59. Heme Biosynthesis: Molecular Pathways and Pathology
Imagine a microscopic trap designed exclusively to capture the most vital element for human survival: oxygen. Without this molecular net, complex aerobic life would simply cease to exist. This concept lies at the heart of the slide deck, which systematically aims to illuminate the complex enzymatic pathways, structural transformations, and clinical pathologies defining cellular respiration. Understanding the stages of heme biosynthesis is absolutely fundamental for college and medical school students. It bridges foundational organic chemistry with profound clinical realities, showing how atoms assemble into the breath of life.
Slide 1: Introduction to Heme Biosynthesis: The Molecule That Breathes Life
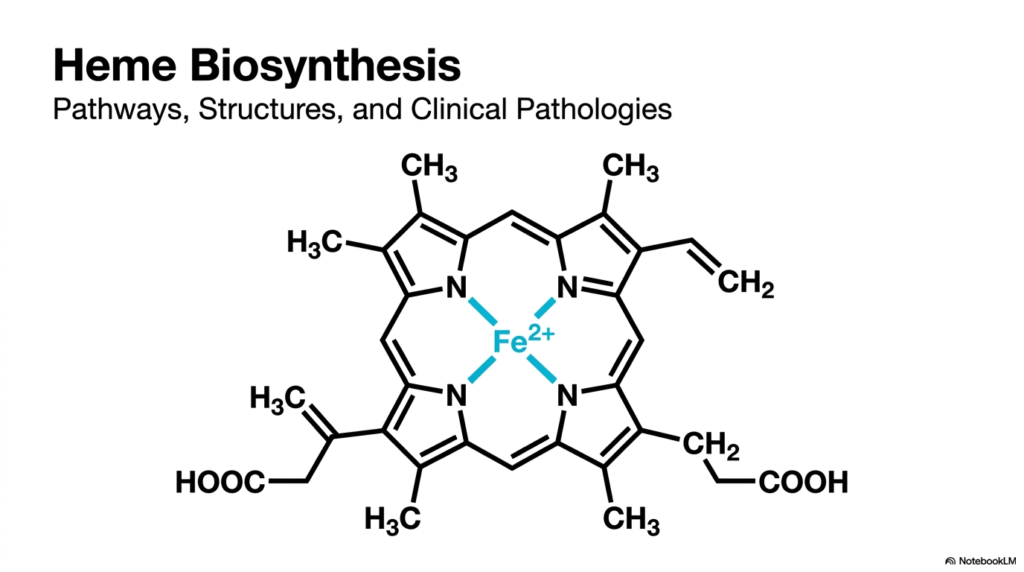
The provided visualization displays the structural masterpiece produced through the successful completion of heme biosynthesis. The foundation is a massive macrocycle known as a tetrapyrrole ring, an intricate chemical cage composed of four smaller pyrrole subunits securely bound together. At the exact geometric center of this biological wonder sits a single ferrous iron atom, denoted as Fe2+. This iron acts as the functional core, conferring on the molecule its unique ability to bind oxygen reversibly. It does this while avoiding irreversible oxidation, meaning the iron works without permanently rusting.
Surrounding this metallic center, students will notice specific side chains jutting outward, specifically methyl, vinyl, and propionate functional groups. These chemical branches are far from random; they are meticulously crafted at each stage of heme biosynthesis to serve a critical structural role. They dictate exactly how the finished molecule docks into larger protein structures, such as hemoglobin within red blood cells and myoglobin in muscle tissues. The precise positioning of these groups ensures the compound anchors perfectly into hydrophobic protein pockets, preventing toxic reactions.
For future clinicians and researchers, analyzing this finished biochemical blueprint is just the starting line. Mastering the exact step-by-step assembly line of heme biosynthesis offers profound insights into metabolic regulation and energy production. When this complex molecular factory functions at peak efficiency, it sustains the energy needs of every organ. Conversely, the slightest structural error during assembly cascades into debilitating blood and neurological disorders. Therefore, grasping this initial architectural overview is an essential prerequisite for diagnosing and treating associated metabolic conditions in medical practice.
Slide 2: Molecular Architecture of Porphyrins in Heme Biosynthesis: Building the Biological Cage
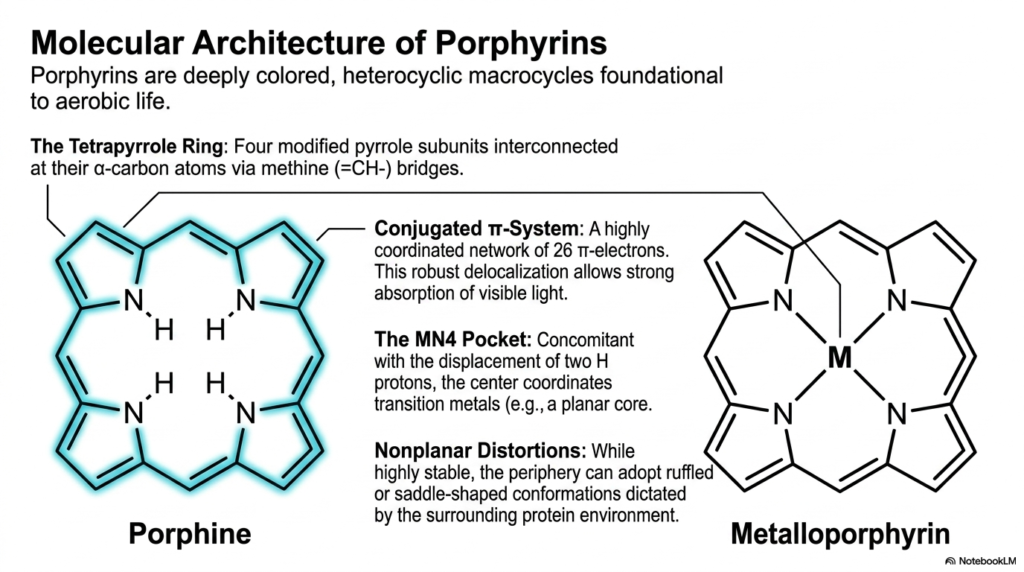
What gives blood its deep red hue and plants their vibrant green color? The answer lies in porphyrins, deeply colored macrocycles that form the chemical backbone of aerobic life. The core purpose of this slide deck is to decode these molecular structures and trace their origin through the heme biosynthesis pathway. Medical and college students must grasp this fundamental architecture, as these rings are the foundational scaffolding that nature uses to trap light and transport gases across a vast array of living organisms.
The illustration breaks down the porphine structure, the simplest foundational model formed before complex heme biosynthesis occurs. It highlights the tetrapyrrole ring, composed of four modified pyrrole subunits. These four nitrogen-containing rings are interconnected at specific carbon atoms via methine bridges. This creates a highly rigid and stable cyclic framework. Within this rigid border lies a highly coordinated network of 26 electrons, known as a conjugated pi system. This extensive electron delocalization is exactly what allows the molecule to absorb visible light so intensely.
At the center of this ring lies the MN4 pocket, an electron-rich void perfectly sculpted for capturing transition metals. During the later stages of heme biosynthesis, two hydrogen protons are displaced from the central nitrogen atoms to make room for a metal ion. When a metal occupies this central void, the molecule graduates from a simple porphine into a metalloporphyrin. This transformation is critical, as the captured metal dictates biological function, whether magnesium for photosynthesis or iron for oxygen transport in humans.
Interestingly, while the basic porphyrin ring is notoriously stable, it is not completely flat or rigid in biological systems. The periphery of the ring can experience nonplanar distortions. Depending on the surrounding protein environment, the macrocycle can adopt ruffled, saddled, or domed conformations. This flexibility is a vital feature of heme biosynthesis, allowing the final metalloporphyrin to bend and shift when oxygen binds or releases. Understanding this dynamic architecture prepares students to comprehend hemoglobin’s mechanical breathing at the molecular level.
Slide 3: The Bipartite Pathway of Heme Biosynthesis: A Tale of Two Cellular Compartments
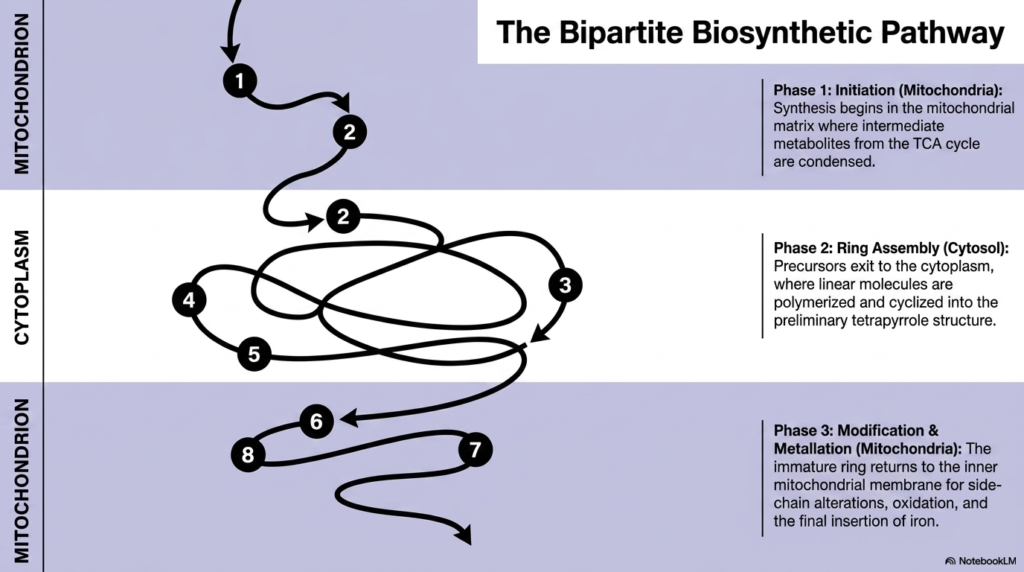
Imagine a highly secure factory where half the manufacturing happens in a fortified underground vault, and the other half occurs on a bustling open floor. This spatial division is the reality of cellular metabolism. The core purpose of this slide deck is to map this complex choreography, illustrating that heme biosynthesis is uniquely bipartite. College and medical students must understand this spatial division, as the constant shuttling of metabolites between cellular compartments is central to metabolic regulation and disease manifestation.
The schematic outlines the fascinating journey a molecule takes during heme biosynthesis, shifting between the mitochondrion and the cytoplasm. Phase one initiates deep within the mitochondrial matrix. Here, basic precursor molecules from the tricarboxylic acid cycle condense to form the initial building blocks. The mitochondrion provides a specialized, energy-rich environment necessary to jumpstart the entire operation. However, the initial product is highly reactive and requires processing machinery that is found only outside this organelle, necessitating its export into the cellular fluid.
Once out in the cytoplasm, phase two begins. This stage of heme biosynthesis focuses entirely on ring assembly. The previously linear precursor molecules are stitched together, polymerized, and carefully folded into a preliminary cyclic tetrapyrrole structure. The cytoplasm offers a spacious environment and specific soluble enzymes needed to handle these bulky intermediate compounds. Constructing the initial ring outside the mitochondrion prevents dangerous premature reactions with mitochondrial iron reserves, showcasing an elegant evolutionary strategy for chemical safety and spatial organization.
Finally, phase three requires the immature ring to return to its birthplace. The macrocycle crosses back into the inner mitochondrial membrane for the final modifications. This includes critical side-chain alterations, vital oxidation steps, and the culminating insertion of the central iron atom. This constant transmembrane shuttling ensures that the final step of heme biosynthesis occurs exactly where the product is most needed, specifically near the electron transport chain components that rely heavily on functional metalloporphyrins to generate cellular energy.
Slide 4: Step 1 of Heme Biosynthesis: The Rate-Limiting Synthesis of Delta-Aminolevulinic Acid
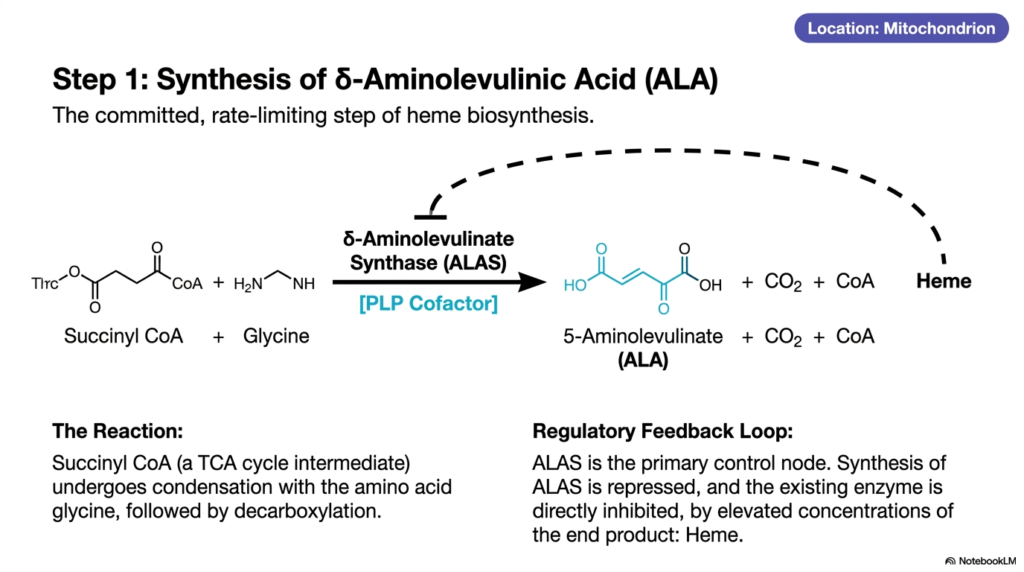
Every grand construction project requires a decisive first step that dictates the pace of the entire build. In metabolic chemistry, this concept is known as the rate-limiting step. The core purpose of this slide deck is to highlight these pivotal regulatory bottlenecks. For medical students, understanding the first reaction in heme biosynthesis is critical, as this single enzymatic step controls the overall rate of production and serves as the primary node for pharmacological intervention and metabolic feedback mechanisms.
The reaction begins inside the mitochondrion, using a key intermediate from the energy-producing tricarboxylic acid cycle, succinyl-CoA. This molecule undergoes a complex condensation reaction with the simple amino acid glycine. The enzyme driving this monumental first step of heme biosynthesis is delta-aminolevulinate synthase, commonly abbreviated as ALAS. Through this highly specific enzymatic union and a subsequent decarboxylation, the molecules merge to form 5-aminolevulinate (also known as delta-aminolevulinic acid, or simply ALA), along with carbon dioxide and free CoA.
To function properly, the ALAS enzyme absolutely requires a specialized chemical assistant. This reaction depends entirely on a cofactor known as pyridoxal phosphate (PLP), an active derivative of vitamin B6. The PLP cofactor forms a temporary chemical bridge that stabilizes the transition state of glycine, thereby allowing it to attack the succinyl-CoA molecule efficiently. Without sufficient vitamin B6, this initial stage of heme biosynthesis halts, leading to systemic energy failure and profound clinical manifestations such as sideroblastic anemia.
Furthermore, this specific reaction features a brilliant regulatory feedback loop. Because it is the committed gateway step, ALAS acts as the primary control node for the entire sequence. As the end product accumulates, it sends a biochemical signal to shut down the factory. High concentrations of the final product physically repress the synthesis of new ALAS enzymes and directly inhibit the activity of existing ones. This elegant feedback mechanism prevents the wasteful and potentially toxic overproduction of intermediates during heme biosynthesis.
Slide 5: Step 2 of Heme Biosynthesis: Condensing into Porphobilinogen and the Threat of Toxicity
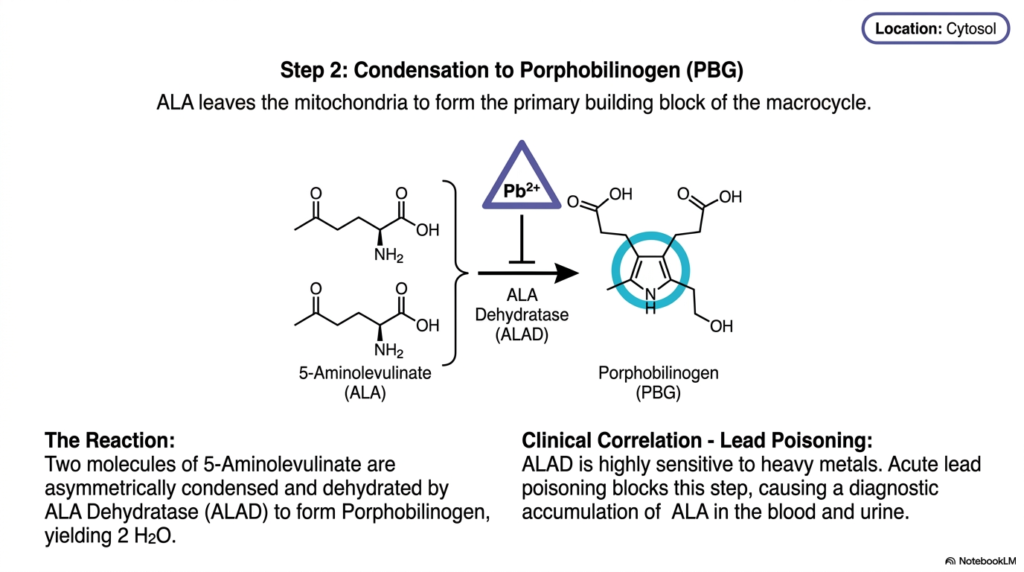
What happens when a delicate biological assembly line is suddenly halted by heavy metal exposure? The consequences are immediate and severe. The core purpose of this slide deck is to trace both the functional assembly of vital molecules and the points of catastrophic failure. For students of pathology, step two of heme biosynthesis is exceptionally important because it bridges the gap between normal metabolic chemistry and the real-world clinical crisis of environmental poisoning, explicitly demonstrating how external toxins disrupt internal equilibrium.
Having been successfully synthesized in the mitochondrion, delta-aminolevulinic acid must now travel out into the cellular cytoplasm. Here, the second major phase of heme biosynthesis commences. Two distinct molecules of 5-aminolevulinate converge and interact with an enzyme called ALA Dehydratase, or ALAD. This enzyme performs an asymmetric condensation, expertly fusing the two precursor molecules while simultaneously dehydrating them, stripping away two water molecules. This specific reaction yields porphobilinogen, a crucial pyrrole ring that serves as the primary building block for the final macrocycle.
Porphobilinogen, abbreviated as PBG, is a singular molecular puzzle piece. It features a five-membered nitrogen-containing ring surrounded by specific side chains. The creation of this distinct molecule marks a monumental shift in heme biosynthesis, transitioning from simple, linear carbon chains into complex cyclic precursors. The cellular machinery will eventually need four of these precise PBG units to stitch together the larger tetrapyrrole cage, making this enzymatic condensation an absolutely vital prerequisite for all subsequent architectural steps.
However, the ALAD enzyme possesses a glaring physiological vulnerability: it is exceptionally sensitive to heavy metals. Acute lead poisoning directly attacks and neutralizes this specific enzyme, physically blocking the active site. When this crucial bottleneck in heme biosynthesis is jammed, the upstream precursor molecules cannot proceed. Consequently, massive amounts of delta-aminolevulinic acid rapidly accumulate within the patient’s bloodstream and urine. For medical professionals, measuring this diagnostic accumulation is the definitive biological footprint used to identify and treat severe heavy metal toxicity.
Slide 6: Steps 3 and 4 of Heme Biosynthesis: The Meticulous Art of Polymerization and Cyclization
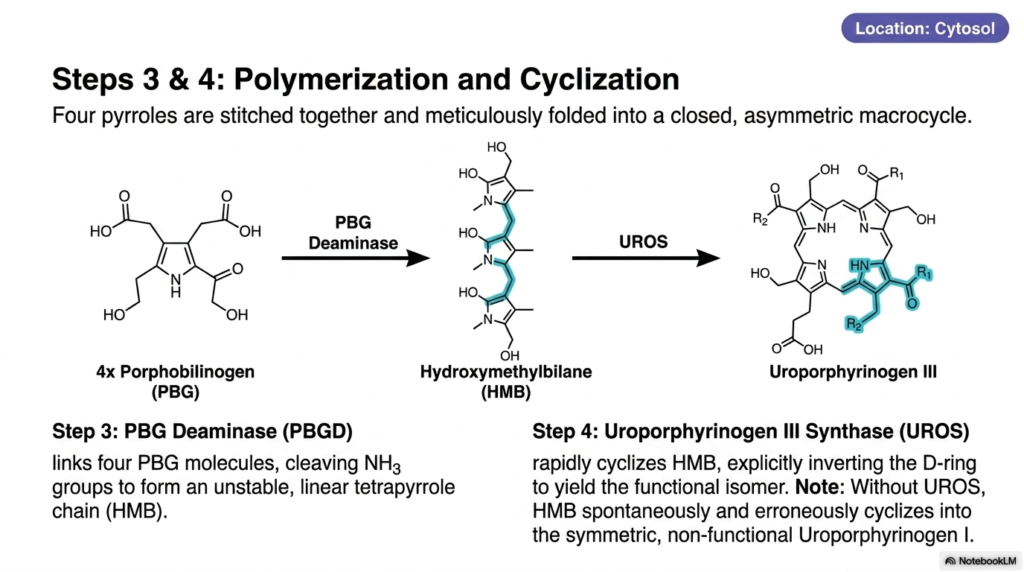
Constructing a closed molecular loop from linear pieces requires absolute precision; a single misplaced chemical bond results in complete biological dysfunction. The core purpose of this slide deck is to illustrate nature’s incredible enzymatic accuracy. Medical and biochemistry students must study steps three and four of heme biosynthesis closely. These stages demonstrate how cellular machinery avoids deadly symmetric errors, relying on exquisite structural inversion to create an asymmetrical ring capable of sustaining complex biological life.
Out in the cytoplasm, the molecular assembly line accelerates. Step three of heme biosynthesis utilizes an enzyme called Porphobilinogen Deaminase. This enzyme is tasked with linking four porphobilinogen molecules. As it stitches these individual pyrrole units into a chain, it actively cleaves off ammonia groups. The result is a highly unstable, linear tetrapyrrole chain known as hydroxymethylbilane. Because this long chemical chain is incredibly fragile, the cell must process it immediately before it degrades or folds incorrectly.
Before the linear chain can spontaneously react, step four of heme biosynthesis takes over. A remarkably fast enzyme, Uroporphyrinogen III Synthase, captures the unstable chain and forces it to cyclize. However, it does not simply snap the ends together. The enzyme explicitly grabs the fourth segment, known as the D-ring, and physically inverts it before closing the loop. This deliberate structural twist is paramount, yielding the uniquely functional, asymmetric isomer known as uroporphyrinogen III.
The necessity of this structural inversion cannot be overstated. If the synthase enzyme is defective or absent, the delicate intermediate chain will spontaneously and erroneously cyclize. This spontaneous reaction creates uroporphyrinogen I, a purely symmetric isomer that is completely biologically useless. This failed product accumulates in the tissues and disrupts the entire flow of heme biosynthesis, leading to devastating congenital diseases. Thus, the deliberate twisting action of the synthase enzyme is a matter of profound cellular survival.
Slide 7: Step 5 of Heme Biosynthesis: Decarboxylation and the Pursuit of Hydrophobicity
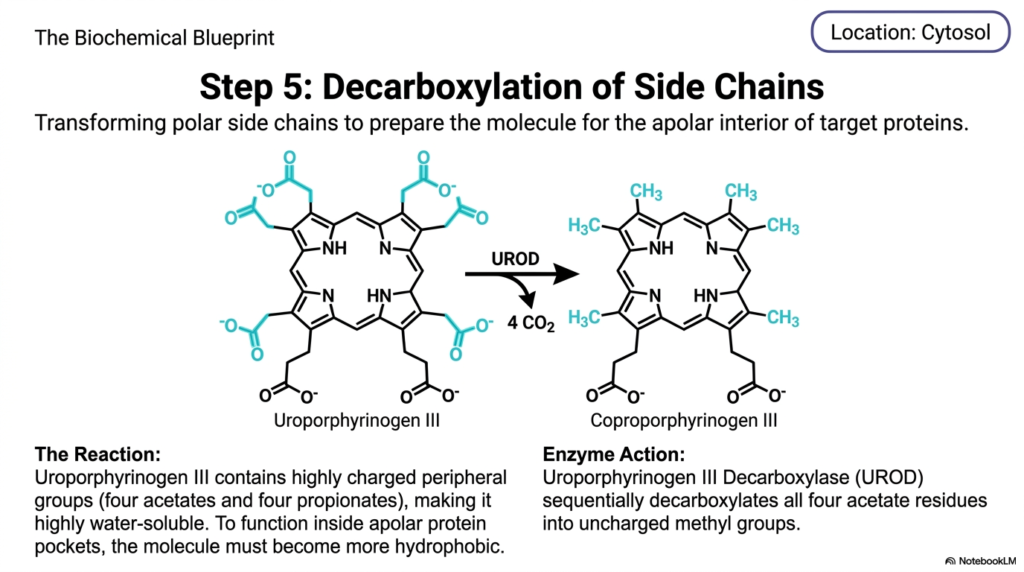
To fit securely into the oily, water-repelling pockets of cellular proteins, a molecule must first shed its water-loving characteristics. The core purpose of this slide deck is to highlight these transformative chemical makeovers. In the context of medical education, this specific stage of heme biosynthesis is a masterclass in solubility modification. Students learn exactly how enzymes systematically strip away charged chemical groups to prepare a highly polar molecule for its final destination within apolar, lipid-rich environments.
At this juncture in the cytoplasm, the developing intermediate is known as uroporphyrinogen III. This specific molecular structure features highly charged peripheral groups jutting out from the central ring, specifically four heavy acetates and four long propionates. Because of these heavily oxygenated, polar appendages, the entire intermediate molecule is exceptionally water-soluble. However, the ultimate functional destination for the finished products of heme biosynthesis lies deep within the hidden, hydrophobic, and apolar interiors of target proteins. Therefore, the intermediate molecule must undergo rigorous chemical tailoring to drastically reduce its polarity and make it much more lipid-soluble.
To achieve this necessary physical transformation, an enzyme named uroporphyrinogen III decarboxylase initiates step five of heme biosynthesis. This enzyme performs a sequential and methodical operation, targeting only the four acetate residues hanging off the perimeter of the macrocycle. By breaking specific carbon bonds, the enzyme liberates four molecules of carbon dioxide. This critical enzymatic action fully converts the charged, water-loving acetate groups into uncharged, non-polar methyl groups, fundamentally altering the molecule’s physical properties.
The direct result of this enzymatic trimming is the creation of coproporphyrinogen III. By eliminating a significant portion of its polar surface area, the molecule becomes far more hydrophobic and robust. This precise transition during heme biosynthesis ensures the intermediate can safely interact with the lipid-dense membranes of cellular organelles. For medical students, understanding this shift in solubility is crucial, as enzymatic failures here lead to the toxic accumulation of water-soluble porphyrins that severely damage the skin and liver.
Slide 8: Step 6 of Heme Biosynthesis: Formation of Vinyl Residues in the Mitochondrion
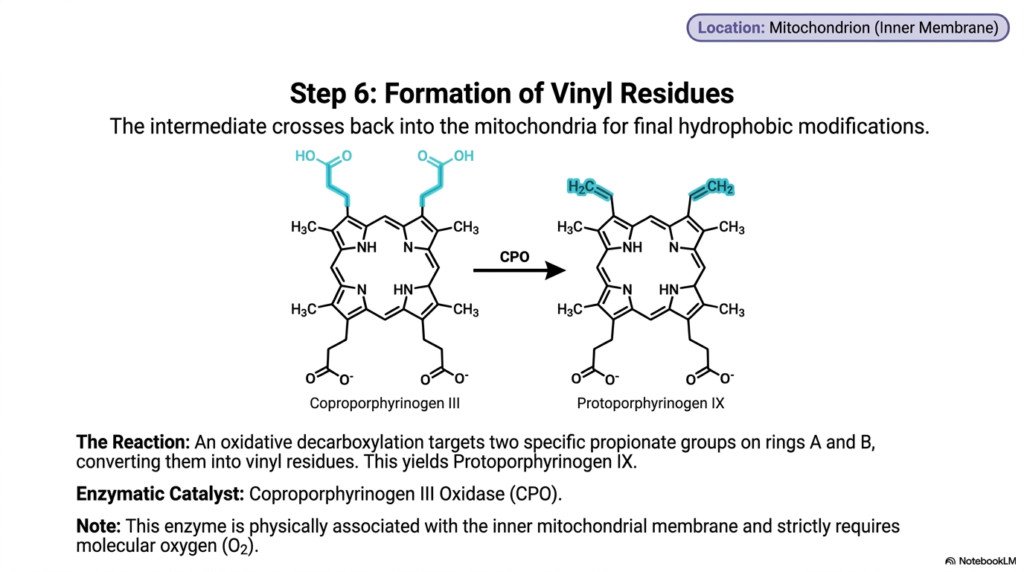
Just as a spaceship must dock with a space station for its final upgrades, metabolic intermediates often journey to specialized cellular membranes for advanced processing. The core purpose of this slide deck is to trace this spectacular biochemical navigation. For aspiring medical professionals, step six of heme biosynthesis is a prime example of organelle-dependent chemistry. It demonstrates how intermediates must physically migrate and interact with specialized mitochondrial enzymes that use ambient oxygen to carry out complex structural refinements.
Having achieved sufficient hydrophobicity in the previous stage, the coproporphyrinogen III molecule leaves the aqueous cytoplasm and crosses back into the mitochondrion. Specifically, it navigates to the highly selective inner mitochondrial membrane, where the next crucial phase of heme biosynthesis occurs. The environment here is drastically different from the cytosol, providing the exact structural support and essential chemical conditions required for advanced oxidation. The molecule must intimately interact with Coproporphyrinogen III Oxidase, an enzyme physically tethered to the mitochondrial architecture.
This specialized oxidase catalyzes an intricate oxidative decarboxylation reaction, targeting highly specific sites on the macrocycle. It selectively focuses on just two of the propionate groups located specifically on rings A and B of the porphyrin structure. By stripping away carbon and utilizing molecular oxygen, the enzyme efficiently converts these two propionate chains into highly reactive vinyl residues. This sophisticated chemical remodeling during heme biosynthesis yields the next vital intermediate, known as protoporphyrinogen IX.
An incredibly important aspect of this specific reaction is its strict requirement for molecular oxygen. Because the oxidase enzyme is completely dependent on oxygen to function, this stage of heme biosynthesis is inextricably linked to the overall aerobic state of the cellular environment. Furthermore, the newly formed vinyl groups add essential structural dynamics to the ring, priming the molecule for the massive electron shifts that occur in the final steps. For researchers, this oxygen dependency represents a critical metabolic checkpoint.
Slide 9: Step 7 of Heme Biosynthesis: Massive Conjugation and the Birth of Pigment
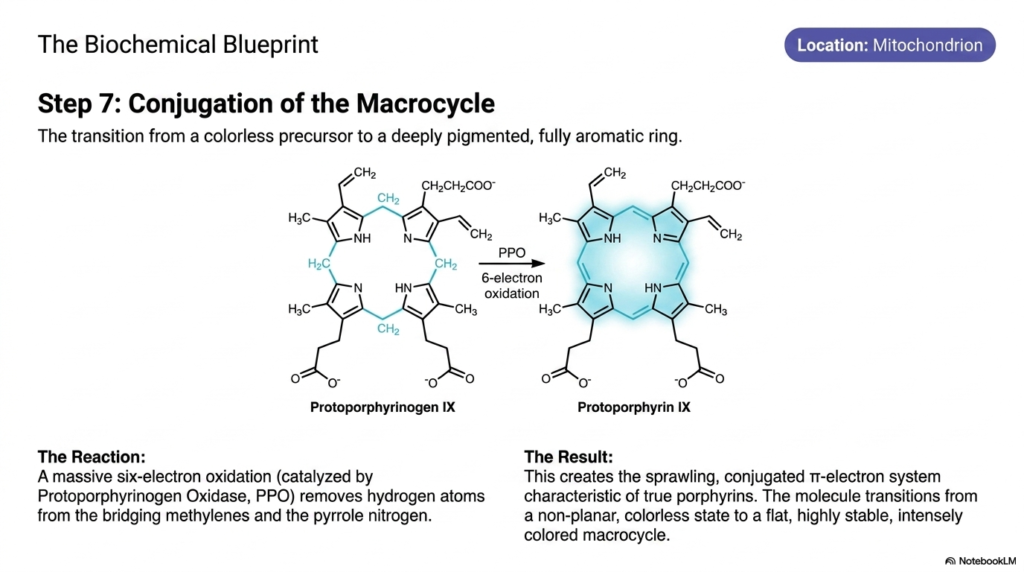
How does a completely colorless chemical precursor suddenly transform into a deeply pigmented molecule capable of sustaining life? The answer lies in the dramatic removal of electrons. The core purpose of this slide deck is to demystify these spectacular energetic transformations. For university students, step seven of heme biosynthesis represents a phenomenal leap in chemical stability and visual biology. It illustrates the exact moment an invisible metabolic intermediate undergoes massive oxidation to become a sprawling, colored, light-absorbing macrocycle.
Still nestled deep within the mitochondrion, the protoporphyrinogen IX molecule undergoes a profound chemical transformation. This specific step of heme biosynthesis is entirely governed by an enzyme called protoporphyrinogen oxidase. The enzyme initiates a massive, coordinated removal of atoms, executing a staggering six-electron oxidation. It aggressively removes crucial hydrogen atoms from both the bridging methylene carbons that connect the separate rings and from the central pyrrole nitrogen atoms, forcing the molecule into a new geometric reality.
The direct biochemical result of this intense enzymatic stripping is the creation of a molecule known as protoporphyrin IX. By ruthlessly removing those specific hydrogen atoms, the enzyme forces the spontaneous creation of alternating double bonds throughout the entire chemical structure. This immediately creates a sprawling conjugated pi-electron system, the ultimate defining characteristic of all true porphyrins produced during late-stage heme biosynthesis. The electrons are now fully delocalized, racing freely around the entire perimeter of the massive molecular cage in a beautifully coordinated fashion.
This profound electron conjugation triggers a spectacular physical transformation. The molecule rapidly shifts from a non-planar, entirely colorless precursor state into a geometrically flat, incredibly stable, and intensely colored macrocycle. This newfound flatness is an absolute mechanical requirement for the imminent insertion of metal. Understanding this dramatic chemical flattening in the final stages of heme biosynthesis allows medical students to appreciate how form dictates function, setting the biological stage for the terminal activation of the entire pathway.
Slide 10: Step 8 of Heme Biosynthesis: The Terminal Insertion of Iron
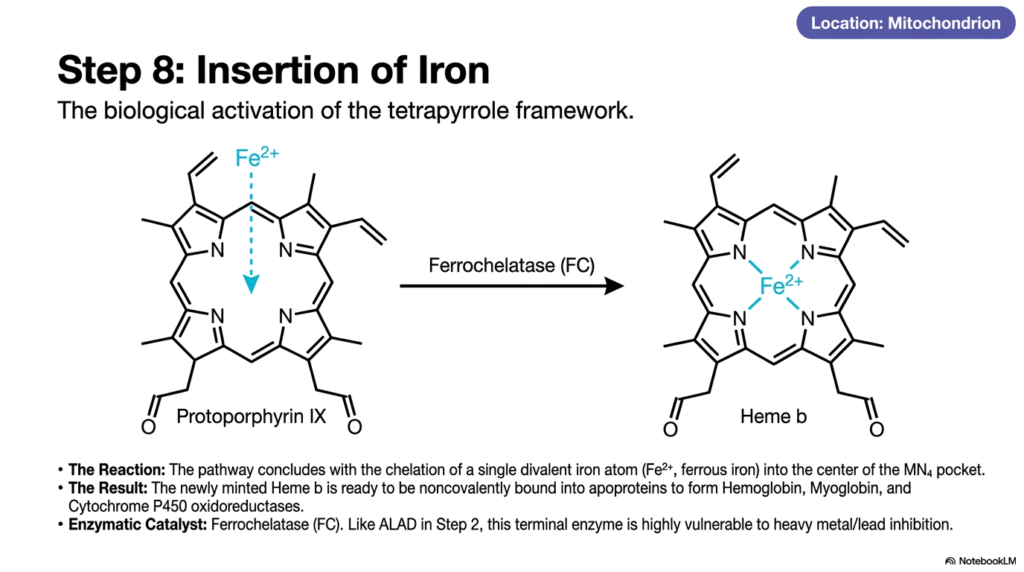
Every great molecular machine requires an engine block to finally bring it to life. In the realm of metabolism, that engine is a single metal atom. The core purpose of this slide deck is to showcase this ultimate biological activation. For future physicians and scientists, the final step in heme biosynthesis is of profound significance. It is the culminating moment when organic architecture captures an inorganic element, forging a biological hybrid molecule that is absolutely essential for human respiration and pharmacological metabolism.
The long, complex journey concludes dramatically within the mitochondria with the precise chelation of a single metal ion. A specialized enzyme known as ferrochelatase orchestrates the final reaction of heme biosynthesis. This enzyme delicately holds the newly flattened protoporphyrin IX ring and facilitates the exact insertion of a single divalent iron atom, specifically in its ferrous state. The iron atom is guided directly into the center of the MN4 pocket, where it binds securely to the inner nitrogen atoms.
This terminal union produces the finished molecule known as Heme b. The biological activation of the tetrapyrrole framework is now fully complete. This newly minted product of heme biosynthesis is entirely primed and ready to be exported and noncovalently bound into massive apoproteins. Once integrated into larger structures, it forms the functional core of vital oxygen-carrying proteins such as hemoglobin and myoglobin, as well as the critically important cytochrome P450 oxidoreductases used for drug metabolism in the liver.
However, students must recognize a critical clinical vulnerability at this terminal juncture. Much like the early stages of the pathway, this final enzyme, ferrochelatase, is exceptionally vulnerable to toxic interference. Heavy metals severely inhibit its activity. When lead poisoning completely blocks this terminal step of heme biosynthesis, iron cannot be inserted. The pathway fails at the very last second, causing massive cellular anemia and a devastating systemic inability to transport oxygen, underscoring the delicate chemical balance of human life.
Slide 11: Beyond Heme Biosynthesis: The Degradation and Bilirubin Pathway
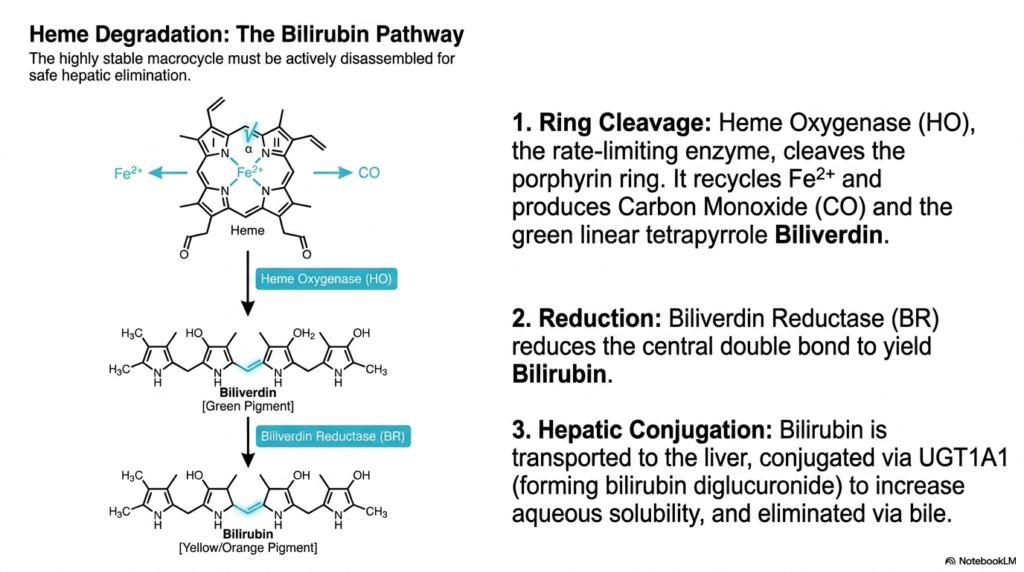
Nature operates on a strict policy of recycling; what takes vast energy to build must eventually be safely dismantled. The core purpose of this slide deck is to provide a comprehensive metabolic overview that inherently includes the breakdown of cellular products. For medical students, looking beyond the construction phase of heme biosynthesis is absolutely necessary. Understanding how the remarkably stable macrocycle is actively disassembled allows future clinicians to diagnose complex liver diseases, jaundice, and hemolytic pathologies resulting from metabolic waste.
Because the tetrapyrrole ring is incredibly robust and highly stable, it cannot simply fall apart on its own. It requires dedicated machinery for active disassembly, distinct from the enzymes used during heme biosynthesis. The process begins with Heme Oxygenase, the primary rate-limiting enzyme of degradation. This enzyme attacks and forcefully cleaves the porphyrin ring. During this destructive event, the central iron atom is safely extracted and recycled for future use, while a surprisingly toxic gas, carbon monoxide, is released as a byproduct.
The physical cleavage of the ring converts the cyclic molecule into a green, linear tetrapyrrole chain known as biliverdin. Immediately following this, an enzyme called Biliverdin Reductase steps in. It chemically reduces the central double bond of the green pigment, transforming it into a yellow-orange pigment known as bilirubin. Unlike the highly water-soluble intermediates generated midway through heme biosynthesis, this resulting bilirubin is highly toxic and insoluble in water, requiring an urgent transport mechanism to prevent systemic neurological damage.
To safely eliminate this hazardous waste product, the insoluble bilirubin binds tightly to carrier proteins in the blood and is transported directly to the liver. Within the hepatic system, a specialized enzyme called UGT1A1 catalyzes the conjugation of bilirubin with glucuronic acid. This vital step significantly increases the compound’s aqueous solubility. Unlike the carefully assembled products of heme biosynthesis, this conjugated waste product is rapidly secreted into the bile duct, eventually exiting the human body safely.
Slide 12: Clinical Pathologies of Heme Biosynthesis: Unmasking the Porphyrias
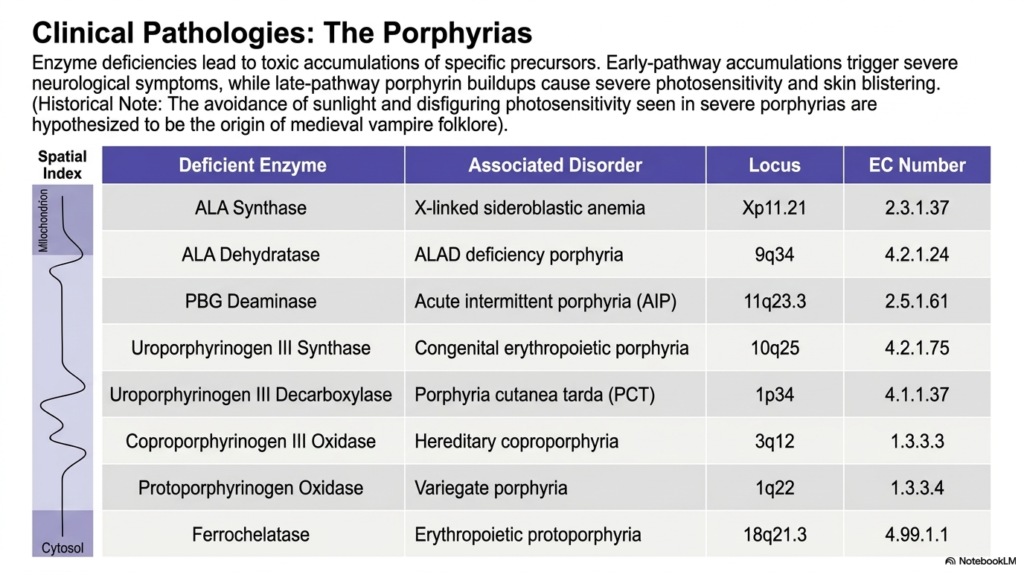
What happens when a single genetic typo derails a massive biological manufacturing plant? The result is an explosive buildup of toxic, unfinished metabolic products. The core purpose of this slide deck is to connect microscopic chemistry directly to observable human disease. For medical trainees, understanding the pathologies associated with defective heme biosynthesis is an essential diagnostic skill. By analyzing the specific enzyme deficiencies known as the porphyrias, students can directly link the accumulation of metabolic intermediates to terrifying neurological and dermatological symptoms.
The slide provides a highly detailed diagnostic table mapping out specific spatial indices, deficient enzymes, and their associated disorders. When a genetic mutation damages an enzyme, the entire progression of heme biosynthesis is drastically compromised. Because the pathway operates like a linear assembly line, a broken machine causes all precursor molecules from earlier steps to accumulate violently within cells and the bloodstream. The exact physical location of this molecular traffic jam dictates the grim clinical presentation seen in patients.
The physiological symptoms are starkly divided by where the error occurs in the sequence. If the enzymatic failure occurs early in the heme biosynthesis pathway, the accumulations are mostly small, neurotoxic precursor molecules such as delta-aminolevulinic acid and porphobilinogen. These specific early-stage buildups rapidly breach the nervous system, triggering severe, acute episodes of agonizing abdominal pain, terrifying hallucinations, and profound psychiatric disturbances. Acute intermittent porphyria is a classic example of this devastating, invisible neurological chaos caused by a truncated pathway.
Conversely, if the enzymatic deficiency strikes much later in the process of heme biosynthesis, completely different symptoms emerge. Late-pathway bottlenecks cause a massive accumulation of fully formed, light-absorbing porphyrin rings in the skin and tissues. When these trapped molecules absorb ambient sunlight, they generate lethal oxygen radicals that physically destroy the surrounding tissue. This results in horrific dermatological photosensitivity, causing severe skin blistering and disfigurement upon sunlight exposure, a tragic phenomenon historically hypothesized to be the real-world origin of ancient vampire folklore.
Slide 13: Harnessing Heme Biosynthesis Intermediates: The Power of Photodynamic Therapy
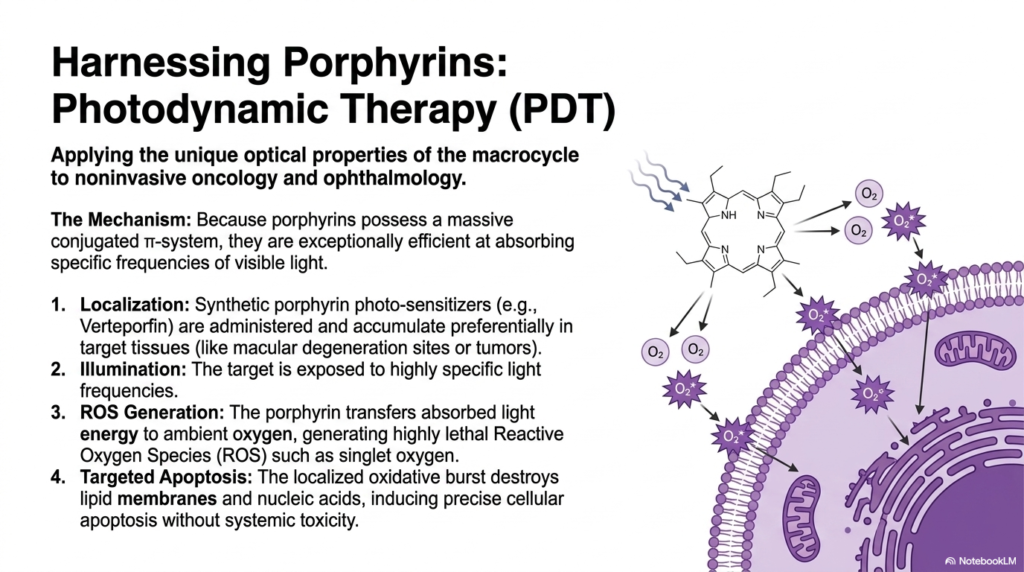
Can humanity weaponize a naturally occurring biological molecule to hunt down and destroy rogue cancer cells? Medical science has achieved exactly that by manipulating the basic laws of light absorption. The core purpose of this slide deck is to show how ancient biological designs can inspire modern medical miracles. College and medical students exploring the visual properties of molecules produced during heme biosynthesis will be fascinated by this clinical application, where intense photochemistry is successfully harnessed for noninvasive oncology and advanced ophthalmology.
The mechanism relies entirely on the unique optical properties of the massive macrocycles formed during the latter stages of heme biosynthesis. Because these porphyrins possess an incredibly large and complex conjugated pi-electron system, they are exceptionally efficient at capturing specific, high-energy frequencies of visible light. In Photodynamic Therapy, or PDT, medical professionals synthesize specific porphyrin photosensitizers in the laboratory. These compounds, such as verteporfin, are carefully administered to the patient, where they preferentially accumulate within highly vascularized target tissues, such as rapidly dividing tumors.
Once the synthetic derivatives of heme biosynthesis have successfully localized inside the targeted disease sites, the physician initiates the attack. The specific tissue is illuminated with a highly calibrated, concentrated beam of light. The trapped porphyrin molecules absorb this massive influx of light energy. However, rather than dissipating this energy harmlessly, the molecule forcefully transfers it to ambient oxygen molecules lingering in the tissue. This violent energy transfer generates an explosive burst of highly lethal reactive oxygen species, primarily singlet oxygen.
This localized oxidative burst acts as a microscopic, targeted bomb. The extremely aggressive reactive oxygen species rapidly rip apart the delicate lipid membranes and nucleic acids of the surrounding diseased cells. This severe internal damage forces the targeted cancer or macular degeneration cells to undergo precise cellular apoptosis, effectively destroying the threat from the inside out. By brilliantly weaponizing the structural legacy of heme biosynthesis, physicians can eradicate localized tumors without inflicting the widespread, catastrophic systemic toxicity typically associated with traditional chemotherapy.
Please read our Content Disclaimer Statement.
Check out our social media channels: