57. Nucleotide Metabolism: Purine and Pyrimidine Biosynthesis
One can imagine trying to build a towering skyscraper without any foundational bricks or structural steel. In the biological world, nucleotides serve as essential structural materials. Understanding purine and pyrimidine biosynthesis is vital because these pathways provide the indispensable building blocks for DNA and RNA. This introductory slide sets the stage for a comprehensive exploration of how cells manufacture these critical molecules. The study of purine and pyrimidine biosynthesis represents an exciting journey into the heart of molecular biology, revealing the elegance of cellular machinery.
Slide 1: The Foundations of Life’s Code – Purine and Pyrimidine Biosynthesis
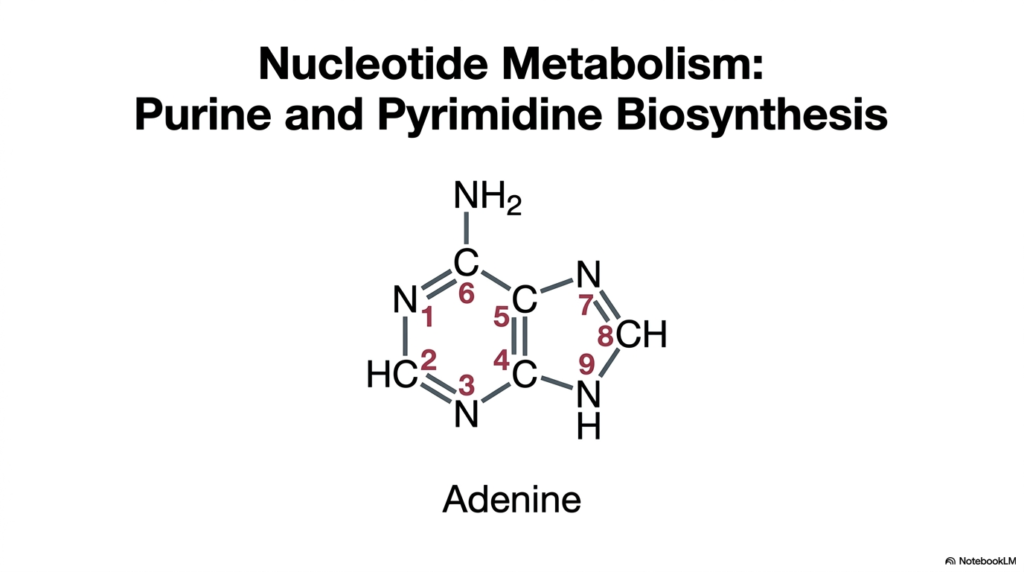
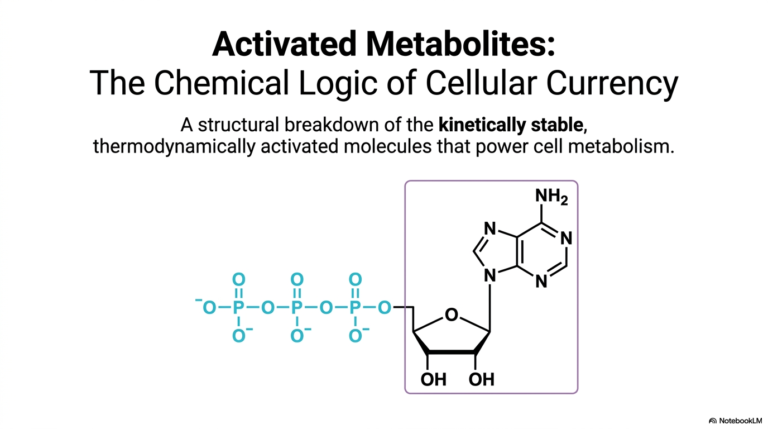
The title slide introduces the overarching topic of nucleotide metabolism by showcasing the precise chemical structure of adenine, a classic double-ringed nitrogenous base. As a student dives into the intricate nuances of purine and pyrimidine biosynthesis, recognizing the distinct molecular architectures becomes immediately crucial. Adenine serves as a prime example of the complex organic chemistry at play. It features a sophisticated, fused ring system that requires a highly orchestrated sequence of enzymatic reactions to assemble from scratch within the active cellular environment.
This specific arrangement of carbon and nitrogen atoms is far from random; every single atom in the adenine structure is meticulously sourced from specific amino acids and single-carbon donors during purine and pyrimidine biosynthesis. The intricate numbering system shown on the slide, mapping from N1 to N9, allows biochemists to precisely trace the origin of each atom. For instance, specific nitrogens are derived from amino acids like aspartate and glutamine, highlighting the deep, functional interconnectedness of different metabolic pathways within human physiology.
Mastering this foundational topic is indispensable for medical and biochemistry students because defects in these pathways consistently lead to profound clinical consequences. When the delicate balance of these biosynthetic processes is disrupted, it can result in severe metabolic disorders, devastating immune deficiencies, or even promote the uncontrolled cellular proliferation commonly observed in cancer. Therefore, carefully analyzing this introductory structural slide is the crucial first step toward understanding the profound pharmacological and pathological implications of nucleotide metabolism.
Slide 2: The Anatomy of a Nucleotide: Building Blocks of Information – Purine and Pyrimidine Biosynthesis
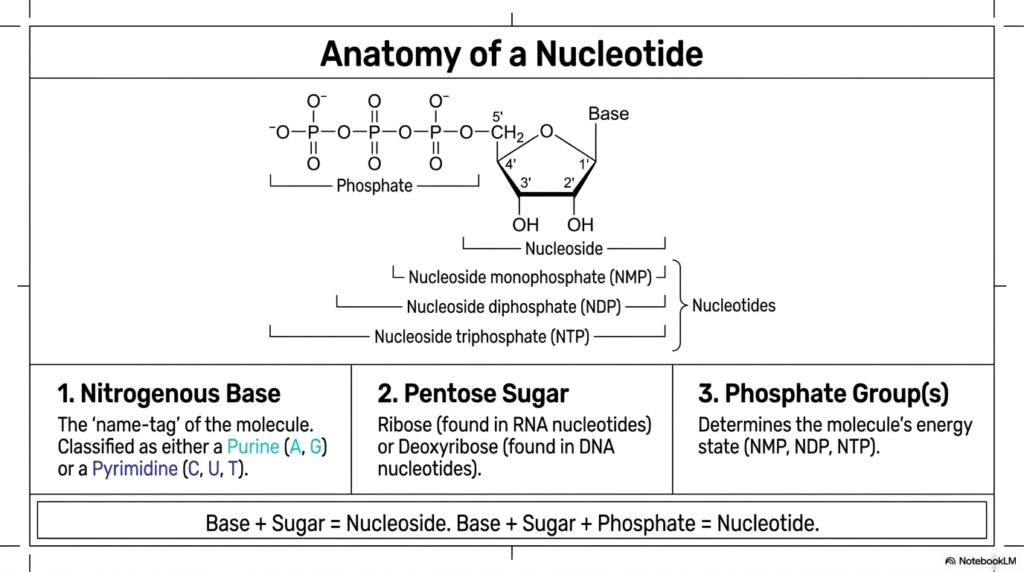
Every complex language is built upon a simple alphabet, and the genetic code is certainly no exception. Before diving deep into the intricate enzymatic steps of purine and pyrimidine biosynthesis, an observer must thoroughly understand the anatomy of the biochemical letters themselves. This foundational knowledge is essential for grasping how cellular energy is managed and how genetic information is safely propagated. A strong grasp of nucleotide structure is the absolute cornerstone for mastering the subsequent complexities of purine and pyrimidine biosynthesis.
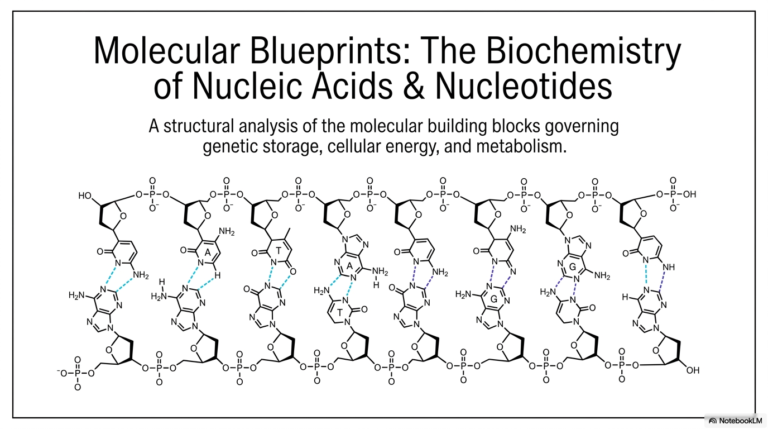
The slide clearly dissects the three fundamental components of every single nucleotide: a nitrogenous base, a pentose sugar, and one or more phosphate groups. The base acts as the unique identifying biological “name-tag” of the molecule. The distinction between a nucleoside and a nucleotide is also visually clarified; a nucleoside contains only the base and the sugar, whereas the addition of phosphate groups transforms it into a fully active nucleotide. This structural hierarchy remains a recurring theme throughout purine and pyrimidine biosynthesis.
The pentose sugar forms the critical central hub of the entire molecule. In RNA, the sugar is ribose, whereas DNA uses deoxyribose, which lacks the oxygen atom at the 2′ position. This seemingly subtle chemical difference drastically alters the stability and physiological function of the resulting nucleic acid polymers. As the energetic processes of purine and pyrimidine biosynthesis naturally unfold, these specific sugars are eventually attached to the synthesized nitrogenous bases to form the diverse array of molecules required for cellular survival.
Finally, the attached phosphate groups are the critical determinants of the molecule’s overall energy state. Nucleotides exist natively as mononucleotides, dinucleotides, or highly energetic triphosphate nucleotides. The high-energy bonds between these phosphate groups determine the molecule’s utility as a cellular currency. Understanding this specific anatomy is profoundly important for clinicians and researchers, as numerous antiviral and anticancer medications are meticulously designed as synthetic nucleotide analogs to intentionally disrupt pathogenic replication or halt malignant tumor growth.
Slide 3: Nitrogenous Bases and The Two Great Families – Purine and Pyrimidine Biosynthesis
To truly appreciate the chemical language of genetics, one must reliably recognize the distinct structural characteristics of its biological consonants and vowels. The foundational step in mastering purine and pyrimidine biosynthesis is distinguishing between the two primary families of nitrogenous bases. These molecules are not just passive structural elements; they participate heavily in dynamic cellular interactions. Exploring these structural families highlights the underlying chemical logic that actively drives the complex enzymatic pathways of purine and pyrimidine biosynthesis.
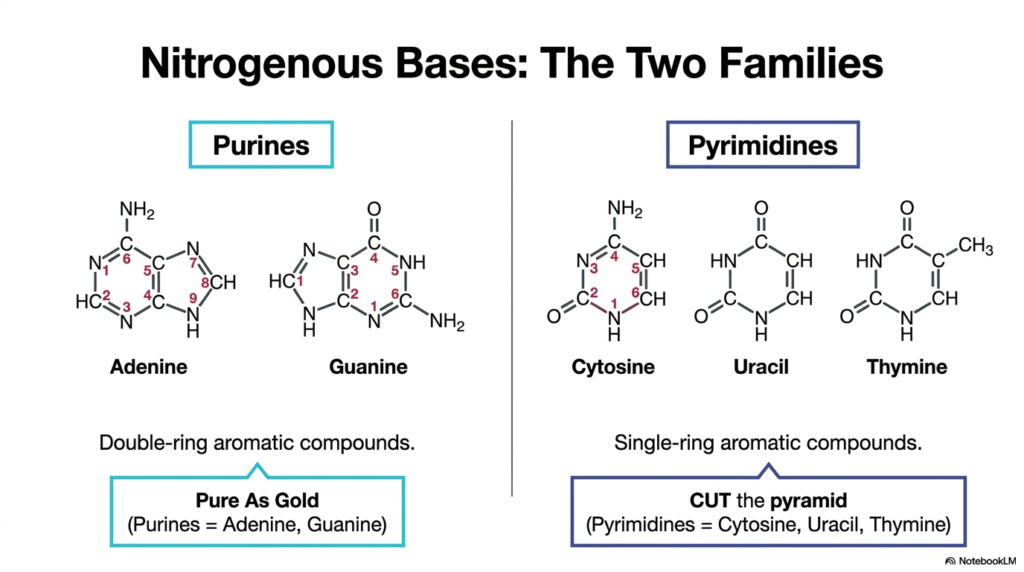
This slide beautifully illustrates the stark structural dichotomy between the purines and the pyrimidines. Purines, specifically adenine and guanine, are characterized visually by their larger, double-ring aromatic structures. In stark contrast, pyrimidines—which include cytosine, uracil, and thymine—are notably smaller, consisting of a single six-membered aromatic ring. The visual distinction provided here is vital for understanding exactly why entirely separate specific enzymes are required for each molecular pathway during purine and pyrimidine biosynthesis.
To aid memory retention, the slide provides invaluable mnemonic devices widely used by medical students worldwide. The phrase “Pure As Gold” perfectly encapsulates the purine family, directly representing Adenine and Guanine. Conversely, the phrase “CUT the pyramid” seamlessly links Cytosine, Uracil, and Thymine to the single-ringed pyrimidine family. These simple, memorable aids are incredibly effective tools for efficiently navigating the incredibly dense, reaction-heavy material inherent to the detailed study of purine and pyrimidine biosynthesis.
The actual physical sizes of these molecules rigidly dictate the base-pairing rules essential for maintaining DNA double-helix stability. A large, bulky purine must always pair with a smaller pyrimidine to maintain a uniform width along the entire DNA molecule. If these geometric rules are severely violated by mutations, the entire genomic structure becomes functionally unstable. Therefore, a profound understanding of these two chemical families remains critically crucial for comprehending genetics and modern molecular biology.
Slide 4: Origin Pathways: De Novo versus Salvage – Purine and Pyrimidine Biosynthesis

Biological cells are ultimate models of manufacturing efficiency, constantly balancing the tremendous energetic cost of synthesizing new materials against the metabolic benefits of active recycling. When one delves deeply into purine and pyrimidine biosynthesis, it quickly becomes evident that organisms rely on two distinct strategies to secure their nucleotide supply. Understanding this dual metabolic approach is critical for predicting exactly how different bodily tissues respond to biological stress. The study of purine and pyrimidine biosynthesis requires an appreciation for this delicate balance.
The slide clearly and effectively contrasts the intense “De Novo” synthesis pathway with the highly efficient “Salvage” pathway. De novo synthesis literally means making a molecule “anew.” This mechanism involves carefully constructing complex nucleotide structures from very simple building blocks such as sugars, amino acids, and carbon dioxide. This process is inherently extremely energy-intensive. This heavy energetic toll explains why de novo purine and pyrimidine biosynthesis is predominantly active only in rapidly dividing cells undergoing the S phase.
In stark contrast, biological salvage pathways operate beautifully like an advanced cellular recycling center. Instead of building energetically expensive molecules from scratch, these specialized pathways actively rescue preformed nitrogenous bases and nucleosides generated directly from the natural degradation of existing cellular nucleic acids. By constantly recycling these vital components, resting or fully mature cells manage to save an enormous amount of metabolic energy. The delicate interplay between these two routes in purine and pyrimidine biosynthesis ensures optimal nucleotide pools.
The underlying physiological significance of distinguishing these two pathways truly cannot be overstated in modern clinical medicine. Because malignant cancer cells are inherently highly proliferative, they become dangerously heavily dependent on the energy-draining de novo pathways to fuel their relentless, unchecked division. This profound vulnerability makes the de novo enzymes excellent targets for intensive pharmacological intervention, as specific chemotherapy drugs are designed to effectively starve tumor cells of the crucial nucleotides they desperately need.
Slide 5: The Blueprint of Purine Ring Assembly – Purine and Pyrimidine Biosynthesis
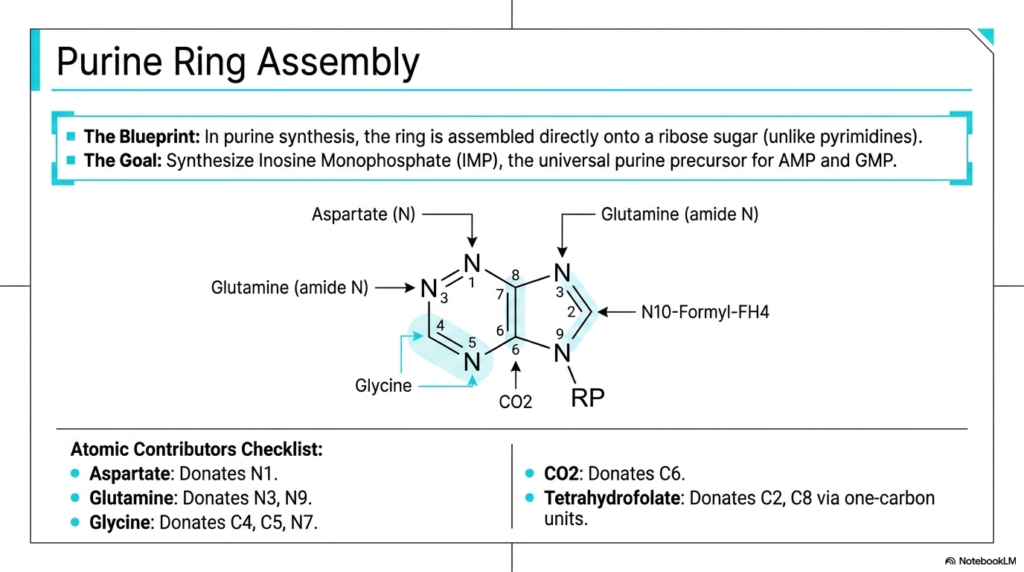
Successfully building a complex molecular structure strictly requires a precise architectural blueprint and an incredibly reliable supply chain of raw chemical materials. In the dense metabolic realm of purine and pyrimidine biosynthesis, the sequential assembly of the double-ring structure represents a marvel of cellular engineering. Analyzing the precise biological origin of every atom in this ring fully reveals the deep metabolic integration required to sustain complex life. This slide details the specific contributors driving purine and pyrimidine biosynthesis.
This helpful visual thoroughly breaks down the step-by-step construction of the purine ring, directly highlighting a fascinating mechanistic feature: the ring is actually assembled directly onto a pre-existing ribose sugar anchor. The ultimate functional goal of this intricate, multi-step pathway is the successful production of Inosine Monophosphate (IMP). This molecule serves as the essential universal precursor for both major purine nucleotides, AMP and GMP. Understanding this blueprint is a primary cornerstone of successfully mastering purine and pyrimidine biosynthesis.
The comprehensive “Atomic Contributors Checklist” meticulously details the various molecular donors involved. Specific amino acids are the primary biological sources of necessary nitrogen; aspartate donates N1, while glutamine provides the critical amides for N3 and N9. The amino acid glycine is heavily involved, effectively contributing a larger carbon-carbon-nitrogen backbone. Furthermore, carbon dioxide provides the essential C6 carbon, and tetrahydrofolate is absolutely crucial for donating necessary single-carbon units. This sourcing highlights the complexity of purine and pyrimidine biosynthesis.
Why should a student carefully map every single biological atom? Because each specific contributing molecule ultimately represents a potential metabolic bottleneck or a highly viable pharmacological target. For example, severe folate deficiency directly impairs the steady supply of tetrahydrofolate, completely crippling the cell’s ability to seamlessly provide carbons for positions C2 and C8. By fully understanding these precise atomic contributions, medical professionals can diagnose nutritional deficiencies much more effectively and successfully utilize targeted antifolate therapies.
Slide 6: Initiating the Process with PRPP – Purine and Pyrimidine Biosynthesis
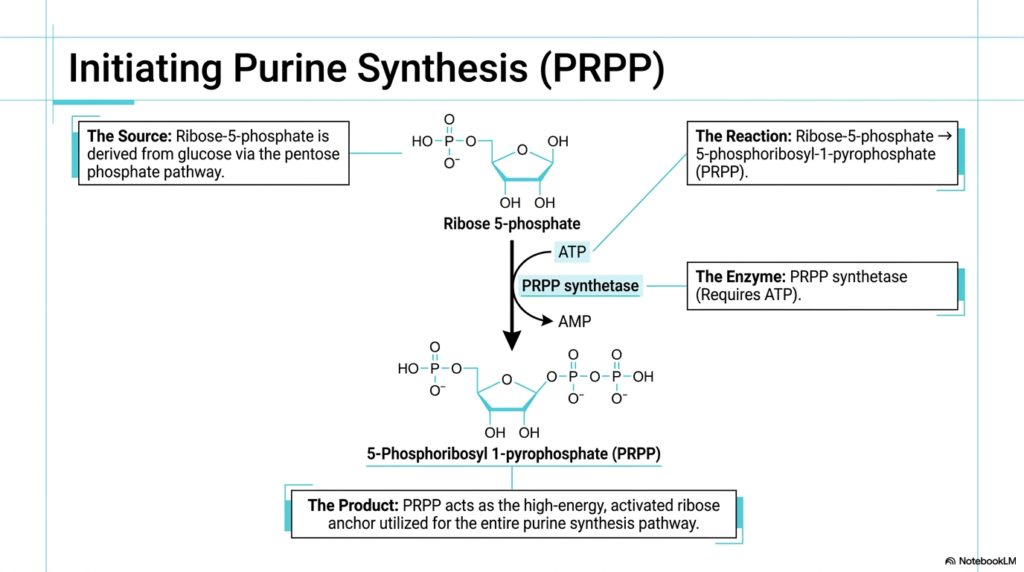
Every great biochemical journey necessarily requires a definitive starting point, a structural foundation upon which everything else must be reliably built. In the highly complex cellular landscape of purine and pyrimidine biosynthesis, the absolutely essential starting material is a specifically activated sugar molecule. Understanding exactly how this critical molecular anchor is formed is the true key to unlocking the entire metabolic pathway. The initiation phase of purine and pyrimidine biosynthesis highlights the critical intersection between basic carbohydrate and nucleotide metabolism.
The slide prominently introduces 5-Phosphoribosyl-1-pyrophosphate, commonly known as PRPP. This specific molecule serves directly as the high-energy, chemically activated ribose anchor utilized extensively throughout the entire purine synthesis pathway. The initiation process officially begins with Ribose-5-phosphate, a simple sugar molecule naturally diverted from the pentose phosphate pathway. This crucial metabolic handoff remains a fundamentally defining characteristic of purine and pyrimidine biosynthesis, smoothly demonstrating how the functioning cell seamlessly coordinates its different energetic and structural needs.
The enzymatic conversion of Ribose-5-phosphate to PRPP is precisely catalyzed by the vital enzyme PRPP synthetase. This specific reaction is not merely a passive cellular event; it demands a highly significant energetic investment, strictly requiring the active consumption of ATP to successfully attach a high-energy pyrophosphate group. This newly attached pyrophosphate functions exceptionally well as a highly effective leaving group, perfectly priming the sugar molecule for the subsequent reactions driving the advancing stages of purine and pyrimidine biosynthesis.
PRPP is definitely not merely a passive structural cellular foundation; it functions concurrently as a profoundly important metabolic regulator. Its intracellular concentration directly acts as a master biological switch, tightly controlling the baseline rate of nucleotide production. Abnormalities resulting in extreme PRPP synthetase overactivity can lead to a dangerous pathological overproduction of purines, eventually resulting in the debilitating, excruciating joint pain clinically characteristic of severe gout.
Slide 7: The Rate-Limiting Step Controlled by GPAT – Purine and Pyrimidine Biosynthesis
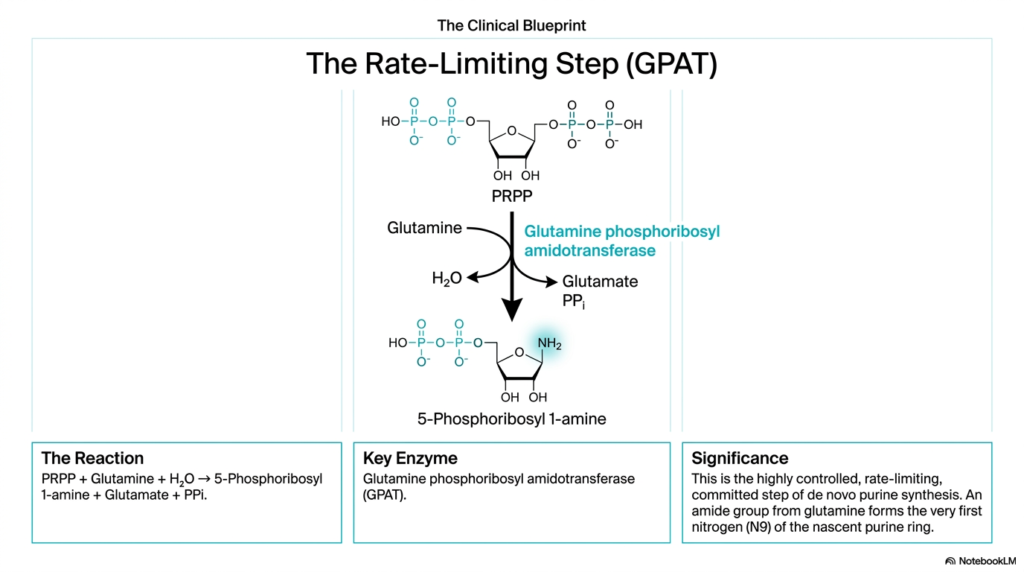
In any highly complex biochemical manufacturing process, there is always one master control point that tightly dictates the maximum overall speed of cellular production. Within the remarkably intricate metabolic pathways of purine and pyrimidine biosynthesis, precisely identifying this biochemical bottleneck is crucial for a deep understanding of metabolic regulation. This specific slide explores the pivotal, committed step in the pathway, revealing the elegant enzymatic mechanisms that strongly govern purine and pyrimidine biosynthesis.
The critical academic spotlight here appropriately falls entirely on the enzyme Glutamine phosphoribosyl amidotransferase, which is widely abbreviated simply as GPAT. This specific enzyme controls the highly regulated, explicitly rate-limiting, and officially committed step of de novo purine synthesis. In this critical biochemical reaction, the activated sugar anchor PRPP reacts directly with the important amino acid glutamine. The enzyme successfully catalyzes the displacement of the high-energy pyrophosphate group, thereby propelling the cycle of purine and pyrimidine biosynthesis forward.
The profound physiological and clinical significance of this specific, highly targeted reaction lies completely in the successful creation of 5-Phosphoribosyl 1-amine. The newly attached amide nitrogen atom will ultimately purposefully become the vital N9 position of the final, fully assembled functional purine ring. Because this specific enzymatic step remains highly thermodynamically irreversible under completely normal physiological conditions, the cell remains fully energetically committed to finishing the entire pathway, a key concept when studying purine and pyrimidine biosynthesis.
Functioning explicitly as the primary rate-limiting enzyme, GPAT naturally serves directly as the absolute primary gateway for external pharmacological intervention. Because the intrinsic activity of this specific enzyme strictly, fundamentally dictates the overall systemic speed of cellular DNA building block production, directly inhibiting GPAT rapidly and effectively halts excessive cellular proliferation. By diligently studying this highly regulated biochemical bottleneck, modern future physicians naturally gain completely deep, actionable insights into how powerful targeted biochemical therapies truly work.
Slide 8: Master Feedback Control of Purine Synthesis – Purine and Pyrimidine Biosynthesis
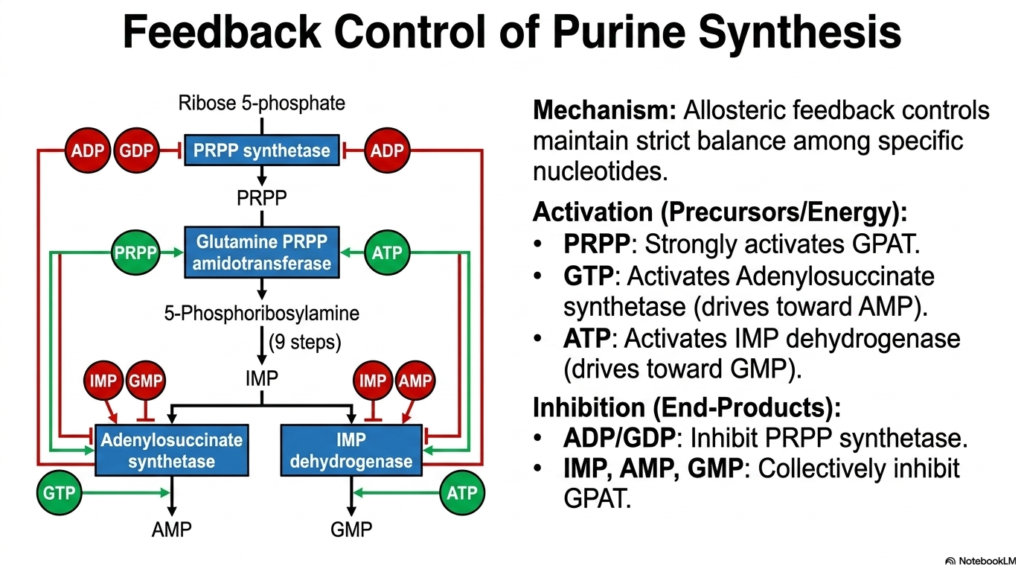
A cellular factory operating continuously without a strict regulatory biological management system will quickly overproduce unneeded inventory and rapidly exhaust its highly valuable raw chemical materials. The stable cellular environment successfully prevents this chaotic metabolic outcome through deeply elegant, complex allosteric feedback mechanisms. Exploring these precise regulatory networks remains a core component of successfully mastering purine and pyrimidine biosynthesis. The intricate study of purine and pyrimidine biosynthesis reveals a highly sophisticated, constant balancing act.
This specific, detailed slide accurately maps out the profoundly complex allosteric feedback controls absolutely necessary to seamlessly maintain a uniquely strict biochemical balance among entirely specific cellular nucleotides. The regulatory mechanisms are carefully divided functionally into forward activation, primarily driven by structural precursors, and biological inhibition, heavily driven by eventual end-product accumulation. When foundational precursors like highly energetic PRPP are extremely plentiful, they very strongly activate the critical GPAT enzyme, successfully pushing purine and pyrimidine biosynthesis dynamically forward.
The specific regulatory biochemical branching point occurring directly at Inosine Monophosphate (IMP) remains particularly fascinating from a functional standpoint. To ensure a meticulously balanced internal pool of both major biological purines, the synthesis of AMP strictly requires substantial energy input directly from GTP. Concurrently, the vital synthesis of GMP is uniquely powerfully powered entirely by ATP. Furthermore, the final cellular end-products collectively exert powerful, necessary negative feedback, thereby ensuring vital metabolic homeostasis during purine and pyrimidine biosynthesis.
This profound cellular concept of metabolic cross-regulation serves perfectly as a truly beautiful, remarkable example of functional evolutionary biochemical design. By cleverly using GTP to synthesize adenine and relying entirely on ATP to synthesize guanine, the biological cell ensures it never produces a toxic, overwhelming excess of a single nucleotide. For all dedicated medical students, a deep understanding of these intricate, highly specific feedback loops remains clinically paramount for diagnosing severe disease.
Slide 9: Purine Salvage and Lesch-Nyhan Syndrome – Purine and Pyrimidine Biosynthesis
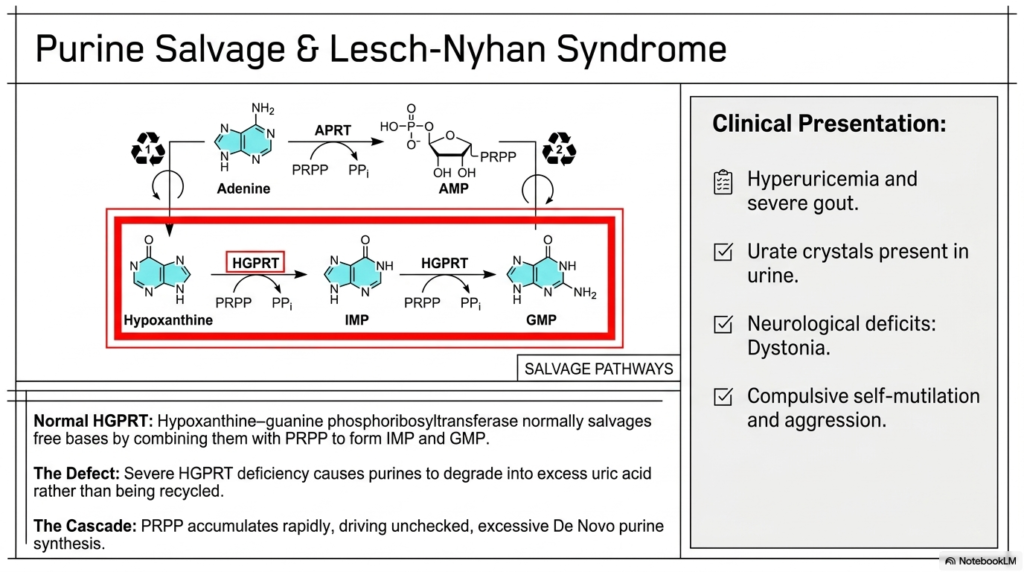
While the remarkable biological creation of entirely new, complex molecules remains completely fascinating, the devastating failure of dedicated cellular metabolic recycling systems routinely leads directly to incredibly severe clinical pathological consequences. The tragic intersection of foundational biochemistry and human pathology is particularly evident when examining the genetic defects inextricably associated with purine and pyrimidine biosynthesis. This highly clinical slide completely underscores exactly why a deep understanding of purine and pyrimidine biosynthesis remains fundamentally clinically vital.
The informative visual clearly highlights the essential purine salvage pathway, with a strong emphasis on the absolutely crucial biological enzyme Hypoxanthine-guanine phosphoribosyltransferase, commonly abbreviated as HGPRT. Normally, this vitally important enzyme actively salvages free, floating purine bases, seamlessly and efficiently combining them with activated PRPP to rapidly reform the highly functional IMP and GMP nucleotides. This brilliant recycling mechanism operates efficiently concurrently with foundational de novo purine and pyrimidine biosynthesis to fully protect mature resting cells from total energetic exhaustion.
The comprehensive slide graphically depicts the truly severe, devastating consequences that naturally result from profound HGPRT deficiency. When this absolutely critical vital recycling enzyme remains absent, the valuable free purine bases simply cannot be naturally salvaged and are unfortunately funneled directly heavily into biological degradation pathways, rapidly resulting in an overwhelming massive overproduction of toxic uric acid. This massive, dangerous excess of PRPP rapidly, forcefully, and directly drives wildly excessive, absolutely unchecked de novo purine and pyrimidine biosynthesis.
This specific devastating biochemical metabolic cascade unfortunately naturally manifests functionally clinically as devastating Lesch-Nyhan syndrome, an incredibly severe X-linked recessive genetic disorder. Affected pediatric patients characteristically present with severe systemic hyperuricemia, extraordinarily excruciating clinical gout, and highly characteristic, visible urate crystals routinely present inside their infant diapers. Most profoundly tragically, the debilitating disease strictly directly causes absolutely profound, highly visible neurological deficits, severe motor dystonia, and a hallmark tragic behavioral symptom of relentless compulsive physical self-mutilation.
Slide 10: The Pyrimidine Ring Assembly Blueprint – Purine and Pyrimidine Biosynthesis
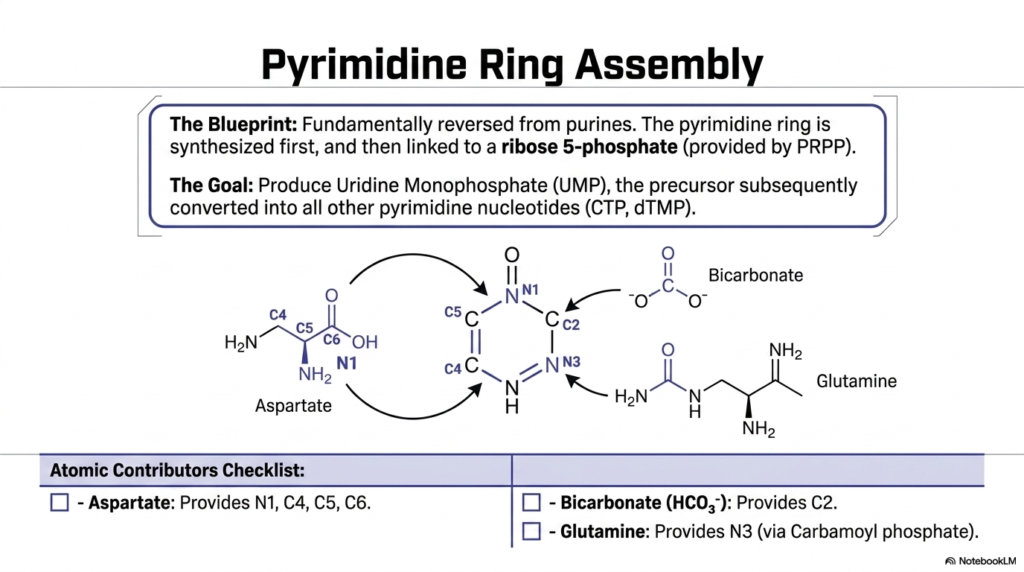
Having fully successfully completely conquered the heavily complex, remarkably dense double-ringed molecular structures, the vital academic focus must absolutely necessarily briefly shift completely to the somewhat slightly physically smaller, single-ringed molecular components of the genetic code. The core metabolic logic underlying the successful biological creation of these specific pyrimidines remains remarkably structurally distinct. Analyzing these stark differences remains an absolutely vital step, particularly for mastering the broader biological landscape of purine and pyrimidine biosynthesis.
This highly specific, detailed slide thoroughly and visually illustrates the essential, fundamental blueprint required for reliable pyrimidine ring assembly, significantly highlighting a profound structural reversal strategy that is unique compared to standard purines. Instead of systematically building the complex molecule directly onto a sugar anchor, the entire pyrimidine ring is first biochemically synthesized independently. This specific alternate, unique, sequential pathway, which naturally drives purine and pyrimidine biosynthesis, reliably produces vital Uridine Monophosphate.
The incredibly helpful, meticulously detailed “Atomic Contributors Checklist” deeply and actively simplifies the wildly diverse, structurally complex molecular origins necessary to completely build the specific pyrimidine ring. The essential amino acid aspartate functions remarkably as the absolutely massive, major, foundational structural contributor, rapidly providing a huge, significant, vital block of specific structural atoms. Tracking these precise, highly specific, individual atomic structural sources is crucially, vitally, fundamentally important for all dedicated biology students intently studying purine and pyrimidine biosynthesis.
Deeply actively fully understanding this entirely critically fundamentally reversed sequential biological assembly order remains definitely absolutely not simply just a completely purely highly pedantic abstract intellectual exercise; it has massive, incredible structural implications deeply for modern complex drug design. Because the foundational pyrimidine ring is heavily actively fundamentally successfully physically built completely absolutely independently first, specific isolated uniquely necessary enzymes can successfully effectively strategically be chemically medically heavily pharmacologically biologically fundamentally actively safely therapeutically fully specifically completely cleanly targeted.
Slide 11: Initiating Pyrimidine Synthesis with CPS-II – Purine and Pyrimidine Biosynthesis
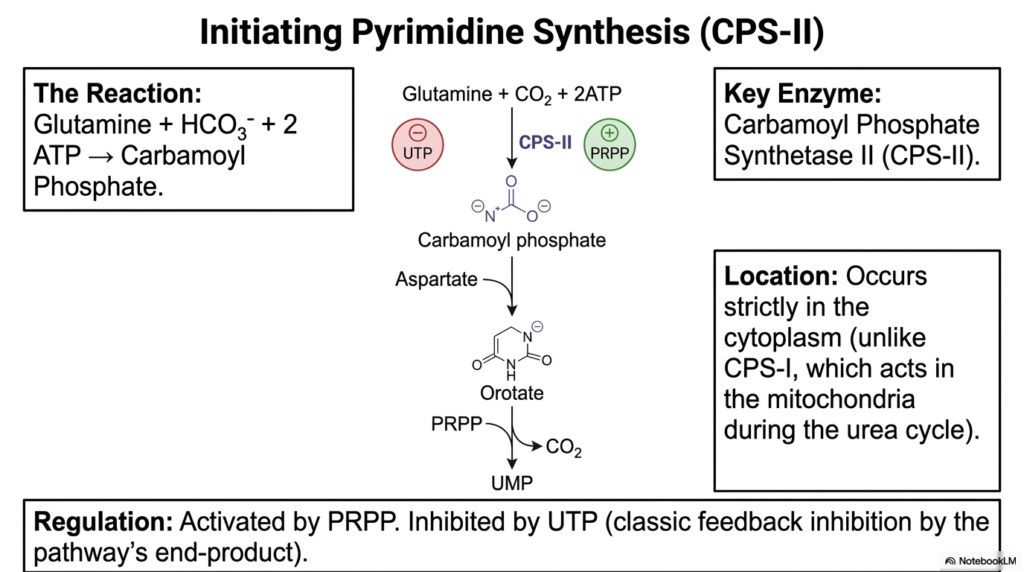
Every biological engine needs a spark to start running. In the cellular world, the creation of single-ringed genetic bases begins with a specific enzymatic ignition. This slide explores the vital initiation phase of pyrimidine production, focusing squarely on the master regulatory enzyme Carbamoyl Phosphate Synthetase II. By mastering this critical gateway, medical students can better understand how the complex pathways of Purine and Pyrimidine Biosynthesis are tightly controlled to sustain rapid cellular replication and prevent metabolic chaos.
The featured reaction shows glutamine, bicarbonate, and ATP combining to form carbamoyl phosphate. This crucial committed step is driven entirely by Carbamoyl Phosphate Synthetase II, or CPS-II. As the primary rate-limiting enzyme for the pyrimidine branch of Purine and Pyrimidine Biosynthesis, it dictates the overall pace of production. Notably, this reaction occurs exclusively within the cellular cytoplasm. This strict physical compartmentalization is essential to prevent interference with CPS-I, a structurally similar enzyme that operates within the mitochondria to carry out the urea cycle.
Following its successful synthesis, carbamoyl phosphate merges with the amino acid aspartate to form an intermediate structure known as orotate. As a defining characteristic of this pathway, orotate is attached only to the activated PRPP sugar anchor to finalize the creation of Uridine Monophosphate. UMP then serves as the universal foundational precursor for all other necessary pyrimidines, including CTP and dTMP. Understanding this precise sequence in Purine and Pyrimidine Biosynthesis is critical for grasping how cells manage their internal nucleotide inventories during the intense demands of cellular division.
The allosteric regulation of CPS-II represents a masterclass in biological feedback logic. The enzyme is strongly activated by elevated PRPP levels. This activation serves as a chemical signal indicating that the cell has an abundance of foundational sugar anchors ready for assembly. Conversely, CPS-II is directly inhibited by UTP, a final end product of the pathway. This classic negative feedback loop ensures that the energetic cellular machinery powers down once sufficient single-ring structures have formed, conserving valuable ATP for other essential life processes.
Slide 12: De Novo Synthesis Comparison – Purine and Pyrimidine Biosynthesis
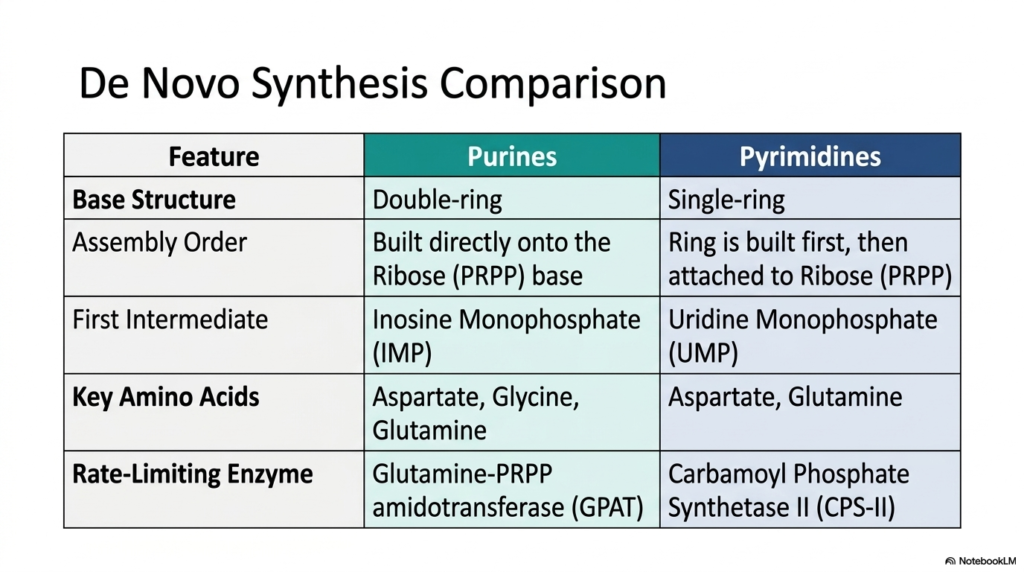
A good scientist always compares and contrasts to see the bigger picture. After exploring the detailed molecular steps of nucleotide metabolism, it is time to synthesize this knowledge into a cohesive overview. This final summary slide presents a clear, comparative look at the two distinct manufacturing strategies that define Purine and Pyrimidine Biosynthesis. For medical students, memorizing these core differences is essential for board exams and for understanding the specific targets of various pharmacological therapies.
The slide clearly outlines the foundational structural differences between the two base families. Purines are built as bulky, double-ring structures, whereas pyrimidines consist of a much smaller single ring. The assembly order represents the most profound divergence within Purine and Pyrimidine Biosynthesis. For the larger bases, the ring is constructed atom by atom directly onto the PRPP ribose anchor. In contrast, the single ring is built independently first and attached to the PRPP sugar only upon completion. This fundamental difference dictates the unique enzymatic requirements for each respective pathway.
Another key comparison involves the first stable intermediate product generated by each sequence. The complex double-ring pathway yields Inosine Monophosphate, which then branches out to form AMP and GMP. The single-ring sequence produces Uridine Monophosphate, the vital precursor for all subsequent pyrimidine variations. Additionally, the specific amino acid contributors vary. While both pathways require aspartate and glutamine for nitrogen atoms, the double-ring structure also demands the amino acid glycine to complete its larger carbon backbone. This underscores the intense nutritional demands of Purine and Pyrimidine Biosynthesis.
Finally, the summary highlights the distinct rate-limiting enzymes that serve as the master control switches for each process. Glutamine phosphoribosyl amidotransferase governs the double-ring assembly, while Carbamoyl Phosphate Synthetase II regulates the single-ring production. Understanding the split nature of Purine and Pyrimidine Biosynthesis gives future physicians actionable insights. Because these distinct pathways use entirely different enzymes and assembly sequences, targeted chemotherapies can be designed to inhibit a single branch, starving cancer cells of specific genetic letters while minimizing systemic toxicity.
Please read our Content Disclaimer Statement.
Check out our social media channels: