36. Understanding Photosynthesis: Light and Dark Reactions
Imagine a cellular powerhouse capable of pulling invisible gas from the air and transforming it into the physical building blocks of life. This fundamental biological process essentially powers our entire biosphere. The core purpose of this slide deck is to provide college and medical school students with an accessible, detailed roadmap of this intricate molecular machinery. By exploring these mechanisms, students will gain a foundational understanding of how cellular systems generate, transport, and utilize energy in the natural world.
Slide 1: Introduction to Photosynthesis and Energy Transduction

This title slide serves to introduce the overarching biochemical theme of the academic presentation: Photosynthesis is fundamentally an awe-inspiring process of biochemical energy transduction. At its biological core, Photosynthesis represents the single most important metabolic mechanism on Earth, actively converting ephemeral solar energy into tangible organic matter. The presentation aims to demystify this massive biochemical undertaking for advanced university students, breaking down its complexity into digestible visual concepts.
Energy transduction is a thermodynamic principle highly familiar in physics, but in biochemistry, it takes on a beautifully specific meaning. The biological machinery involved does not simply passively absorb light; it actively translates the quantum energy of photons into high-energy chemical bonds. This slide sets the fundamental stage for understanding Photosynthesis as an incredible feat of bioenergetics, successfully bridging the distinct gap between quantum physics and complex organic chemistry.
The mechanistic process detailed extensively throughout this deck will show exactly how solar radiation is systematically captured and creatively utilized. By successfully transducing light, autotrophic organisms are incredibly able to power the otherwise thermodynamically impossible reactions required to synthesize highly complex molecules. Therefore, Photosynthesis acts as the ultimate energetic foundation of nearly all terrestrial and marine food webs across the planet.
For students actively delving into cellular and molecular biology, viewing this process directly through the focused lens of energy conversion is exceptionally critical. It decisively shifts the academic perspective from merely memorizing a simple biological equation to deeply appreciating a sophisticated energy-capturing network. The remaining slides will expand significantly upon this complex transduction, breaking down the exact molecular components directly involved in making this vital conversion entirely possible.
Recognizing the sheer global scale of this biochemical transformation is undeniably awe-inspiring. Millions of these microscopic energy-conversion events happen every single millisecond within a tiny section of a single leaf. The successful biochemical transduction of solar energy into organic matter directly sustains heterotrophic life globally, clearly making it a pivotal area of ongoing study in both modern biochemistry and medicine.
Slide 2: Photosynthesis Compartmentalization and Chloroplast Anatomy
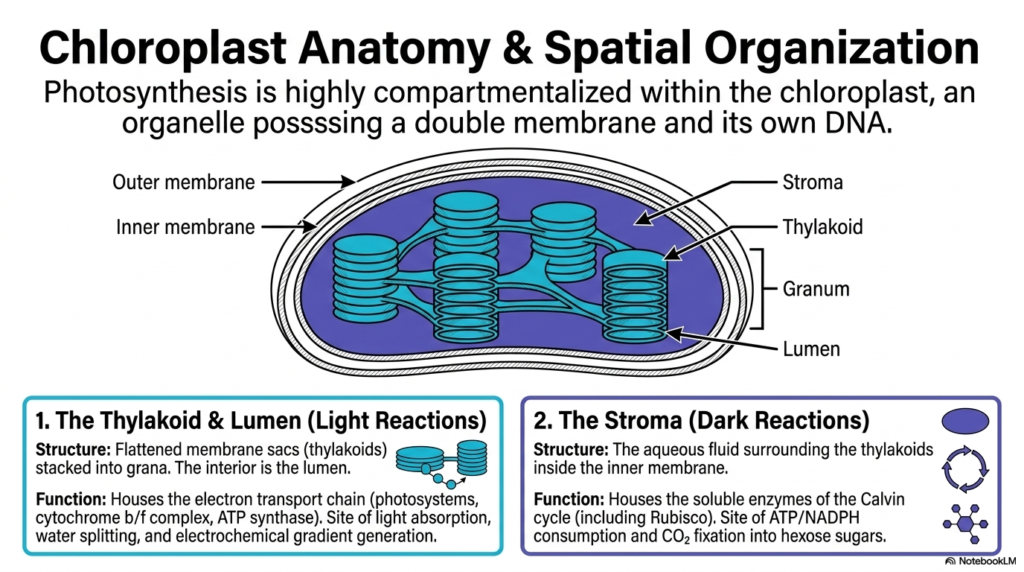
The study of Photosynthesis must begin with a thorough understanding of its physical environment. This initial slide details the highly compartmentalized anatomy of the chloroplast. Photosynthesis requires a precisely structured arena to separate conflicting chemical environments, functioning much like a specialized biochemical laboratory. The chloroplast is an intricate organelle encased in a double membrane and containing its own distinct DNA. This structural complexity allows the organelle to orchestrate delicate biochemical reactions with incredible efficiency and precise regulation.
The physical structural organization perfectly mirrors the division of labor within the complex process of Photosynthesis. The internal architecture reveals flattened membrane sacs known as thylakoids, which are strategically stacked into structures called grana. The thylakoid membrane and its internal space, known as the lumen, serve as the dedicated sites for the light-dependent reactions. Here, the sophisticated electron transport chain operates, capturing solar energy to split water and generate a powerful electrochemical proton gradient.
Conversely, the surrounding aqueous fluid, known as the stroma, serves an entirely different purpose for Photosynthesis. The stroma houses the highly soluble enzymes required for the dark reactions, which include the critical steps of the Calvin cycle. Within this fluid environment, the energetic currencies generated by the thylakoids are actively consumed to fix atmospheric carbon dioxide into stable hexose sugars. This spatial separation ensures that light-driven energy harvesting never interferes with subsequent carbohydrate synthesis.
Understanding this vital spatial organization is strictly essential for medical and biochemistry students aiming to grasp how electrochemical gradients are successfully maintained. The physical boundary provided by the thylakoid membrane allows protons to be forcibly trapped within the lumen. Without this precise physical compartmentalization, the vital biochemical transduction of solar energy would simply dissipate as useless heat, making cellular life utterly impossible.
By analyzing this dual-compartment system, one can genuinely appreciate the evolutionary brilliance of the chloroplast structure. It is not merely a disorganized bag of enzymes, but rather a highly engineered biological organelle. The exact placement of photosystems within the lipid membranes, versus the free-floating nature of the carbon-fixing enzymes in the stroma, dictates the entire rhythmic metabolic flow of the process.
Slide 3: The Chemical Balance and Challenges of Photosynthesis
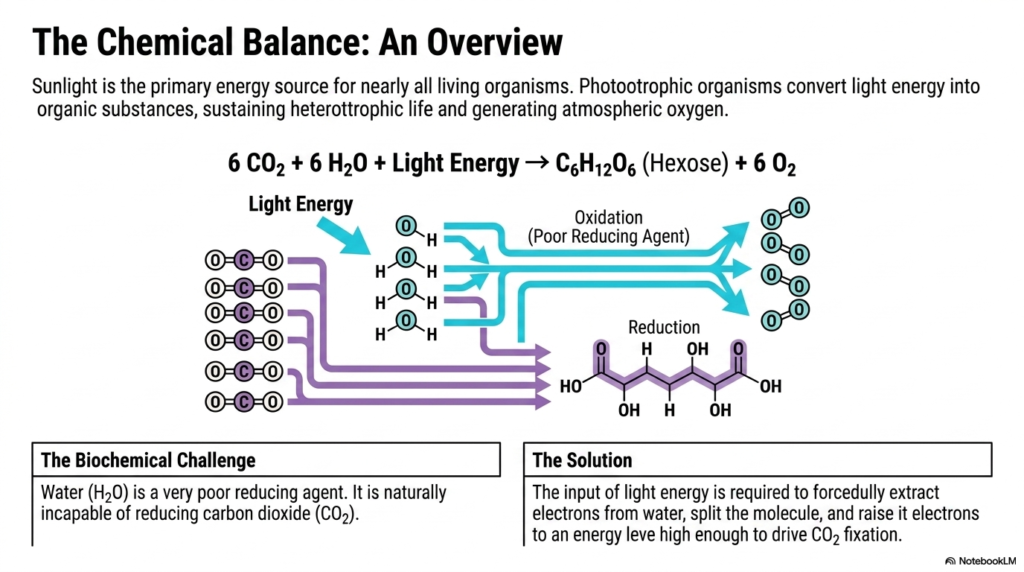
When exploring the chemical balance of life, one must recognize that sunlight is the primary energy source for nearly all living organisms. Photoautotrophic organisms brilliantly convert light energy into complex organic substances, effectively sustaining heterotrophic life while simultaneously generating the atmospheric oxygen we breathe. This slide introduces the fundamental overarching equation of Photosynthesis, highlighting the transformation of carbon dioxide and water into a stable hexose sugar and oxygen.
However, examining this equation reveals a massive biochemical challenge intrinsic to Photosynthesis. Water is remarkably stable and acts as a very poor reducing agent under normal biological conditions. It is naturally entirely incapable of spontaneously reducing carbon dioxide to form carbohydrates. The covalent bonds holding the water molecule together are incredibly tight, meaning that coaxing it to surrender its vital electrons requires a tremendous, targeted energetic push.
The brilliant evolutionary solution to this severe biochemical hurdle lies at the very heart of Photosynthesis. The direct input of intense light energy is strictly required to forcefully extract electrons from the stable water molecules. This light energy is used to physically split the molecule and raise the extracted electrons to an energy level high enough to successfully drive carbon dioxide fixation in later steps.
By breaking down the oxidation and reduction pathways, the visual representation cleanly illustrates the massive flow of electrons. The oxidation of water releases highly necessary protons and electrons, leaving oxygen as a convenient byproduct. Simultaneously, the difficult reduction of carbon dioxide forms the highly structured, energy-rich carbon skeleton of the resulting hexose sugar molecule.
Understanding this difficult, energetic uphill battle is completely crucial for any advanced biochemistry student. It highlights exactly why light is not just a passive signal but an active chemical reactant acting as the driving force. Without the continuous immense input of solar energy, the critical reduction of atmospheric carbon dioxide would simply remain a thermodynamic impossibility, bringing the biological carbon cycle to a sudden halt.
Slide 4: Chlorophyll Structure in Photosynthesis
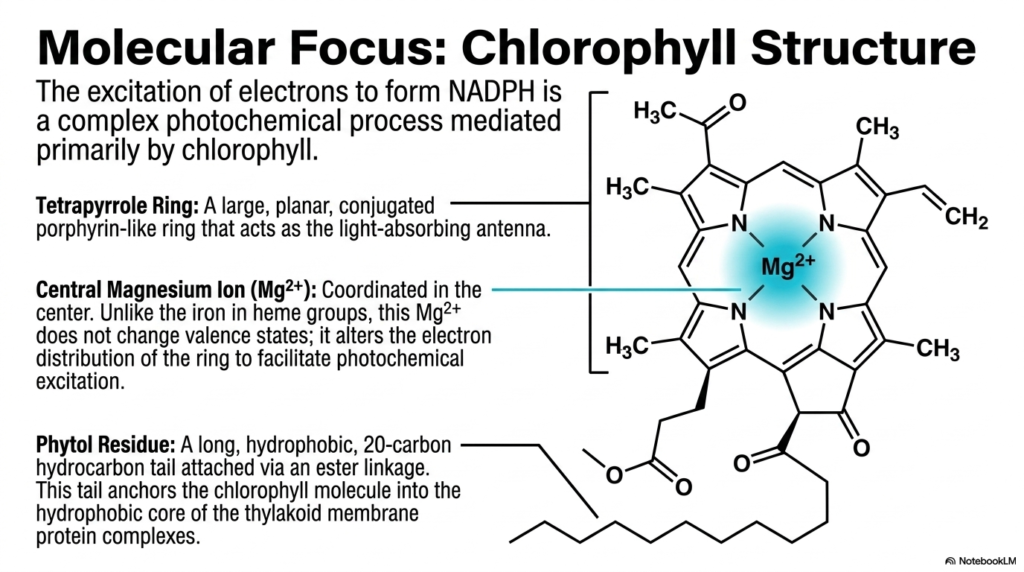
The actual excitation of electrons necessary to eventually form NADPH is a highly complex photochemical process. This intricate process within Photosynthesis is mediated primarily by a remarkable green pigment known as chlorophyll. This slide expertly details the highly specialized molecular structure of chlorophyll, revealing precisely how its physical anatomy dictates its unique biological function in capturing fleeting light energy.
The most prominent feature is the large tetrapyrrole ring, which serves as the primary light-absorbing antenna in photosynthesis. This expansive, planar, conjugated porphyrin-like ring features alternating single and double bonds, creating a delocalized electron cloud. This specific delocalization makes it incredibly efficient at absorbing specific wavelengths of visible solar light and entering a highly energetic excited state upon photon absorption.
At the very center of this planar ring sits a crucial, coordinated magnesium ion. Unlike the central iron atom typically found in blood heme groups, this specific magnesium ion purposefully does not change its valence states during the chemical process. Instead, its primary biochemical role is to actively alter the delicate electron distribution across the entire ring, thereby dramatically facilitating the necessary photochemical excitation when a quantum of light strikes.
Anchoring this powerful molecular antenna is the long, highly hydrophobic phytol residue. This twenty-carbon hydrocarbon tail is firmly attached via a specialized ester linkage to the main porphyrin structure. The primary function of this hydrophobic tail is to securely anchor the entire chlorophyll molecule deep into the incredibly hydrophobic lipid core of the thylakoid membrane protein complexes.
The beautiful synergy of these structural components ensures that Photosynthesis operates with maximal physical efficiency. The antenna portion is optimally positioned to capture light, while the phytol tail ensures the molecule remains firmly embedded exactly where fast-paced electron transfer must occur. Students must closely appreciate this structural biology to understand how perfectly form meets function in this vital energy-harvesting apparatus.
Slide 5: Matrix of Light vs. Dark Reactions in Photosynthesis

To fully comprehend the massive scope of the process, Photosynthesis is elegantly divided into two tightly coupled, mutually dependent metabolic phases. This incredibly helpful matrix neatly contrasts the differences between the light and dark reactions. While they occur sequentially, understanding their distinct physical locations, specific inputs, and thermodynamic properties is vital to mastering the overall biochemical pathway.
The light reactions, also known strictly as the photochemical phase of Photosynthesis, occur specifically within the thylakoid membrane and its internal lumen. These reactions are strongly endergonic, meaning they absolutely require an active input of energy, which is generously provided by incoming solar light. Using water, light energy, NADP+, and ADP, this incredibly dynamic phase generates atmospheric oxygen and the highly energetic molecules ATP and NADPH.
The core biochemical function of these photochemical reactions is to forcefully split water to capture excited electrons and strategically generate a steep proton gradient. The generated energy currencies are precisely what power the second half of Photosynthesis. Without the light reactions continuously running and supplying these volatile, high-energy intermediates, the subsequent carbon-fixing machinery would quickly grind to an absolute, unceremonious halt.
In stark contrast, the dark reactions, formally known as the Calvin Cycle, occur entirely in the aqueous chloroplast stroma. These reactions are thermodynamically driven completely by the rapid hydrolysis of the previously generated ATP and the oxidation of NADPH. By taking in atmospheric carbon dioxide alongside these energy currencies, the complex stroma machinery patiently synthesizes stable hexose sugars.
This matrix makes it abundantly clear that the core function of the dark reactions is to fundamentally fix atmospheric carbon into incredibly stable organic carbohydrate bonds. It is important to note that if adequate ATP, NADPH, and the appropriate enzymes are readily available, these specific reactions can proceed perfectly well in the total absence of light, highlighting their functional independence.
Slide 6: The Z-Scheme and Energetics of Photosynthesis
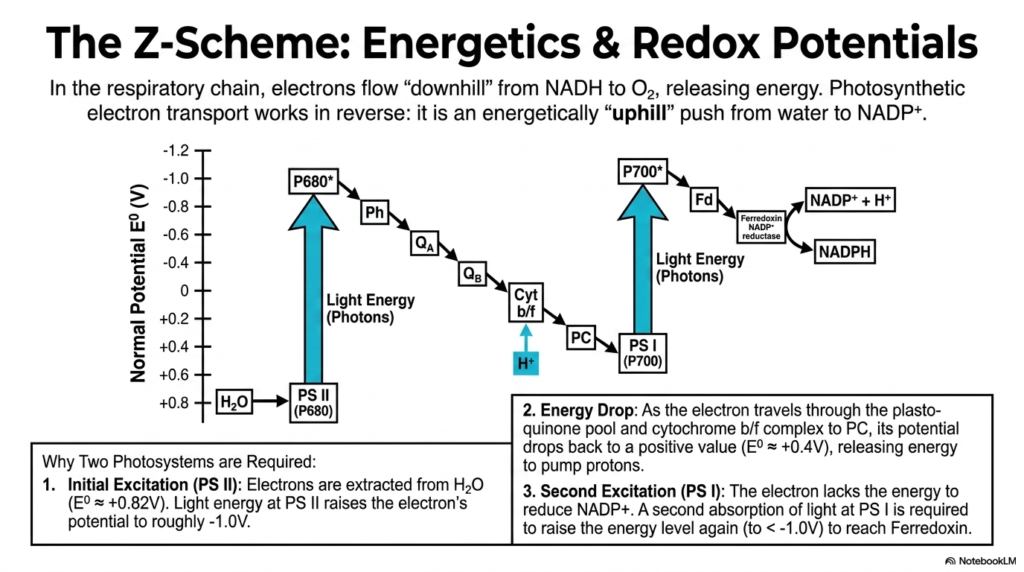
Energetics govern every biological process, and the classic Z-Scheme beautifully illustrates the extreme thermodynamic journey of an electron during Photosynthesis. In a standard respiratory chain, electrons flow easily “downhill” from molecules such as NADH to oxygen, thereby naturally releasing energy. However, photosynthetic electron transport miraculously works in direct reverse, requiring a massive, energetically “uphill” push to move electrons from stable water to NADP+.
The complex visual graph perfectly tracks the normal electrical potential as the electrons boldly traverse the sophisticated machinery of Photosynthesis. The essential journey begins with a massive initial excitation at Photosystem II. Electrons are forcefully extracted from water at a highly positive electrical potential and are suddenly violently raised to an incredibly negative, high-energy state by the forceful absorption of light quanta.
Following this dramatic initial excitation, a significant energy drop naturally occurs. As the highly energetic electron sequentially travels through the mobile plastoquinone pool and the integral cytochrome b/f complex, its redox potential gradually returns to a relatively positive value. This specific downward slide is not wasted; instead, it aggressively releases vital kinetic energy, which is deliberately used to pump protons.
However, the exhausted electron now lacks sufficient energy to reduce NADP+. Therefore, a critical second excitation phase specifically at Photosystem I is biologically required to make Photosynthesis functionally complete. A second targeted absorption of solar light successfully raises the electron’s specific energy level once again, forcefully propelling it high enough to finally reach the terminal carrier, ferredoxin.
This distinctive zigzag shape visually explains exactly why two separate, dedicated photosystems are absolutely required in sequence. A single quantum of light simply cannot provide enough direct energetic thrust to both strip an electron from stable water and successfully push it all the way to NADP+. This slide definitively clarifies the complex energetic choreography underpinning this remarkable life-giving biochemical process.
Slide 7: Structure of Photosystem II in Photosynthesis
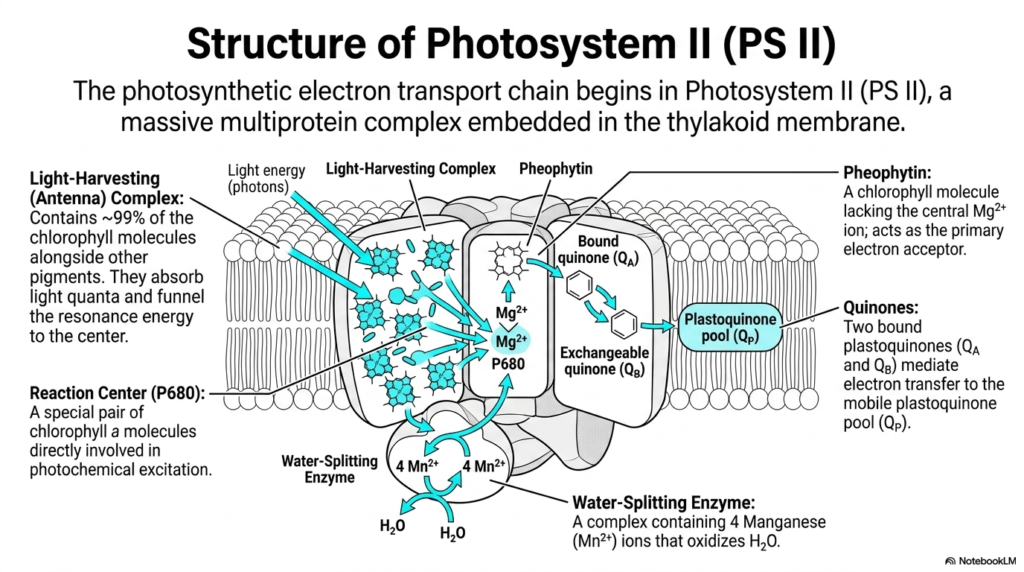
The incredibly complex photosynthetic electron transport chain officially begins its journey deep within Photosystem II. This slide offers a highly detailed view of this massive, multi-protein complex that sits securely embedded within the vital thylakoid membrane. Photosystem II is the biochemical engine of Photosynthesis, the exact site where incoming light energy first makes its decisive chemical impact.
The structure features an expansive light-harvesting antenna complex that brilliantly contains roughly ninety-nine percent of the total chlorophyll molecules. These tightly packed accessory pigments eagerly absorb incoming light quanta and quickly and efficiently funnel the resulting resonant energy directly toward the very center. This acts precisely like a biological satellite dish, aggressively concentrating diffuse incoming signals into one single, highly focused focal point.
At this critical focal point lies the specialized reaction center, widely known as P680. This is a unique pair of targeted chlorophyll a molecules that are directly and intimately involved in the primary photochemical excitation event of Photosynthesis. When the concentrated resonant energy finally reaches P680, an electron is violently promoted to an excited state, rapidly initiating the critical sequential electron-transfer cascade.
Surrounding the central reaction center are several absolutely vital specialized electron acceptors. Pheophytin, which is essentially a specialized chlorophyll molecule conveniently lacking the central magnesium ion, acts as the very first primary electron acceptor. From there, the excited electron is passed to tightly bound quinones, which actively mediate the crucial transfer to the mobile, highly hydrophobic plastoquinone pool lingering nearby.
Crucially, the massive complex also securely houses the powerful water-splitting enzyme containing four essential manganese ions. Because P680 becomes incredibly positively charged after rapidly losing an electron, it becomes the strongest known biological oxidizing agent. It violently rips replacement electrons directly from nearby water molecules, resulting in the life-sustaining biological release of atmospheric oxygen gas.
Slide 8: The Quantum Steps of Photosynthesis at PS II
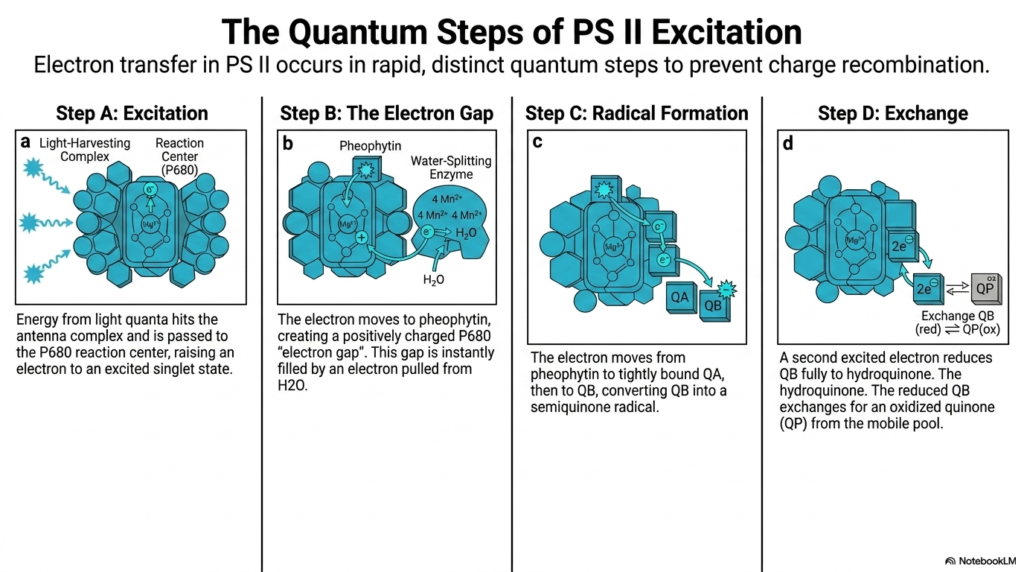
To truly grasp the phenomenal efficiency of Photosystem II, one must closely examine the rapid quantum steps of electron transfer. If the violently excited electron immediately fell back to its original state, the captured energy of Photosynthesis would be lost entirely as useless heat or fluorescence. Therefore, electron transfer rigidly occurs in distinct, rapid quantum steps to strictly prevent wasteful charge recombination.
The sequential process officially begins with Step A, which is the initial rapid excitation phase. Ambient energy from incoming light quanta collides violently with the massive antenna complex and is quickly transferred to the sensitive P680 reaction center. This instantly raises a single-target electron to an extremely excited singlet state, ready to leave the central porphyrin ring and begin its important journey.
Almost instantaneously, Step B initiates the crucial creation of the electron gap. The highly excited electron physically moves rapidly to the nearby waiting pheophytin molecule. This specific movement violently creates a highly positive, desperate “electron gap” back at the P680 center, a gap so powerfully oxidizing that it is instantly, forcefully filled by a reluctant electron ripped violently from a neighboring water molecule.
Following this, Step C dictates the subsequent radical formation necessary for Photosynthesis. The captured electron rapidly moves from the waiting pheophytin molecule to the tightly bound quinone, QA, and then to QB. This specific transfer momentarily converts the waiting QB molecule into a highly reactive semiquinone radical, holding the specific charge safely in place without losing it.
Finally, Step D illustrates the vital chemical exchange phase. A second newly excited electron rapidly travels down the identical pathway, ultimately reducing the waiting QB completely to a stable hydroquinone state. This fully reduced QB rapidly exchanges physically with an entirely oxidized quinone directly from the mobile plastoquinone pool, effectively carrying the captured solar energy safely away for the next steps.
Slide 9: The Electron Transport Chain in Photosynthesis
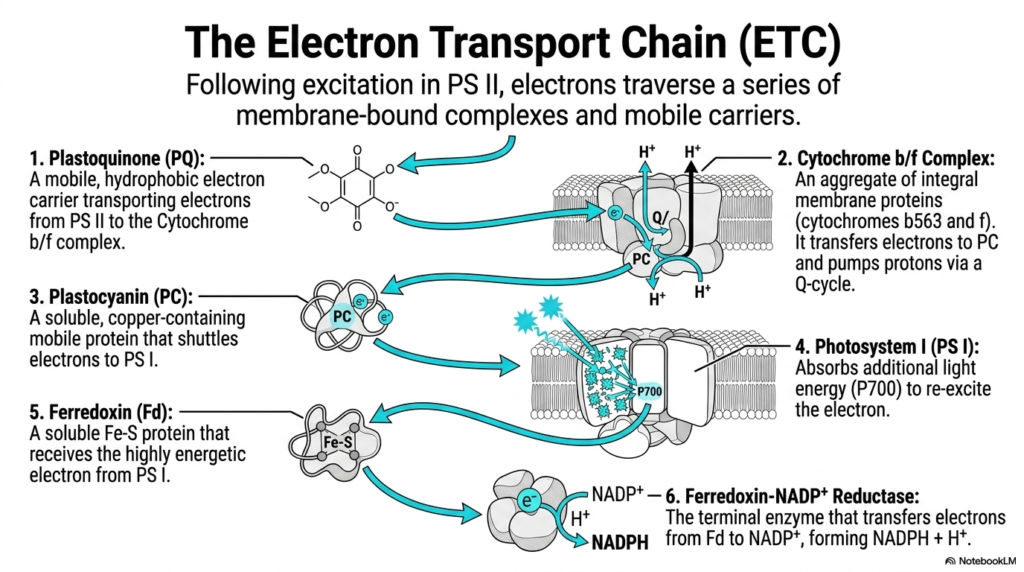
Following the initial dramatic excitation in Photosystem II, the highly energetic electrons must actively traverse a complex series of specialized carriers. The electron transport chain in photosynthesis brilliantly uses both stationary membrane-bound complexes and highly mobile carriers to shuttle electrons efficiently. This slide expertly maps the exact sequence of these vital transfers, highlighting the massive interconnected biochemical highway embedded within the thylakoid membrane.
The vital journey continues specifically with the mobile, highly hydrophobic plastoquinone pool. These specialized lipid-soluble molecules actively transport captured electrons safely away from Photosystem II directly to the waiting Cytochrome b/f complex. The Cytochrome b/f complex is a massive aggregate of integral membrane proteins that actively transfers electrons while simultaneously pumping critical protons forcefully across the membrane via an intricate Q-cycle.
Once the electrons successfully exit the massive cytochrome complex, they are quickly picked up by plastocyanin. Plastocyanin is a small, highly soluble, copper-containing mobile protein that efficiently shuttles the now low-energy electrons directly over to Photosystem I. This incredibly efficient biological relay race ensures that electrons flow seamlessly between the massive, stationary transmembrane complexes without ever being accidentally lost to the surrounding environment.
At Photosystem I, the exhausted electrons finally receive a necessary second burst of incoming solar energy. This specific complex forcefully absorbs additional light energy at the P700 wavelength, violently re-exciting the arriving electron. This crucial secondary energetic boost in Photosynthesis is mandatory to provide the electron with the phenomenal reducing power strictly required for the final, difficult biochemical synthetic steps.
The heavily energized electron is then passed to ferredoxin, a highly soluble iron-sulfur protein. Finally, the critical terminal enzyme, known as Ferredoxin-NADP+ Reductase, successfully catches these highly energetic electrons. It actively transfers them from ferredoxin directly to the waiting NADP+ molecule, successfully forming the highly stable, energy-dense NADPH required for the upcoming carbon-fixing dark reactions.
Slide 10: Chemiosmosis and the Gradient in Photosynthesis
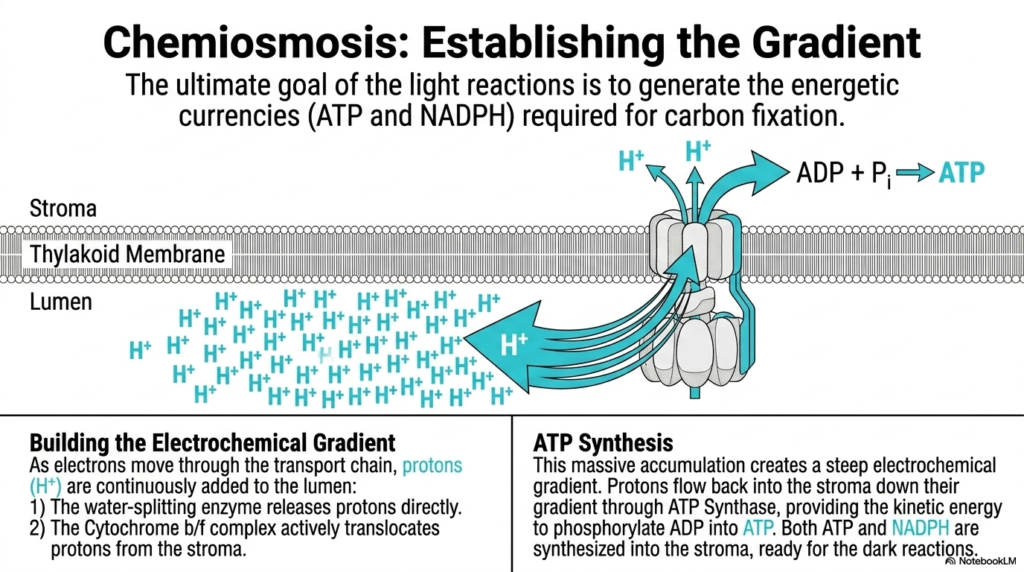
The ultimate biological goal of the light-dependent reactions in Photosynthesis is to generate essential energy currencies. ATP and NADPH are strictly required to power the subsequent difficult carbon fixation steps. This slide brilliantly illustrates the phenomenon of chemiosmosis, demonstrating exactly how the frantic movement of electrons is coupled to the massive accumulation of highly concentrated protons.
As excited electrons rapidly move through the incredibly complex electron transport chain, protons are continuously, aggressively added specifically to the confined internal thylakoid lumen. First, the incredibly violent water-splitting enzyme physically releases its protons directly into this tightly enclosed space. Second, the massive Cytochrome b/f complex actively and continuously translocates additional protons specifically from the outer stroma directly into the confined lumen.
This massive, continuous physical accumulation of positive charges creates an incredibly steep, volatile electrochemical gradient directly across the thin thylakoid membrane. The high concentration of trapped protons in the tiny lumen passionately seeks to flow back out into the stroma to restore chemical and electrical equilibrium. This stored potential energy is the true biological battery powering the incredible engine of Photosynthesis.
The brilliant resolution to this extreme electrical tension is the massive ATP Synthase complex. As the trapped protons eagerly flow forcefully back into the stroma down their massive gradient, they literally spin the mechanical rotor of the ATP Synthase enzyme. This remarkable kinetic movement directly provides the necessary energy to forcefully phosphorylate ambient ADP into stable, energy-rich ATP.
Consequently, through this brilliant mechanism of chemiosmosis, both vital ATP and high-energy NADPH are successfully synthesized directly into the awaiting stroma. They are now perfectly positioned and completely ready to actively power the complex, demanding dark reactions. This beautiful coupling of electron flow to proton pumping is a masterclass in elegant biochemical energy conversion.
Slide 11: Dynamic Routing and Electron Flow in Photosynthesis
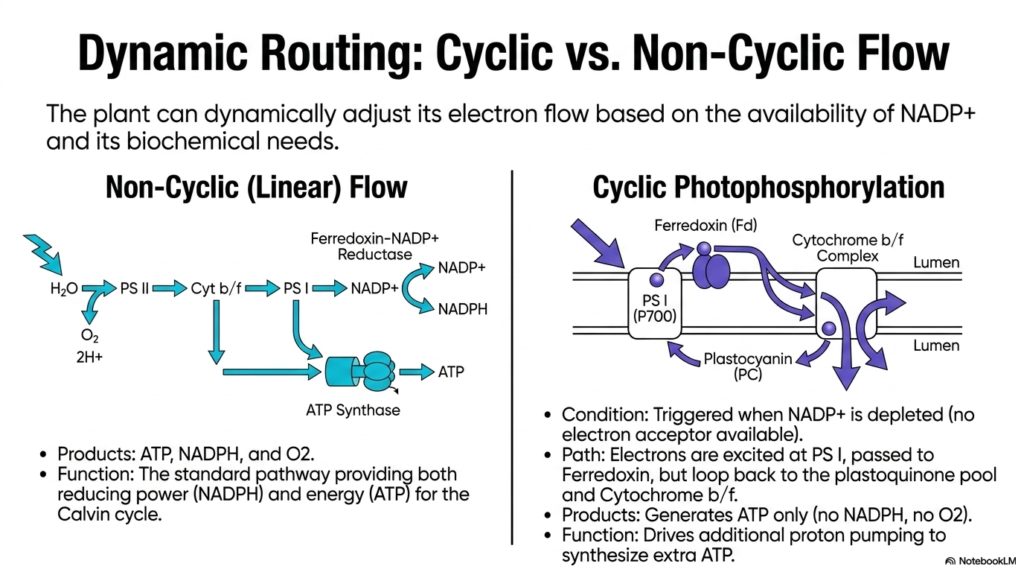
Plants and autotrophic organisms possess a remarkable biological ability to dynamically adapt to varying metabolic conditions. The complex machinery of Photosynthesis can dynamically adjust its electron flow based on the current cellular availability of NADP+ and shifting biochemical needs. This vital slide vividly contrasts the standard non-cyclic linear flow with the incredibly adaptive, specialized process of cyclic photophosphorylation.
The standard linear flow is the most commonly recognized pathway in Photosynthesis. Electrons travel cleanly from water through Photosystem II, down the cytochrome complex, into Photosystem I, and finally neatly terminate at the NADP+ reductase enzyme. This highly standard pathway reliably produces a balanced ratio of ATP, NADPH, and oxygen, beautifully providing both the reducing power and strict energy required for the Calvin cycle.
However, cellular conditions constantly change, and the dark reactions occasionally consume significantly more ATP than NADPH. When ambient NADP+ is heavily depleted and strictly unavailable to act as an electron acceptor, the ingenious cyclic photophosphorylation pathway is instantly triggered. In this highly specialized condition, the excited electrons actively bypass their normal terminal destination to purposefully prevent dangerous cellular blockages.
During this specific cyclic path, electrons are violently excited at Photosystem I and successfully passed to ferredoxin as normal. However, instead of reducing NADP+, they loop entirely backward to the mobile plastoquinone pool and pass through the Cytochrome b/f complex once again. This massive biological detour elegantly forces the active machinery to continuously pump more protons without ever generating any extra NADPH.
The brilliant function of this cyclic routing within Photosynthesis is to aggressively drive additional proton pumping, thereby synthesizing the extra ATP needed to successfully balance the skewed cellular ratios. It is a stunning display of metabolic flexibility, ensuring the overall biochemical factory never fundamentally stalls simply because one specific energetic currency temporarily outpaces the specific production of the other.
Slide 12: The Calvin Cycle in Photosynthesis
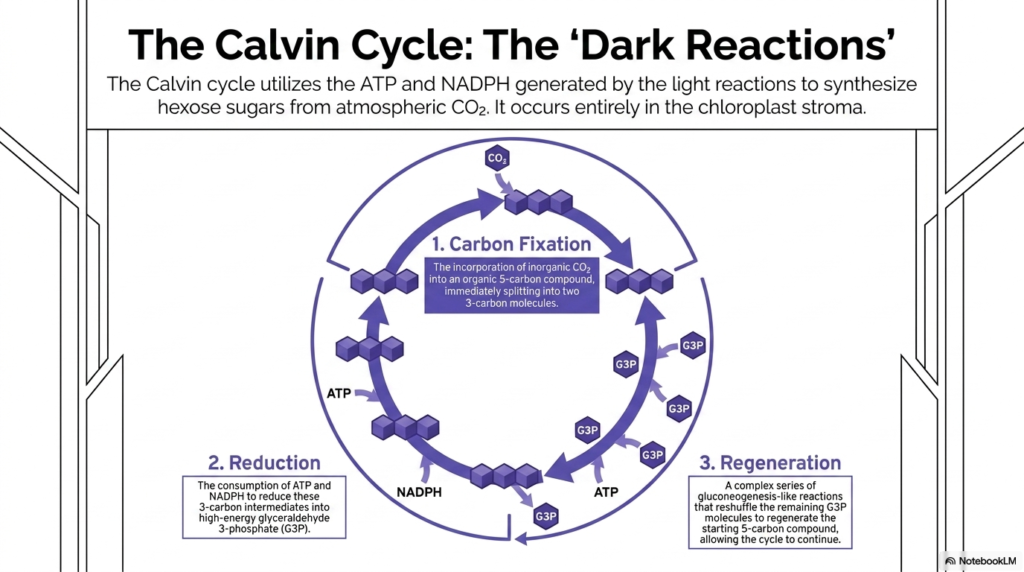
Moving away from the energetic light reactions, the biochemical focus now dramatically shifts to the incredible synthetic power of the dark reactions. The legendary Calvin Cycle beautifully utilizes the heavily concentrated ATP and highly energetic NADPH previously generated by the light reactions. Its ultimate metabolic goal within Photosynthesis is to synthesize highly complex hexose sugars entirely from simple, inert atmospheric carbon dioxide.
It is biochemically vital to clearly note that this highly intricate cyclic process occurs entirely free-floating within the aqueous fluid of the chloroplast stroma. The complete cycle is traditionally divided into three highly distinct, critical metabolic phases: carbon fixation, chemical reduction, and molecular regeneration. These three phases must operate in perfect synchronization to ensure the continuous, uninterrupted biological production of vital, stable carbohydrates.
The very first specific phase is carbon fixation, a truly monumental biochemical step in Photosynthesis. This critical step successfully incorporates completely inorganic atmospheric carbon dioxide gas directly into an existing, stable organic five-carbon molecule. This highly unstable, newly formed six-carbon intermediate violently splits into two highly stable three-carbon molecules, permanently trapping the free gas within the biological realm.
The subsequent vital phase is the heavy reduction step. Here, the previously stored chemical energy from the massive light reactions is finally aggressively spent. The cycle rapidly consumes massive amounts of stored ATP alongside the reducing power of NADPH to forcefully reduce these simple three-carbon intermediates. This incredibly heavy energetic investment successfully transforms them into highly energetic glyceraldehyde 3-phosphate molecules.
Finally, the highly complex regeneration phase ensures that the cycle can persist continuously. A massive, complex series of difficult gluconeogenesis-like enzymatic reactions successfully reshuffles the vast majority of the remaining intermediate molecules. This heavy reshuffling successfully regenerates the exact starting five-carbon compound, allowing the massive wheel of Photosynthesis to turn endlessly, continually fixing more gas into stable solid matter.
Slide 13: Carbon Fixation and Rubisco in Photosynthesis
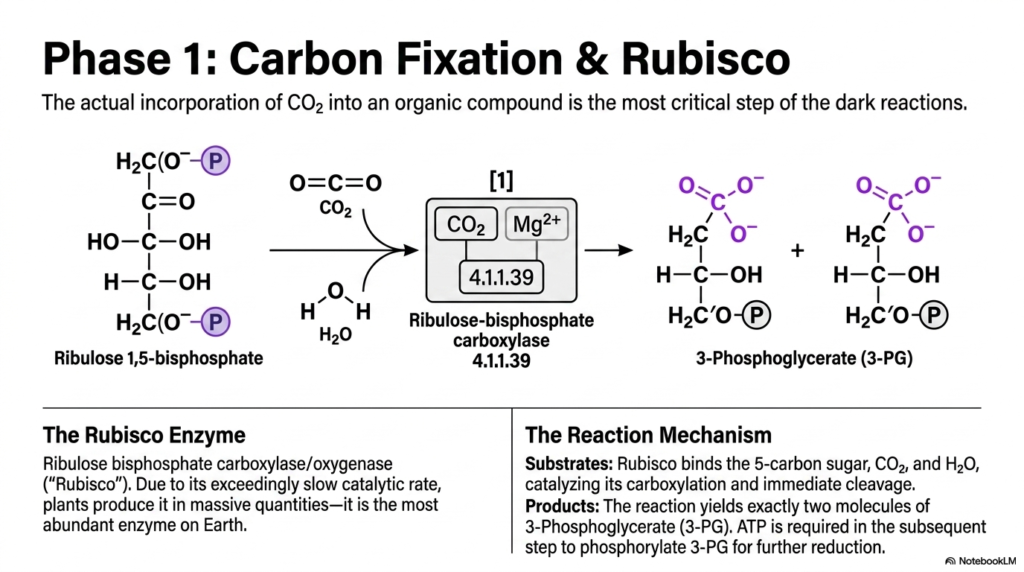
The actual successful incorporation of inert atmospheric carbon dioxide into a living organic compound is arguably the most biologically critical single step of the dark reactions. This particular slide zooms in incredibly closely on the precise molecular mechanics of phase one of the Calvin cycle. Understanding this highly specific chemical pathway is essential for students studying the broader macro-scale impacts of Photosynthesis on global carbon cycles.
The central reacting substrate is a dedicated five-carbon sugar known precisely as Ribulose 1,5-bisphosphate. When a single molecule of free carbon dioxide and a single molecule of water are successfully introduced, they combine with this specific substrate. This incredibly specific reaction seamlessly merges inorganic gas into the existing organic biological carbon skeleton, officially initiating the difficult, energy-intensive process of producing stable life-sustaining sugars.
This monumental biological task is actively catalyzed exclusively by the massive Rubisco enzyme, formally known as Ribulose bisphosphate carboxylase/oxygenase. Surprisingly, despite its incredible importance to Photosynthesis, Rubisco is notorious for possessing an exceedingly slow, clumsy catalytic rate. To actively compensate for this severe molecular inefficiency, autotrophic plants are forced to produce it in absolutely massive, staggering cellular quantities.
Consequently, Rubisco is widely considered by biochemists to be the single most abundant biological enzyme present on planet Earth. Its highly specific reaction mechanism dictates that it must carefully bind the five-carbon sugar, capture ambient carbon dioxide, and actively catalyze both its immediate carboxylation and subsequent cleavage. It is an incredibly heavy molecular burden placed squarely upon a surprisingly inefficient biological protein complex.
The immediate terminal products of this vital reaction are exactly two distinct molecules of 3-Phosphoglycerate. However, the vital biochemical process is far from entirely complete at this stage. Massive amounts of energetic ATP are immediately strictly required in the very next immediate step to forcefully phosphorylate these newly formed stable molecules for the severe further reduction reactions required in Photosynthesis.
Slide 14: The Stoichiometry of Sugar Synthesis in Photosynthesis
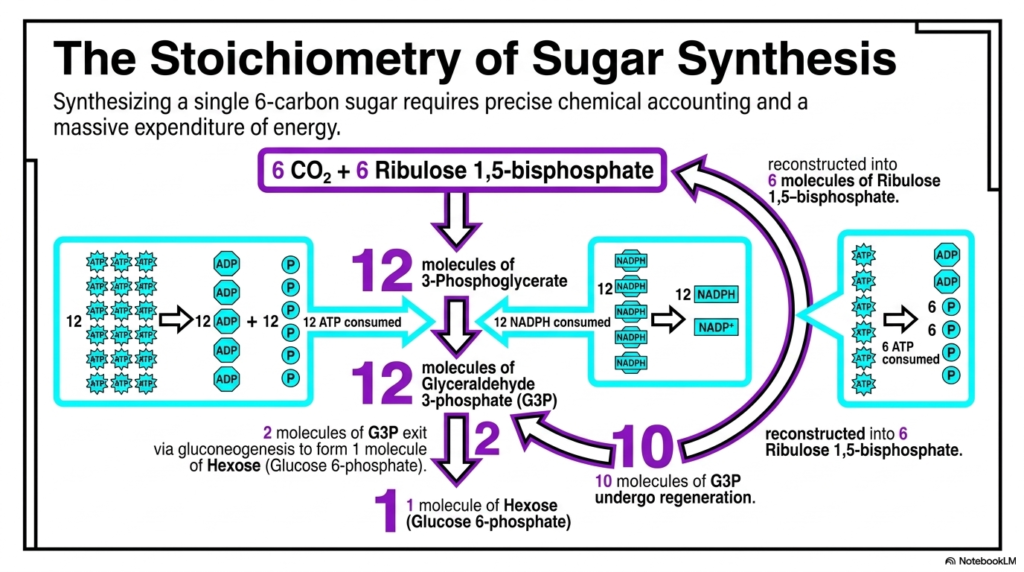
Biochemistry is fundamentally an exact science of strict chemical accounting, and synthesizing even a single stable six-carbon sugar requires precise energetic mathematics. This slide beautifully breaks down the staggering, massive expenditure of cellular energy strictly required to successfully complete the dark reactions. The overall stoichiometry of Photosynthesis reveals exactly how incredibly energetically expensive it is to forcefully build highly complex organic molecules from scratch.
To successfully synthesize exactly one single molecule of a stable hexose sugar, the complex cycle must precisely incorporate exactly six individual molecules of atmospheric carbon dioxide. These actively combine with exactly six molecules of the starting Ribulose 1,5-bisphosphate substrate, thereby yielding exactly twelve molecules of 3-Phosphoglycerate. This massive accumulation of starting material prepares the system for the severe energetic investment required.
The incredible energy drain of Photosynthesis becomes completely obvious during the massive reduction phase. To successfully convert those twelve specific molecules into twelve energetic molecules of Glyceraldehyde 3-phosphate, the cycle must forcefully and aggressively consume exactly twelve molecules of ATP and exactly twelve molecules of NADPH. This represents a truly massive drain on the energetic currencies violently generated earlier by the thylakoid light reactions.
Of the twelve newly formed complex G3P molecules, only exactly two are ever allowed to safely exit the ongoing cycle. These two highly precious molecules proceed via distinct gluconeogenesis-like pathways to ultimately form a single molecule of a stable hexose. This single sugar molecule represents the final ultimate physical prize of the entire incredibly exhaustive, energy-intensive biochemical process.
The remaining ten distinct molecules of G3P must rapidly undergo the highly complex regeneration phase. They are meticulously structurally rearranged and reconstructed specifically back into exactly six molecules of the starting Ribulose 1,5-bisphosphate. This tedious regeneration forcefully consumes an additional six molecules of ATP, pushing the total massive thermodynamic cost of Photosynthesis to a staggering eighteen ATP and twelve NADPH per sugar.
Slide 15: The Coupled Economy of Photosynthesis
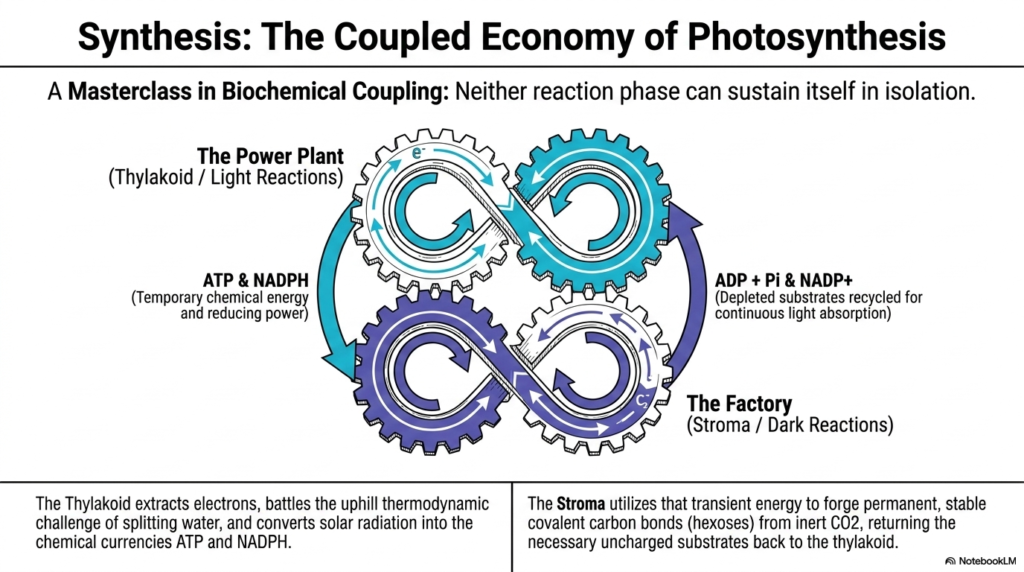
The final slide brilliantly synthesizes the entire presentation, illustrating the beautifully coupled economy of Photosynthesis. It perfectly frames the entire biochemical cascade as an absolute masterclass in strict biological biochemical coupling. Students must clearly understand that neither the light reaction phase nor the dark reaction phase can ever successfully sustain itself in total isolation.
The thylakoid membrane acts exactly as the ultimate cellular Power Plant during Photosynthesis. It aggressively extracts stubborn electrons, successfully battles the severe uphill thermodynamic challenge of violently splitting water, and brilliantly converts ambient solar radiation into highly usable chemical currencies. The exclusive biological output of this massive power plant is the steady, vital supply of the temporary chemical energy found in ATP and NADPH.
However, this massive generated energy is entirely temporary and highly volatile. The surrounding fluid stroma acts perfectly as the heavily dependent cellular Factory. It eagerly uses that massive, transient energy to forge permanent, highly stable covalent carbon bonds from the inert carbon dioxide gas. The factory successfully locks the fleeting quantum energy safely away into highly reliable, storable hexose carbohydrates.
Crucially, the relationship is entirely perfectly reciprocal. As the massive stroma factory forcefully consumes the supplied ATP and NADPH, it actively generates entirely depleted substrates in the form of ADP, inorganic phosphate, and empty NADP+. These severely depleted molecules are carefully recycled directly back to the active thylakoid power plant, ensuring the necessary uncharged substrates are permanently available for continuous, uninterrupted solar light absorption.
This beautifully intertwined, endlessly cycling dynamic perfectly encapsulates the absolute true majesty of Photosynthesis. The aggressive thermodynamic battle, successfully won by the thylakoids, directly enables the phenomenal synthetic construction performed in the stroma. Together, these two perfectly coupled halves execute the single most important, life-sustaining biochemical energy-transduction mechanism known in the natural biological world.
Please read our Content Disclaimer Statement.
Check out our social media channels: