49. Cholesterol Biosynthesis: Pathways, Regulation, and Clinical Impact
Imagine a molecule so essential that human life is biologically impossible without it, yet so dangerous that its systemic dysregulation remains a leading global cause of mortality. This fascinating dual nature makes studying this specific lipid fundamentally compelling for future medical professionals. The following presentation systematically unpacks the complex molecular mechanisms, enzymatic pathways, and severe clinical implications of lipid metabolism. Designed specifically for medical and college students, this comprehensive educational guide strictly illuminates the intricate cellular machinery required to maintain human health.
Slide 1: Molecular Mechanisms and Transport in Cholesterol Biosynthesis
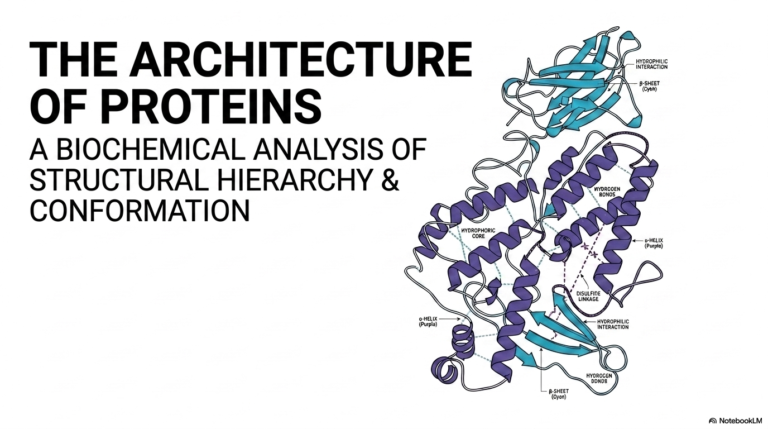
The overarching journey into Cholesterol Biosynthesis begins with understanding its profound biological duality. The introductory slide displays the iconic twenty-seven-carbon sterol structure, composed of four fused hydrocarbon rings, a short hydrocarbon tail, and a crucial hydroxyl group. This specific structural composition confers the molecule’s unique amphipathic properties, enabling it to interface seamlessly with both aqueous environments and hydrophobic lipid bilayers. The intricate study of Cholesterol Biosynthesis represents a fundamental cornerstone of modern biochemistry, effectively bridging basic cellular physiology with widespread clinical pathology. Students must approach this topic recognizing that the endogenous cellular production of this lipid requires substantial energetic investment, using numerous molecules of ATP and NADPH.
Mastering the intricate pathway of Cholesterol Biosynthesis demands a deep appreciation for systemic metabolic integration and cellular compartmentalization. The sterol molecule pictured on the title slide is not merely a static structural component, but rather a dynamic and deeply regulated metabolic hub. Remarkably, every single carbon atom in this incredibly complex molecular architecture originates from acetyl-CoA, a simple two-carbon precursor. The complete cellular synthesis involves dozens of highly specific, sequential enzymatic reactions that occur primarily within the cytoplasm and the smooth endoplasmic reticulum of hepatic cells. This sophisticated cellular machinery is constantly acting to balance the physiological demand for new sterol molecules against the dietary influx of exogenous lipids.
The clinical and therapeutic implications of mastering Cholesterol Biosynthesis are vast and medically vital. Dysregulation in the meticulously controlled pathways of this essential sterol directly contributes to the pathogenesis of atherosclerosis and cardiovascular diseases, making this a critical, high-yield topic for future healthcare professionals. Thoroughly understanding the molecular mechanisms detailed in the subsequent slides provides the essential foundational knowledge required to fully grasp modern pharmacology, systemic pathology, and complex endocrinology. By meticulously dissecting these biochemical steps, learners will discover how targeted medical interventions can inhibit specific metabolic enzymes, thereby restoring systemic equilibrium and preventing progressive cardiovascular disease.
Slide 2: Structural and Endocrine Roles of Cholesterol Biosynthesis
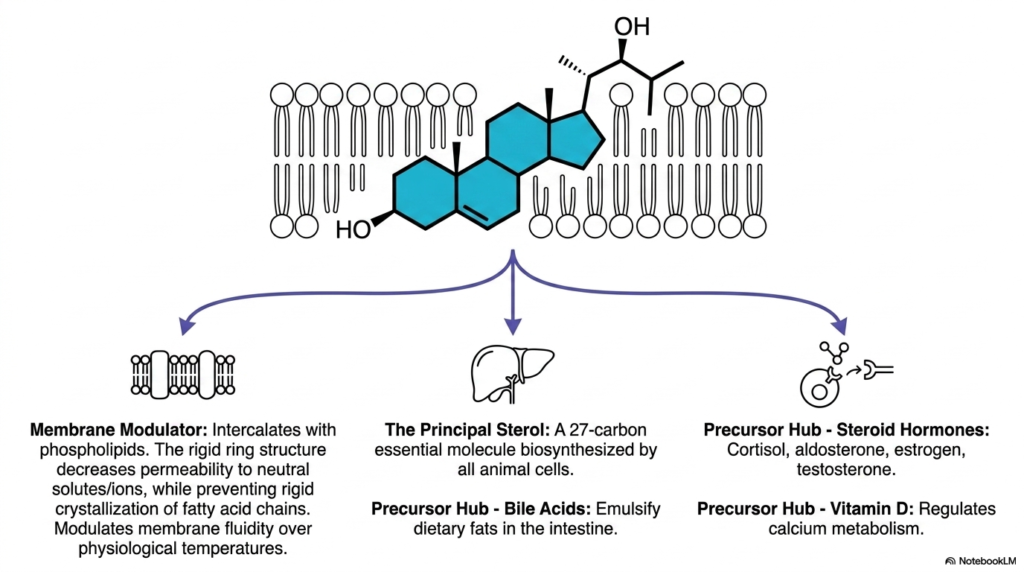
Before delving into the enzymatic steps of Cholesterol Biosynthesis, it is crucial to understand why the body invests so much energy in producing this sterol. The second slide outlines its three primary physiological roles, beginning with its function as a critical membrane modulator. The rigid, planar tetracyclic ring structure intercalates among the flexible fatty acid tails of membrane phospholipids. This precise positioning acts as a bidirectional thermal buffer for the plasma membrane. At higher temperatures, it decreases membrane permeability and prevents excessive fluidity, while at lower temperatures, it physically prevents the rigid crystallization of adjacent fatty acid chains, ensuring cellular viability.
The sheer volume of endogenous lipid generated through the pathway of Cholesterol Biosynthesis is staggering, as it is a mandatory structural component for human survival. Beyond its role in cellular architecture, the liver heavily utilizes this abundant 27-carbon molecule as a fundamental building block for synthesizing bile acids. These amphipathic biological derivatives, such as cholic acid, are massively secreted into the intestinal lumen, where they act as powerful biological detergents. They effectively emulsify complex dietary fats, greatly increasing the surface area available to pancreatic lipases, thereby facilitating proper intestinal digestion and vital caloric absorption.
The diverse molecular products derived directly from Cholesterol Biosynthesis are enzymatically cleaved and modified to produce the entire spectrum of steroid hormones. The slide emphasizes this indispensable role as a universal precursor hub for biologically active compounds. Within the adrenal cortex and gonads, this core sterol is converted into glucocorticoids such as cortisol, mineralocorticoids such as aldosterone, and critical sex hormones, including estrogen and testosterone. Furthermore, through a specialized photolytic reaction in the skin induced by ultraviolet radiation, a direct metabolic intermediate is converted into Vitamin D, which is absolutely vital for maintaining systemic calcium homeostasis and bone health throughout the human lifespan.
Slide 3: The Four Major Stages of Cholesterol Biosynthesis
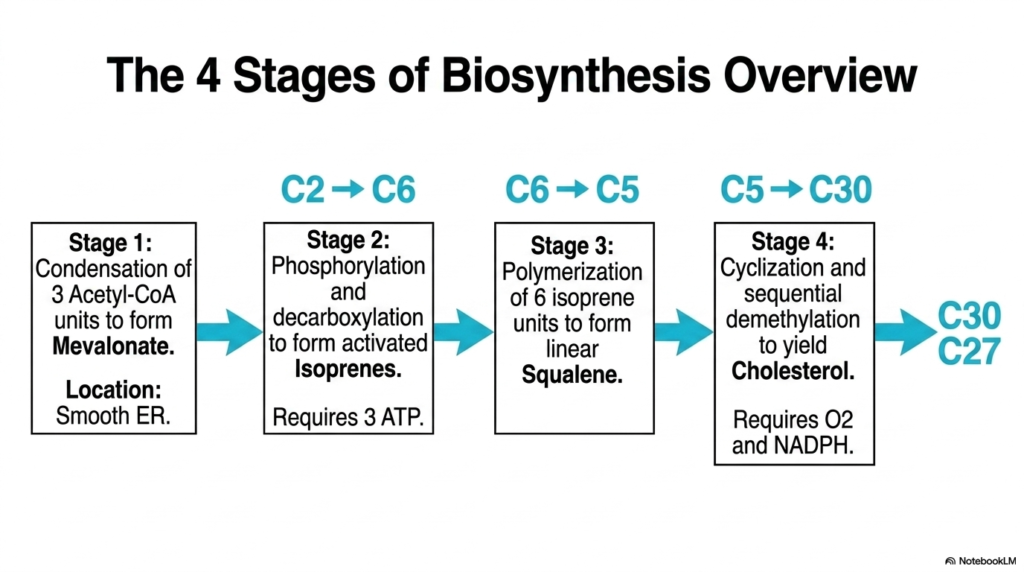
The overarching biochemical process of Cholesterol Biosynthesis is metabolically massive and requires a logical division into four distinct, highly coordinated stages. The third slide provides a broad macro-level overview of this multi-step pathway, tracking the progressive assembly of carbon units. Stage one initiates deep in the cytoplasm and smooth endoplasmic reticulum, where three two-carbon acetyl-CoA molecules are sequentially condensed to form the six-carbon intermediate known as mevalonate. This initial enzymatic fusion represents the foundational building-block phase, committing the cellular metabolic machinery to the energy-intensive lipid production pathway, provided that the cell’s systemic energetic status remains sufficiently high.
The complex pathway of Cholesterol Biosynthesis then proceeds to the second and third stages, which focus intensely on energetic activation and massive linear polymerization. In stage two, the six-carbon mevalonate undergoes serial phosphorylation by powerful kinases, aggressively consuming three molecules of ATP, followed by a sudden decarboxylation event to generate active five-carbon isoprenes. Stage three features the remarkable condensation of six of these five-carbon isoprene units. This rapid enzymatic polymerization yields a linear thirty-carbon hydrocarbon chain called squalene. This elegant mathematical progression highlights the highly conserved evolutionary efficiency of widespread isoprenoid metabolic pathways.
The spectacular final act of Cholesterol Biosynthesis occurs in stage four, in which the linear, highly hydrophobic squalene molecule undergoes a remarkable structural transformation. This intricate stage requires significant input from molecular oxygen and the reducing power of NADPH to facilitate a massive, concerted cyclization reaction. The thirty-carbon chain violently snaps into the characteristic four-ring sterol structure, initially forming lanosterol. Following this immense ring closure, a complex series of enzymatic modifications painstakingly trims the thirty-carbon lanosterol down to the final twenty-seven-carbon sterol product. This rigorous multi-step transformation perfectly illustrates the biochemical complexity required to synthesize such a specialized biological molecule.
Slide 4: Enzymatic Rate-Limiting Steps in Cholesterol Biosynthesis
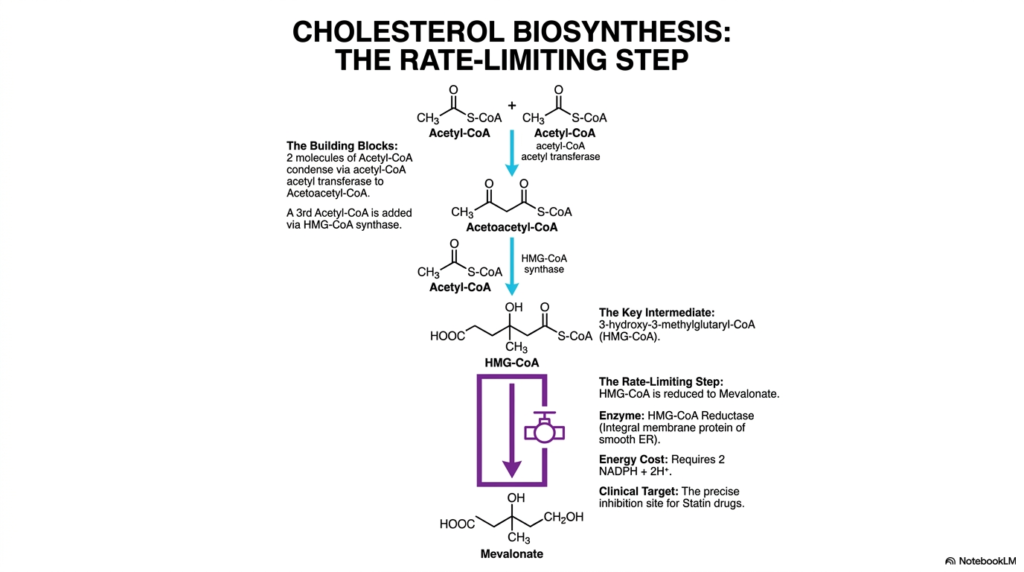
The intricate metabolic pathway of Cholesterol Biosynthesis is not continuously active; it requires strict metabolic governance, primarily exerted at a single enzymatic bottleneck. Slide four visually details this critical rate-limiting step, starting directly with the foundational building blocks. Initially, two molecules of acetyl-CoA are seamlessly joined by acetyl-CoA acetyltransferase to form acetoacetyl-CoA. Subsequently, HMG-CoA synthase adds a third acetyl-CoA molecule, yielding the vital six-carbon intermediate, 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA). This exact enzymatic sequence elegantly demonstrates how the basic metabolic currency, ordinarily used in the rapid citric acid cycle, is dynamically diverted toward complex lipid synthesis.
The true focal regulatory point of Cholesterol Biosynthesis lies with the enzyme HMG-CoA Reductase. This integral membrane protein, securely anchored deep within the smooth endoplasmic reticulum, catalyzes the irreversible reduction of HMG-CoA directly into mevalonate. This catalytic event is highly energy-dependent, strictly requiring the rapid oxidation of two NADPH molecules to systematically strip the thioester linkage and reduce the carboxylate group to a primary alcohol. Because this step is thermodynamically irreversible and highly energy-intensive, it represents the committed step of the pathway. Once mevalonate is successfully formed, the cellular machinery is essentially obligated to carry the reaction through to completion.
Clinically, the rigid pharmacological control of Cholesterol Biosynthesis at this exact biochemical juncture has revolutionized modern cardiovascular medicine. Because HMG-CoA Reductase dictates the overall velocity of the entire metabolic sequence, it serves as the ultimate pharmacological target. The incredibly famous class of lipid-lowering medications known as statins is explicitly designed to act at this active site. By understanding the specific molecular structure of HMG-CoA, medical students can readily visualize how competitive inhibitors effectively block this enzymatic choke point. This precise targeted inhibition continues to save millions of lives globally by halting the excessive accumulation of dangerous systemic lipids.
Slide 5: Activation and Squalene Assembly in Cholesterol Biosynthesis
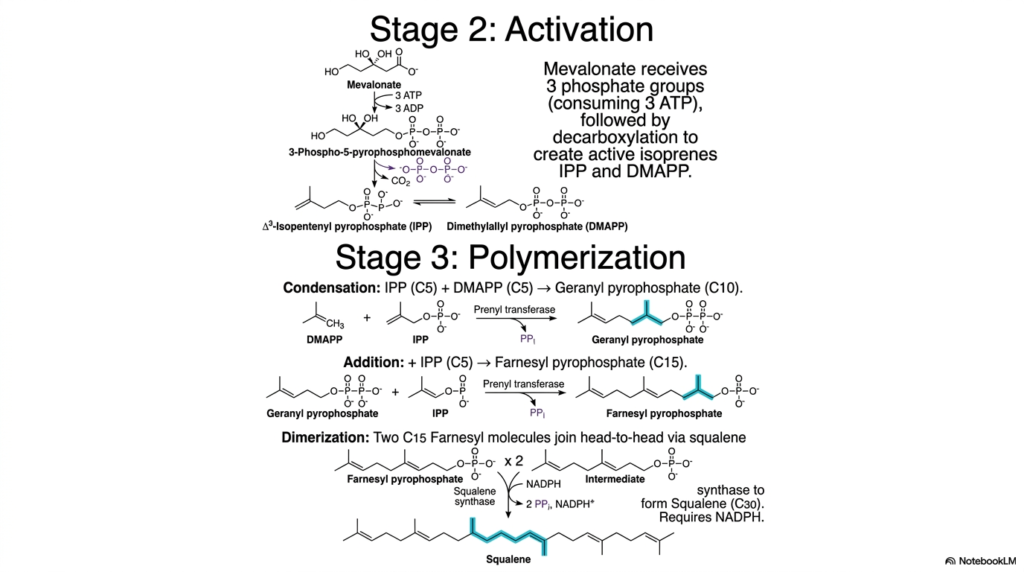
Following the committed step, the pathway of Cholesterol Biosynthesis accelerates into a dynamic phase of molecular activation and rapid elongation. Slide five focuses tightly on Stage two, where the stable six-carbon mevalonate molecule must be chemically activated before it can be used for structural assembly. This vital activation is achieved through the sequential enzymatic addition of three distinct phosphate groups, a demanding process that inherently consumes three massive molecules of cellular ATP. This massive energy investment destabilizes the molecule, perfectly priming it for a critical decarboxylation event that releases carbon dioxide and generates the activated five-carbon isoprenes.
The extreme biochemical elegance of Cholesterol Biosynthesis is vividly displayed during Stage three, the massive polymerization phase. The newly activated five-carbon isoprenes cleverly serve as modular building blocks, using their high-energy pyrophosphate groups as effective molecular leaving groups to drive complex condensation reactions forward. An isopentenyl pyrophosphate molecule merges physically with a dimethylallyl pyrophosphate to form a larger ten-carbon geranyl pyrophosphate. A subsequent massive condensation with another five-carbon unit yields the vital fifteen-carbon farnesyl pyrophosphate. These prenyltransferase reactions efficiently build the massive hydrocarbon chain while continuously hydrolyzing the energetic pyrophosphate bonds to guarantee strict thermodynamic favorability.
The final dramatic sequence in this polymerization phase of Cholesterol Biosynthesis requires the profound catalytic action of the enzyme squalene synthase. Two massive molecules of the fifteen-carbon farnesyl pyrophosphate are brought together in a highly unique head-to-head dimerization reaction. This specific structural alignment absolutely requires the powerful reducing capability of NADPH to permanently fuse the two massive lipid chains, while simultaneously releasing the two remaining pyrophosphate groups. The breathtaking culmination of this intricate biochemical choreography is the immediate formation of squalene, a completely linear thirty-carbon hydrocarbon. The successful creation of this incredibly long molecule completely isolates the lipid deeply within the cellular membrane.
Slide 6: Cyclization and Structural Finalization in Cholesterol Biosynthesis
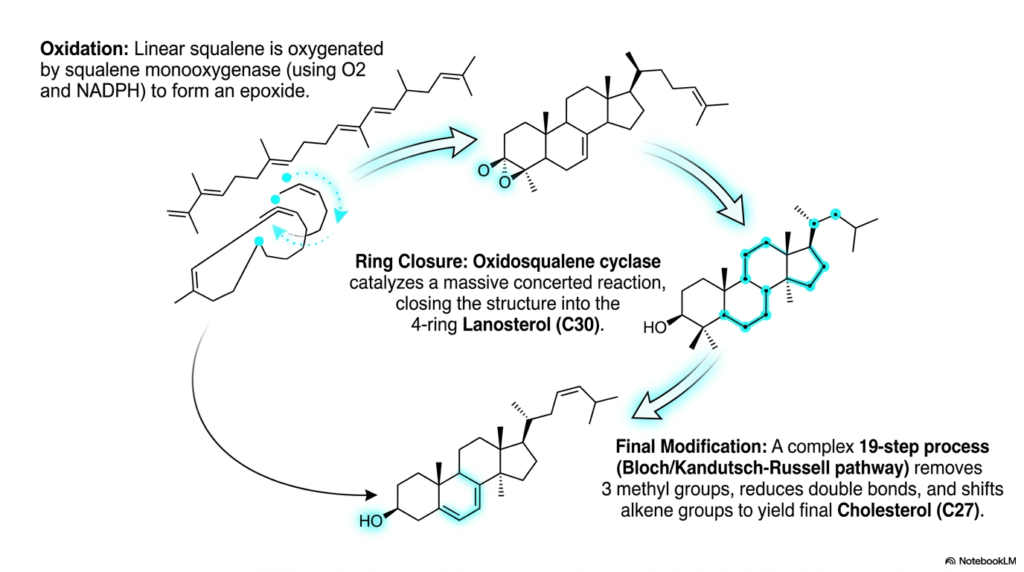
The massive transition from a linear lipid to a complex rigid ring structure represents the most chemically fascinating phase of Cholesterol Biosynthesis. Slide six illustrates this remarkable structural metamorphosis, initiating with the targeted oxidation of squalene. The specific enzyme squalene monooxygenase utilizes molecular oxygen and the reducing equivalent NADPH to carefully introduce a highly reactive oxygen atom directly across a double bond, forming squalene epoxide. This critical biochemical step effectively installs a tightly wound thermodynamic spring within the target molecule. The epoxide ring possesses immense molecular strain, actively storing potential energy required to drive the upcoming structural rearrangement.
The absolutely defining catalytic moment of Cholesterol Biosynthesis is flawlessly executed by the massive enzyme oxidosqualene cyclase. This remarkable structural protein carefully folds the linear squalene epoxide into a highly specific three-dimensional conformation completely buried within its hydrophobic active site. Through a breathtaking single concerted enzymatic reaction, the delicate epoxide ring bursts open, triggering a cascade of electron shifts along the entire thirty-carbon hydrocarbon chain. In a mere fraction of a second, four incredibly stable hydrocarbon rings forcefully snap into molecular existence, yielding lanosterol. This intricate bio-organic mechanism is among the most structurally complex reactions in human biology.
Despite the successful formation of the rigid sterol core, the comprehensive pathway of Cholesterol Biosynthesis is not quite complete. The newly minted lanosterol molecule must immediately undergo a highly tedious, complex nineteen-step enzymatic refinement process, academically referred to as the Bloch or Kandutsch-Russell pathway. These subsequent vital modifications primarily occur deeply within the smooth endoplasmic reticulum and involve the systematic chemical removal of three protruding methyl groups, the delicate reduction of specific double bonds, and the careful shifting of alkene groups. The cellular machinery painstakingly executes these chemical steps with absolute precision to avoid the lethal accumulation of toxic intermediate sterols.
Slide 7: Metabolic and Hormonal Regulation of Cholesterol Biosynthesis
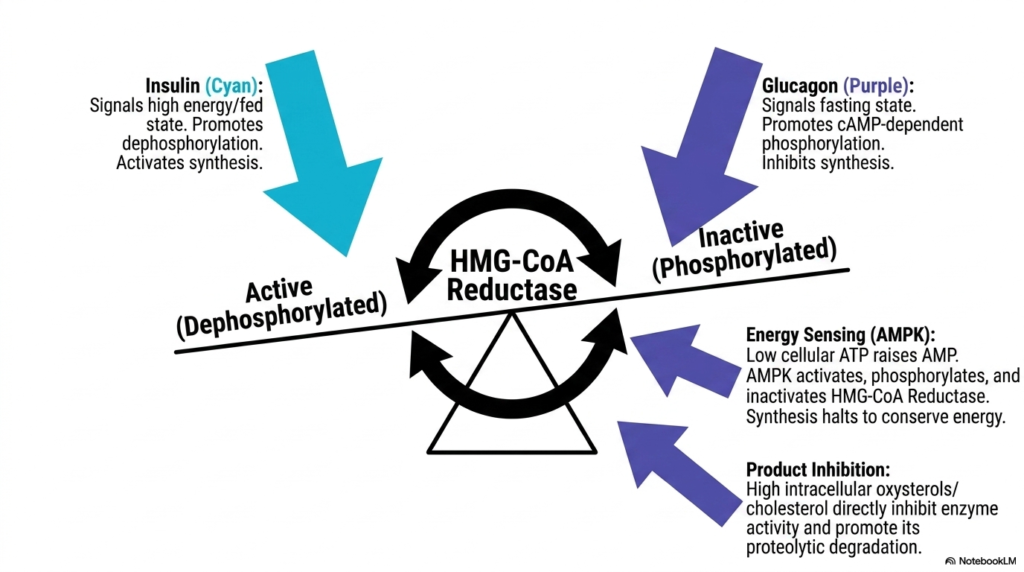
Because the endogenous cellular production of sterols is extremely energetically taxing, Cholesterol Biosynthesis must be meticulously governed by global energetic signals and changing hormonal states. Slide seven breaks down the intensely complex regulatory web surrounding HMG-CoA Reductase, the primary biochemical control valve of the pathway. The absolute most immediate mechanism of cellular control is rapid covalent modification via phosphorylation. Hormones signaling a high-energy or fed state, notably pancreatic insulin, aggressively activate cellular phosphatases that swiftly strip phosphate groups from HMG-CoA Reductase. This targeted dephosphorylation rapidly forces the essential enzyme into its highly active conformational state, massively upregulating rapid sterol production.
Conversely, during times of starvation or cellular stress, the intensive pathway of Cholesterol Biosynthesis is immediately shut down to actively conserve vital energy. Glucagon, the primary systemic fasting hormone, instantly triggers a massive cAMP-dependent signaling cascade that results in the intense phosphorylation and subsequent rapid inactivation of the reductase enzyme. Furthermore, an internal cellular energy sensor, AMP-activated protein kinase, serves as a vital metabolic safeguard. When intracellular ATP levels drop dangerously low, rising AMP levels strongly activate this specific kinase, which directly phosphorylates and firmly halts the enzyme, strictly ensuring the cell does not waste scarce biological energy.
The complex biological regulation of Cholesterol Biosynthesis also features a brilliant negative feedback loop directly driven by the final sterol product itself. When intracellular concentrations of sterols or oxygenated sterol derivatives rapidly rise, they physically bind to the massive reductase enzyme and its associated regulatory membrane proteins. This specific binding not only profoundly inhibits the enzyme’s intrinsic catalytic activity through structural allosteric changes but also dramatically accelerates its proteolytic degradation. The cellular machinery efficiently ubiquitinates the specific enzyme, aggressively tagging it for rapid destruction within the proteasome. This multi-layered regulatory network effectively prevents the highly lethal accumulation of cellular lipids.
Slide 8: Transcriptional Control and SREBP in Cholesterol Biosynthesis
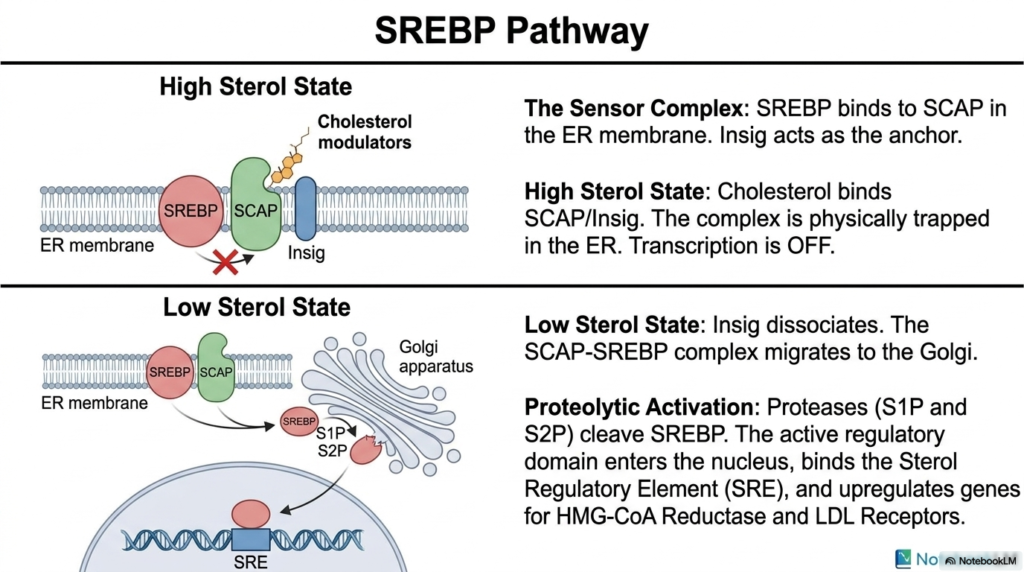
While enzymatic phosphorylation offers immediate metabolic control, long-term regulation of Cholesterol Biosynthesis requires actively altering the transcription of key metabolic enzymes. Slide eight beautifully illustrates the elegant Sterol Regulatory Element-Binding Protein pathway, an incredibly sophisticated intracellular lipid sensing mechanism. SREBP is a crucial transcription factor that is continuously synthesized as an inactive precursor, securely anchored within the dense endoplasmic reticulum membrane. It is tightly bound to a specialized chaperone-sensing protein called SCAP, which, in turn, is strictly anchored by a retention protein called Insig. During states of high sterol concentration, this trimeric protein complex remains physically trapped.
When the intracellular environment is severely depleted of sterols, the vital genetic machinery driving Cholesterol Biosynthesis must be rapidly activated. In a dangerously low-sterol state, the complex’s structural conformation changes, causing it to completely dissociate from the highly restrictive Insig anchor. Freed entirely from its molecular tether, the newly liberated SCAP-SREBP complex is quickly packaged into specific transport vesicles and dynamically migrates from the endoplasmic reticulum directly toward the Golgi apparatus. This massive spatial translocation deep within the active cell is the absolutely critical prerequisite for the subsequent genetic activation required to restore healthy lipid levels.
Upon arriving at the Golgi, the ultimate genetic activation of Cholesterol Biosynthesis occurs through precise proteolytic cleavage. Two highly specific resident membrane proteases, known clinically as S1P and S2P, systematically snip the dense SREBP protein, physically severing its highly active regulatory domain straight from the lipid membrane anchor. This newly liberated, highly soluble transcription factor travels swiftly deep into the central cellular nucleus. There, it actively binds to specific genetic promoters perfectly known as Sterol Regulatory Elements, massively upregulating the genetic transcription of the HMG-CoA Reductase gene. This brilliant genetic mechanism elegantly ensures the cell can simultaneously synthesize essential structural lipids.
Slide 9: Lipoprotein Packaging Following Cholesterol Biosynthesis
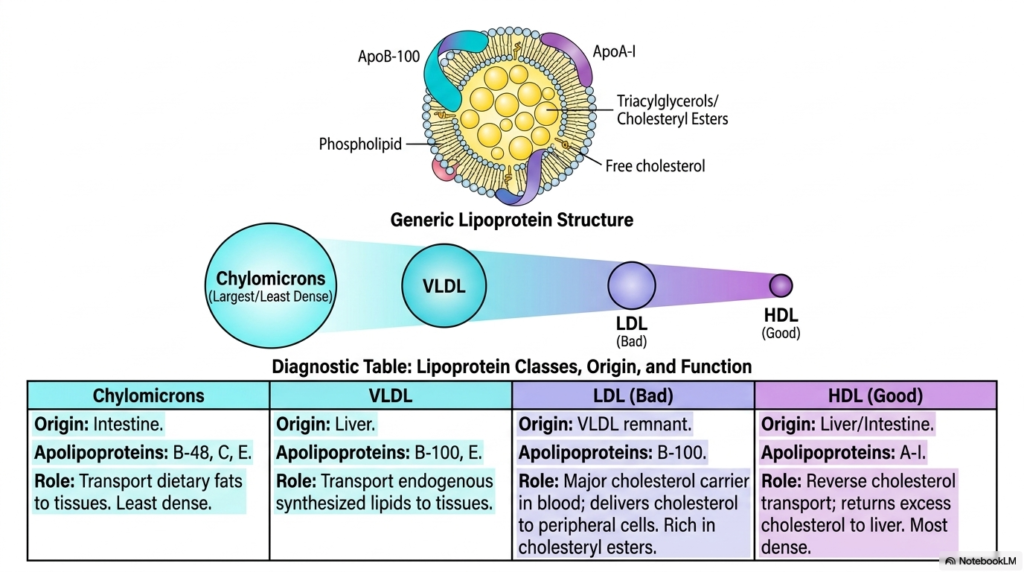
The high hydrophobicity of the mature sterols produced via Cholesterol Biosynthesis prevents them from readily dissolving in the aqueous environment of human blood. To fully address this strict physiological problem, the human body actively utilizes highly specialized spherical transport vehicles, formally known as lipoproteins, as detailed on slide nine. These elegant biological complexes feature a highly protective outer shell composed exclusively of amphipathic phospholipids and specific targeting apolipoproteins, which effectively shields a highly hydrophobic inner core tightly packed with triacylglycerols and large esterified sterols. This specific molecular architecture enables the safe distribution of vital hepatic lipids to distant human tissues.
Understanding the major classes of complex lipoproteins is essential for linking the fundamental pathway of Cholesterol Biosynthesis to human physiology. Chylomicrons are structurally the largest and least dense circulating particles, exclusively responsible for packaging dietary fats directly from the human intestine and delivering them throughout the vast vascular body. The human liver, strictly acting as the central metabolic command center, forcefully exports its massive endogenously synthesized sterols inside Very Low-Density Lipoproteins. As these massive lipid particles rapidly travel through the systemic human circulation, they are progressively enzymatically stripped of their internal triacylglycerols, causing them to physically shrink.
The massive systemic distribution of the crucial lipids generated by Cholesterol Biosynthesis ultimately relies on the continuous creation of Low-Density Lipoprotein, the infamous “bad” systemic cholesterol carrier. LDL is naturally formed directly from the exhausted remnants of VLDL and is uniquely structurally characterized by its solitary ApoB-100 surface protein. Its primary, strictly biological mandate is to effectively deliver massive payloads of vital esterified sterols to hungry peripheral tissues. Recognizing these highly unique biological roles, diagnostic apolipoprotein markers, and varying physical particle densities is absolutely paramount for dedicated medical students actively learning to accurately interpret complex clinical lipid panels.
Slide 10: Cellular Uptake and Receptor Recognition After Cholesterol Biosynthesis
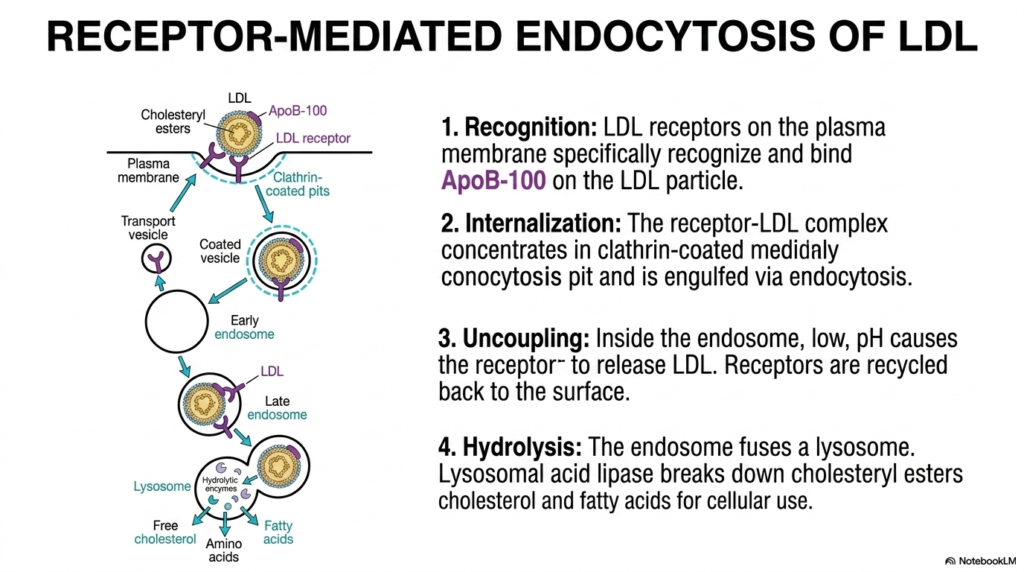
The abundant systemic lipids generated from active hepatic Cholesterol Biosynthesis must successfully cross the external plasma membrane of peripheral cells to be properly utilized. Slide ten visually details the highly elegant mechanism of receptor-mediated endocytosis, an incredibly specific cellular import process. Cells that require large amounts of structural sterols or essential hormonal precursors strongly upregulate the surface expression of specialized target LDL receptors. These critical cellular receptors possess a highly specific biochemical affinity perfectly tuned for the ApoB-100 protein, which is exclusively found on the outer protective shell of circulating LDL particles. This incredibly precise lock-and-key recognition ensures that cells import only specific lipid packages.
Once the cellular receptor firmly binds its specific target, the mature products of Cholesterol Biosynthesis are physically dragged directly into the deep intracellular space. The external cell membrane invaginates at specialized zones heavily coated with the scaffolding protein clathrin, forming a highly concentrated endocytic pit. This specific membrane section violently pinches off, swiftly forming a highly secure intracellular transport vesicle that perfectly contains the intact LDL particle. As this specialized vesicle structurally matures into an early endosome, powerful molecular proton pumps rapidly lower the internal pH, thereby altering the three-dimensional conformation of the target receptor.
The highly coordinated final stage of cellular acquisition following active Cholesterol Biosynthesis involves rigorous enzymatic digestion and internal recycling. The completely unbound, delicate LDL receptors are efficiently clustered into specific transport vesicles and rapidly recycled back to the external human cell surface to seamlessly continue scavenging. Meanwhile, the dark endosome, which completely contains the isolated LDL particle, fuses directly with a cellular lysosome. Powerful lysosomal acid lipases aggressively hydrolyze the particle, rapidly breaking apart the dense hydrophobic core of cholesteryl esters. This violent enzymatic action rapidly releases pure sterol molecules directly into the active cytoplasm for immediate cellular use.
Slide 11: Reverse Transport and the Clearance of Cholesterol Biosynthesis Products
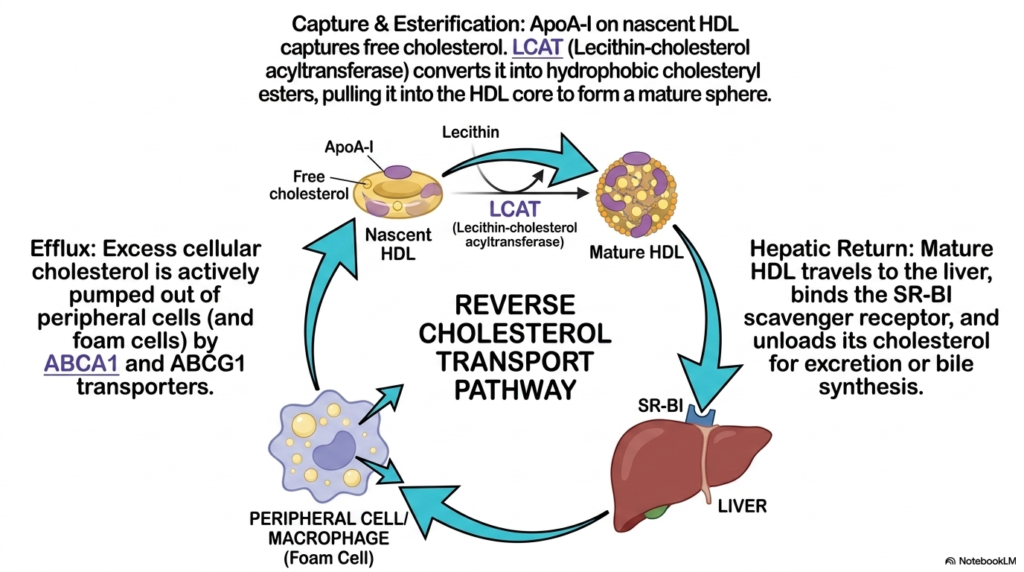
Because peripheral tissues entirely lack the necessary enzymatic machinery to fully degrade the resilient molecular products of Cholesterol Biosynthesis, the body requires a dedicated extraction mechanism. Slide eleven expertly maps out the Reverse Cholesterol Transport pathway, a highly crucial physiological process that aggressively prevents widespread cellular lipid toxicity. This vital salvage operation is spearheaded by High-Density Lipoprotein, which begins its systemic lifecycle as a flattened, nascent disc synthesized by the liver. This empty lipid particle aggressively patrols the peripheral circulation, utilizing its specific ApoA-I surface protein to physically interact with specialized cellular ABCA1 flippase transporters located strictly on lipid-loaded macrophages.
The massive physiological retrieval of the hydrophobic molecules produced by active Cholesterol Biosynthesis requires an aggressive enzymatic trapping mechanism. As the peripheral cellular transporters actively pump large free sterol molecules out of the bloated cell and onto the nascent protective HDL particle, a specialized enzyme known as LCAT immediately springs into violent action. LCAT aggressively esterifies these massive free sterol molecules, chemically attaching a highly bulky fatty acid chain directly to the exposed hydroxyl group. This massive biochemical modification permanently transforms the target molecule into a highly hydrophobic cholesteryl ester, forcing it to dive deep into the particle’s core.
The ultimate physiological goal of this vital pathway is to safely return the excess molecular products of Cholesterol Biosynthesis back to the hepatic system for proper biological disposal. The fully engorged, massive mature HDL sphere travels incredibly swiftly through the vast systemic bloodstream directly toward the human liver. Upon arrival, the dense HDL particle physically docks tightly with a highly specialized hepatic scavenger receptor, specifically SR-BI. The human liver aggressively and selectively extracts the massive payload of dense sterol esters straight from the particle’s internal core without completely destroying the protective protein shell. This dynamic recycling system strictly maintains human homeostasis.
Slide 12: Pathological Atherosclerosis and the Failure of Cholesterol Biosynthesis Regulation
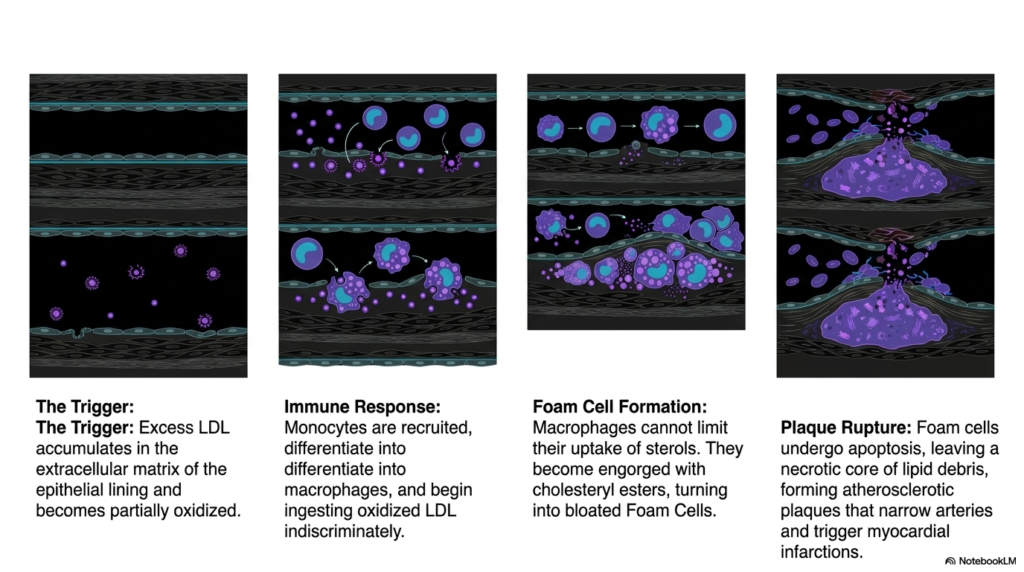
When the carefully balanced machinery of active Cholesterol Biosynthesis and systemic lipid transport fails completely, the resulting physiological consequences are devastating. Slide twelve darkly illustrates the progressive, deadly pathophysiology of atherosclerosis, the unquestioned leading cause of widespread global cardiovascular mortality. The highly insidious trigger for this deadly vascular disease strictly occurs when an immense excess concentration of circulating LDL particles slowly penetrates the physically damaged internal endothelial lining of major human arteries. Once tightly trapped within the subendothelial matrix, these highly delicate lipid particles are constantly bombarded by local destructive oxygen species, becoming heavily oxidized into toxic molecular debris.
This progressive pathological accumulation of intensely oxidized lipids resulting from unregulated Cholesterol Biosynthesis triggers a massive, systemic immune crisis. Circulating immune monocytes are rapidly recruited directly to the active arterial injury site, forcefully squeezing through the delicate endothelial cell gaps. Once deep within the intimate, dark arterial layer, these active cells physically differentiate exclusively into aggressive macrophages that begin frantically ingesting the highly toxic oxidized LDL. Because these specific primitive immune receptors deliberately and violently bypass normal cellular feedback inhibition, the highly active macrophages continue to gorge themselves endlessly on the massive sterol esters, rapidly transforming into completely bloated, highly dysfunctional Foam Cells.
The ultimate lethal systemic consequence of completely deranged Cholesterol Biosynthesis is the massive structural failure of the delicate arterial wall. As countless bloated Foam Cells rapidly undergo programmed cellular death via apoptosis, they violently spill their highly toxic lipid contents directly into the intimate arterial space, rapidly creating a massive, deadly necrotic core of dark, greasy debris. This highly progressive accumulation rapidly forms a physically unstable atherosclerotic plaque that physically intensely narrows the internal, delicate arterial lumen. Understanding this highly pathological cascade is absolutely critical for medical students strictly preparing to actively treat severe catastrophic cardiovascular diseases in complex clinical practice.
Slide 13: Pharmacological Inhibition and Clinical Targets of Cholesterol Biosynthesis
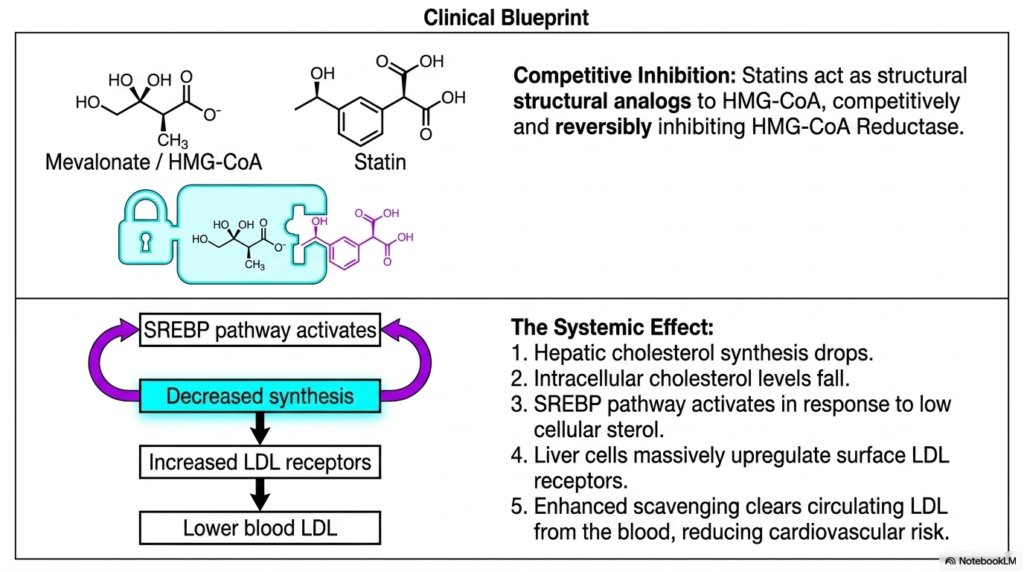
The immense clinical danger posed by excessive systemic lipids has driven the intensive development of powerful pharmacological interventions specifically targeting active Cholesterol Biosynthesis. Slide thirteen visually and flawlessly decodes the precise, complex molecular mechanism of the famous statin medications, the unquestioned modern gold standard of cardiovascular pharmacotherapy. As beautifully depicted, the highly distinct chemical structure of a synthetic statin drug perfectly mimics the natural three-dimensional conformation of the natural HMG-CoA intermediate. Because of this massive structural mimicry, the pharmaceutical molecule aggressively acts as a highly effective competitive inhibitor, physically violently jamming itself incredibly deeply into the active catalytic enzymatic site.
The targeted biochemical disruption of hepatic Cholesterol Biosynthesis utilizing this precise competitive inhibition triggers a profound, systemic metabolic cascade. By successfully completely halting the highly essential rate-limiting step strictly within the dense hepatic smooth endoplasmic reticulum, internal cellular synthesis of complex new sterol molecules abruptly plummets. This sudden drug-induced massive intracellular lipid deficit deliberately and rapidly triggers a widespread cellular panic response. The human hepatic cell rapidly detects a critically low internal sterol environment, immediately uncoupling the sensitive SCAP complex and dramatically activating the powerful SREBP pathway. The human liver intensely upregulates the rapid genetic transcription of crucial surface receptor proteins.
The true clinical brilliance of pharmacologically manipulating endogenous Cholesterol Biosynthesis lies in the powerful secondary systemic effects of this targeted genetic activation. Forced exclusively by the statin medication to rapidly acquire massive amounts of necessary structural lipids directly from circulating sources, the hepatic cells massively increase the physical biological density of active LDL receptors on their external cellular membranes. This highly intense physical receptor upregulation brilliantly transforms the human liver into a massive, powerful biological vacuum, aggressively and constantly pulling dangerous circulating LDL particles directly and violently out of the human bloodstream, thereby severely reducing the risk of catastrophic cardiovascular events.
Slide 14: Physiological Excretion and Vital Utilization of Cholesterol Biosynthesis Products
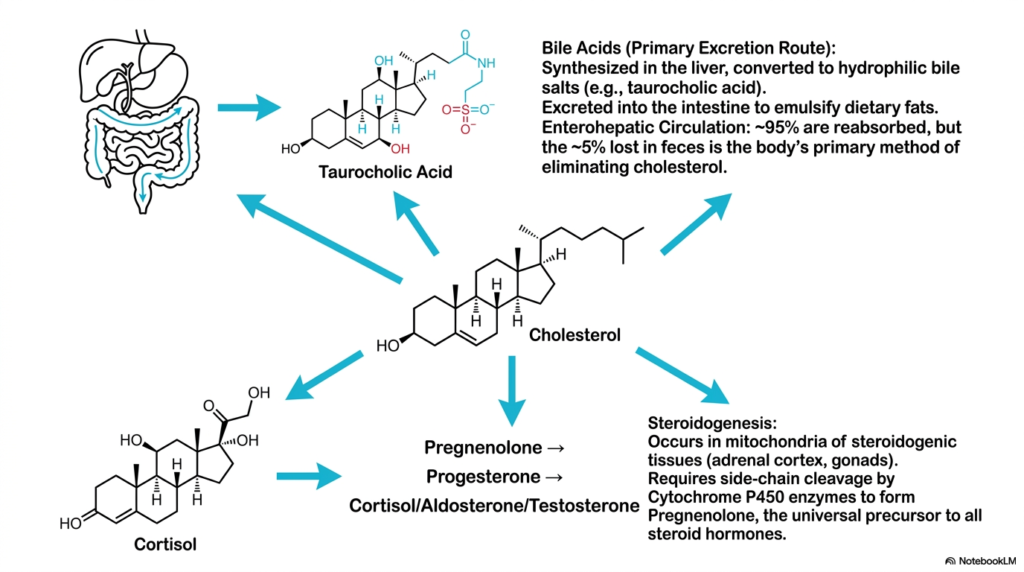
While significant modern medical attention is rightfully focused on severe cardiovascular pathology, the vital physiological fates of the specific lipids generated by Cholesterol Biosynthesis must not be overlooked. Slide fourteen cleanly maps out the incredibly intricate biological utilization and complex excretion pathways of this highly essential sterol. The primary biological route for massive systemic lipid elimination is the intense hepatic production of essential bile acids. The human liver actively hydroxylates the dense sterol core, converting it into highly soluble, highly amphipathic bile salts that act as potent biological detergents necessary for digesting complex, massive dietary fats.
The complex physiological handling of the numerous molecules constantly generated by hepatic Cholesterol Biosynthesis features a highly efficient biological recycling system universally known as enterohepatic circulation. Despite immense daily intestinal biliary secretion, the human body aggressively reabsorbs approximately 95% of these highly valuable bile salts directly from the terminal ileum, returning them straight back to the liver for continuous vital reuse. It is strictly the remaining five percent that physically escapes extensive biological reabsorption and is permanently lost in the feces. This incredibly minor elimination represents the absolute body’s primary physiological mechanism for strict lipid disposal.
Beyond simple physical excretion, the rigid molecular products of active Cholesterol Biosynthesis are absolutely essential for driving intense human endocrine function. The detailed slide expertly illustrates the highly fundamental concept of profound steroidogenesis, a highly complex enzymatic process occurring exclusively within the dense mitochondria of the human adrenal cortex and vital gonadal tissues. Specialized vital Cytochrome P450 enzymes meticulously biochemically cleave the aliphatic side chain of the massive sterol to generate pregnenolone. These incredibly diverse biochemical destinies uniquely highlight the universally multifaceted incredible biological importance of this specific massive molecular cellular lipid framework across human physiological systems.
Slide 15: Familial Hypercholesterolemia and Genetic Disruption of Cholesterol Biosynthesis
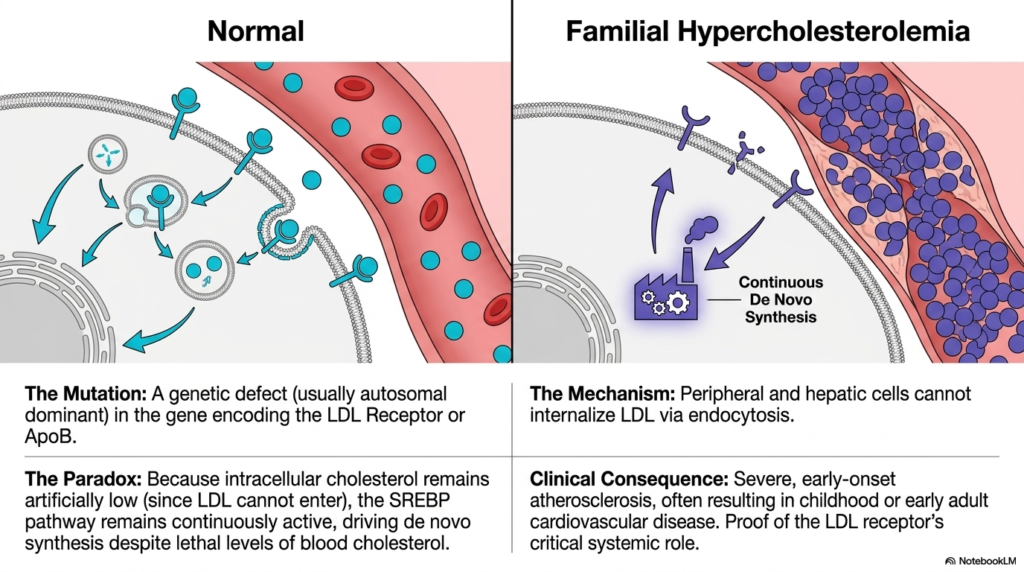
To fully appreciate the vital physiological importance of proper lipid regulation, medical students must examine specific genetic pathologies where the feedback inhibition of normal Cholesterol Biosynthesis completely fails. Slide fifteen clinically details severe Familial Hypercholesterolemia, a highly deadly, incredibly life-threatening genetic disorder distinctly characterized by dangerously massive internal systemic lipid concentrations. This highly devastating condition is most frequently directly caused by a dominant genetic mutation severely affecting either the complex cellular LDL receptor gene or the specific massive structural ApoB surface protein. Consequently, peripheral human cells physically completely lose their fundamental biological ability to properly safely internalize massive circulating LDL particles.
The profound biochemical tragedy of this specific genetic defect lies in the paradoxical continuous activation of widespread hepatic Cholesterol Biosynthesis. Because the incredibly abundant circulating systemic LDL particles physically cannot cross the highly defective cellular plasma membrane to enter the active cytoplasm, the internal cellular environment remains completely starved of essential structural lipids. The internal cellular sensors physically detect a massive, severe physiological lipid deficit and instantly trigger the highly continuous, dangerously unregulated, massive genetic activation of the SREBP cellular pathway. The human liver highly aggressively continues massive unregulated synthesis of lethal structural lipid particles.
The devastating clinical consequences of this complete systemic breakdown in the negative feedback regulation of active Cholesterol Biosynthesis are extremely severe and incredibly rapid. Human patients actively suffering from this highly aggressive genetic mutation frequently violently experience severe, aggressively rapid early-onset atherosclerosis, physically distinctly manifesting as massive intense lipid deposits strictly under the human skin. Without highly extreme modern medical intervention to aggressively artificially block immense endogenous cellular lipid synthesis, uniquely affected individuals uniquely suffer massive, rapidly fatal early myocardial infarctions. This strict pathology clearly illustrates why a fundamentally deep understanding of intricate, complex molecular-cellular metabolic pathways remains absolutely essential.
Please read our Content Disclaimer Statement.
Check out our social media channels: