48. Biochemistry of Lipogenesis and Complex Lipid Synthesis
Ever wonder how an extra slice of pizza transforms into long-term cellular energy? The answer lies in the elegant and highly regulated pathways of lipid biosynthesis. This comprehensive guide breaks down the complex biochemistry behind fat creation, offering a visually driven exploration of structural biology, enzymatic assembly lines, and hormonal oversight. Designed for medical and biochemistry students, this presentation demystifies the molecular mechanisms that sustain life, build cellular membranes, and manage metabolic energy flux.
Slide 1: Introduction to Lipogenesis and Complex Lipid Biosynthesis
Welcome to the fascinating world of cellular metabolism, with a focus on the biosynthesis of complex lipids. This visual journey explores the overarching mechanisms by which biological systems convert raw nutrients into crucial structural and energetic molecules. At the absolute core of this transformation lies lipogenesis, a highly orchestrated and magnificent sequence of biochemical reactions. The initial slide sets the stage for an in-depth exploration of pathways, structural biology, and the metabolic regulation of fats. Understanding these foundations is essential for any medical or university student stepping into the realm of biochemistry.
When examining the fundamental nature of lipid creation, it becomes immediately apparent how essential these molecules are for sustaining human life. The structural diagram presented highlights the intricate chemical nature of phospholipids, which feature hydrophilic heads and hydrophobic tails. As the study of lipogenesis unfolds throughout this module, learners will discover exactly how simple two-carbon precursors are intricately assembled into these complex structures. The precise coordination required to build such molecules from scratch beautifully showcases the evolutionary elegance of human cellular machinery.
Why does this metabolic mastery matter in a clinical or academic context? Beyond mere energy storage, the end-products synthesized during lipogenesis construct the very plasma membranes that define every single cell’s boundaries. A deep dive into these pathways reveals not just how the human body stores excess caloric intake, but how cellular compartments maintain their architectural integrity and communicate. The continuous interplay among dietary intake, hormonal signaling, and genetic expression converges directly on these critical lipid assembly lines.
To fully master the upcoming content, one should pay close attention to the recurring biochemical themes of precursor activation, metabolic branch points, and allosteric feedback loops. The subsequent slides will thoroughly dissect the specific enzymes, cofactors, and substrates involved in every distinct phase of lipogenesis. By breaking down these complex pathways into logical, digestible steps, the overarching logic of metabolic regulation becomes incredibly clear. Prepare to transition from a macroscopic view of cellular biology into the dynamic world of lipid assembly.
Slide 2: Defining Lipogenesis: Cellular Compartmentalization and Flow
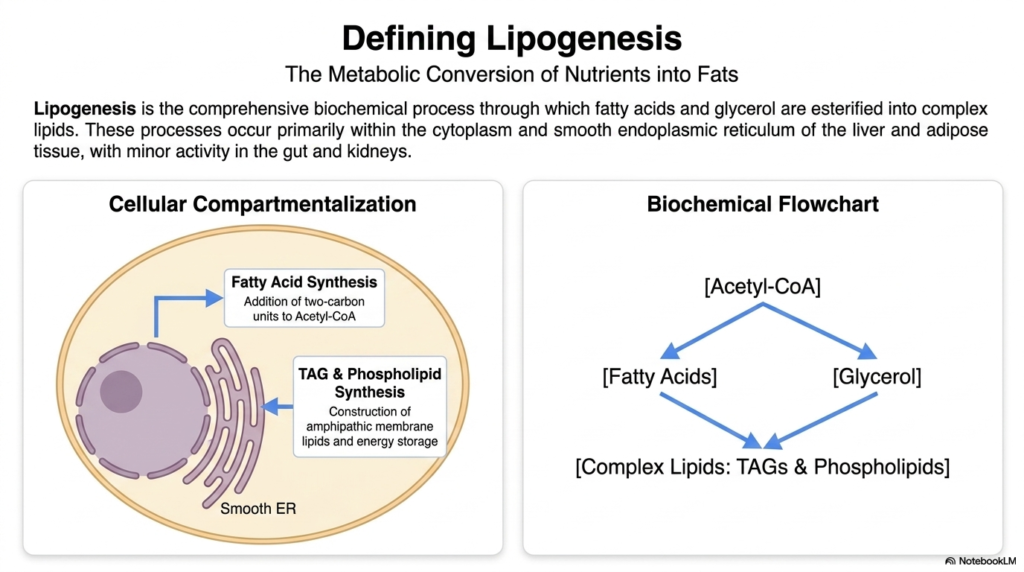
To understand fat synthesis, one must first clearly define the parameters and location of the process. Lipogenesis is the comprehensive biochemical phenomenon through which free fatty acids and glycerol are esterified to construct complex lipids. The provided slide perfectly illustrates the cellular compartmentalization of these events, showing their primary occurrence within the cytoplasm and the smooth endoplasmic reticulum. While the liver and adipose tissues act as the primary anatomical hubs for these reactions, minor activities also occur in the intestines and kidneys.
The biochemical flowchart presented here strips the complexity down to its most fundamental components. It demonstrates how a simple molecule such as Acetyl-CoA serves as the absolute starting material for the entire system. From Acetyl-CoA, the pathway diverges to supply the necessary fatty acid chains. Simultaneously, the cell must generate a glycerol backbone. This dual-pronged approach is the defining characteristic of lipogenesis, as both the fatty acid tails and the glycerol backbone must be synthesized independently before they can be joined.
Visualizing the smooth endoplasmic reticulum’s role is crucial for medical students, as this organelle is the specialized factory for the synthesis of triacylglycerol (TAG) and phospholipids. The construction of amphipathic membrane lipids requires a highly specific environment separate from the general aqueous cytoplasm. When lipogenesis is actively occurring, the smooth ER enzymes work tirelessly to attach the newly minted fatty acids to the glycerol framework. This compartmentalization prevents unwanted side reactions and ensures that the highly hydrophobic products are handled correctly by the cell.
Ultimately, the metabolic conversion of nutrients into complex fats represents a massive energetic investment by the cell. It takes continuous energy and reducing equivalents to fuel the demanding machinery of lipogenesis. Understanding this fundamental biochemical flowchart—from Acetyl-CoA down to complex TAGs and phospholipids—provides the necessary scaffolding for the entire topic. As we progress, we will examine precisely how each arrow in this simplified flowchart is driven by specific, highly regulated enzymatic catalysts.
Slide 3: Acquiring the Backbone for Lipogenesis: Glycerol 3-Phosphate
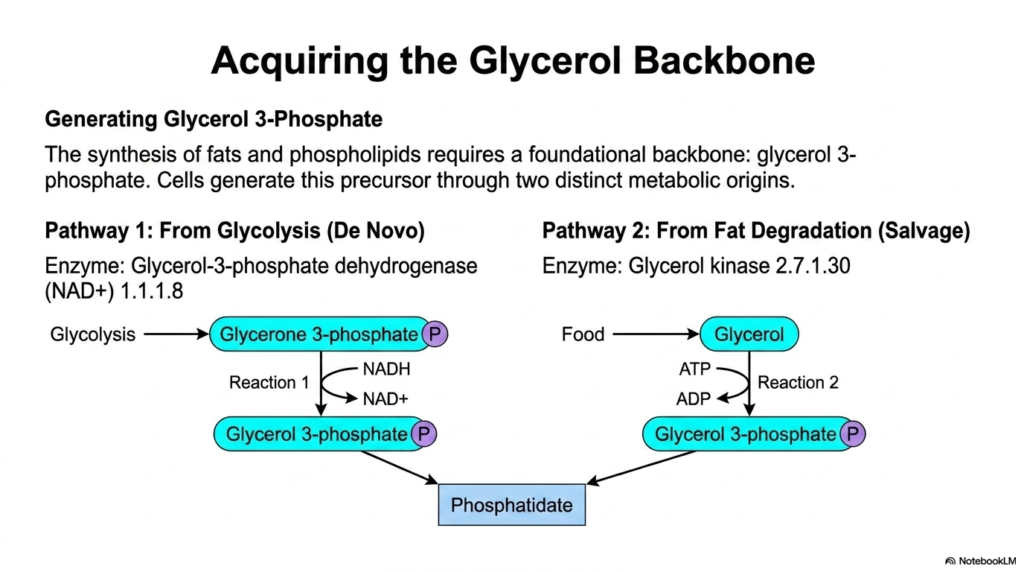
Before a cell can build a complex fat, it needs a strong structural foundation. The synthesis of both neutral fats and active phospholipids strictly requires a foundational backbone, glycerol 3-phosphate. The presentation brilliantly outlines how human cells generate this vital precursor through two highly distinct metabolic origins. Without this essential phosphorylated sugar derivative, the entire machinery of lipogenesis would instantly grind to a halt, making the acquisition of this backbone a primary and indispensable metabolic priority.
The first critical route is the de novo pathway, directly linking lipid synthesis to carbohydrate metabolism. As glucose is broken down during glycolysis, an intermediate called glycerone 3-phosphate (also known as DHAP) is produced. The crucial enzyme glycerol-3-phosphate dehydrogenase utilizes NADH to reduce this intermediate directly into glycerol 3-phosphate. This powerful reaction inextricably links the rate of lipogenesis to the availability of dietary glucose, explaining why high-carbohydrate diets so readily translate into robust fat synthesis and energy storage.
Alternatively, cells can utilize the salvage pathway to recycle materials derived from fat degradation or direct dietary intake. In this second route, free glycerol obtained from digested food is phosphorylated directly. The enzyme glycerol kinase uses a molecule of ATP to attach a phosphate group, immediately yielding glycerol 3-phosphate. This bypasses glycolysis entirely, ensuring the necessary backbone remains readily available even if direct glucose metabolism fluctuates. This dual-pathway redundancy ensures a constant supply of raw materials.
By mastering these two distinct origins, students gain a deeper appreciation for metabolic flexibility. The convergence of glycolysis and fat salvage onto a single molecule—glycerol 3-phosphate—highlights the brilliant interconnectivity of human biochemistry. As the foundational backbone flows downstream, it prepares to accept fatty acid chains in the subsequent steps of lipogenesis. This initial acquisition phase is the quiet but absolutely essential setup required to construct every single membrane lipid and fat droplet in the body.
Slide 4: Phosphatidate: The Universal Precursor in Lipogenesis
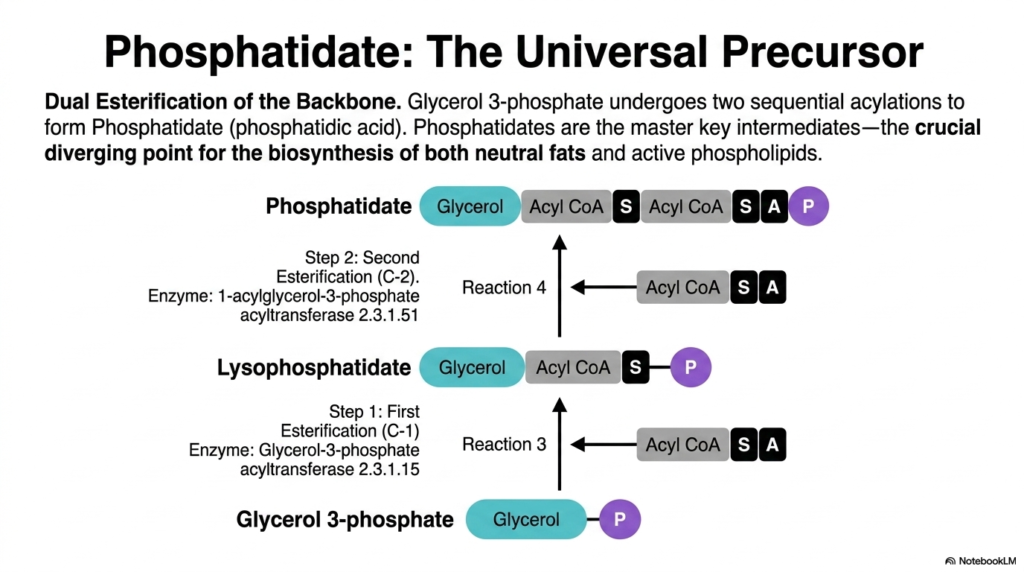
With the glycerol 3-phosphate backbone successfully synthesized, the true assembly line of fat creation begins. This stage involves the dual esterification of the backbone to form a molecule called phosphatidate (phosphatidic acid). Phosphatidate serves as the ultimate master-key intermediate in lipogenesis. It serves as the single crucial point of divergence in the biosynthesis of both energy-storing neutral fats and structurally vital phospholipids. Understanding this specific molecule is absolutely paramount for any aspiring biochemist.
The first biochemical step requires attaching a single fatty acid chain to the first carbon (C-1) of the glycerol backbone. The enzyme glycerol-3-phosphate acyltransferase carefully facilitates this reaction, utilizing an activated acyl-CoA molecule. This initial esterification produces lysophosphatidate, an intermediate that now possesses a single hydrophobic tail. This critical addition marks the exact moment when the water-soluble backbone undergoes a definitive transformation into a highly hydrophobic, nonpolar lipid structure characteristic of active lipogenesis.
Following the formation of lysophosphatidate, a second, equally important esterification must occur rapidly. The enzyme 1-acylglycerol-3-phosphate acyltransferase transfers another activated fatty acid from acyl-CoA onto the second carbon (C-2) position. This perfectly executes the dual esterification process, culminating in the birth of the universal precursor, phosphatidate. Structurally, phosphatidate now features a glycerol backbone, two long-chain fatty acyl tails, and a charged phosphate group at the C-3 position, making it incredibly versatile.
Because phosphatidate stands at the biochemical crossroads, it dictates the lipid’s future fate. At this critical juncture, the cell must make a tightly regulated decision about which type of complex lipid is currently required. Will it become a neutral storage fat, or will it become a structural membrane component? The robust activity of the lipogenesis pathway absolutely depends on the steady generation of phosphatidate to supply all downstream lipid branches effectively and efficiently.
Slide 5: The DAG Branch Point in Lipogenesis Pathways
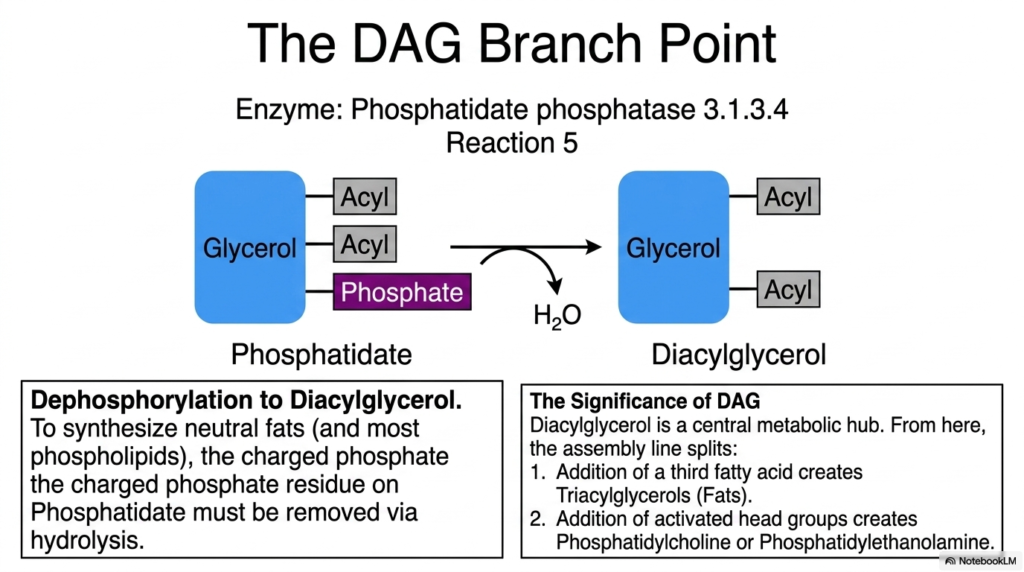
As we navigate deeper into the lipid assembly line, we arrive at the highly significant Diacylglycerol (DAG) branch point. To synthesize the vast majority of neutral fats and predominant phospholipids, the charged phosphate residue on phosphatidate must be completely removed. This vital dephosphorylation is carried out by the enzyme phosphatidate phosphatase, which uses a water molecule to hydrolyze the phosphate bond. This specific reaction drives lipogenesis forward by converting phosphatidate directly into diacylglycerol.
The significance of diacylglycerol cannot be overstated within cellular metabolism. DAG acts as a central metabolic hub where the assembly line formally splits into two major destinies. The removal of the bulky, charged phosphate group renders the molecule significantly more hydrophobic, perfectly preparing the C-3 position for its final modifications. From this specific hub, the cell can seamlessly dictate whether to prioritize long-term energetic storage or the immediate expansion of its complex structural membranes.
If the cellular environment dictates energy conservation, DAG takes the first route. The addition of a third fatty acid chain transforms the DAG into a mature triacylglycerol, commonly known as a neutral fat. This is the classic endpoint most commonly associated with lipogenesis, resulting in the dense lipid droplets that pack our adipose tissue. It represents the ultimate metabolic savings account for future periods of nutrient scarcity.
Conversely, if the cell is actively dividing or needs to repair its boundaries, DAG takes the second prominent route. Instead of adding a third fatty acid, highly activated head groups are carefully attached to the open C-3 position. This alternative pathway yields critical structural phospholipids such as phosphatidylcholine and phosphatidylethanolamine. Therefore, the delicate enzymatic control at the DAG branch point represents a beautiful intersection of structural biology and energy management within the lipogenesis framework.
Slide 6: Triacylglycerols: The Energetic Climax of Lipogenesis
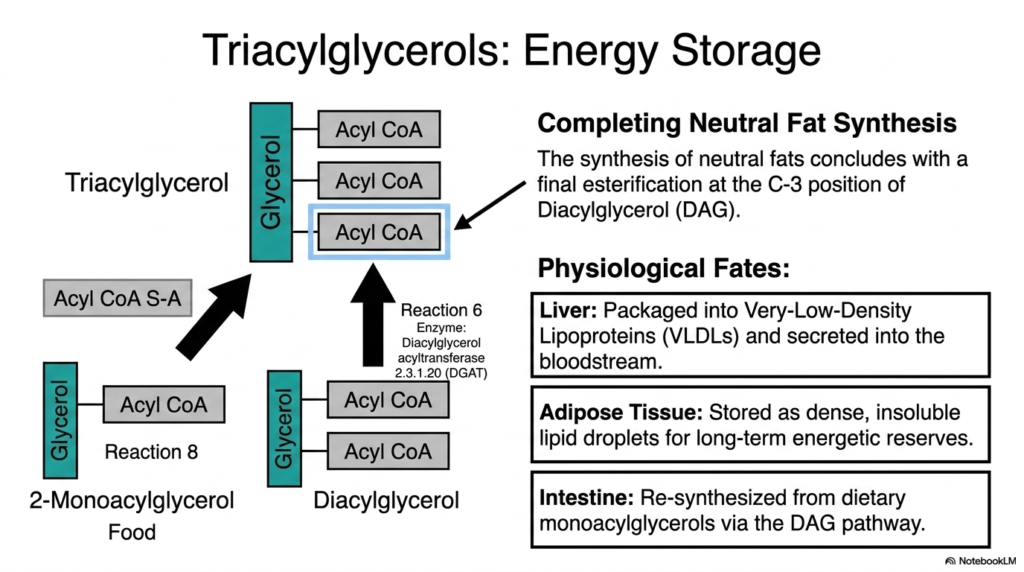
When the cellular mandate is energy storage, the pathway proceeds to synthesize triacylglycerols (TAGs). The synthesis of neutral fats concludes with a decisive final esterification exactly at the C-3 position of diacylglycerol. The key enzyme, diacylglycerol acyltransferase (DGAT), skillfully transfers a final fatty acid from an activated acyl-CoA directly onto the DAG molecule. This specific reaction represents the formal completion of the neutral fat branch of lipogenesis, yielding a fully formed, highly dense triacylglycerol molecule.
The physiological fates of these newly minted triacylglycerols are fascinating and highly dependent on their tissue of origin. In the liver, the primary organ of systemic metabolism, these dense fats cannot simply remain free-floating. Instead, they are meticulously packaged into Very-Low-Density Lipoproteins (VLDLs) and actively secreted into the general bloodstream. This sophisticated transport mechanism ensures that the massive energetic bounty generated by hepatic lipogenesis can be safely distributed to distant tissues throughout the entire human body.
In contrast, when this pathway occurs directly within adipose tissue, the end goals are entirely different. Adipocytes function as specialized metabolic vaults, actively storing the synthesized triacylglycerols as immense, completely insoluble lipid droplets. These magnificent droplets represent the body’s primary long-term energy reserves, capable of sustaining human life during prolonged fasting. The robust capacity of adipose tissue to perform lipogenesis guarantees that excess caloric energy is never wasted but carefully preserved.
Interestingly, the intestinal tract also utilizes a variation of this biochemistry. During the absorption of dietary fats, the intestines re-synthesize triacylglycerols from digested monoacylglycerols via the DAG pathway. This ensures that dietary fats are properly repackaged before entering the lymphatic system. Across these distinct tissues, the synthesis of triacylglycerols is a magnificent evolutionary adaptation, seamlessly converting transient nutritional abundance into stable, life-preserving energy stores.
Slide 7: Phospholipids via the DAG Hub in Lipogenesis
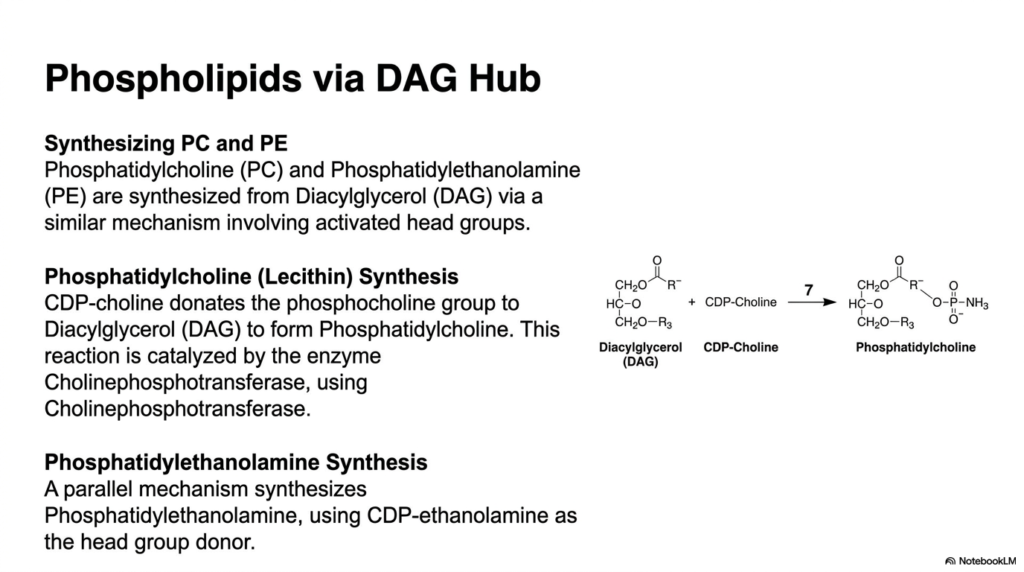
While energy storage is crucial, cells must also relentlessly build and maintain their physical boundaries. Returning to the pivotal diacylglycerol (DAG) hub, we now examine the biosynthesis of the primary structural phospholipids. This branch of lipogenesis is entirely dedicated to membrane architecture. The synthesis of phosphatidylcholine (PC), often called lecithin, and phosphatidylethanolamine (PE) relies on an incredibly fascinating biochemical mechanism where the incoming head groups must be highly energized before they can be attached to the DAG backbone.
To create phosphatidylcholine, the molecule choline must first be activated into CDP-choline. This activated cytidine diphosphate derivative serves as a high-energy donor. The enzyme 1-alkyl-2-acetylglycerolcholine phosphotransferase seamlessly transfers the phosphocholine head group directly onto the open C-3 hydroxyl of diacylglycerol. This critical step in lipogenesis locks the hydrophilic head in place alongside the hydrophobic tails, thereby finalizing the magnificent amphipathic structure that is essential for fluid cellular membranes.
The synthesis of phosphatidylethanolamine follows a striking parallel mechanism. In this parallel track, an activated CDP-ethanolamine complex donates its specific head group to the exact same diacylglycerol hub. This elegant redundancy in the biochemical assembly line allows the cell to rapidly produce different lipid classes using the very same foundational machinery. By simply swapping the activated nucleotide donor, the specific character and surface charge of the resulting membrane lipid are radically altered to meet cellular demands.
Mastering these phospholipid pathways provides medical students with profound insight into membrane biology. Phosphatidylcholine and phosphatidylethanolamine are not just passive barriers; they are dynamic participants in cellular signaling and structural integrity. The intricate enzymatic processes that link these distinct head groups to the DAG hub represent the true structural side of lipogenesis, proving that fat synthesis is just as much about building cellular architecture as it is about storing reserve energy.
Slide 8: Phospholipid Interconversion: Flexibility in Lipogenesis
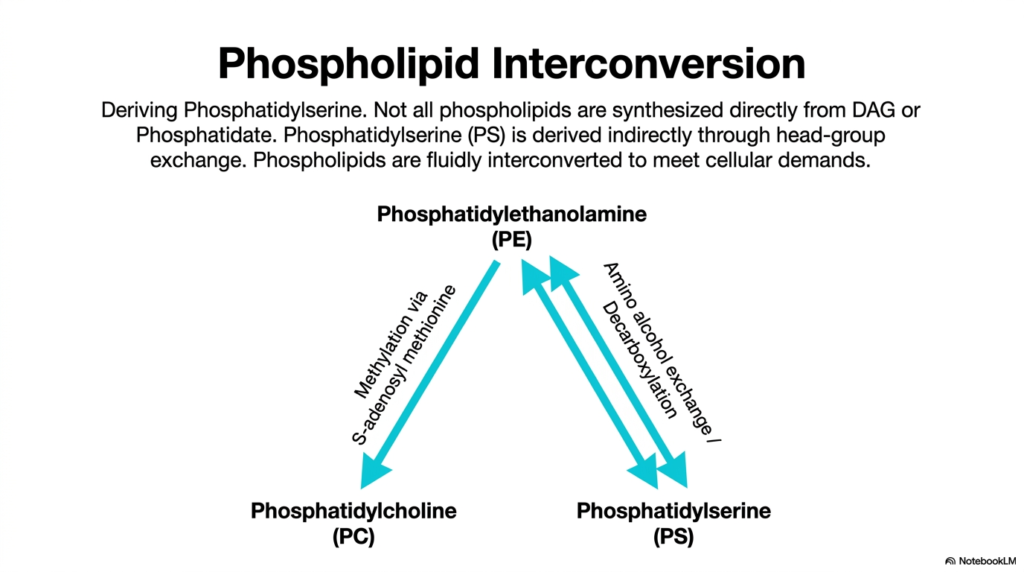
The biochemical flexibility of a cell is heavily dependent on its ability to rapidly adapt to environmental changes. Strikingly, not all major membrane components are synthesized directly from DAG or phosphatidate. Instead, certain crucial lipids like phosphatidylserine (PS) are derived entirely through an indirect process known as head-group exchange. This phenomenon of fluid interconversion represents a sophisticated auxiliary branch of lipogenesis, in which preexisting structural phospholipids are continuously remodeled to meet shifting cellular demands.
The primary mechanism for generating phosphatidylserine involves directly altering a fully formed phosphatidylethanolamine (PE) molecule. Through a highly specific enzymatic exchange reaction, the ethanolamine head group is simply swapped out for the amino acid serine. Additionally, a reverse reaction involving strict decarboxylation allows phosphatidylserine to be converted right back into phosphatidylethanolamine. This elegant, bidirectional loop ensures that the cell can precisely maintain the delicate ratio between these two critical membrane components without constantly synthesizing them from scratch.
Another vital interconversion pathway involves the derivation of phosphatidylcholine directly from phosphatidylethanolamine. This is achieved through a sequential methylation process in which the enzyme uses S-adenosylmethionine (SAM) as the universal methyl donor. Transferring three methyl groups to the ethanolamine head group completely transforms the lipid into phosphatidylcholine. This alternative route highlights the astonishing metabolic versatility inherent in lipogenesis, ensuring that highly specialized lipids can be generated even when direct precursor pathways are temporarily compromised.
Understanding these fluid interconversions is fundamental for grasping the dynamic nature of cellular membranes. Lipids are not static bricks in a wall; they are constantly being synthesized, exchanged, and chemically modified. This continuous remodeling ensures optimal membrane fluidity, proper curvature, and correct protein anchoring. The interconversion pathways beautifully complement the primary lipogenesis assembly lines, providing an essential layer of metabolic plasticity required for the continuous growth and survival of complex human cells.
Slide 9: The Alternate Route in Lipogenesis: CDP-DAG
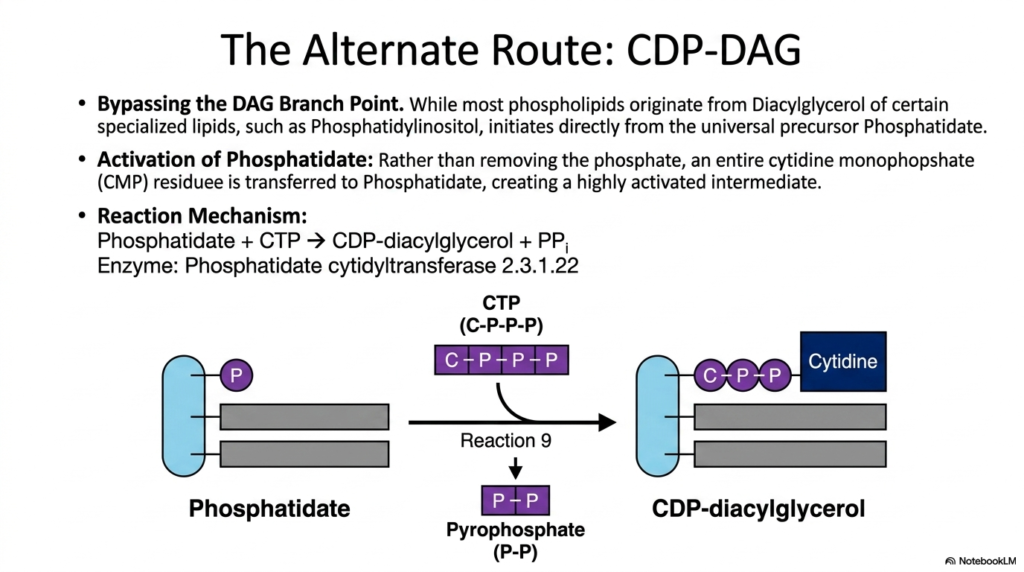
While the diacylglycerol hub serves as the primary gateway for most complex lipids, a fascinating biochemical detour exists. This alternate route entirely bypasses the standard DAG branch point. Instead, specialized lipids are synthesized directly from the universal precursor, phosphatidate. This distinct divergence within lipogenesis is vital for generating highly specific intracellular signaling molecules. Rather than removing the phosphate group from phosphatidate, this alternative pathway seeks to supercharge the entire molecule for specialized construction.
The defining reaction of this alternate route is the intense activation of phosphatidate itself. The enzyme phosphatidate cytidylyltransferase facilitates the transfer of a cytidine monophosphate (CMP) residue directly onto the phosphate group of phosphatidate. This reaction consumes a CTP molecule and releases highly energetic pyrophosphate (PPi). The magnificent result is the creation of CDP-diacylglycerol, a highly activated, energy-rich intermediate that is fully primed to accept specialized head groups in subsequent biochemical reactions.
The creation of CDP-diacylglycerol highlights a brilliant inversion of the standard metabolic strategy. In the primary DAG pathway, the incoming head groups (such as CDP-choline) are activated prior to attachment. However, in this alternative branch of lipogenesis, the massive lipid backbone itself becomes the activated nucleotide carrier. This powerful structural activation provides the necessary thermodynamic driving force to attach complex, bulky, or highly specialized ring structures, like inositol, in the next phase of biosynthesis.
For biochemistry students, distinguishing between the DAG and CDP-DAG pathways is a classic and essential learning objective. These two distinct routes perfectly demonstrate how cells utilize different thermodynamic strategies to achieve diverse structural endpoints. The CDP-DAG intermediate stands as a vital testament to the metabolic complexity of lipogenesis, serving as the exclusive, high-energy launchpad for the synthesis of the critical signaling lipids we will explore on the next slide.
Slide 10: Synthesizing Signaling Lipids via Lipogenesis
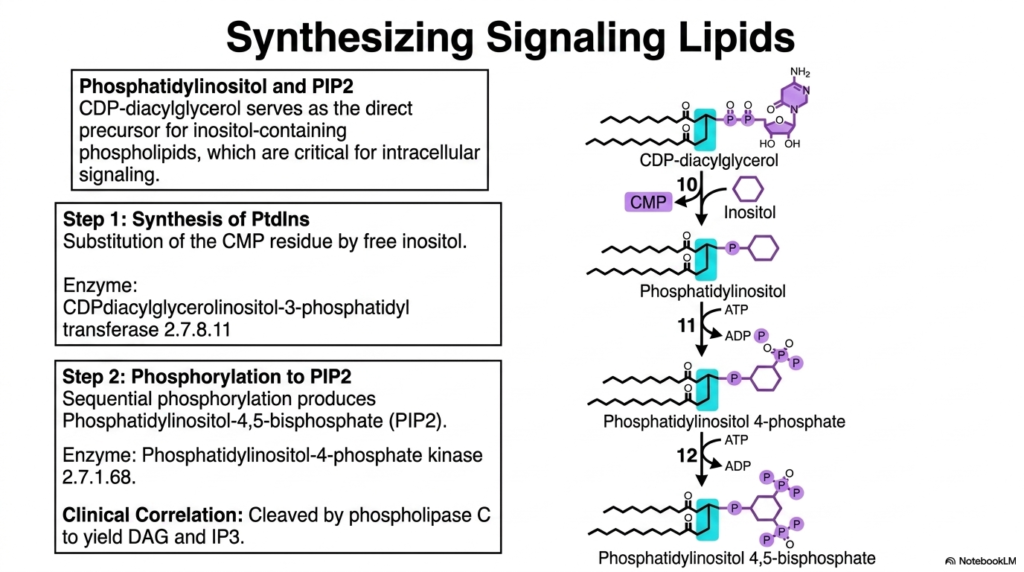
Building on the alternative CDP-DAG route, we now explore the biosynthesis of the incredibly important intracellular signaling molecules. The highly activated CDP-diacylglycerol intermediate serves as the direct and exclusive precursor for inositol-containing phospholipids. These specialized molecules are not merely structural components of the membrane; they are critical functional participants in complex cellular communication cascades. The ultimate goal of this specific branch of lipogenesis is to generate phosphatidylinositol and its heavily phosphorylated derivatives.
The first dedicated step involves the synthesis of phosphatidylinositol (PtdIns). The enzyme CDP-diacylglycerol-inositol 3-phosphatidyltransferase meticulously replaces the entire activating CMP residue with a free inositol sugar alcohol. This reaction releases the CMP and permanently attaches the inositol ring directly to the lipid backbone. This precise step in the lipogenesis pathway produces the fundamental phosphatidylinositol molecule, securely anchoring the massive cyclic sugar directly into the hydrophobic environment of the plasma membrane.
Once phosphatidylinositol is synthesized, it must undergo rapid, sequential phosphorylation to become fully active. Specific kinases, such as phosphatidylinositol-4-phosphate kinase, utilize ATP to selectively add phosphate groups to specific hydroxyls on the inositol ring. This sequential mechanism predictably produces phosphatidylinositol-4,5-bisphosphate, universally known in medical literature as PIP2. PIP2 is a towering biochemical celebrity, functioning as an absolutely essential substrate for countless hormonal and intracellular receptor signaling pathways.
The clinical correlation of PIP2 synthesis is profound and directly relevant to medical students. When specific extracellular hormones bind to the cell, the enzyme phospholipase C rapidly cleaves PIP2. This dramatic cleavage instantly yields two incredibly potent secondary messengers: DAG, which activates protein kinase C, and IP3, which actively triggers massive intracellular calcium release. Therefore, this highly specialized offshoot of lipogenesis is directly responsible for generating the chemical language that allows human cells to respond dynamically to their environment.
Slide 11: Hormonal Regulation of Lipogenesis: Insulin Activation
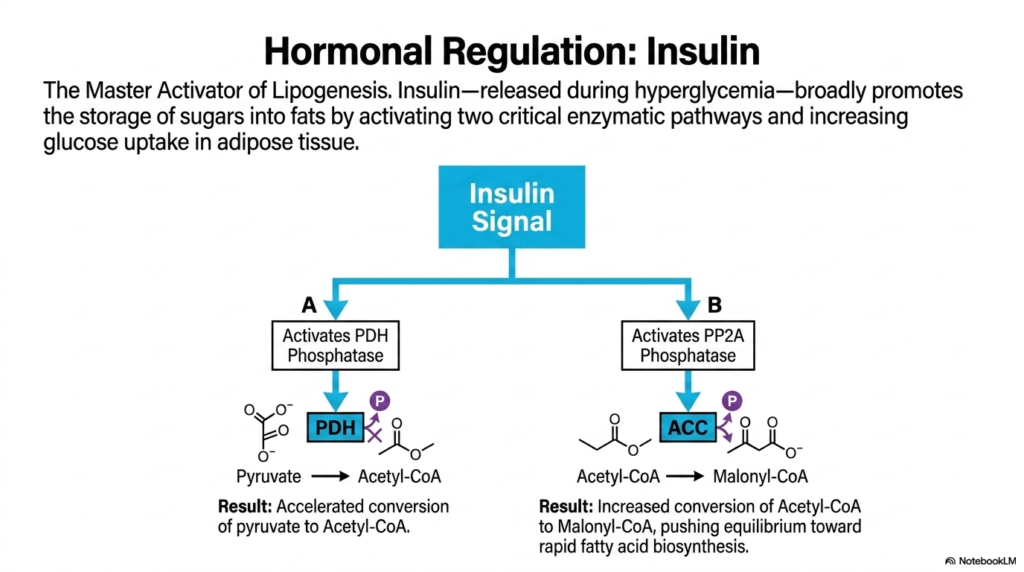
A biological process as energetically expensive as fat synthesis cannot be left unregulated. Hormonal control acts as the ultimate master switch, and insulin is undoubtedly the supreme activator of lipogenesis. Released precisely during states of nutrient abundance and hyperglycemia, insulin broadly and aggressively promotes the conversion of excess sugars into stored fats. It achieves this metabolic dominance by systematically increasing glucose uptake in adipose tissues and forcefully activating two distinctly critical enzymatic pathways within the cell.
The first major target of the insulin signaling cascade is the Pyruvate Dehydrogenase (PDH) complex. Insulin signaling specifically activates PDH phosphatase, an enzyme that subsequently removes an inhibitory phosphate group from the PDH complex itself. This crucial dephosphorylation dramatically accelerates the conversion of glycolysis-derived pyruvate directly into Acetyl-CoA. By aggressively opening this primary metabolic floodgate, insulin ensures a massive, continuous supply of the fundamental building blocks required to sustain high rates of lipogenesis.
Simultaneously, insulin signaling targets a second, equally important bottleneck. It strongly activates PP2A phosphatase, which targets and dephosphorylates Acetyl-CoA Carboxylase (ACC). Dephosphorylated ACC is the highly active form of the enzyme, rapidly catalyzing the vital conversion of Acetyl-CoA into Malonyl-CoA. This specific reaction strongly drives the metabolic equilibrium toward rapid fatty acid biosynthesis. The combined, synergistic activation of PDH and ACC ensures that every available carbon is aggressively funneled into fat production.
Understanding the precise biochemical actions of insulin is absolutely critical to understanding the pathology of metabolic diseases such as diabetes and obesity. By driving dephosphorylation cascades, insulin effectively signals the cellular machinery that energy is abundant and should be conserved for the future. The sheer power of insulin to orchestrate this massive, systemic shift toward lipogenesis perfectly illustrates the beautiful and profound connection between macroscopic dietary intake and microscopic enzymatic regulation.
Slide 12: Hormonal Inhibition of Lipogenesis Pathways
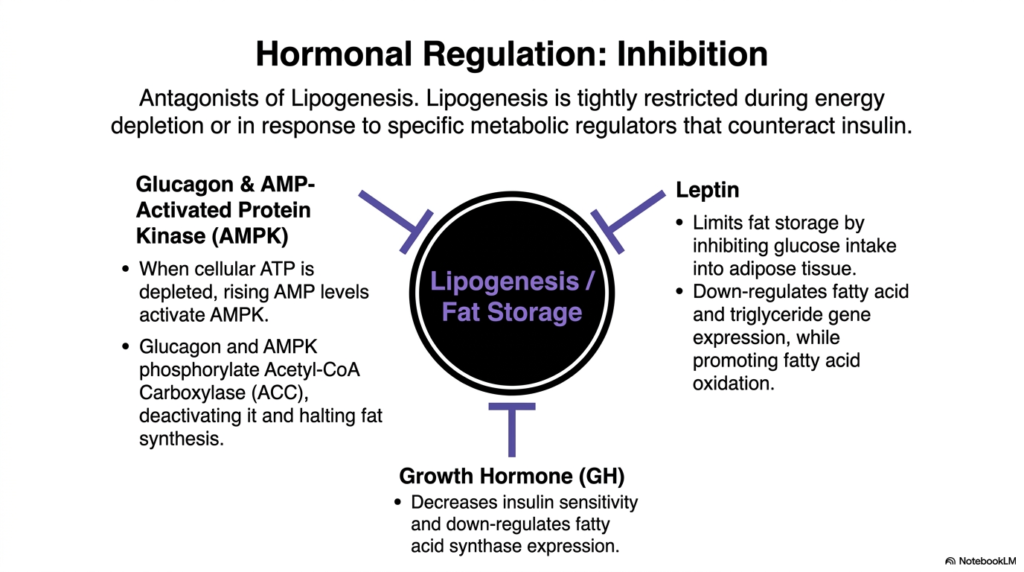
Metabolic balance requires equally powerful brakes to counteract the accelerators. Lipogenesis is strictly and aggressively restricted during periods of energy depletion, fasting, or acute physiological stress. A specialized array of hormonal antagonists precisely shuts down fat synthesis when resources are scarce. These potent inhibitors, primarily glucagon and AMP-Activated Protein Kinase (AMPK), work in direct, absolute opposition to insulin, ensuring that no precious energy is ever wasted on fat storage during times of cellular starvation.
When cellular ATP is severely depleted, rising AMP levels strongly activate AMPK. Simultaneously, a fall in blood glucose triggers the systemic release of glucagon. Together, glucagon and AMPK heavily phosphorylate the critical enzyme Acetyl-CoA Carboxylase (ACC). Unlike insulin’s activating dephosphorylation, this targeted phosphorylation instantly and completely deactivates ACC. This rapid biochemical blockade immediately halts the production of Malonyl-CoA, severely crippling the entire lipogenesis machinery and preserving vital carbon molecules for immediate energetic use.
Beyond acute kinase activity, long-term regulatory hormones such as leptin also play a defining role. Leptin, secreted directly by adipose tissue, actively limits systemic fat storage by powerfully inhibiting excess glucose intake into adipocytes. Furthermore, leptin strongly downregulates the gene expression required to synthesize fatty acids and triglycerides, while simultaneously promoting fatty acid oxidation. This powerful hormonal feedback loop prevents unchecked lipogenesis, acting as a crucial molecular thermostat for total body fat mass.
Growth Hormone (GH) also heavily contributes to the physiological suppression of fat synthesis. GH actively decreases systemic insulin sensitivity and significantly downregulates the expression of the fatty acid synthase complex. By thoroughly understanding these diverse inhibitory mechanisms—from rapid enzyme phosphorylation by AMPK to the long-term genetic suppression by Leptin—medical students can deeply appreciate the stunning complexity of metabolic homeostasis. Halting lipogenesis is just as vital for human survival as initiating it.
Slide 13: Transcriptional Regulation in Lipogenesis
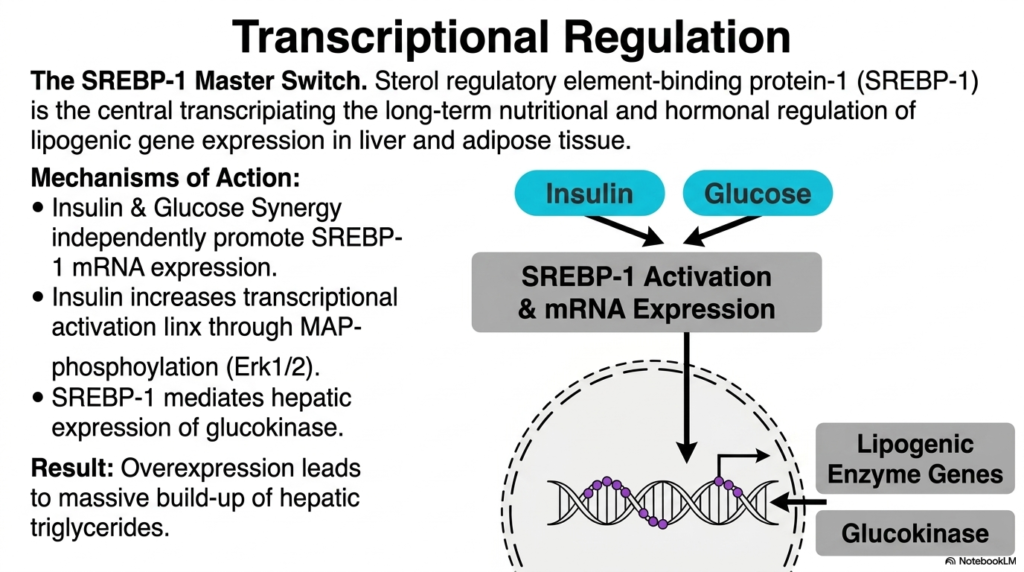
While kinases and phosphatases handle the immediate, second-to-second regulation of enzymes, the cell also possesses a brilliant master switch for long-term control. The Sterol Regulatory Element-Binding Protein-1 (SREBP-1) is the central transcription factor dictating the prolonged nutritional and hormonal regulation of lipogenesis. Operating primarily in the liver and adipose tissues, SREBP-1 serves as the primary genetic architect, physically modifying the cellular blueprint to greatly increase the machinery dedicated to fat synthesis during periods of prolonged abundance.
The mechanisms of SREBP-1 action highlight a beautiful synergy between hormones and nutrients. Both insulin and high glucose independently and strongly promote the rapid transcription of SREBP-1 mRNA. Furthermore, insulin dramatically increases the transcriptional activation capacity of the SREBP-1 protein through complex MAP kinase phosphorylation cascades (specifically, Erk1/2). This intense, multi-tiered activation ensures that the nucleus receives a highly amplified, undeniable signal to massively scale up the entire process of lipogenesis.
Once strongly activated, SREBP-1 directly binds to DNA, aggressively mediating the robust hepatic expression of crucial enzymes such as glucokinase and numerous lipogenic genes. By physically increasing the total number of enzyme complexes available in the cytoplasm, the cell’s maximum capacity for fat synthesis skyrockets. The clinical result of this severe genetic overexpression is a massive, relentless build-up of hepatic triglycerides, a phenomenon fundamentally linked to the development of fatty liver disease.
Mastering the concept of transcriptional regulation provides a profoundly complete picture of metabolism. It bridges the critical gap between acute hormonal signals and long-term phenotypic changes in the human body. SREBP-1 is the ultimate molecular reason why consistently high-calorie diets physically alter the liver’s underlying metabolic architecture. Understanding this specific genetic master switch is entirely essential for any medical professional attempting to combat the sweeping modern epidemics of obesity and hyperlipidemia driven by unchecked lipogenesis.
Slide 14: Complex Lipids Matrix: A Lipogenesis Summary
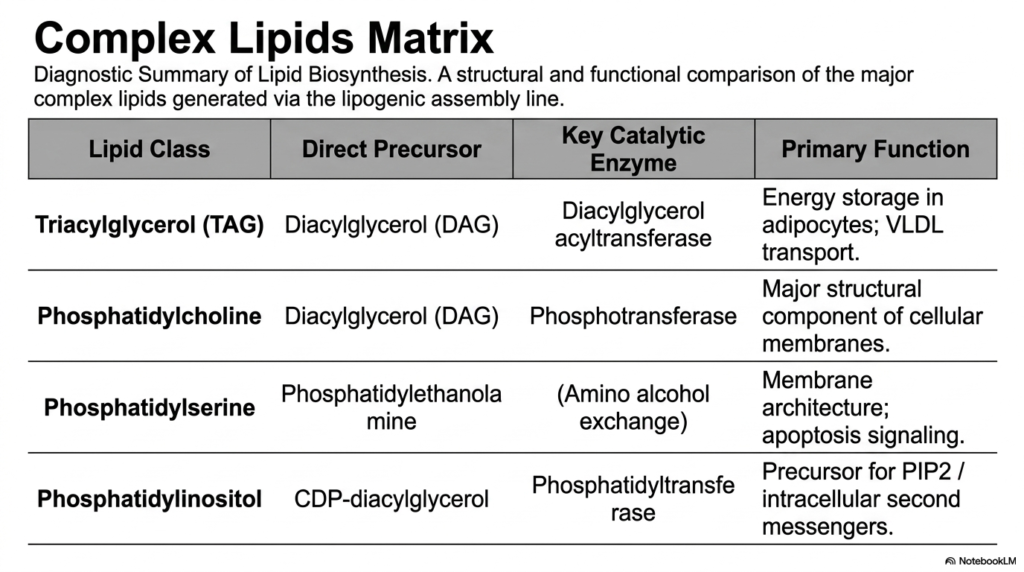
To effectively consolidate this immense volume of biochemical information, a diagnostic summary of lipid biosynthesis is incredibly helpful. This complex lipid matrix provides a high-yield structural and functional comparison of the major molecules generated via the lipogenesis assembly line. By systematically organizing the lipid classes, their direct molecular precursors, the key catalytic enzymes, and their primary physiological functions, students can rapidly integrate these distinct pathways into a cohesive, completely unified understanding of metabolic biology.
The matrix perfectly reinforces the critical branch points we previously explored. Triacylglycerol (TAG) and Phosphatidylcholine share Diacylglycerol (DAG) as their direct precursor, underscoring the central role of the DAG hub. Meanwhile, the synthesis of Phosphatidylserine is explicitly linked to the amino alcohol exchange with Phosphatidylethanolamine, emphasizing the dynamic interconversion pathways. Finally, Phosphatidylinositol is traced directly back to the highly activated CDP-diacylglycerol intermediate, confirming the existence of the vital alternate structural route in lipogenesis.
Reviewing the primary functions outlined in the matrix effectively bridges the gap between molecular structure and human physiology at the macroscopic level. While TAGs are exclusively dedicated to massive energy storage in adipocytes and VLDL transport, the phospholipids serve entirely different biological mandates. From forming major cellular membranes to mediating severe apoptosis signaling and serving as vital precursors for PIP2 intracellular messengers, the incredibly diverse physiological fates of these distinct molecules are breathtaking.
This summary table serves as the ultimate high-yield study tool for biochemistry examinations. By memorizing the specific associations among precursors, enzymes, and functions, a student fully grasps the overarching logic of lipid assembly lines. The magnificent diversity of lipid classes demonstrates how the fundamental biochemical process of lipogenesis can be carefully modified, adapted, and regulated to address a wide array of biological and structural challenges in the human body.
Slide 15: Metabolic Integration of Lipogenesis
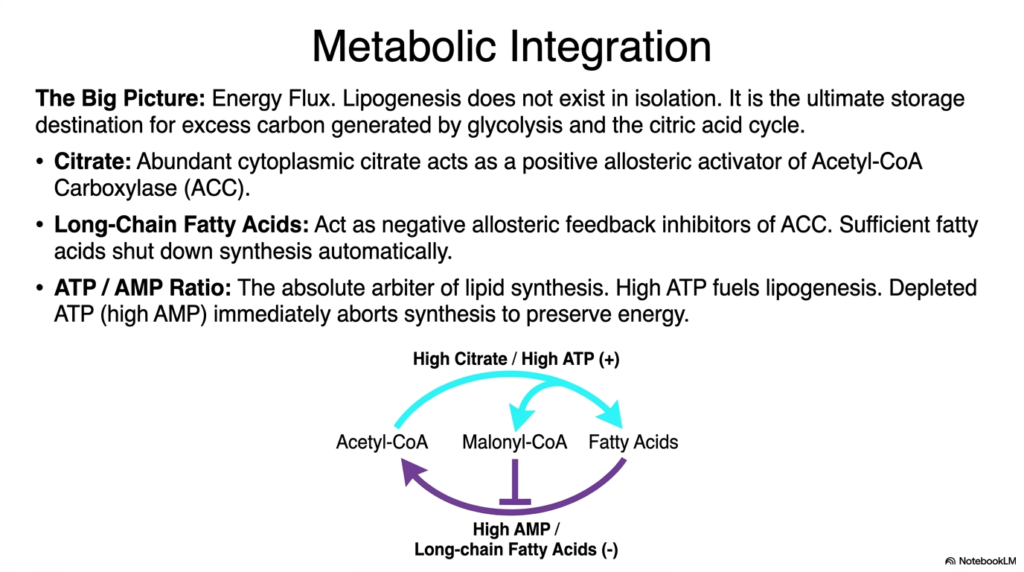
As we conclude our comprehensive study, we must step back to visualize the magnificent big picture of energy flux. Lipogenesis absolutely does not exist in cellular isolation; rather, it is the ultimate, massive storage destination for excess metabolic carbon originally generated by glycolysis and the citric acid cycle. The integration of fat synthesis with the broader metabolic network is heavily dependent on dynamic, fluctuating intracellular metabolites that act as powerful allosteric regulators, constantly assessing the cell’s immediate energetic status.
Abundant cytoplasmic citrate acts as the premier positive allosteric activator. When the citric acid cycle is overwhelmed by excess energy, citrate leaks into the cytoplasm and aggressively binds to and activates Acetyl-CoA Carboxylase (ACC). This beautiful feed-forward mechanism ensures that excess energy is instantly funneled directly into lipogenesis. Conversely, long-chain fatty acids act as devastating negative allosteric feedback inhibitors. When sufficient fatty acids have accumulated, they physically bind to ACC, automatically shutting down further synthesis to prevent toxic lipid overload.
Ultimately, the absolute arbiter of all lipid synthesis is the precise intracellular ATP-to-AMP ratio. High ATP levels fuel and actively drive lipogenesis, signaling that energy is abundant and safely storable. Conversely, depleted ATP, reflected by high levels of AMP, immediately and aggressively aborts fat synthesis to furiously preserve the remaining energy for critical survival functions. This constant, intricate tug-of-war elegantly ensures that the complex machinery of fat biosynthesis aligns perfectly with the organism’s physiological needs.
The profound integration of citrate, fatty acids, and nucleotide ratios perfectly highlights the breathtaking sophistication of human metabolism. Every single pathway, enzyme, and intermediate we have extensively studied operates within this highly sensitive, deeply interconnected biochemical web. By fully understanding the exquisite macroscopic metabolic integration of lipogenesis, future physicians and scientists can truly appreciate how the human body masterfully balances energy consumption, structural integrity, and long-term survival at the absolute molecular level.
Please read our Content Disclaimer Statement.
Check out our social media channels: