of Acetyl-CoA perfectly feeds the downstream citric acid cycle, explicitly linking lipid metabolism to global cellular respiration. This cyclic engine stands as one of the most elegant and efficient energy-generating systems in human biology.
Slide 7: Step 1 Dehydrogenation in Fatty Acid Oxidation
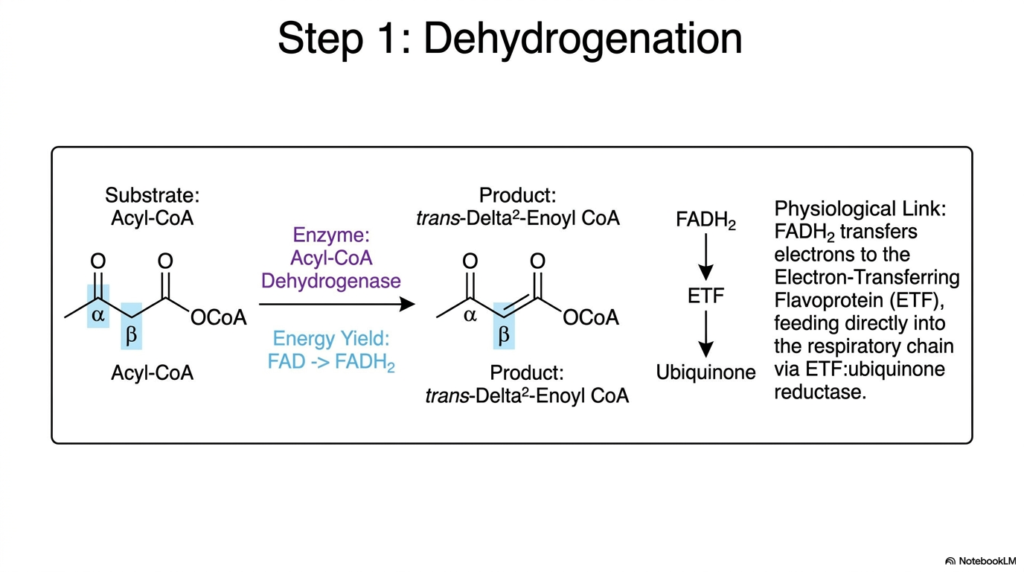
The metabolic engine officially kicks into high gear with a critical initial desaturation reaction. The absolute first step of beta-centric Fatty Acid Oxidation is catalyzed by the enzyme Acyl-CoA Dehydrogenase. This specific enzyme targets the single carbon-carbon bond located directly between the alpha and beta carbons of the saturated lipid substrate. By forcibly removing two hydrogen atoms and their associated electrons, the enzyme creates a new carbon-carbon double bond, radically altering the molecule’s chemical landscape.
The immediate structural outcome of this enzymatic attack is the formation of trans-Delta-2-Enoyl CoA. It is essential to note the specific stereochemistry of the new double bond; it exists exclusively in the trans configuration. Concurrently, the valuable electrons extracted during this initial phase of Fatty Acid Oxidation are securely transferred to an enzyme-bound flavin adenine dinucleotide cofactor, reducing FAD to FADH2. This deliberate electron transfer represents the first direct energy harvest from the lipid hydrocarbon chain.
The generated FADH2 does not simply float away; it is part of a highly integrated mitochondrial metabolic network. It efficiently transfers its freshly acquired electrons to a specialized Electron-Transferring Flavoprotein. This unique protein serves as a physiological bridge, linking the initial biochemical steps of Fatty Acid Oxidation directly to the mitochondrial respiratory chain via an oxidoreductase complex. Ultimately, these electrons flow straight to ubiquinone, powering the proton pumps that drive cellular ATP synthesis.
Understanding the specific nature of Acyl-CoA Dehydrogenase is absolutely crucial for modern clinical diagnosis. The human body utilizes several distinct isozymes of this dehydrogenase, each tailored to specific lipid chain lengths. Genetic mutations resulting in medium-chain acyl-CoA dehydrogenase deficiency are among the most common inborn errors of metabolism. Recognizing this specific enzymatic step helps future physicians understand why these patients cannot tolerate prolonged fasting and require strict dietary management to prevent sudden metabolic collapse.
Slide 8: Step 2 Hydration in Fatty Acid Oxidation
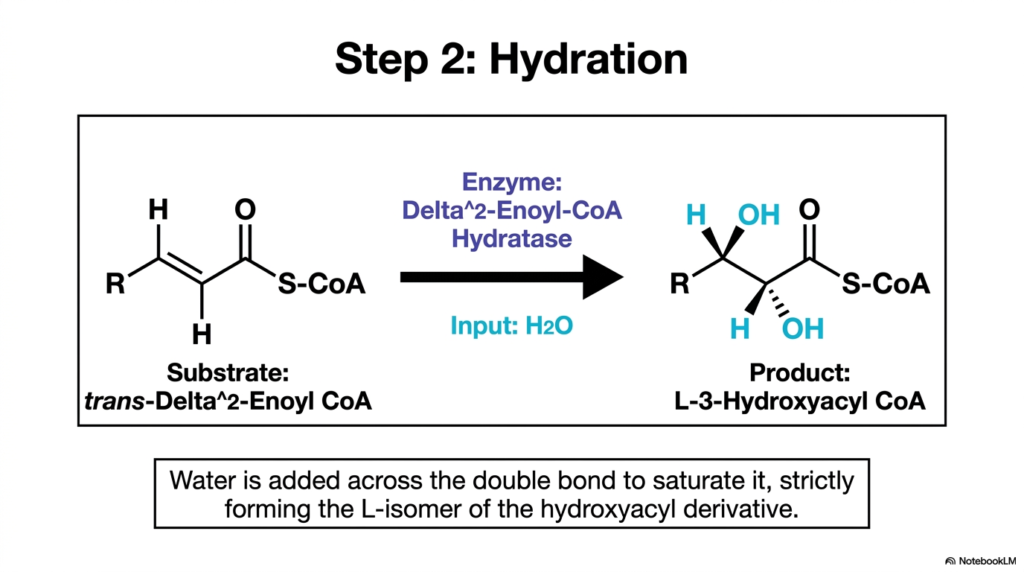
Following the successful creation of the trans double bond, the chemically altered lipid substrate undergoes its second transformation. The second phase of core Fatty Acid Oxidation is a hydration reaction catalyzed by the enzyme Enoyl-CoA Hydratase. This enzyme uses a free water molecule to break the newly formed double bond between the alpha and beta carbons. By carefully adding water across this specific site, the enzyme perfectly prepares the molecule for the critical second oxidation step.
The addition of this water molecule is highly specific and strictly regulated by rigid stereochemical rules. The hydroxyl group from the water molecule is added exclusively to the beta-carbon, while the remaining proton firmly attaches to the alpha-carbon. In the precise context of Fatty Acid Oxidation, this specific mechanism exclusively produces the L-isomer of 3-Hydroxyacyl CoA. The enzyme will accept only the trans-Delta-2 configuration generated in the first step, perfectly demonstrating the absolute lock-and-key specificity of these mitochondrial pathways.
This vital hydration step is essential for drastically altering the electronic structure of the lipid intermediate. By deliberately introducing an electronegative oxygen atom to the beta-carbon, the molecule becomes highly susceptible to further oxidative attack. While this specific step of Fatty Acid Oxidation does not directly yield any energy in the form of electron carriers, it is fundamentally necessary for the progression of the cycle. Without this exact hydration, the subsequent dehydrogenase enzyme would have no proper chemical substrate to act upon.
For absolute academic mastery, medical students must appreciate how biological systems use simple molecules like water to orchestrate incredibly complex chemical rearrangements. Enoyl-CoA hydratase operates with near-perfect catalytic efficiency, seamlessly converting the lipid from an alkene to an alcohol. This transformation beautifully highlights how cellular enzymes manipulate chemical bonds to extract maximum energy later. The newly formed hydroxyl group gracefully sets the perfect stage for the final energy-yielding reaction of the four-step metabolic cycle.
Slide 9: Step 3 Dehydrogenation in Fatty Acid Oxidation
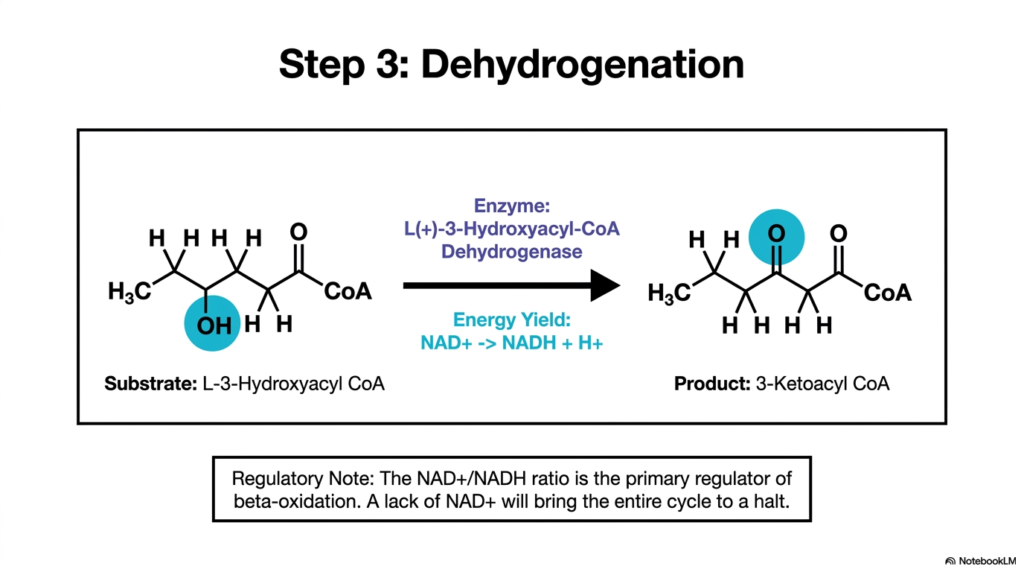
With the hydroxyl group successfully placed on the beta-carbon, the lipid molecule faces its second aggressive oxidative attack. The third crucial step of Fatty Acid Oxidation is carefully catalyzed by L-3-Hydroxyacyl-CoA Dehydrogenase. This specific enzyme specifically targets the freshly added hydroxyl group, forcibly removing two protons and two electrons. This chemical oxidation dynamically converts the simple hydroxyl group into a highly reactive ketone group, radically shifting the thermodynamic potential of the remaining carbon backbone.
The direct biochemical product of this aggressive oxidation is 3-Ketoacyl CoA. Crucially, the electrons extracted during this specific phase of Fatty Acid Oxidation are rapidly accepted by the soluble electron carrier NAD+, effectively reducing it to NADH and releasing a free proton into the matrix. This new NADH molecule is a substantial thermodynamic prize for the cell, as it will rapidly flow to Complex I of the electron transport chain to drive substantial ATP production via oxidative phosphorylation.
This secondary dehydrogenation step actively represents a pivotal regulatory checkpoint for the entire metabolic pathway. The prevailing NAD+/NADH ratio in the mitochondrion is the primary metabolic regulator of Fatty Acid Oxidation. If cellular energy levels are already high, NADH accumulates rapidly, depriving the dehydrogenase of its necessary cofactor. Consequently, a distinct lack of available NAD+ will immediately bring the entire degradative cycle to a grinding halt, perfectly preserving lipid stores for later use.
Medical students should pay incredibly close attention to this regulatory mechanism, as it permanently links lipid breakdown to oxygen availability. Because NADH must be subsequently oxidized by the respiratory chain, the entire process is strictly aerobic. In dangerous conditions of hypoxia or ischemia, such as a myocardial infarction, the electron transport chain stalls, NAD+ is depleted, and lipid degradation abruptly stops. Understanding this molecular bottleneck is essential for comprehending the metabolic consequences of cellular suffocation.
Slide 10: Step 4 Thiolytic Cleavage in Fatty Acid Oxidation
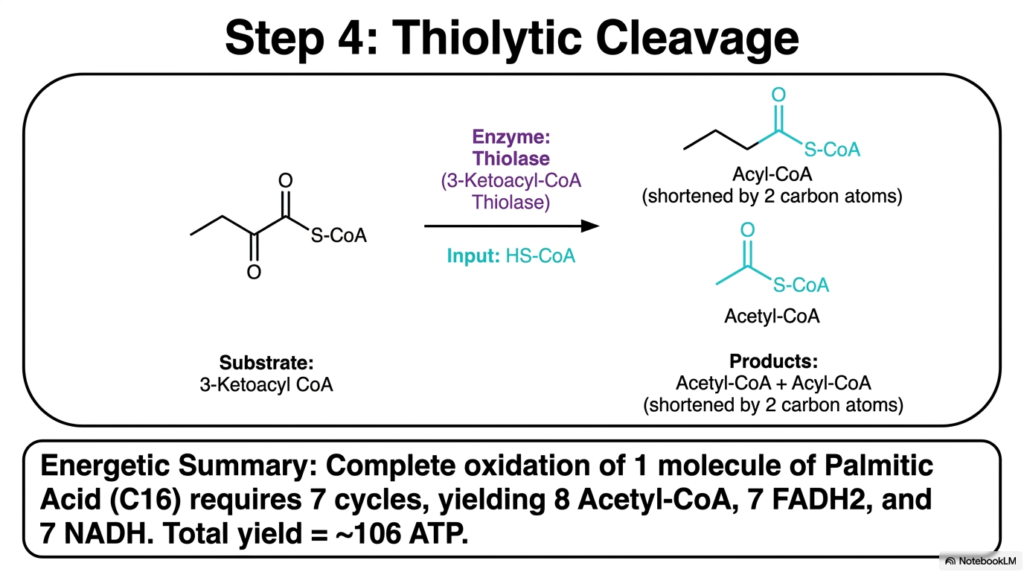
The remarkable metabolic engine safely culminates in a powerful chemical fracture. The fourth and absolute final step of the Fatty Acid Oxidation cycle is brilliantly catalyzed by the enzyme Thiolase, specifically 3-Ketoacyl-CoA Thiolase. The prior introduction of the ketone group severely weakened the chemical bond located between the alpha and beta carbons. Thiolase flawlessly exploits this engineered weakness by using a fresh Coenzyme A molecule to cleave the lipid chain precisely at this molecular junction.
This specific thiolytic cleavage yields two distinct and thoroughly crucial biochemical products. First, it efficiently releases a two-carbon fragment as Acetyl-CoA, a central metabolic hub molecule. Second, it produces a brand new Acyl-CoA molecule that has been successfully shortened by exactly two carbon atoms. This newly shortened product is now perfectly primed to directly re-enter the very first step of Fatty Acid Oxidation, continuously continuing the cycle until the entire hydrocarbon tail is dismantled.
The energetic summary of this repetitive cellular process is astonishingly high. For example, the complete Fatty Acid Oxidation of one molecule of the 16-carbon Palmitic acid requires exactly seven full turns of the metabolic cycle. This comprehensive degradation brilliantly yields eight molecules of Acetyl-CoA, seven molecules of FADH2, and seven molecules of NADH. When fully processed by the respiratory chain and the citric acid cycle, this remarkable breakdown generates a massive net yield of approximately 106 ATP.
Grasping the pure arithmetic of this metabolic engine is a fundamental requirement for biochemistry students. It mathematically illustrates precisely why fats are the body’s undisputed premier energy reserve. The incredible cellular yield of 106 ATP vastly outperforms the energetic output of simple glucose metabolism. The mass production of Acetyl-CoA directly links lipid breakdown to carbohydrate metabolism and ketone body synthesis, demonstrating that cellular pathways are interconnected energetic webs rather than isolated metabolic silos.
Slide 11: Edge Case I – Polyunsaturated Fatty Acid Oxidation
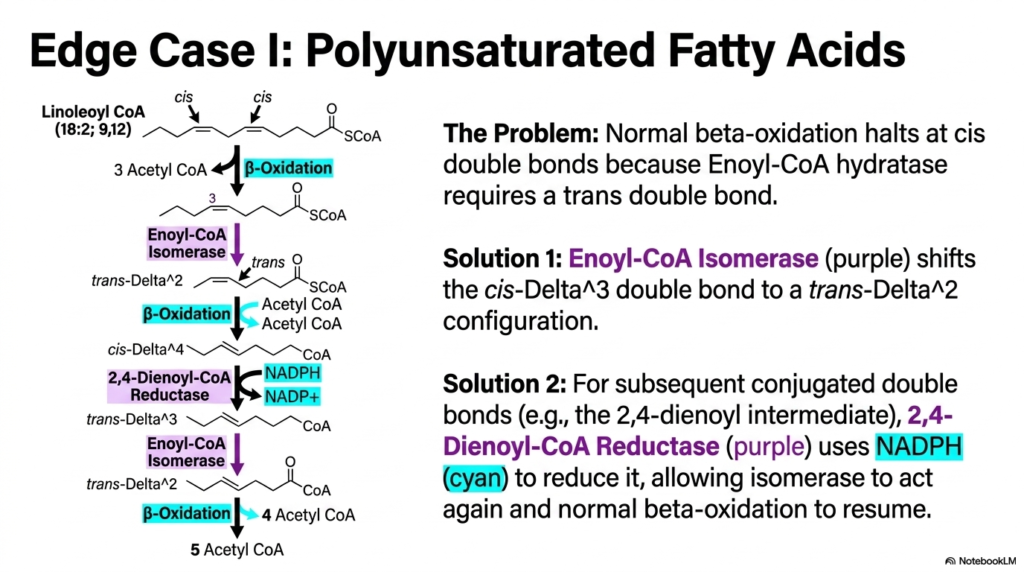
While the standard four-step engine efficiently handles saturated lipids, natural dietary fats are rarely so chemically uniform. The molecular breakdown of polyunsaturated lipids presents a highly unique structural challenge for standard Fatty Acid Oxidation. The standard enzymatic machinery, specifically Enoyl-CoA hydratase, absolutely requires a double bond in the trans configuration between carbons 2 and 3. However, most naturally occurring unsaturated fats contain double bonds explicitly in the cis configuration, which immediately halts the normal metabolic machinery.
To functionally overcome this architectural blockade, the cell intelligently deploys specialized auxiliary enzymes. The first brilliant solution utilizes Enoyl-CoA Isomerase, which physically shifts the problematic cis-Delta-3 double bond smoothly into a compliant trans-Delta-2 configuration. Once isomerized, the altered substrate can easily re-enter the standard pathway of Fatty Acid Oxidation. This relatively simple molecular rearrangement enables the cell to continue extracting energy from common monounsaturated fats such as oleic acid without losing significant metabolic momentum.
For highly complex polyunsaturated molecules with multiple double bonds, an additional enzyme is required. When conjugated double bonds physically form, the cell expertly utilizes 2,4-Dienoyl-CoA Reductase. This remarkable enzyme consumes a molecule of NADPH to reduce the troublesome diene structure, smoothly creating a single double bond that the isomerase can then strategically reposition. These elegant auxiliary steps ensure that no dietary lipid is wasted during Fatty Acid Oxidation, regardless of its initial molecular geometry.
Understanding these specific isomerase and reductase enzymes perfectly highlights the profound adaptability of human metabolism. Medical students must recognize that an optimal clinical diet actively includes many of these beneficial unsaturated fats, such as linoleic acid. The biological cell’s ability to seamlessly integrate these highly complex molecules into its primary energy-generating pathways is a testament to evolutionary efficiency. It demonstrates scientifically that structural variations in biological substrates can be readily manipulated using specialized biochemical tools.
Slide 12: Edge Case II – Odd-Numbered Fatty Acid Oxidation
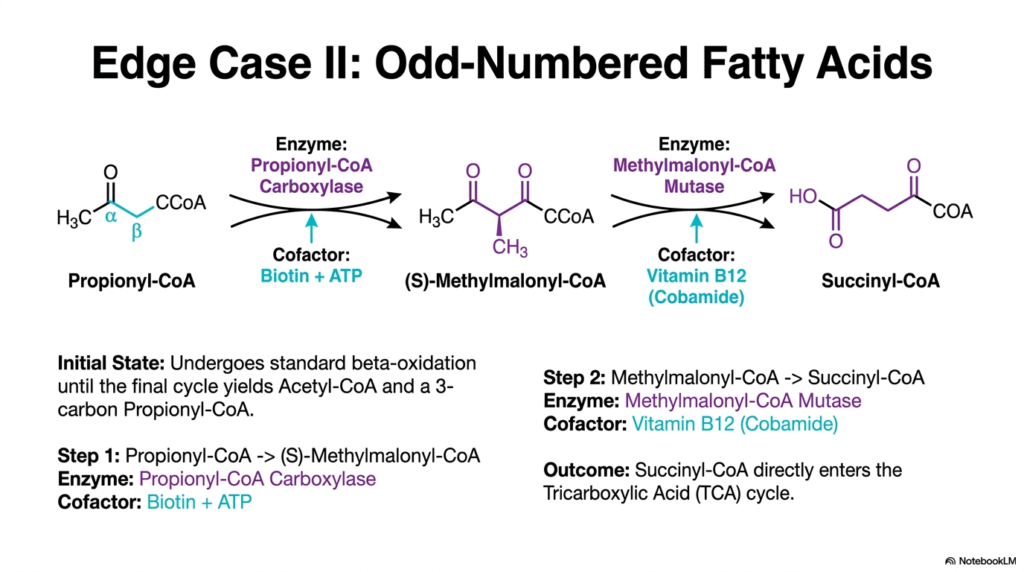
Another truly fascinating exception to standard lipid metabolism involves chains with an odd number of carbon atoms. While the vast majority of dietary lipids are even-numbered, odd-chain variants widely exist in certain plant types and marine organisms. These peculiar molecules successfully undergo standard Fatty Acid Oxidation, continuously cycling through the four-step engine and releasing normal two-carbon Acetyl-CoA units. However, the final enzymatic cycle inevitably leaves behind a highly unique three-carbon fragment famously known as Propionyl-CoA.
The human cell cannot directly utilize Propionyl-CoA for standard energy production, so it initiates a highly specialized biochemical conversion sequence. The very first step firmly involves Propionyl-CoA Carboxylase, an enzyme that uses the vital cofactor biotin and cellular ATP to add a carbon atom, rapidly forming S-Methylmalonyl-CoA. This highly specific intermediate is subsequently manipulated and ultimately acted upon by Methylmalonyl-CoA Mutase. This final crucial step of odd-chain Fatty Acid Oxidation completely requires Vitamin B12 to successfully rearrange the carbon skeleton.
The ultimate outcome of this highly unique enzymatic detour is the direct cellular production of Succinyl-CoA. Unlike the Acetyl-CoA easily generated by standard Fatty Acid Oxidation, Succinyl-CoA is a direct and vital intermediate of the Tricarboxylic Acid (TCA) cycle. This fact means that the biological remnants of odd-chain lipids can contribute to the net synthesis of new glucose via gluconeogenesis, a rare feat that standard even-chain lipids cannot achieve. This provides an essential bridge between lipid and carbohydrate metabolism.
The clinical implications of this specific pathway are profound and are frequently tested on medical board exams. The absolute biological reliance of the mutase enzyme on Vitamin B12 means that B12 deficiencies directly lead to the dangerous accumulation of methylmalonic acid in the blood and urine. Thorough understanding of this specific edge case in lipid metabolism allows astute clinicians to accurately diagnose highly complex nutritional deficiencies and severe demyelinating neurological disorders.
Slide 13: Edge Case III – Alpha-Oxidation and Refsum’s Disease
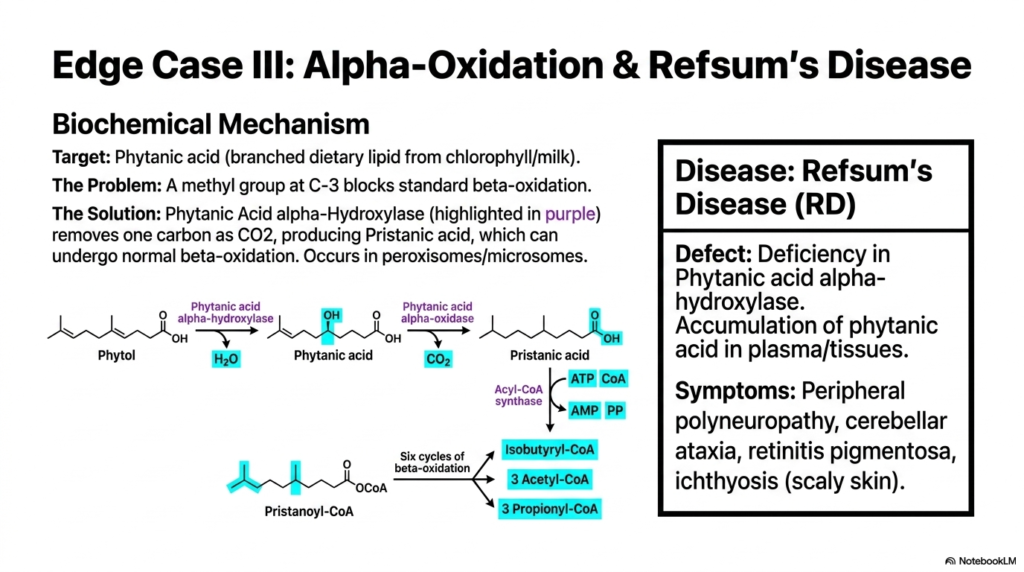
Not all normal dietary fats, unfortunately, possess a simple, unbranched hydrocarbon tail. Phytanic acid, a heavily branched-chain lipid that is highly abundant in chlorophyll-rich diets and dairy, consistently poses a severe metabolic challenge. A bulky methyl group firmly attached to the third carbon completely and physically blocks the standard enzymatic machinery. To bypass this chemical obstruction, the body efficiently directs the complex lipid to peroxisomes and uses an entirely distinct pathway, famously known as alpha-directed Fatty Acid Oxidation.
The precise biochemical solution involves totally sidestepping the blocked beta-carbon altogether. The highly specialized enzyme Phytanic Acid alpha-Hydroxylase selectively targets the alpha-carbon instead. Through this specialized alternative form of Fatty Acid Oxidation, a distinct enzyme hydroxylates the molecule and subsequently removes a single carbon atom as carbon dioxide. This highly unique one-carbon amputation completely converts phytanic acid into pristanic acid, effectively shifting the stubborn methyl branches out of the way of the metabolic enzymes.
Once the physical structural blockage is successfully alleviated, the newly resulting pristanic acid can smoothly undergo standard cycles of beta-cleavage. This completely elegant molecular workaround actively ensures that heavily branched plant lipids can still yield valuable cellular energy. While this minor alternative pathway of Fatty Acid Oxidation technically handles only a small fraction of total dietary intake, its physiological importance becomes tragically clear when the specific alpha-hydroxylase enzyme is genetically defective or entirely absent in a patient.
A severe genetic deficiency in this precise alpha-hydroxylation pathway leads directly to Refsum’s Disease, a brutal neurological condition. Without this specific enzyme, highly toxic levels of phytanic acid rapidly accumulate in the plasma and vulnerable neuronal tissues. Medical students should easily associate this tragic biochemical defect with severe clinical symptoms like peripheral polyneuropathy, cerebellar ataxia, and blinding retinitis pigmentosa. A thorough understanding of this pathway perfectly illustrates how a single missing molecular tool can severely disrupt systemic physiological function.
Slide 14: Edge Case IV – Omega-Oxidation in the Endoplasmic Reticulum
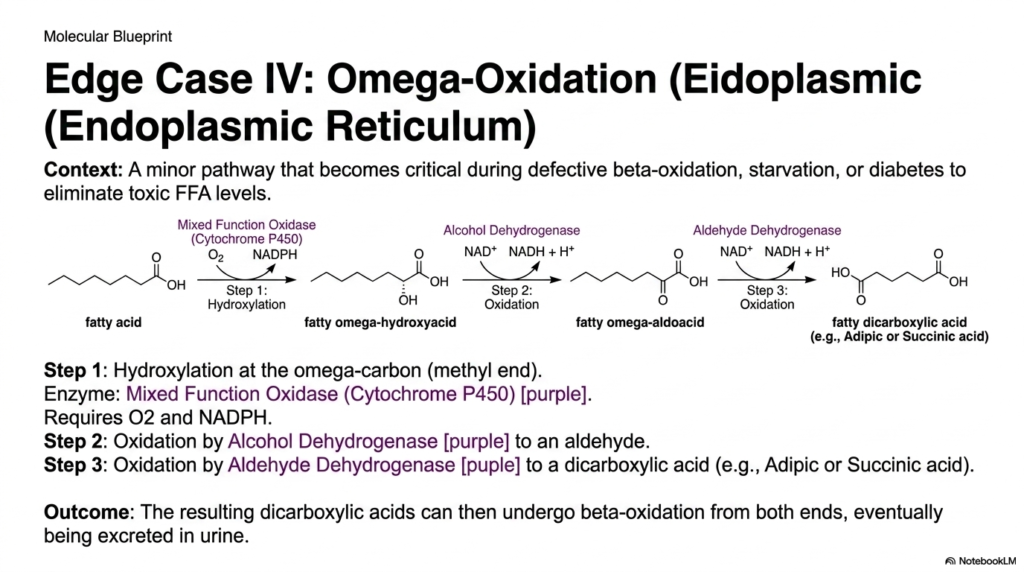
When the primary mitochondrial metabolic pathways become heavily overwhelmed or functionally compromised, the biological cell must rely on emergency rescue systems. Omega-centric Fatty Acid Oxidation is a highly minor, auxiliary physiological pathway strictly located in the Endoplasmic Reticulum. Unlike the standard mitochondrial pathways that aggressively target the carboxyl end, this system attacks the absolute opposite end of the molecule, the omega-carbon. This specific pathway becomes highly biochemically active during prolonged starvation, poorly controlled diabetes, or severe defects in primary lipid metabolism.
The intricate biochemistry of this emergency response is uniquely driven by a specialized series of distinct oxidative enzymes. The very first step involves a Mixed Function Oxidase, a member of the Cytochrome P450 family, which efficiently uses oxygen and NADPH to forcibly hydroxylate the terminal omega-carbon. Following this, the specialized pathway of Fatty Acid Oxidation uses Alcohol Dehydrogenase and Aldehyde Dehydrogenase to sequentially strip electrons. This aggressive oxidation perfectly transforms the simple methyl end into a second, highly reactive carboxyl group.
The absolute ultimate cellular outcome of this specialized Fatty Acid Oxidation pathway is the net production of a highly soluble fatty dicarboxylic acid, such as adipic or succinic acid. Because the lipid molecule now essentially possesses a carboxyl group on both ends, it can smoothly undergo standard beta-cleavage from either direction simultaneously. This rapid degradation strategy brilliantly helps the cell rapidly clear toxic accumulations of medium-chain fats that could otherwise tragically destroy cellular membranes or heavily induce severe metabolic acidosis.
For the diligent future physician, explicitly recognizing the high presence of these specific dicarboxylic acids directly in a patient’s urine is a massive diagnostic clue. It loudly signals that the primary mitochondrial machinery has functionally failed and the Endoplasmic Reticulum rescue pathway is working continuously to prevent systemic lipid toxicity. A thorough understanding of this biochemical failsafe provides critical insight into exactly how the human body adapts to severe metabolic crises, desperately utilizing every available organelle to aggressively maintain energetic homeostasis.
Slide 15: The Diagnostic Matrix of Fatty Acid Oxidation
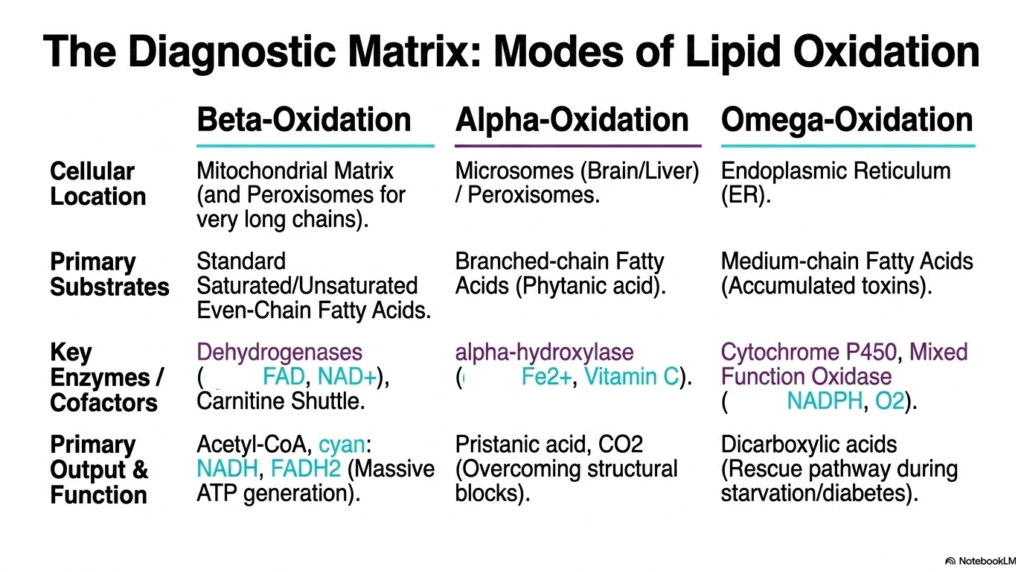
To truly and utterly master complex lipid metabolism, one must efficiently synthesize all available degradation pathways into a highly cohesive physiological framework. This diagnostic matrix clearly serves as the definitive academic summary of the highly diverse modes of Fatty Acid Oxidation. It brilliantly contrasts the standard core mitochondrial engine perfectly with the specialized peroxisomal and endoplasmic reticulum auxiliary pathways. By cleanly organizing these mechanisms by cellular location and primary substrate, students can rapidly identify the correct biochemical tool for any metabolic scenario.
The classic standard beta-cleavage pathway is the dominant pathway in the mitochondrial matrix, successfully handling the vast majority of normal saturated and unsaturated even-chain dietary fats. It is definitively characterized by its strict reliance on the carnitine shuttle and powerful dehydrogenase enzymes. The primary biological function of this central Fatty Acid Oxidation pathway is the massive, sustained ATP generation purely via NADH and FADH2 production. It steadily remains the completely undisputed powerhouse of long-term cellular energy conservation and human endurance.
In stark biological contrast, the alpha- and omega-centric alternative forms of Fatty Acid Oxidation absolutely serve highly specialized, explicitly life-saving metabolic roles. Alpha-oxidation occurs exclusively in peroxisomes, which deploy specialized hydroxylases to efficiently overcome branched-chain structural blocks such as phytanic acid, steadily releasing CO2 in the process. Meanwhile, auxiliary omega-oxidation in the endoplasmic reticulum efficiently uses cytochrome P450 to convert toxic fat accumulations into readily excretable dicarboxylic acids. These pathways are absolutely not primary energy generators, but rather essential molecular detoxification systems.
Thoroughly mastering this distinct diagnostic matrix bridges the significant educational gap between basic biochemistry and clinical pathology. By completely understanding which highly specific form of lipid degradation precisely handles which biological substrate, medical professionals can accurately trace highly complex clinical symptoms directly back to a single defective organelle or missing enzyme. This comprehensive cellular view of lipid metabolism empowers students to see metabolic pathways not as isolated memorization tasks but as a deeply integrated, highly dynamic biological defense system.
Please read our Content Disclaimer Statement.
Check out our social media channels: