60. Heme Degradation: The Complete Guide to Bilirubin Metabolism
Have you ever wondered what happens to red blood cells when they reach the end of their lifespan? The human body runs a remarkably efficient recycling program to manage this cellular waste. This introductory slide outlines the deck’s core purpose: mapping the molecular journey from a functional oxygen-carrying molecule to its final breakdown products. It sets the stage for understanding the intricate biochemical pathway and the profound clinical significance of this highly regulated physiological process.
Slide 1: Introduction to Heme Degradation: The Body’s Ultimate Recycling Program
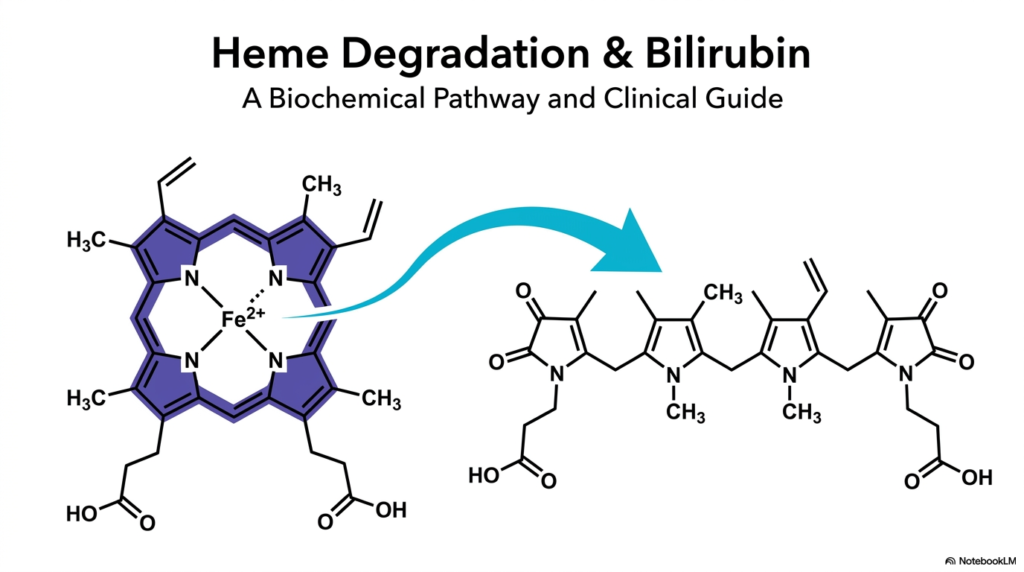
Heme Degradation represents a fundamental biochemical process that prevents the potentially lethal buildup of cellular components. Every single day, the human organism gracefully breaks down hundreds of millions of aging red blood cells. The hemoglobin housed within these cells must be carefully dismantled. This controlled breakdown actively prevents rampant oxidative stress and subsequent tissue damage. The featured visual elegantly captures the initial dramatic molecular transformation. It shows the shift from a robust, cyclic porphyrin ring into a linear chain, preparing the molecule for safe disposal.
In the fascinating context of Heme Degradation, the spotlight falls squarely on these precise structural changes. The starting molecule possesses a rigid architecture specifically designed to cradle iron. Once the cell expires, this once-helpful structure instantly becomes a dangerous liability. Specialized enzymes must crack the protective ring wide open. This precise molecular surgery guarantees that the body can salvage its precious iron reserves. Simultaneously, it readies the remaining organic skeleton for a complex journey of transport, processing, and eventual elimination.
Mastering the intricate stages of Heme Degradation is essential for any medical professional. If a single metabolic step fails, the sudden buildup of toxic byproducts routinely leads to severe clinical presentations, such as severe jaundice. This comprehensive presentation systematically demystifies these complex molecular shifts step by step. By thoroughly internalizing this essential pathway, students construct a formidable foundation. This knowledge is vital for accurately diagnosing and effectively treating various metabolic and hepatic disorders encountered in clinical practice.
Slide 2: The Macro Overview of Heme Degradation: Scale and Scope
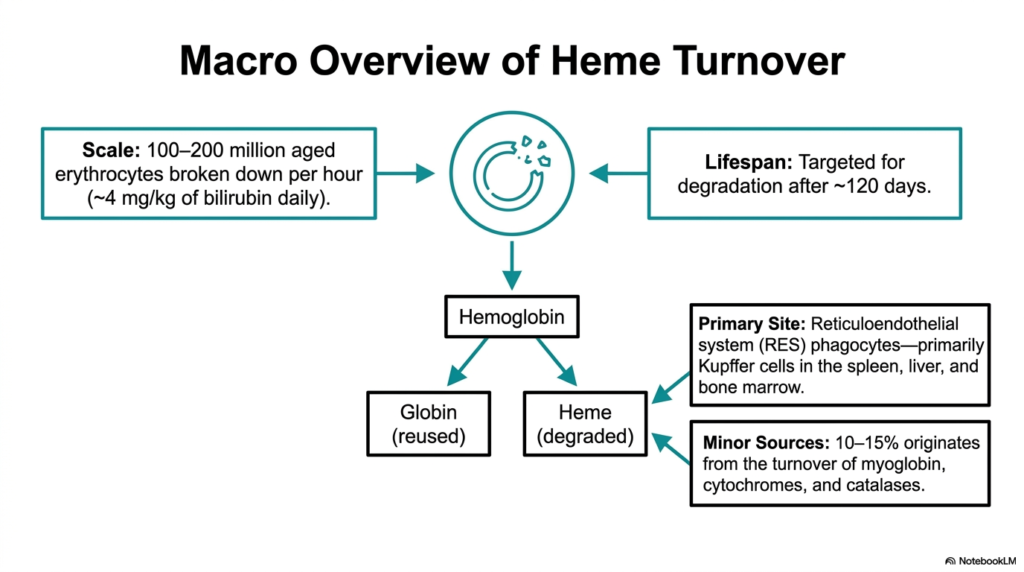
Imagine a busy factory that must safely dispose of millions of specialized tools every single hour without a single shutdown. The human body accomplishes exactly this staggering feat during regular cellular turnover. This slide provides a macro-level perspective on the sheer scale of the system. It outlines the massive daily volume of erythrocyte breakdown and pinpoints the exact anatomical locations where this crucial cellular demolition occurs, emphasizing the continuous, high-volume nature of the system.
The sheer volume of Heme Degradation occurring within a healthy body is truly spectacular to consider. Between 100 and 200 million aged erythrocytes are meticulously disassembled every hour. This translates to roughly 4 milligrams of bilirubin produced per kilogram of body weight per day. The primary site for this monumental task is the reticuloendothelial system. Here, specialized phagocytes, particularly Kupffer cells in the spleen, liver, and bone marrow, initiate the complex disassembly process.
The systematic process of Heme Degradation initially splits the abundant hemoglobin molecule into two distinct parts. The globin protein portion is efficiently recycled into individual amino acids for future protein synthesis. Conversely, the heme ring itself must be systematically degraded and removed. While aged red blood cells provide the vast majority of the substrate, roughly ten to fifteen percent originates from minor sources. These additional inputs include the regular turnover of myoglobin, various cytochromes, and cellular catalases.
Understanding the massive scope of Heme Degradation provides crucial context for appreciating the workload placed on the liver and spleen. The reticuloendothelial system acts as an incredibly efficient biological filter. It continuously identifies and engulfs red blood cells that have reached their hundred-and-twenty-day lifespan. This precise identification mechanism ensures that only exhausted cells are targeted. By visualizing the scale of this daily turnover, students can better comprehend how easily hepatic or hemolytic diseases can overwhelm the body.
Slide 3: The Starting Molecule of Heme Degradation: Analyzing the Porphyrin Ring
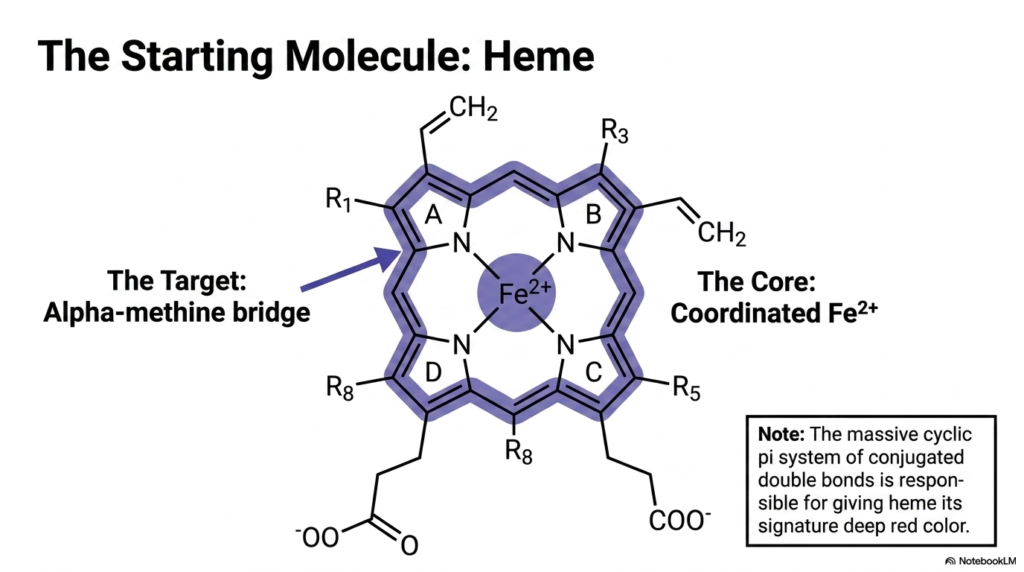
To dismantle a complex structure safely, one must first deeply understand its architectural blueprints. Before exploring the breakdown process, it is vital to examine the specific molecular target itself. This slide introduces the intricate structure of the starting molecule and highlights its unique chemical properties. By focusing on the conjugated pi system and the coordinated central metal ion, students can clearly visualize exactly why specialized enzymatic machinery is absolutely required to process this stable, resilient compound safely.
The initial substrate for Heme Degradation is a highly stable, tightly coordinated tetrapyrrole ring system. This impressive cyclic structure features a massive conjugated double-bond system. These alternating double bonds create a continuous pi electron cloud that effectively absorbs specific wavelengths of light. This extensive conjugation is precisely what gives the molecule its signature deep red pigmentation. At the absolute center of this flat, disc-like molecular stage sits a single, securely coordinated ferrous iron atom.
The opening act of Heme Degradation targets a very specific structural vulnerability within this otherwise robust chemical fortress. The alpha-methine bridge acts as the designated weak point connecting the pyrrole rings. Because the molecule is so chemically stable and highly lipophilic, it cannot simply be dissolved or casually flushed from the system. The body must deploy targeted oxidative force directly at this methine bridge to crack the ring open and release the deeply sequestered iron payload safely.
Without the specific steps involved in heme degradation, the highly reactive central iron atom would cause profound cellular damage. The surrounding porphyrin structure protects the cell from this iron, but it simultaneously makes disposal extremely challenging. Students must recognize that the rigid, planar geometry of the molecule is its defining characteristic. Breaking this geometry is the fundamental prerequisite for transforming the dangerous, lipophilic waste product into a safer, more manageable, and ultimately excretable metabolic compound.
Slide 4: Step One of Heme Degradation: Ring Cleavage by Heme Oxygenase
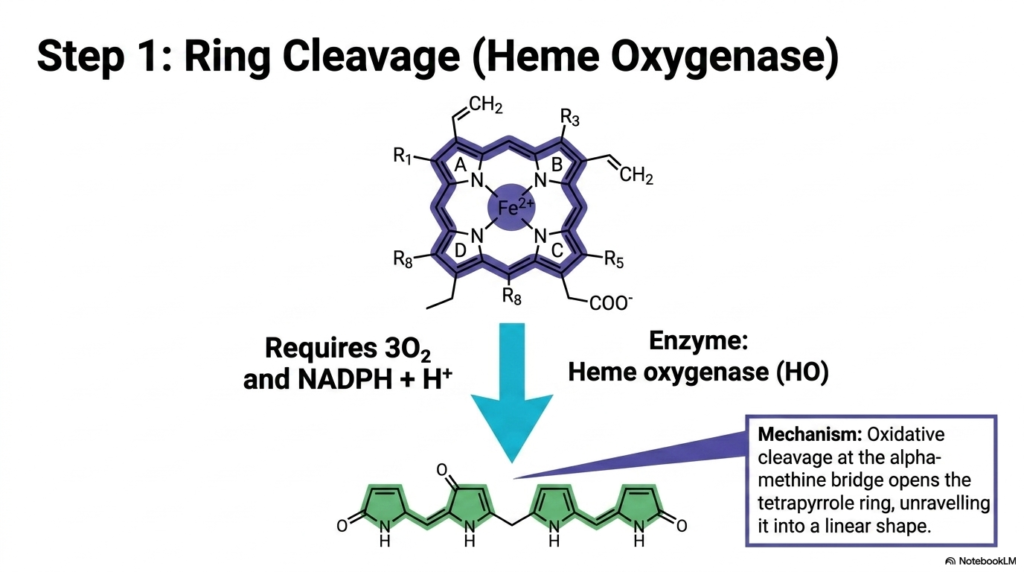
How does a cell safely crack open a chemical vault without spilling its hazardous contents? The answer lies in precision enzymatic machinery. This crucial slide details the very first, irreversible chemical reaction in the breakdown pathway. It focuses squarely on the specific enzyme responsible for prying open the cyclic ring. The objective is to clearly illustrate the biochemical requirements and the precise structural transformation that permanently neutralizes the original porphyrin’s oxygen-carrying capacity.
The initiation of Heme Degradation is a masterpiece of precision biochemical engineering. The pivotal enzyme executing this delicate maneuver is heme oxygenase. This highly specialized protein orchestrates the oxidative cleavage of the tetrapyrrole ring precisely at the alpha-methine bridge. The process is remarkably energy-intensive, strictly requiring three molecules of molecular oxygen, the reducing power of NADPH, and a hydrogen ion. This targeted attack systematically unravels the complex cyclic architecture into a much simpler linear shape.
During this specific phase of Heme Degradation, the dramatic physical restructuring of the molecule yields biliverdin. The enzyme forcefully inserts oxygen atoms into the molecular framework, physically snapping the rigid bonds. The previously flat, circular chemical vault is thus forced open, unfurling into a long, flexible linear chain. This newly formed biliverdin molecule exhibits a distinct green coloration, a direct physical manifestation of the severely altered yet still partially intact system of conjugated double bonds.
This exact stage of heme degradation is the critical point of no return for the molecule. Once the ring is cleaved, the structure can never again carry oxygen. Heme oxygenase operates as a vital guardian, ensuring the safe dismantling of a potentially dangerous compound. For medical students, understanding the rigorous chemical demands of this cleavage step highlights the immense metabolic energy the cell readily sacrifices to safely manage its own highly toxic internal metabolic waste products.
Slide 5: Physiological Byproducts of Heme Degradation: Carbon Monoxide and Iron

When you demolish a building, you inevitably generate debris. But what if that debris contained highly valuable building materials alongside potent toxic gases? This slide shifts the focus from the primary structural remnants to the vital byproducts released during the initial ring cleavage. The core purpose here is to reveal how the body ingeniously repurposes a dangerous heavy metal while simultaneously utilizing a notoriously toxic gas as a powerful internal cellular signaling mechanism.
A fascinating paradox in heme degradation is the endogenous production of carbon monoxide. Most people solely associate this gas with lethal poisoning from faulty furnaces. Yet, roughly 86% of the carbon monoxide naturally present in the human body originates directly from this specific enzymatic cleavage. Far from being mere metabolic waste, this gaseous byproduct acts as a highly potent gasotransmitter. It actively functions as an important endogenous vasodilator and a potent anti-inflammatory signaling agent.
The safe handling of iron is another critical triumph of Heme Degradation. The oxidative cracking of the ring liberates a free ferrous iron atom. Free iron is an incredibly dangerous cellular asset; if left unbound, it rapidly generates lethal free radicals through runaway Fenton reactions. To prevent catastrophic oxidative damage, the newly liberated iron is immediately sequestered by the storage protein ferritin. This rapid containment strategy neutralizes the threat while keeping the element available for reuse.
The flawless execution of heme degradation guarantees that this salvaged iron is fully recycled by the body for de novo synthesis. Rather than constantly absorbing massive amounts of dietary iron, the organism efficiently relies on this internal supply chain. Recognizing the dual nature of these byproducts demonstrates the elegance of human biochemistry. The system masterfully converts potentially fatal metabolic waste products into indispensable physiological tools, ensuring optimal cellular communication and maintaining a stable, safe heavy-metal equilibrium.
Slide 6: The Second Step of Heme Degradation: Reduction to Bilirubin
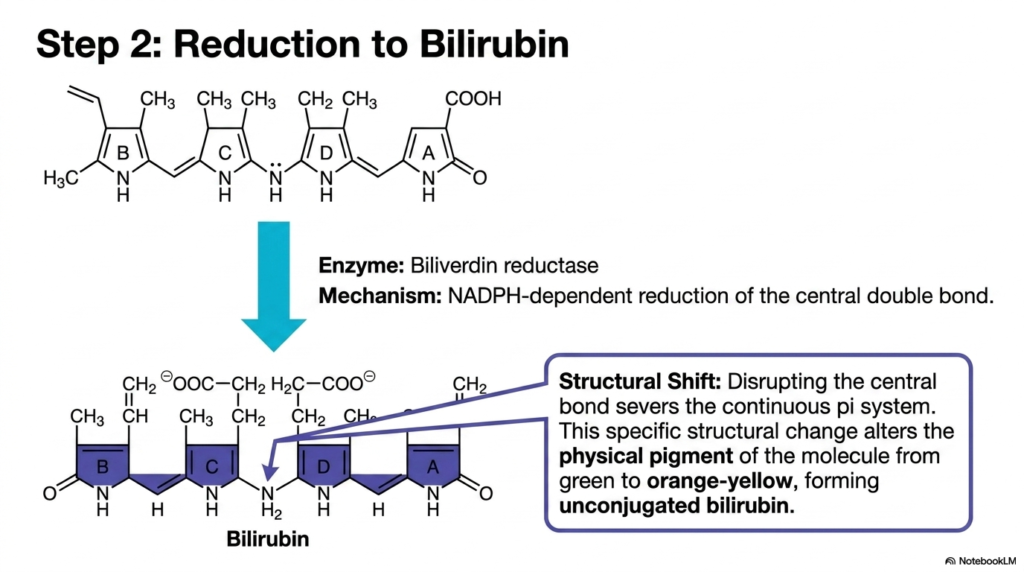
How does a cell transform a stiff, green, linear molecule into a flexible, yellow compound ready for transport? The solution requires a targeted chemical reduction. This slide highlights the second major enzymatic reaction in the pathway. The goal is to explain how a specific enzyme carefully snips the remaining electrical connections within the molecule, fundamentally altering its physical structure and drastically shifting its visible pigment, setting the stage for its complex journey through the bloodstream.
The second major enzymatic hurdle in Heme Degradation is catalyzed by biliverdin reductase. This powerful enzyme acts on the green biliverdin chain produced during the initial cleavage. The mechanism involves a strict NADPH-dependent reduction focused entirely on the molecule’s central double bond. By forcibly pushing hydrogen atoms onto this specific central junction, the enzyme systematically severs the continuous pi-electron system that previously extended the entire length of the extended tetrapyrrole chain.
This targeted reduction step during Heme Degradation initiates a profound structural shift. The disruption of the central double bond effectively divides the large pi system into two smaller, isolated halves. This specific structural alteration profoundly changes how the molecule interacts with visible light. Consequently, the compound’s physical pigment transitions dramatically from its initial vibrant green to a distinct orange-yellow hue. The resulting molecule from this intricate chemical modification is recognized as unconjugated bilirubin.
The formation of unconjugated bilirubin represents a critical juncture in heme degradation. The newly formed molecule is highly lipophilic, causing it to fold tightly upon itself through internal hydrogen bonding. This unique folded geometry makes it incredibly difficult to manage in an aqueous environment. For clinical students, grasping this specific reduction step clarifies the exact chemical origin of the yellow pigmentation universally associated with conditions like jaundice, directly linking molecular structure to observable, real-world diagnostic signs.
Slide 7: The Visual Proof of Heme Degradation: The Anatomy of a Bruise

Have you ever watched a nasty bruise slowly change color over a couple of weeks? You were actually watching a live, macroscopic display of complex enzymatic chemistry. This slide translates abstract metabolic pathways into a highly relatable, everyday clinical observation. The purpose is to map the previously discussed molecular shifts directly onto the predictable sequence of colors visible in a common peripheral tissue hematoma, perfectly bridging the gap between microscopic biochemistry and clinical physical examination.
The shifting colors of a bruise provide a beautiful, real-time window into Heme Degradation. Phase one is characterized by a deep red or purple coloration. This hue perfectly represents the accumulation of intact blood cells trapped beneath the skin, which contain the original, fully functional cyclic pi system of the porphyrin ring. At this stage, the cellular macrophages have just arrived on the scene and are only beginning the monumental task of engulfing the trapped erythrocytes.
As Heme Degradation actively progresses in the tissue, phase two emerges, bringing a distinct green tint to the bruised area. This green coloration directly corresponds to the enzymatic activity of heme oxygenase. The macrophages have successfully cleaved the red rings, unraveling them into linear green biliverdin molecules with intact linear conjugation. The visual shift is a direct, tangible readout of the specific oxidative ring cleavage happening simultaneously across millions of trapped hemoglobin molecules.
The final visible stage of heme degradation manifests as a fading yellow or orange patch on the skin. This marks phase three, indicating the successful deployment of biliverdin reductase. The central double bonds have been reduced, dividing the pi system and forming unconjugated bilirubin. By understanding this chromogenic shift, medical students learn to visually estimate the age of a hematoma simply by observing its color, utilizing biochemistry directly as a rapid, practical diagnostic tool.
Slide 8: Blood Transport in Heme Degradation: The Essential Albumin Shuttle
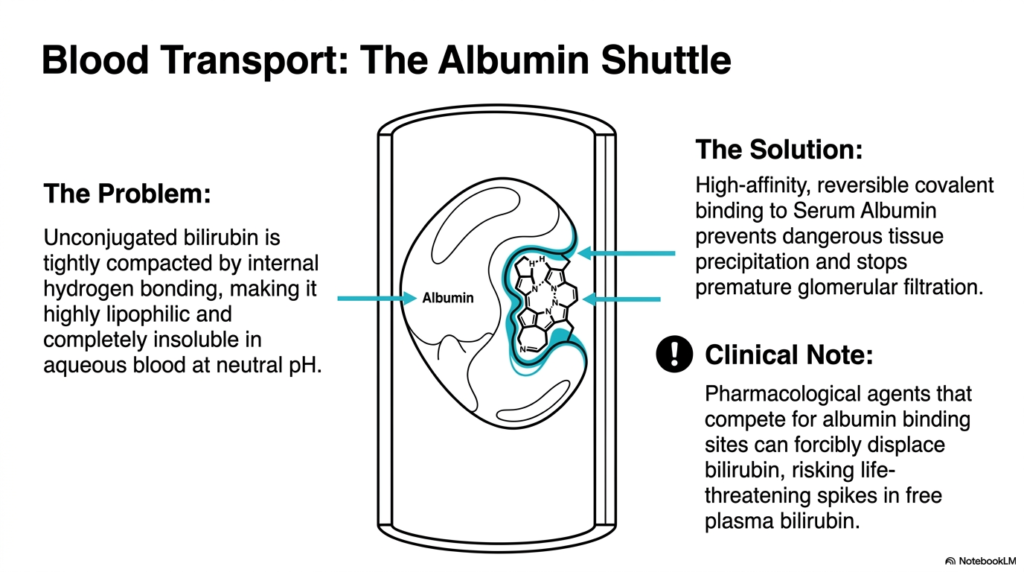
If a metabolic waste product acts like oil, how does the body transport it through the watery rivers of the bloodstream? It requires a specialized, high-capacity cellular cargo ship. This important slide addresses the glaring solubility problem created by the previous enzymatic steps. It clearly illustrates the critical transport mechanism that completely prevents the lipophilic waste from prematurely crystallizing in peripheral tissues or causing severe functional damage to the kidneys, highlighting the delicate balance of plasma protein binding.
A significant logistical challenge within Heme Degradation is the chemical nature of unconjugated bilirubin. Because the molecule is tightly compacted by extensive internal hydrogen bonding, it is highly lipophilic. This means it behaves much like grease and is completely insoluble in the aqueous environment of blood at a neutral physiological pH. Without a dedicated transport system, this oily substance would immediately precipitate out of the blood, rapidly causing catastrophic blockages and widespread, devastating tissue toxicity.
To solve this critical transport issue during heme degradation, the body utilizes serum albumin as a high-capacity molecular shuttle. The unconjugated molecule binds to specific high-affinity sites on the surface of albumin. This reversible but remarkably tight binding safely sequesters the lipophilic waste product. It allows the compound to travel smoothly through the circulatory system, preventing dangerous tissue deposition while simultaneously preventing premature, highly damaging filtration by the delicate renal glomeruli.
Understanding this transport mechanism is vital for mastering the pharmacology of heme degradation. A crucial clinical note involves the introduction of external pharmacological agents. Many common drugs strongly compete for these exact same binding sites on the albumin molecule. If a patient takes these competing medications, the drugs can forcibly displace the bound waste product. This displacement suddenly triggers a massive, life-threatening spike in free plasma bilirubin, quickly precipitating severe neurological crises, especially in vulnerable pediatric patients.
Slide 9: Hepatic Conjugation in Heme Degradation: Unlocking Water Solubility
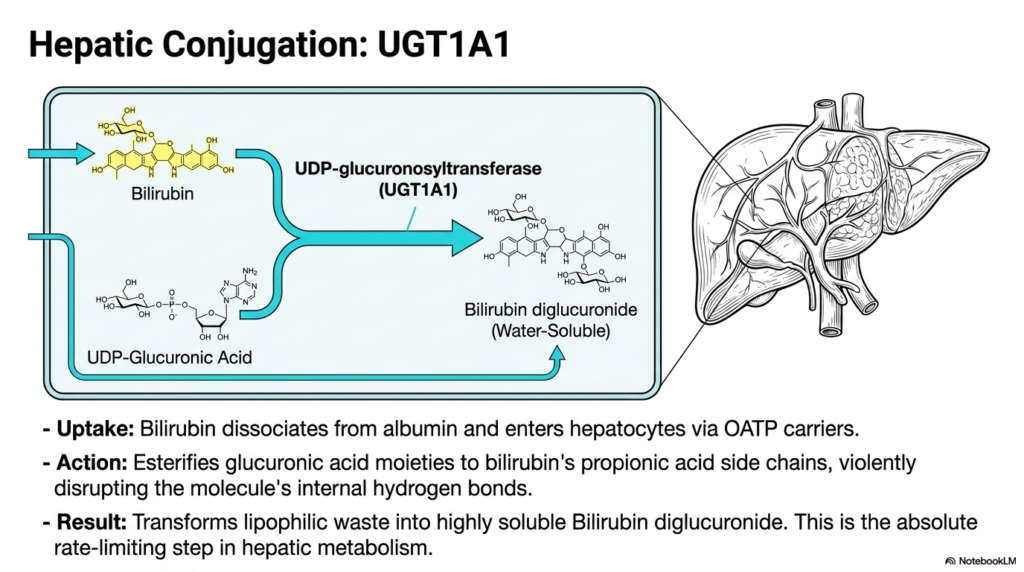
How do the liver’s metabolic factories finally make an oily waste product dissolve in water? The solution is to attach sugar molecules to it forcefully. This slide dives into the endoplasmic reticulum of the hepatocyte to showcase the absolute rate-limiting step of the entire metabolic pathway. The purpose is to vividly explain the esterification process that ultimately breaks the molecule’s internal bonds, preparing it for imminent excretion into the human digestive tract.
The most critical bottleneck in Heme Degradation occurs deep within the liver’s functional tissue. Upon reaching the liver sinusoids, the previously bound cargo cleanly dissociates from its albumin shuttle and rapidly enters hepatocytes via specific organic anion transporters. Once safely inside, the powerful enzyme UDP-glucuronosyltransferase, specifically the vital UGT1A1 isoform, takes absolute control. This enzyme acts as a biological stapler, aggressively attaching highly polar sugar components directly onto the stubborn, lipophilic framework of the metabolic waste product.
This crucial phase of Heme Degradation relies on a steady supply of UDP-glucuronic acid. The enzyme meticulously esterifies these glucuronic acid moieties directly onto the molecule’s propionic acid side chains. This forceful addition of bulky, heavily charged sugar groups acts like a molecular wedge. It violently disrupts the tight internal hydrogen bonds that previously kept the molecule stubbornly folded. The dense, folded structure is finally forced entirely open, exposing its polar edges.
The ultimate result of this central step in heme degradation is the creation of bilirubin diglucuronide. This new molecule is completely transformed; it transitions from an oily, lipophilic hazard into a highly water-soluble compound. Because this conjugation is the absolute slowest step in the entire hepatic metabolism sequence, any slight liver impairment immediately causes a massive systemic backup. Grasping this specific enzyme’s function allows students to perfectly understand the mechanism behind severe hepatocellular jaundice.
Slide 10: Intestinal Metabolism and Excretion in Heme Degradation
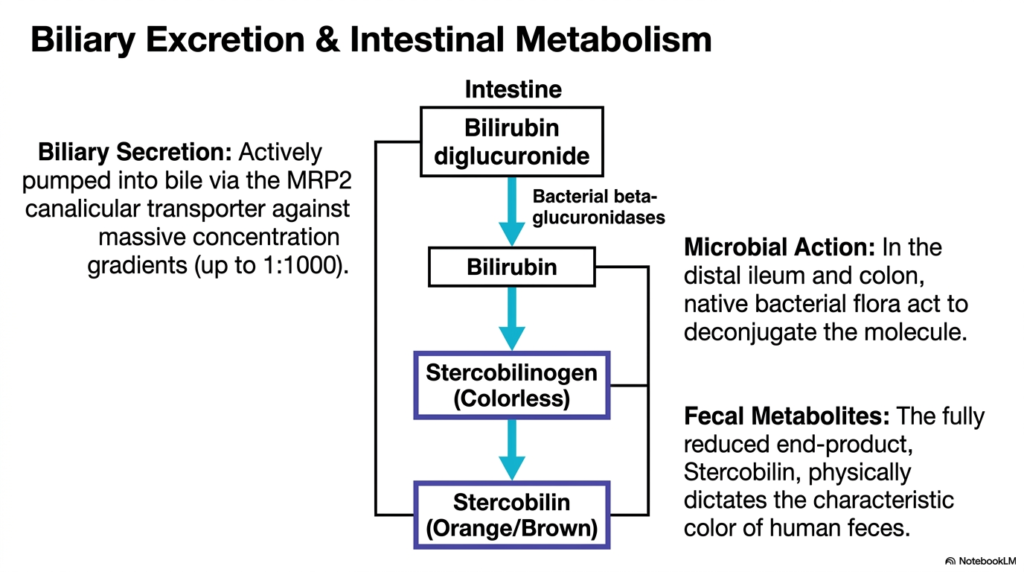
Where does biological waste ultimately exit the body, and why is human waste the color it is? The journey ends in the dark reaches of the human gut. This slide follows the newly water-soluble compound as it leaves the liver and encounters the vast microbiome of the digestive tract. It aims to explain the energetic pumping mechanism required for excretion and the fascinating microbial chemistry that dictates the final characteristic color of fecal matter.
The final disposal phase of Heme Degradation demands a tremendous amount of cellular energy. The freshly conjugated, water-soluble molecule cannot simply diffuse out of the liver. Instead, it must be actively pumped directly into the bile canaliculi. This arduous task is performed by the MRP2 canalicular transporter, which forcefully pushes the compound against a steep concentration gradient, sometimes reaching ratios of 1:1,000. This creates the primary component of dark green human bile.
Once deposited into the intestines, the products of heme degradation face a brand new chemical environment. In the distal ileum and the colon, the massive population of native bacterial flora takes over the metabolic process. These microbes deploy their own powerful beta-glucuronidase enzymes. These bacterial tools swiftly act to deconjugate the molecule, stripping away the previously attached sugar groups. This microbial action reduces the complex compound down into a completely colorless intermediate known as stercobilinogen.
The ultimate conclusion of heme degradation is a visually obvious, everyday phenomenon. The colorless stercobilinogen travels further down the digestive tract, where it undergoes spontaneous oxidation. This final, simple chemical reaction produces a fully reduced end-product called stercobilin. Stercobilin is a distinctly orange or brown compound. Students find it incredibly fascinating that this specific end-product of microscopic blood breakdown physically dictates the characteristic, universal brown color of healthy human feces at the macroscopic level.
Slide 11: Renal Excretion and the Loop of Heme Degradation
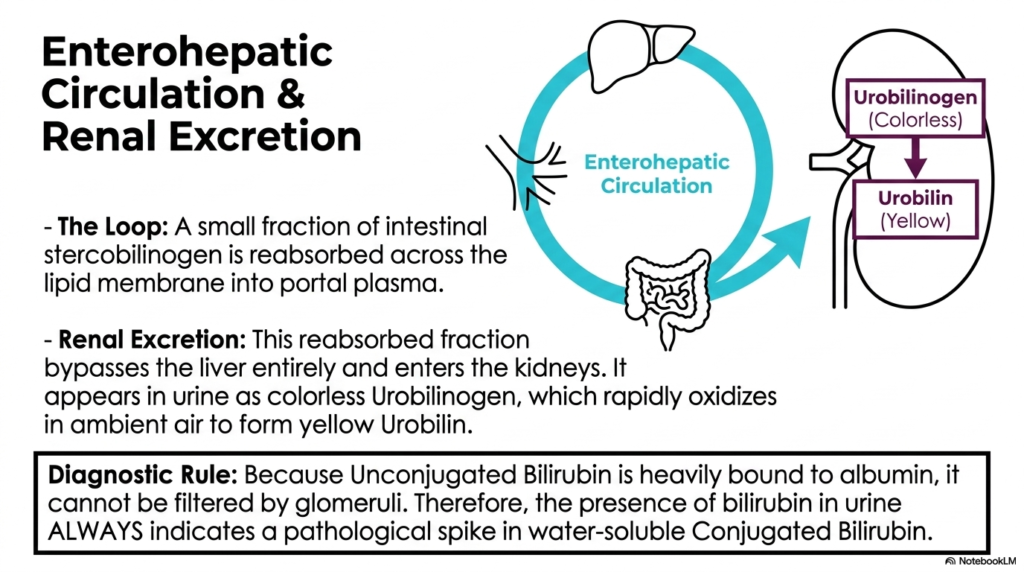
Why does urine sometimes turn a bright, alarming yellow when the liver is struggling? The answer lies in a biological feedback loop and a secondary escape hatch. This slide explores the clever anatomical shortcut that allows a small fraction of waste to bypass the digestive tract entirely. By highlighting the distinct differences in kidney filtration, the presentation equips students with a critical, undeniable diagnostic rule for instantly recognizing severe, dangerous liver pathology.
A fascinating twist in the story of heme degradation is the enterohepatic circulation loop. While the vast majority of the colorless intestinal stercobilinogen continues downward towards fecal excretion, a small fraction takes a detour. This specific fraction is reabsorbed directly across the intestinal lipid membranes and returns to the portal plasma. Instead of returning to the liver for another round, this reabsorbed fraction completely bypasses the hepatic system and travels straight to the kidneys.
This specific renal pathway within Heme Degradation is responsible for the normal color of urine. Upon reaching the kidneys, the compound is filtered and appears in the liquid urine as the colorless molecule urobilinogen. However, the moment this clear liquid is exposed to ambient air, it rapidly oxidizes. This instantaneous oxidation reaction transforms the previously invisible compound into urobilin, the exact chemical responsible for giving normal urine its familiar yellow hue.
The diagnostic implications of this phase of Heme Degradation are absolutely profound for future clinicians. Unconjugated bilirubin is tightly bound to albumin, so it is not normally filtered by healthy glomeruli. Therefore, a steadfast diagnostic rule holds: the sudden appearance of bilirubin in the urine always indicates a pathological spike in water-soluble conjugated bilirubin. This single observation immediately tells the physician that the liver is conjugating the waste but failing to excrete it.
Slide 12: Isoforms of Heme Degradation: The Heme Oxygenase Matrix
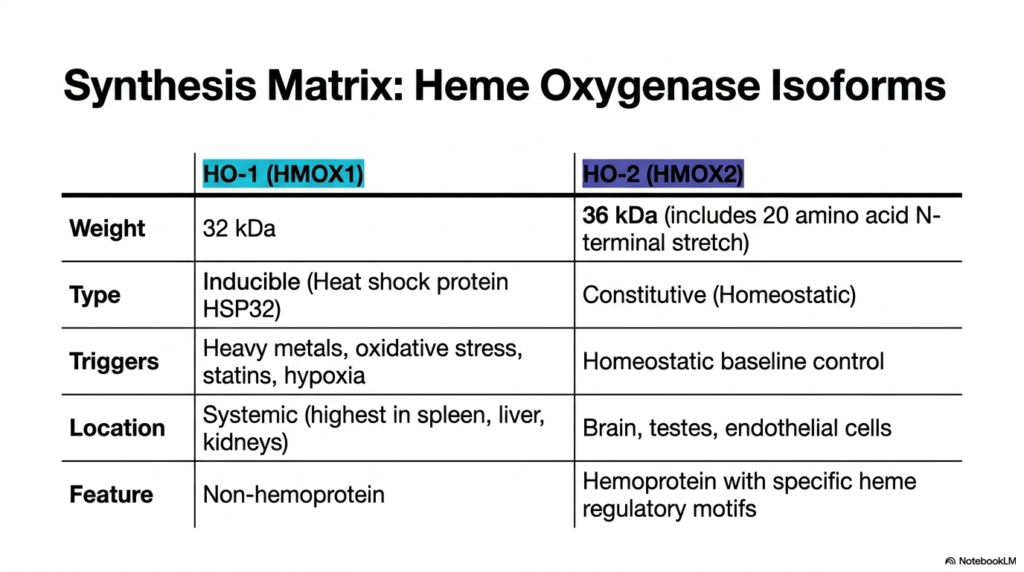
Are all breakdown enzymes built the same way, or does the body customize them for specific emergencies? It turns out the body maintains both a steady baseline and a rapid response team. This slide breaks down the critical differences between the two primary isoforms of the crucial ring-cleaving enzyme. The goal is to provide a comprehensive structural and functional matrix that demonstrates how the body dynamically upregulates this metabolic machinery to survive sudden periods of intense, overwhelming oxidative stress.
The enzymatic machinery driving Heme Degradation is highly adaptable, featuring two distinct primary isoforms. The first is HO-1, also known as HMOX1. This specific isoform is a non-hemoprotein weighing approximately thirty-two kilodaltons. Its most vital characteristic is its high inducibility. Acting essentially as a specialized heat shock protein, it serves as the body’s rapid-response metabolic unit. It is systemically distributed but is found at the highest concentrations in the spleen, liver, and kidneys.
The sudden activation of this specific heme degradation pathway is a brilliant survival mechanism. HO-1 is aggressively triggered by extreme physiological threats, including heavy metal toxicity, overwhelming oxidative stress, statin administration, and severe cellular hypoxia. When the body senses dangerous tissue damage, it instantly floods the system with this enzyme. This rapid upregulation ensures that toxic cellular debris is cleared at an accelerated rate, protecting fragile peripheral tissues from catastrophic, irreversible runaway oxidative damage cascades.
In stark contrast, the second pathway of heme degradation relies on the HO-2 isoform. Weighing roughly thirty-six kilodaltons due to an additional amino acid stretch, this hemoprotein is strictly constitutive, meaning it operates constantly to maintain normal homeostatic baseline control. It is uniquely concentrated in the brain, testes, and endothelial cells. By mastering the distinct triggers and anatomical locations of these two isoforms, students gain a deep, nuanced understanding of how humans dynamically manage complex metabolic stress.
Slide 13: Clinical Pathologies of Heme Degradation: Understanding Jaundice Matrices

How do you pinpoint the exact location of a metabolic traffic jam simply by looking at bloodwork? You must understand the specific choke points of the system. This clinically focused slide categorizes the major failures of the breakdown pathway into three distinct anatomical pathologies. It is designed to train students to instantly correlate specific biochemical spikes in the blood with pre-hepatic, hepatic, or post-hepatic failures, effectively turning complex lab results into a clear, actionable diagnostic map.
When the elegant process of heme degradation fails, the clinical result is hyperbilirubinemia, universally recognized as jaundice. This matrix cleanly divides these failures. Pre-hepatic, or hemolytic, jaundice occurs when increased erythrocyte destruction overwhelms the liver’s natural conjugating capacity. Because the liver simply cannot keep pace with the massive influx of raw material, the predictable biochemical result is a pronounced systemic spike in unconjugated (indirect) bilirubin. Notably, the patient’s urine remains acholuric, entirely free of bilirubin.
The second major failure point in heme degradation occurs directly within the liver tissue itself. Hepatic, or hepatocellular, jaundice is usually caused by impaired enzymatic conjugation, frequently due to severe viral hepatitis or chronic liver cirrhosis. In this scenario, the cellular machinery is broken, leading to another massive spike in unconjugated molecules. The third category, post-hepatic or obstructive jaundice, happens when there is a physical blockage of bile drainage, such as a large gallstone or pancreatic tumor.
This final obstructive failure in Heme Degradation results in a markedly different laboratory presentation. Because the liver successfully conjugates the waste but cannot excrete it, conjugated bilirubin dangerously spikes in the blood, leading to pathologically dark urine. Finally, a dire pediatric warning must be noted. In severe neonatal jaundice, the highly lipophilic unconjugated waste can easily cross an infant’s immature blood-brain barrier. This precipitates irreversible, devastating neurological damage known as kernicterus, making rapid medical intervention absolutely critical.
Slide 14: The Complete Heme Degradation Blueprint: A Multi-Compartment Pipeline
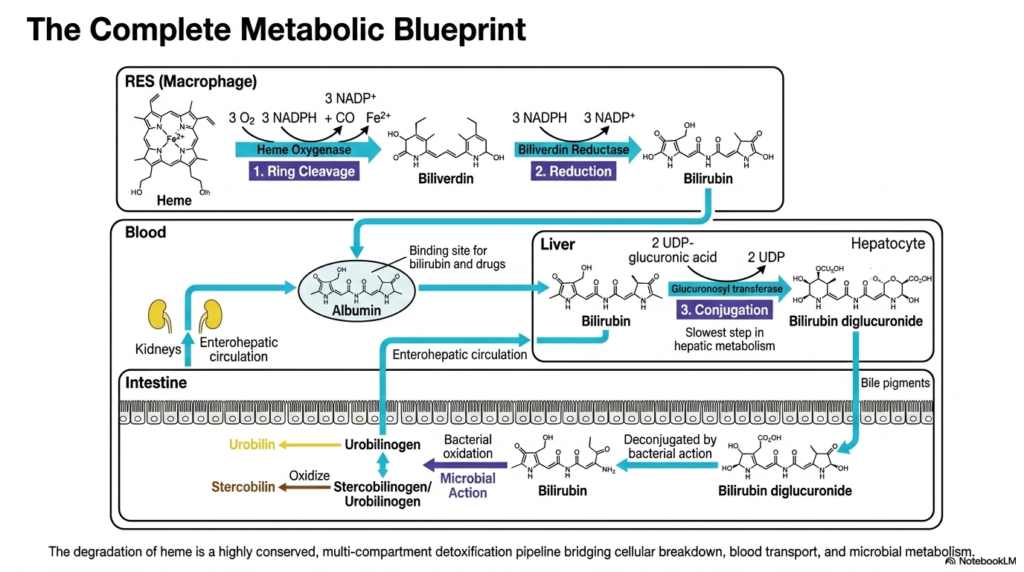
Can we step back and view the entire metabolic assembly line from start to finish? To truly master this topic, one must see the interconnected whole. This final summary slide seamlessly stitches together all isolated biochemical reactions into a unified, multi-compartment detoxification pipeline. The ultimate purpose is to consolidate the student’s newfound knowledge, bridging cellular breakdown, vascular transport, and microbial metabolism into a single, cohesive, and easily digestible physiological master blueprint.
The overarching mastery of Heme Degradation requires visualizing a perfectly synchronized cellular relay race. This comprehensive blueprint begins securely inside the resident macrophages of the reticuloendothelial system. Here, the initial toxic ring is safely pried open by specialized oxygenase enzymes, heavily utilizing molecular oxygen and reducing agents to forge the green intermediate biliverdin. Immediately following this, reductase enzymes rapidly step in, carefully snipping the central bonds to produce the characteristic yellow, highly lipophilic compound known as unconjugated bilirubin.
The complex vascular phase of heme degradation represents the crucial second act of this metabolic pipeline. Because the newly formed waste is entirely hydrophobic, it absolutely requires the dedicated albumin transport shuttle to navigate the watery bloodstream safely. This shuttle flawlessly delivers the dangerous cargo directly to the waiting hepatocytes. Deep inside the liver’s factories, the slow, rate-limiting conjugation step occurs, forcefully attaching bulky sugar molecules to completely transform the dangerous oil into a highly water-soluble, easily excretable substance.
The final, spectacular act of heme degradation beautifully bridges human metabolism with microscopic bacterial ecosystems. Pumped aggressively into the digestive tract, the water-soluble conjugate is eventually stripped of its sugars by thriving microbial colonies. The resulting colorless compounds are ultimately oxidized in ambient air to produce the distinct yellow and brown pigments found in our daily biological waste. This incredible, highly conserved detoxification pipeline brilliantly ensures that potentially lethal cellular garbage is safely and efficiently eliminated from the body forever.
Please read our Content Disclaimer Statement.
Check out our social media channels: