47. Fatty Acid Synthesis: Pathways and Mechanisms
Have you ever wondered how the human body effortlessly converts that extra slice of pizza into long-term energy reserves? The secret lies deep within our cells, governed by an elegant and highly regulated metabolic engine. This comprehensive slide deck explores the molecular mechanics behind lipid production, breaking down the specific enzymes, chemical reactions, and regulatory switches that drive this vital biological pathway. Designed for advanced students, this guide decodes the structural and functional nuances of lipid metabolism.
Slide 1: The Assembly Line of Fatty Acid Synthesis

The complex process of cellular metabolism relies heavily on highly coordinated enzymatic pathways to construct essential macromolecules. The introductory slide of this presentation illustrates the foundational assembly line necessary for Fatty Acid Synthesis. This visual model conceptualizes the cellular machinery as an intricate engine, demonstrating how simple metabolic precursors are systematically linked together to form high-energy storage molecules. The detailed schematic introduces the principal components required for this anabolic pathway, providing students with a holistic view of the biological construction process. Such foundational knowledge is indispensable for any advanced study of lipid biochemistry.
A primary requirement for Fatty Acid Synthesis is the continual supply of the two-carbon building block known as Acetyl-CoA. Once this substrate is introduced into the system, it interacts with the Acyl Carrier Protein, which acts as a central scaffold moving the growing lipid chain between different active sites. The assembly line features several distinct enzymatic functions, including a condensing enzyme, a ketoacyl reductase, a dehydratase, and an enoyl reductase. Each domain performs a highly specific chemical modification, advancing the structural complexity of the developing molecule.
The ultimate objective of this specific iteration of Fatty Acid Synthesis is the production of palmitate, a fully saturated sixteen-carbon lipid molecule. The repetitive elongation cycles continue seamlessly until a specialized thioesterase enzyme cleaves the finalized lipid from the machinery. Understanding this magnificent biochemical assembly line provides medical and university scholars with profound insights into human energy storage. By mastering these intricate steps, the scientific community can better comprehend metabolic health, cellular energy management, and the physiological consequences of metabolic dysfunction.
Slide 2: Cellular Geography of Fatty Acid Synthesis
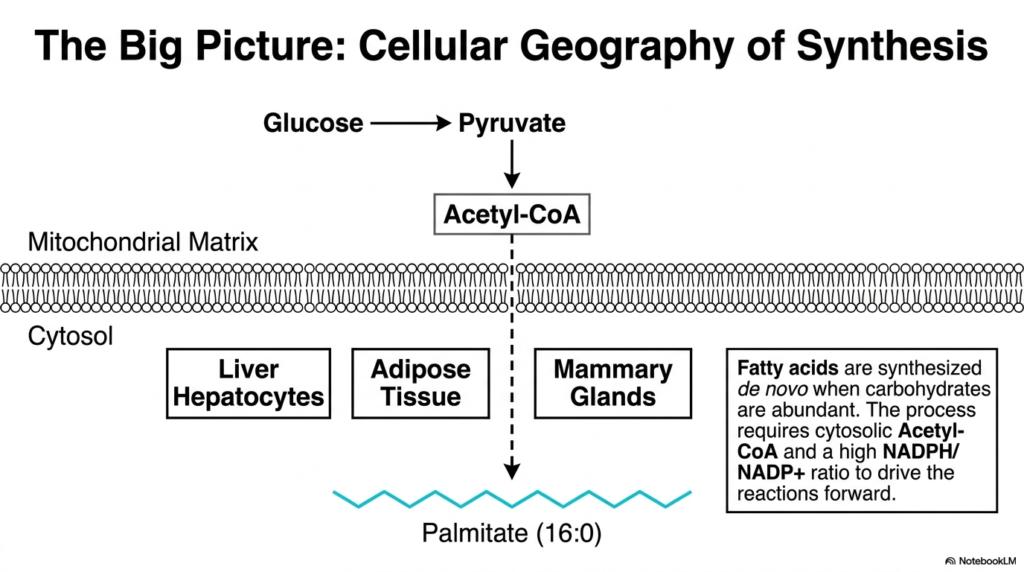
Understanding where biochemical reactions occur is just as important as knowing the reactions themselves. The cellular geography of Fatty Acid Synthesis requires specific environments to proceed efficiently. This process is highly active in distinct tissues, most notably the liver hepatocytes, adipose tissue, and mammary glands. When carbohydrates are abundant, glucose is converted into pyruvate, which then enters the mitochondria to be transformed into Acetyl-CoA. However, the lipogenic machinery resides outside the mitochondria, necessitating specialized transport mechanisms.
The physical separation of these cellular compartments means that Fatty Acid Synthesis must strictly occur within the cytosol of the cell. This specific localization prevents newly formed lipids from immediately undergoing degradation through mitochondrial oxidation pathways. Furthermore, the cytosolic environment provides the necessary biochemical conditions, specifically a high ratio of NADPH to NADP+. This significant concentration of reducing equivalents acts as the vital chemical driving force, pushing the energy-intensive anabolic reactions forward to successfully build long hydrocarbon chains.
Ultimately, the geographic compartmentalization of Fatty Acid Synthesis ensures that energy storage only happens under optimal physiological conditions. When systemic glucose levels are high, cells prioritize the de novo synthesis of palmitate from cytosolic Acetyl-CoA. Medical students must recognize this spatial organization, as it highlights the evolutionary brilliance of cellular design. By segregating opposing metabolic pathways, the cell maintains strict thermodynamic control, ensuring that lipid construction and lipid breakdown do not occur in the exact same cellular space at the same time.
Slide 3: The Nomenclature in Fatty Acid Synthesis
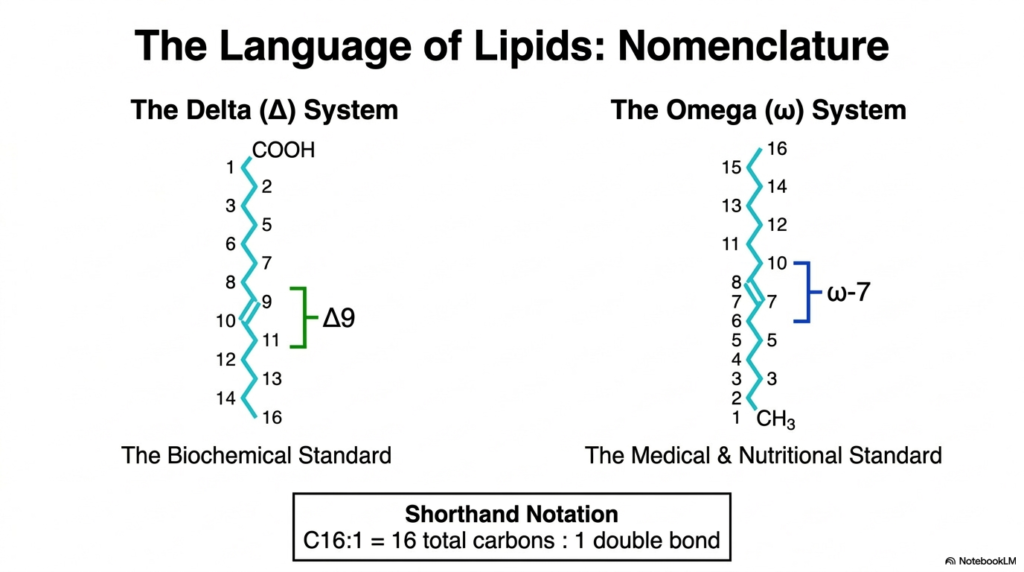
The scientific language used to describe lipid structures is crucial for precise communication in the biomedical field. When discussing the products of Fatty Acid Synthesis, researchers utilize two primary naming conventions: the Delta system and the Omega system. The Delta system represents the strict biochemical standard, beginning its carbon numbering directly at the carboxyl-terminal carbon. This method allows biochemists to precisely map the exact location of every double bond relative to the functional acidic end of the newly synthesized molecule.
Conversely, the Omega system is widely favored in nutritional and medical settings, shifting the perspective to the opposite end of the lipid chain. Rather than starting at the carboxyl group, this standard counts from the terminal methyl group, known as the omega carbon. While Fatty Acid Synthesis primarily generates fully saturated chains, subsequent modifications can introduce double bonds. Identifying these bonds from the methyl end helps clinicians easily classify structurally related dietary lipids, such as omega-3 and omega-6 essential lipids, which play massive roles in systemic inflammation.
Both naming systems are unified by a universal shorthand notation that streamlines the identification of lipids generated through Fatty Acid Synthesis. This standardized notation lists the total number of carbon atoms, followed by a colon, and then the total number of double bonds present in the hydrocarbon chain. For example, a sixteen-carbon chain with one double bond is efficiently written as C16:1. Mastering this structural nomenclature empowers university scholars to effortlessly navigate complex biochemical literature and interpret sophisticated nutritional data with high clinical accuracy.
Slide 4: Crossing the Boundary for Fatty Acid Synthesis
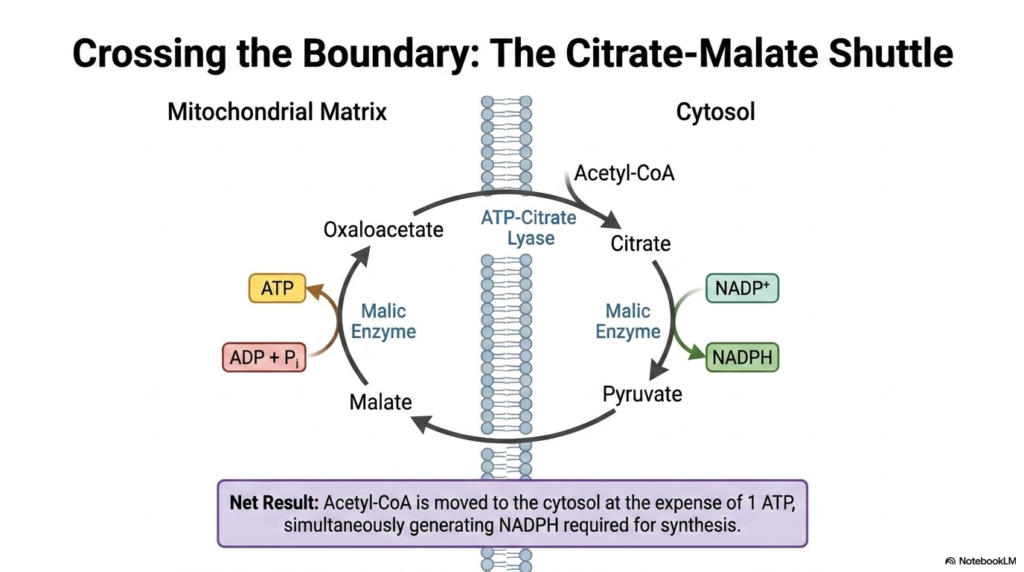
One of the greatest logistical challenges in cellular metabolism is moving vital substrates across impermeable membranes. The primary precursor for Fatty Acid Synthesis is Acetyl-CoA, which is continuously generated deep within the mitochondrial matrix. Unfortunately, the inner mitochondrial membrane is completely impermeable to this critical molecule. To bypass this barrier, the cell employs a brilliant biochemical workaround known as the Citrate-Malate Shuttle, effectively disguising the substrate to allow it safe passage into the cytosol.
To initiate this crucial phase of Fatty Acid Synthesis, Acetyl-CoA is fused with oxaloacetate to form citrate. Unlike its precursor, citrate easily crosses the mitochondrial boundary into the cytosol. Once it reaches its destination, an enzyme known as ATP-Citrate Lyase expends cellular energy to cleave the molecule, regenerating Acetyl-CoA in the exact location where lipogenic enzymes reside. This targeted transportation system ensures that the cytosolic machinery has a continuous, localized supply of two-carbon building blocks required for rapid lipid construction.
Furthermore, the Citrate-Malate Shuttle provides an exceptional secondary benefit essential for Fatty Acid Synthesis. As the remaining malate molecule is recycled back into pyruvate, the malic enzyme simultaneously generates a molecule of NADPH. Because the lipogenic pathway requires massive amounts of reducing power to saturate the growing hydrocarbon chains, this coupled generation of NADPH is an elegant metabolic synergy. The cell seamlessly solves its substrate transportation problem while simultaneously funding the energetic cost of future biochemical reductions.
Slide 5: The Rate-Limiting Step of Fatty Acid Synthesis
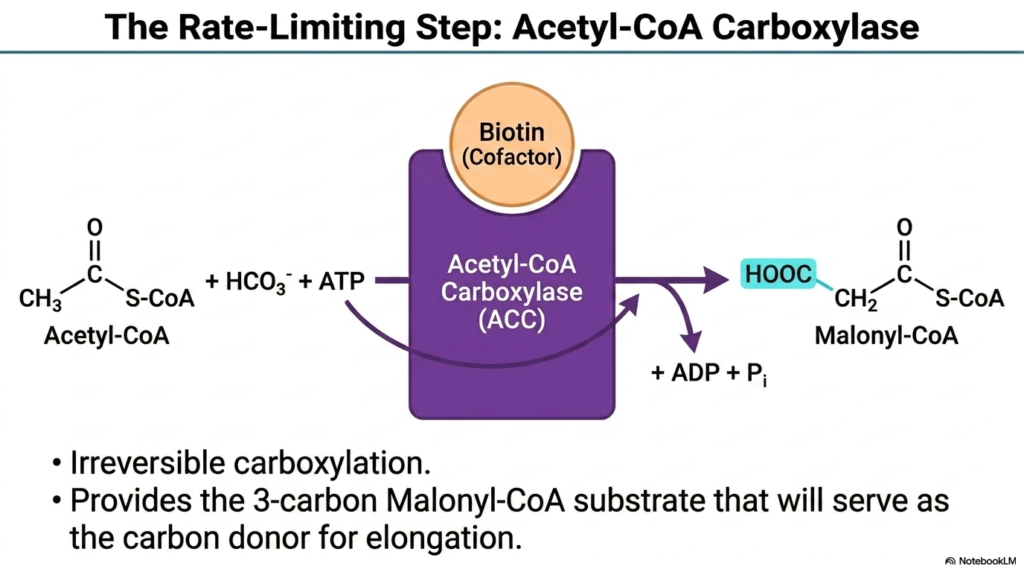
In every major metabolic pathway, there is a singular, highly regulated bottleneck that controls the overall speed of production. The undisputed rate-limiting step of Fatty Acid Synthesis is governed by the enzyme Acetyl-CoA Carboxylase. This massive enzymatic complex acts as the metabolic gatekeeper, dictating exactly when and how rapidly the cell commits its resources to storing energy. Because this step requires the direct consumption of ATP, the cell heavily guards this reaction to prevent unnecessary energy waste during times of starvation.
The primary function of Acetyl-CoA Carboxylase in Fatty Acid Synthesis is to perform an irreversible carboxylation reaction. Using a covalently bound biotin cofactor, the enzyme captures a bicarbonate molecule and forcefully attaches it to the two-carbon Acetyl-CoA substrate. This energy-dependent fusion creates Malonyl-CoA, a structurally critical three-carbon molecule. The irreversible nature of this biochemical event ensures that once the cell commits to lipogenesis, metabolic flux continues in a single, forward direction toward lipid assembly.
The creation of Malonyl-CoA is undeniably the most important chemical commitment in Fatty Acid Synthesis. This newly formed three-carbon molecule will serve as the exclusive carbon donor for all subsequent elongation cycles. By deeply understanding the mechanism of Acetyl-CoA Carboxylase, medical students can grasp how pharmacological interventions might target this exact step to combat metabolic disorders. Inhibiting this specific enzyme halts the entire downstream lipogenic pathway, demonstrating the immense biological significance of this singular regulatory chokepoint.
Slide 6: The Architect of Fatty Acid Synthesis
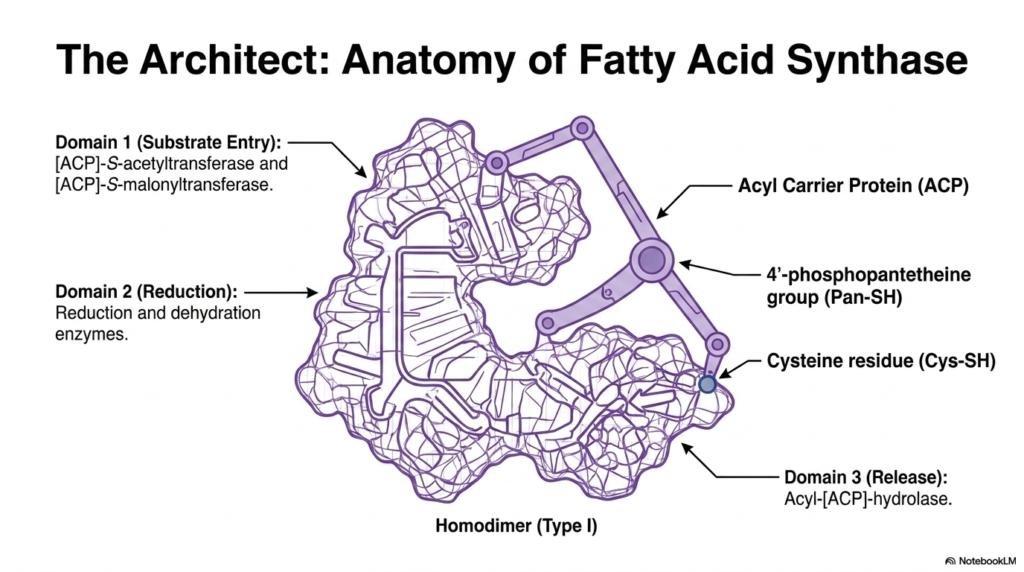
To build complex biological molecules, the cell relies on sophisticated molecular machines with highly specialized architecture. The central engine driving Fatty Acid Synthesis in human cells is a massive, multifunctional enzyme complex known as Fatty Acid Synthase. In mammals, this structure functions as a Type I homodimer, consisting of two identical polypeptide chains that fold together into a highly interactive functional unit. This contiguous design ensures that intermediate molecules never diffuse away during the construction process.
The architecture of the enzyme responsible for Fatty Acid Synthesis is divided into three distinct functional domains. Domain 1 serves as the primary portal for substrate entry, using specific transferases to accept incoming building blocks. Domain 2 houses the critical reduction and dehydration enzymes necessary to modify the growing hydrocarbon chain. Finally, Domain 3 acts as the release mechanism, utilizing a specific hydrolase to detach the completed lipid product. This spatial organization maximizes the catalytic efficiency of the entire lipogenic pathway.
At the very heart of Fatty Acid Synthesis lies the Acyl Carrier Protein, which functions as the flexible robotic arm of the complex. Equipped with a long 4′-phosphopantetheine group, this arm physically carries the growing lipid chain between the different catalytic domains. Working in tandem with a specific cysteine residue, the Acyl Carrier Protein ensures that the highly reactive intermediates are securely tethered to the enzyme complex at all times, preventing unwanted side reactions in the bustling cytosolic environment.
Slide 7: Priming the Machine for Fatty Acid Synthesis
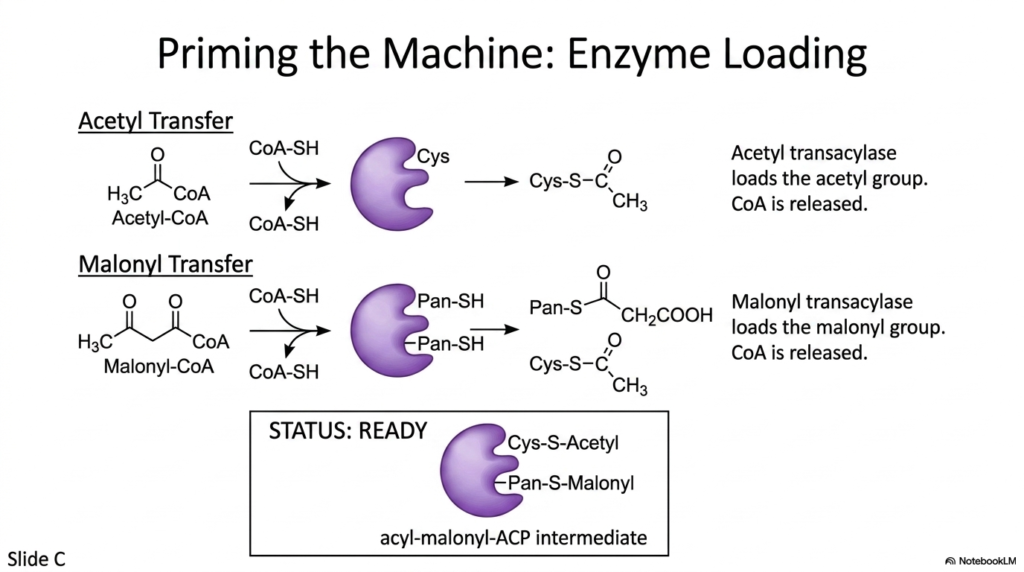
Before a complex biochemical engine can begin its manufacturing cycle, it must be properly loaded with its initial raw materials. The priming phase of Fatty Acid Synthesis involves precisely attaching the starting substrates to their respective docking stations on the enzyme complex. This vital preparatory step ensures that the catalytic domains possess the exact chemical precursors required to initiate the very first carbon-carbon bond formation. Without this highly specific loading mechanism, the lipogenic machinery would remain completely inert.
The first step in priming the machinery for Fatty Acid Synthesis is the transfer of the initial two-carbon starting unit. An enzyme known as acetyl transacylase carefully moves the acetyl group from its Coenzyme A carrier directly onto the highly reactive Cys-SH residue of the enzyme complex. Following this, a separate malonyl transacylase loads the three-carbon malonyl group onto the flexible Pan-SH arm of the Acyl Carrier Protein. During both transfers, the original Coenzyme A carriers are released back into the cellular pool.
Once these specific transfers are complete, the machinery required for Fatty Acid Synthesis is fully primed. The enzyme now holds an acetyl group on its stationary cysteine residue and a malonyl group on its flexible, swinging arm, creating a highly reactive acyl-malonyl-ACP intermediate. This structural alignment perfectly positions the two distinct chemical groups for the impending condensation reaction. Medical students studying this pathway must recognize that this meticulous substrate preparation is the fundamental prerequisite for all subsequent elongation cycles.
Slide 8: Condensation Reaction in Fatty Acid Synthesis
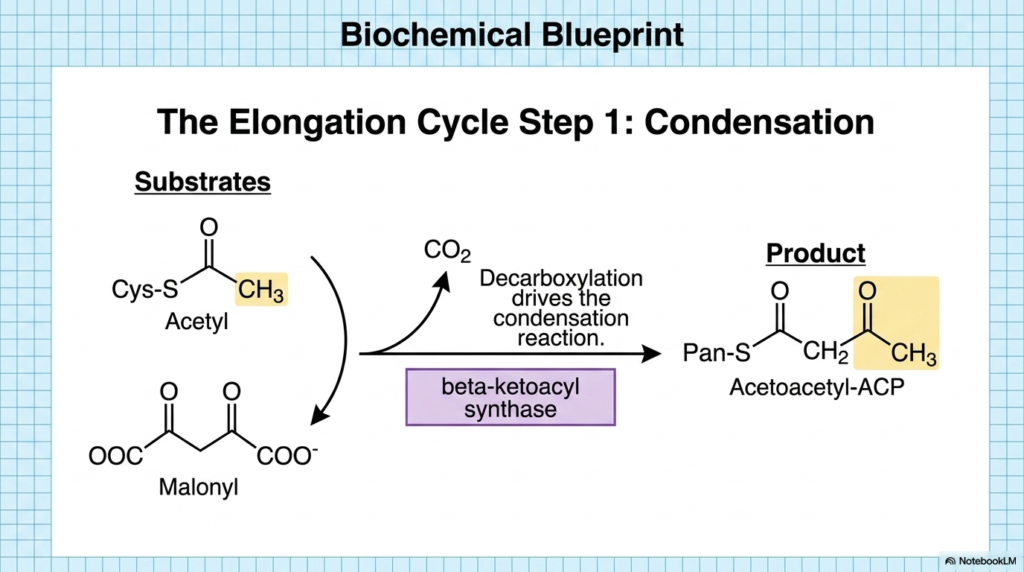
With the molecular machinery fully primed and loaded, the actual assembly of the hydrocarbon chain can commence. The first official step of the elongation cycle in Fatty Acid Synthesis is the condensation reaction. This critical biochemical event is catalyzed by the enzyme beta-ketoacyl synthase, which orchestrates the physical fusion of the two separate carbon substrates. This step marks the defining moment of biosynthesis, in which smaller precursor molecules are permanently linked to form a larger, more complex structural backbone.
The chemical brilliance of this step in Fatty Acid Synthesis lies in its thermodynamic driving force. The three-carbon malonyl group undergoes a rapid decarboxylation, releasing a molecule of carbon dioxide. The energy released by breaking this bond provides the exact thermodynamic driving force to drive the condensation reaction forward. The acetyl group on the stationary cysteine residue is forcefully transferred and linked to the remaining two carbons of the malonyl group, bypassing the energetic barriers that usually prevent such bonds.
The direct result of this elegant condensation mechanism during Fatty Acid Synthesis is the formation of a four-carbon intermediate known as Acetoacetyl-ACP. While the carbon chain has successfully grown, it still bears a highly reactive, oxidized beta-keto group that must be stabilized. Grasping the mechanism of this decarboxylation-driven fusion helps students understand why the cell expends ATP to create Malonyl-CoA in the first place. The temporary addition of carbon dioxide acts purely as an energetic spring, forcing the building blocks together.
Slide 9: The First Reduction Step of Fatty Acid Synthesis
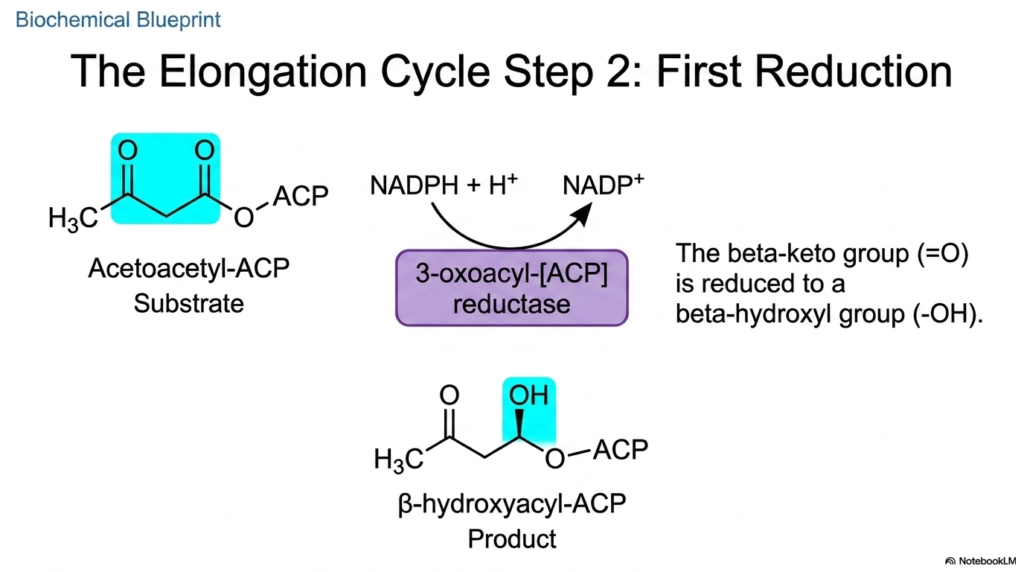
Following the successful fusion of the carbon substrates, the newly formed molecule remains chemically unstable for long-term biological storage. The second step of the elongation cycle in Fatty Acid Synthesis involves an essential reduction reaction to modify the highly reactive beta-keto group. The primary goal of this enzymatic phase is to systematically remove oxygen-containing functional groups from the carbon backbone, thereby pushing the molecule closer to its final, energy-dense, highly reduced lipid state.
To accomplish this critical modification in Fatty Acid Synthesis, the enzyme 3-oxoacyl-[ACP] reductase takes the central stage. This specific domain utilizes NADPH, transferring a hydride ion and a proton to chemically reduce the target beta-keto double bond. The high-energy electrons from NADPH are forcefully incorporated into the developing lipid structure. This specific use of reducing power underscores why the cytosolic environment requires a high ratio of NADPH to NADP+ to sustain anabolic processes.
The chemical product generated by this specific phase of Fatty Acid Synthesis is beta-hydroxyacyl-ACP. The previously oxidized beta-keto group has been successfully converted into a more stable beta-hydroxyl group. This precise molecular alteration prepares the growing hydrocarbon chain for the subsequent dehydration step. For scholars of biochemistry, understanding this continuous electron-transfer process illuminates how the energy originally captured from sunlight and dietary carbohydrates is gradually stored in the dense carbon-carbon bonds of human lipid stores.
Slide 10: Dehydration Step in Fatty Acid Synthesis
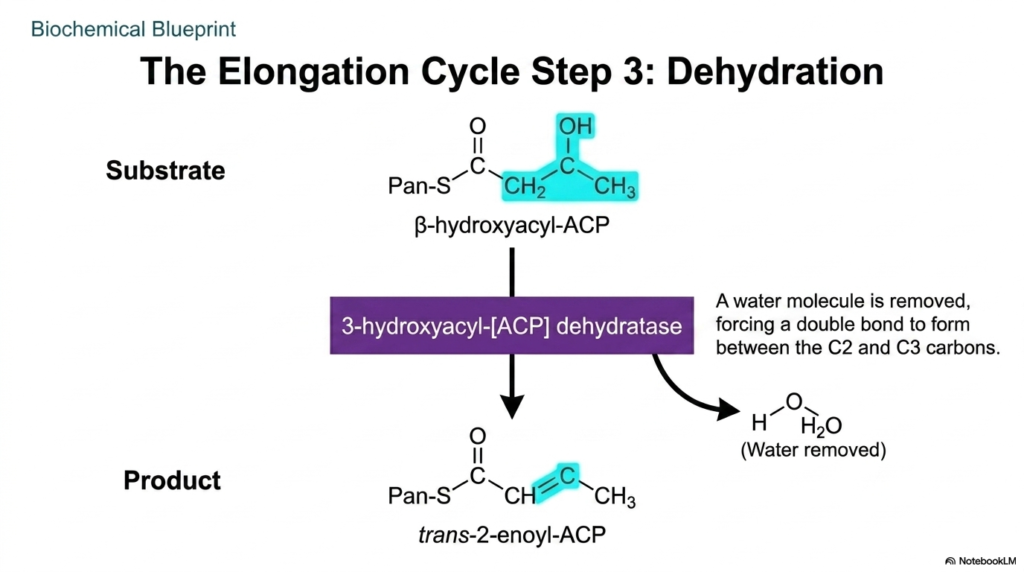
Biological molecules must undergo continuous chemical refinement to achieve their functional anatomical structures. The third sequential step within the elongation cycle of Fatty Acid Synthesis is the dehydration reaction. Having just converted the beta-keto group into a hydroxyl group, the enzyme complex now works to completely strip this remaining oxygen atom from the developing carbon backbone. This structural pruning is absolutely mandatory to create the dense, hydrophobic hydrocarbon tails characteristic of mature, stable cellular lipids.
The specialized enzyme that drives this reaction in Fatty Acid Synthesis is known as 3-hydroxyacyl-[ACP] dehydratase. This catalytic domain precisely targets the beta-hydroxyl group, forcing the complete removal of a water molecule from the growing substrate. By extracting the hydroxyl group from one carbon and a hydrogen atom from the adjacent carbon, the enzyme chemically forces a new carbon-carbon double bond to form. This structural shift effectively eliminates the bulky oxygen atom from the lipid architecture.
The immediate biochemical result of this dehydration phase in Fatty Acid Synthesis is the creation of a trans-2-enoyl-ACP intermediate. Although the molecule has successfully shed its water, the newly formed double bond still leaves the structure slightly unsaturated and kinked. This intermediate state perfectly sets the stage for the final chemical reduction of the cycle. Medical professionals studying lipid pathways note that this systematic removal of oxygen is what ultimately grants fatty acids their incredible caloric density compared to heavily oxygenated carbohydrates.
Slide 11: The Second Reduction of Fatty Acid Synthesis
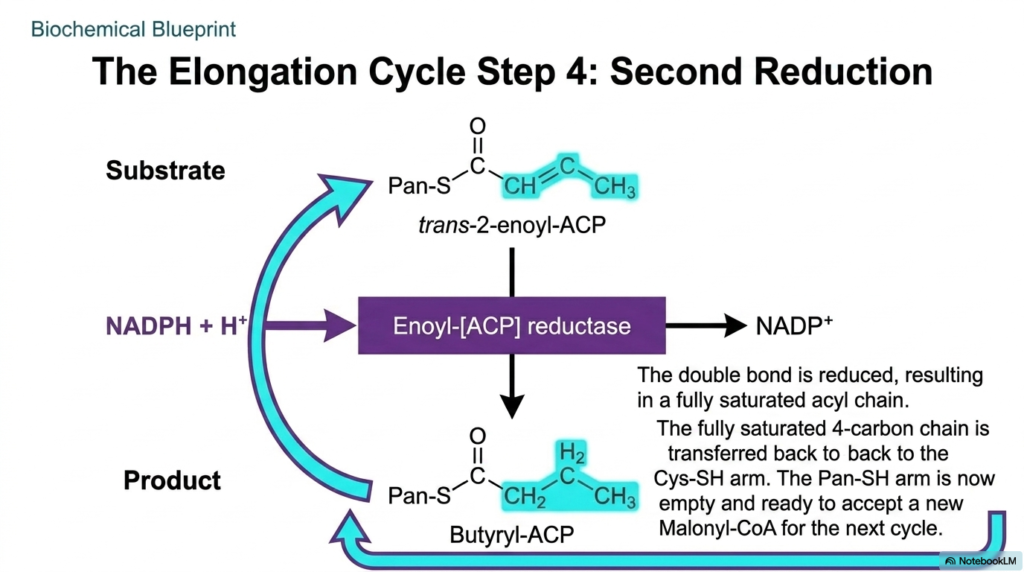
To finalize the structural integrity of the newly lengthened carbon chain, the enzyme complex must perform one last chemical modification. The fourth and final step of the elongation cycle in Fatty Acid Synthesis is the second reduction reaction. The primary objective here is to eliminate the temporary double bond created during the previous dehydration phase. By fully saturating the carbon backbone with hydrogen atoms, the cell guarantees that the developing lipid can pack tightly into energy-dense storage droplets.
This critical saturating reaction within Fatty Acid Synthesis is strictly governed by the enoyl-[ACP] reductase domain. Similar to the first reduction step, this enzyme consumes another molecule of NADPH, transferring its high-energy electrons and protons directly across the trans-double bond. This targeted chemical bombardment successfully breaks the double bond, yielding a fully saturated, straight-chain acyl group. The continuous expenditure of cellular reducing power at this stage highlights the immense energetic investment required for comprehensive lipid biosynthesis.
The completion of this step in Fatty Acid Synthesis yields Butyryl-ACP, a fully saturated four-carbon lipid chain attached to the flexible robotic arm. To prepare for the next round of elongation, this four-carbon chain is physically shifted back onto the stationary Cys-SH residue. This elegant molecular maneuver completely frees the Pan-SH arm, allowing it to accept a brand-new Malonyl-CoA molecule. The cycle is now completely reset, perfectly positioned to continue growing the lipid by another two carbons.
Slide 12: Cycle Completion in Fatty Acid Synthesis
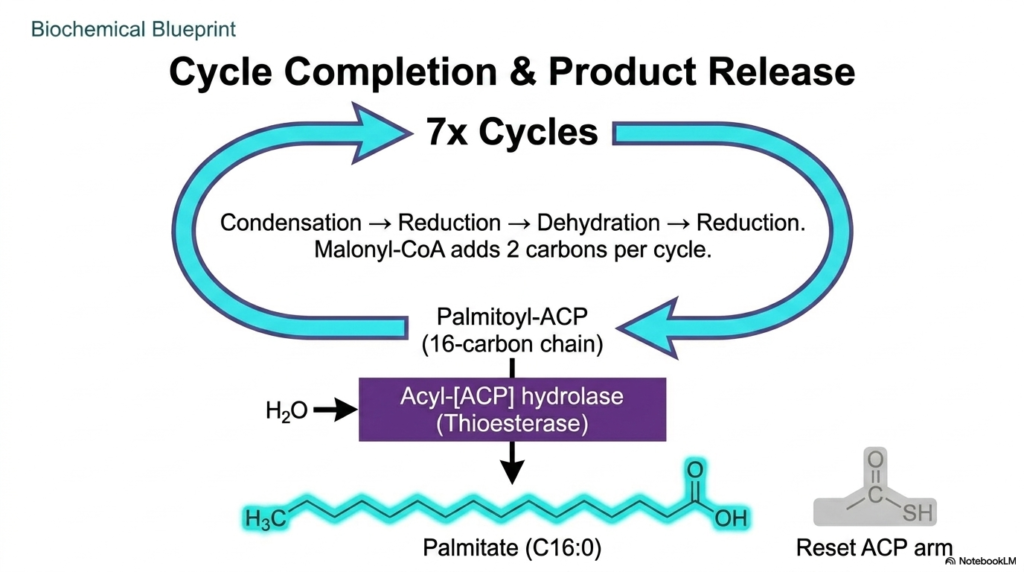
Biochemical manufacturing requires repeated cycles to produce substantial macromolecular structures. The incredible machinery responsible for Fatty Acid Synthesis does not simply stop after creating a four-carbon chain; it continuously cycles through elongation. The sequence of condensation, reduction, dehydration, and a final reduction is repeated over and over. With every single rotation of this metabolic wheel, the malonyl-CoA substrate successfully donates exactly two new carbon atoms to the rapidly growing, highly hydrophobic lipid backbone.
To achieve the ultimate standard product of Fatty Acid Synthesis, the enzyme complex must execute exactly seven complete cycles. This repetitive biochemical hammering systematically lengthens the chain until it reaches the critical threshold of a sixteen-carbon structure, known chemically as palmitoyl-ACP. The cellular machinery is highly sensitive to this specific molecular length, as the internal cavity of the enzyme complex is precisely calibrated to accommodate a hydrocarbon chain of exactly sixteen carbons before structural constraints halt further elongation.
Once the sixteen-carbon threshold in Fatty Acid Synthesis is reached, the cycle-completion phase relies on a specialized termination domain. The acyl-[ACP] hydrolase, commonly referred to as a thioesterase, introduces a water molecule to forcefully cleave the completed lipid from the carrier protein. This specific hydrolytic event officially releases free Palmitate into the cellular environment while simultaneously resetting the entire enzyme complex for a completely new manufacturing run. This tightly controlled termination ensures remarkable product consistency across human metabolism.
Slide 13: The Global Stoichiometry of Fatty Acid Synthesis

Every cellular operation requires a strict biochemical accounting of its metabolic inputs and outputs. The global stoichiometry of Fatty Acid Synthesis provides a comprehensive mathematical receipt of the exact resources required to construct a single lipid molecule. To successfully synthesize one complete molecule of palmitate, the human cell must commit eight molecules of Acetyl-CoA, seven molecules of ATP, and fourteen molecules of NADPH. This massive energetic investment unequivocally demonstrates why lipogenesis only occurs during states of profound caloric abundance.
Analyzing the metabolic receipt of Fatty Acid Synthesis reveals exactly how these resources are distributed. Of the eight required Acetyl-CoA molecules, only one is utilized directly to prime the enzyme, while the remaining seven are converted into malonyl-CoA. The energetic cost of seven ATP is exclusively consumed by Acetyl-CoA Carboxylase to facilitate this critical conversion. This data highlights that the primary energy expenditure in lipid construction is front-loaded into the preparation of carbon substrates rather than into the elongation cycles.
Furthermore, the reducing power required for Fatty Acid Synthesis is immense, requiring exactly fourteen molecules of NADPH. Because each of the seven elongation cycles necessitates two reduction steps, the mathematical correlation is perfectly clear. As the final palmitate molecule is released, the cell also generates multiple byproducts, including water, ADP, and free Coenzyme A. Mastering this stoichiometric balance allows advanced university students to fully appreciate the complex thermodynamic economy operating within the boundaries of human lipid metabolism.
Slide 14: Expanding Beyond Palmitate in Fatty Acid Synthesis
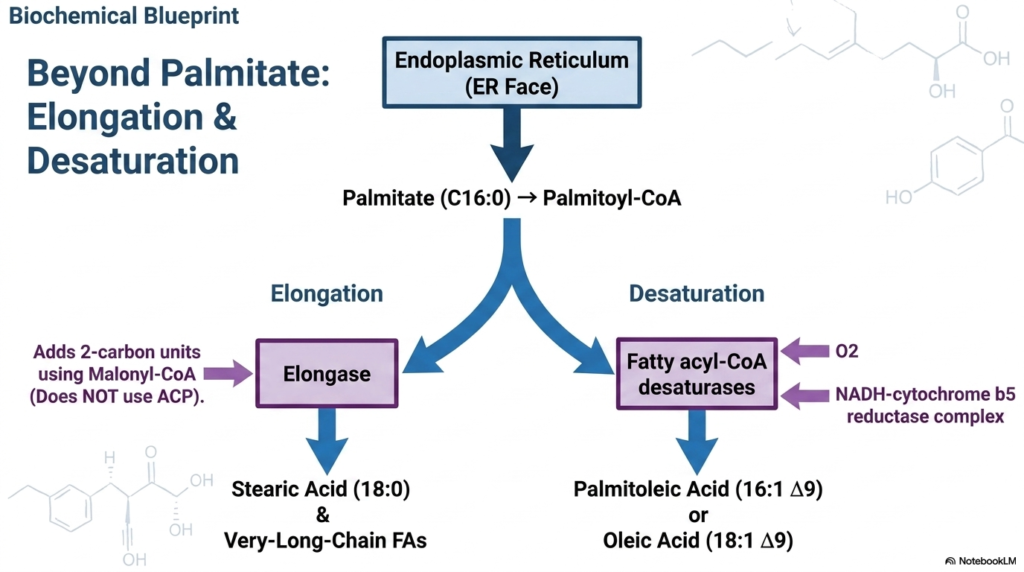
While the primary cytosolic machinery is highly efficient, it produces a strictly limited structural output. The standard product of de novo Fatty Acid Synthesis is exclusively the sixteen-carbon, fully saturated palmitate. However, the human body requires a vast array of significantly longer and chemically varied lipid structures to maintain complex biological functions. To expand upon this basic sixteen-carbon foundation, the newly minted palmitate must immediately interact with entirely different enzyme systems anchored to the cytoplasmic face of the Endoplasmic Reticulum.
To increase the length of the lipid chain after foundational Fatty Acid Synthesis, specialized enzymes known as elongases take control. Unlike the massive cytosolic synthase complex, these discrete elongases add two-carbon units using Malonyl-CoA but strictly operate without the assistance of an Acyl Carrier Protein. This post-synthetic elongation pathway efficiently modifies the original palmitate into stearic acid, an eighteen-carbon lipid, and can continue functioning to generate the vital very-long-chain fatty acids required for specific neurological tissues.
Simultaneously, the cell utilizes specialized fatty acyl-CoA desaturases to introduce structural kinks into the straight chains produced by Fatty Acid Synthesis. Relying on an intricate NADH-cytochrome b5 reductase complex and molecular oxygen, these desaturases chemically strip hydrogen atoms to create highly specific cis-double bonds. This precisely controlled desaturation mechanism transforms rigid palmitate into palmitoleic acid, or stearic acid into oleic acid, providing the vital membrane fluidity necessary for proper cellular dynamics and complex physiological signaling.
Slide 15: Mammalian Limits After Fatty Acid Synthesis

Despite possessing robust molecular machinery, human biochemistry has distinct evolutionary limitations. While mammals excel at foundational Fatty Acid Synthesis and subsequent desaturation, our cellular enzymes are restricted in where they can physically introduce double bonds. The human desaturase zone is strictly limited to the first nine carbons of a lipid chain, starting from the carboxyl end. Beyond carbon 9, mammals entirely lack the specific enzymatic capabilities required to create new points of unsaturation in the hydrocarbon tail.
This strict biochemical limitation following Fatty Acid Synthesis directly establishes the physiological requirement for essential fatty acids. Because human cells cannot modify the distal ends of lipid chains, any polyunsaturated fats featuring double bonds beyond the ninth carbon must be exogenously acquired. Consequently, the human diet must reliably provide specific plant-based lipids, as the cellular machinery is physically incapable of assembling these highly complex, multi-double-bonded structures from scratch, regardless of the availability of foundational metabolic precursors.
The inability to fully modify the products of Fatty Acid Synthesis elevates linoleate and linolenate to critical nutritional status. These exogenously sourced essential lipids are the absolute, mandatory precursors for generating potent biological signaling molecules known as eicosanoids. Without consuming these specific dietary fats, humans cannot synthesize vital prostaglandins or leukotrienes, leading to severe inflammatory and immune system dysfunctions. Understanding this mammalian enzymatic boundary perfectly bridges the gap between complex molecular biochemistry and daily clinical nutrition.
Slide 16: Master Switches Regulating Fatty Acid Synthesis
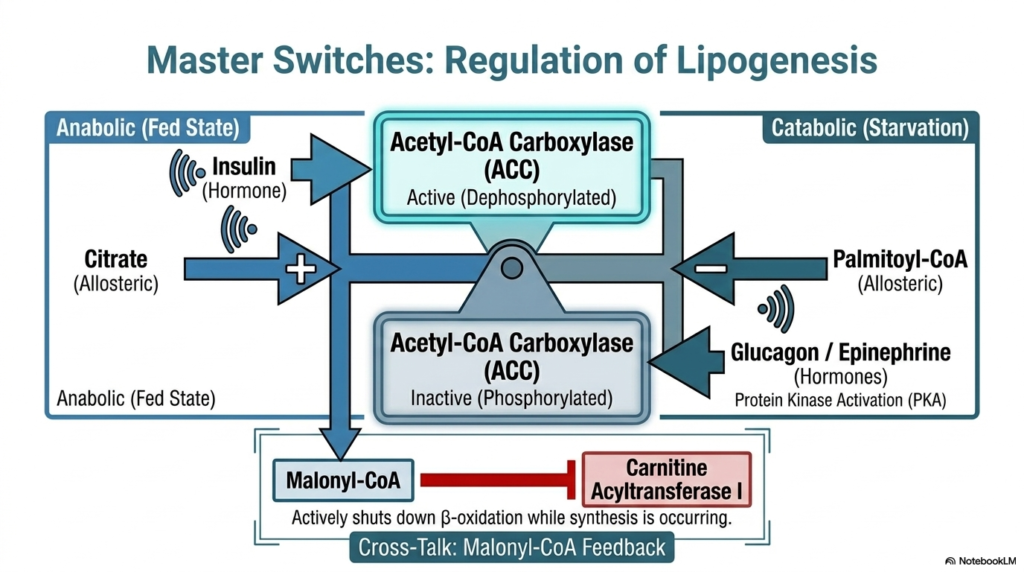
Metabolic pathways must be rigidly controlled to prevent cellular chaos and catastrophic energy waste. The entire cascade of Fatty Acid Synthesis is tightly regulated by powerful allosteric and hormonal master switches. The undisputed central control node for this system is Acetyl-CoA Carboxylase (ACC). In a well-fed, anabolic state, the hormone insulin signals the cell to permanently dephosphorylate ACC. This critical modification forces the enzyme into its highly active state, rapidly accelerating the production of malonyl-CoA to store excess dietary calories.
Conversely, during periods of starvation, the regulation of Fatty Acid Synthesis shifts entirely. Catabolic hormones such as glucagon and epinephrine activate protein kinases, which phosphorylate ACC, thereby shutting down the entire lipogenic pathway. Furthermore, the system utilizes brilliant allosteric feedback loops. High cytosolic concentrations of citrate physically bind to ACC to stimulate lipid construction, while a buildup of the final product, palmitoyl-CoA, directly inhibits the enzyme, ensuring the cell naturally halts production when sufficient energy stores are achieved.
Perhaps the most elegant regulatory crosstalk in Fatty Acid Synthesis involves the newly synthesized malonyl-CoA. As lipogenesis ramps up, the rising concentration of malonyl-CoA actively binds to and inhibits Carnitine Acyltransferase I, the specific enzyme responsible for shuttling lipids into the mitochondria for beta-oxidation. This brilliant chemical feedback mechanism acts as an absolute cellular fail-safe. It ensures that newly synthesized lipid chains are not immediately drawn back into the mitochondrial furnace, thereby separating anabolic synthesis from catabolic degradation.
Slide 17: The Ancient Pathway of Mitochondrial Fatty Acid Synthesis
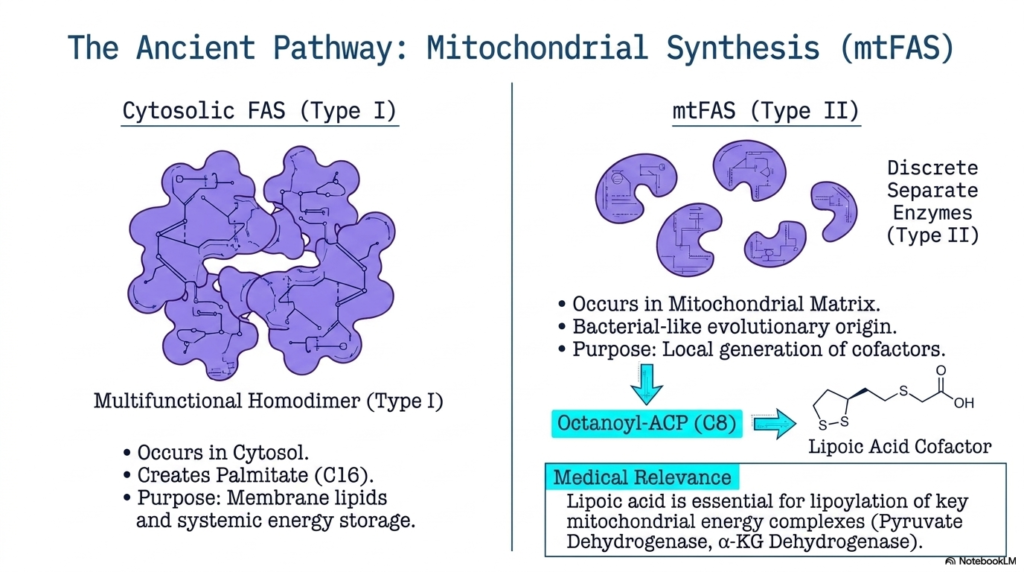
While the vast majority of mammalian lipid construction occurs in the cytosol, a fascinating, secondary system operates deep within the mitochondria. This highly specialized pathway, known as mitochondrial Fatty Acid Synthesis (mtFAS), differs drastically from its cytosolic counterpart. Instead of using a massive, single Type I homodimer, this archaic system relies on a Type II architecture composed of entirely discrete enzymes. This fragmented structural organization strongly mirrors bacterial metabolism, hinting at the profound evolutionary origins of the mitochondrial organelle.
The purpose of this ancient form of Fatty Acid Synthesis is not to generate massive amounts of energy-dense palmitate for systemic storage. Instead, mtFAS operates on a much smaller, highly localized scale. The primary biochemical objective of this secondary pathway is the dedicated generation of octanoyl-ACP, a highly specific eight-carbon intermediate. The mitochondrial machinery intentionally halts elongation at this precise length to provide the exact chemical precursor necessary for the local synthesis of lipoic acid within the matrix.
This alternative variation of Fatty Acid Synthesis holds immense medical relevance for human cellular function. The locally generated lipoic acid is an essential cofactor for several critical mitochondrial energy complexes, most notably Pyruvate Dehydrogenase and Alpha-Ketoglutarate Dehydrogenase. Without the successful operation of this discrete, ancient Type II pathway, the cell would lose its ability to link glycolysis to the citric acid cycle, resulting in catastrophic failure of global cellular respiration and metabolic collapse.
Slide 18: Comparing Fatty Acid Synthesis and Beta-Oxidation
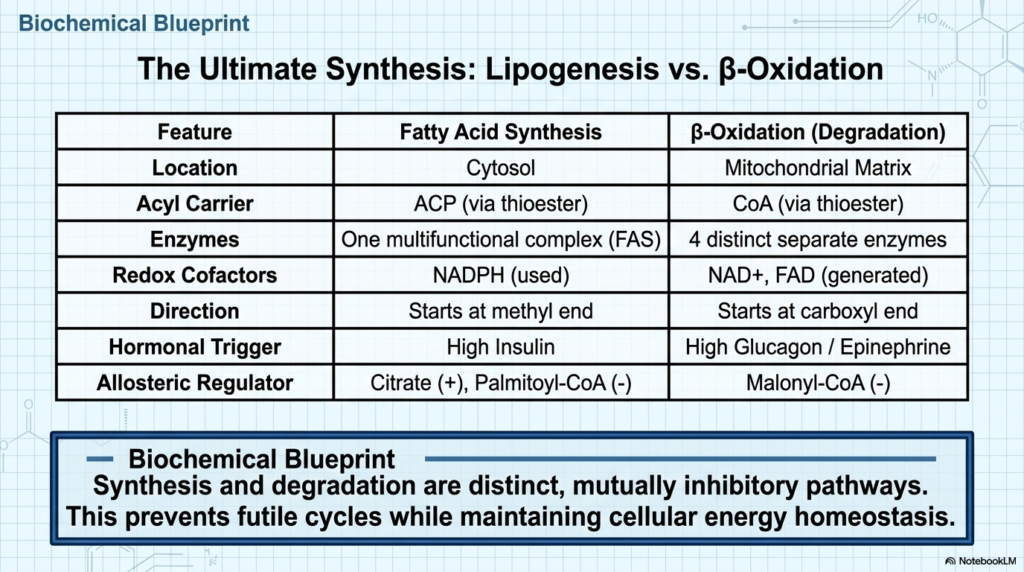
The final slide of this educational series summarizes the ultimate balance between cellular construction and cellular destruction. Comparing the intricate pathway of Fatty Acid Synthesis to its exact metabolic opposite, beta-oxidation, reveals a perfect dichotomy in biological design. Lipogenesis represents the ultimate anabolic state, actively consuming cellular energy to link carbon units into massive macromolecules. Conversely, beta-oxidation represents the ultimate catabolic breakdown, systematically tearing those carbon bonds apart to release trapped energy during periods of systemic starvation.
The physiological differences between Fatty Acid Synthesis and lipid breakdown are stark and highly compartmentalized. While lipogenesis occurs exclusively in the cytosol, using numerous multifunctional enzymes, beta-oxidation is strictly confined to the mitochondrial matrix, using discrete enzymatic units. Furthermore, the construction phase relies on NADPH as its primary electron donor, whereas the oxidative breakdown phase strips electrons to generate NADH and FADH2, which directly fuel the mitochondrial electron transport chain to produce large quantities of ATP.
Understanding the dynamic interplay between Fatty Acid Synthesis and beta-oxidation is the cornerstone of metabolic biochemistry. The human cell must constantly evaluate its environment, utilizing sophisticated hormonal signals and sensitive allosteric feedback loops to seamlessly toggle between these two fundamentally opposed pathways. By ensuring that these processes never operate simultaneously, the biological system maximizes its thermodynamic efficiency. This precise balance ensures that the body can safely store energy during times of plenty and effortlessly mobilize it during severe famine.
Please read our Content Disclaimer Statement.
Check out our social media channels: