32. Intermediary Metabolism Explained: Pathways and Energy
Have you ever wondered how a late-night pizza slice transforms into the energy required to ace a complex medical exam? The answer lies in the microscopic, bustling factories within our cells. This comprehensive slide deck is designed to demystify these intricate biochemical networks for college and medical students. By exploring the core mechanisms of nutrient processing and energy extraction, this guide provides a clear, step-by-step breakdown of how the body sustains human life at the molecular level.
Slide 1: Unveiling Intermediary Metabolism
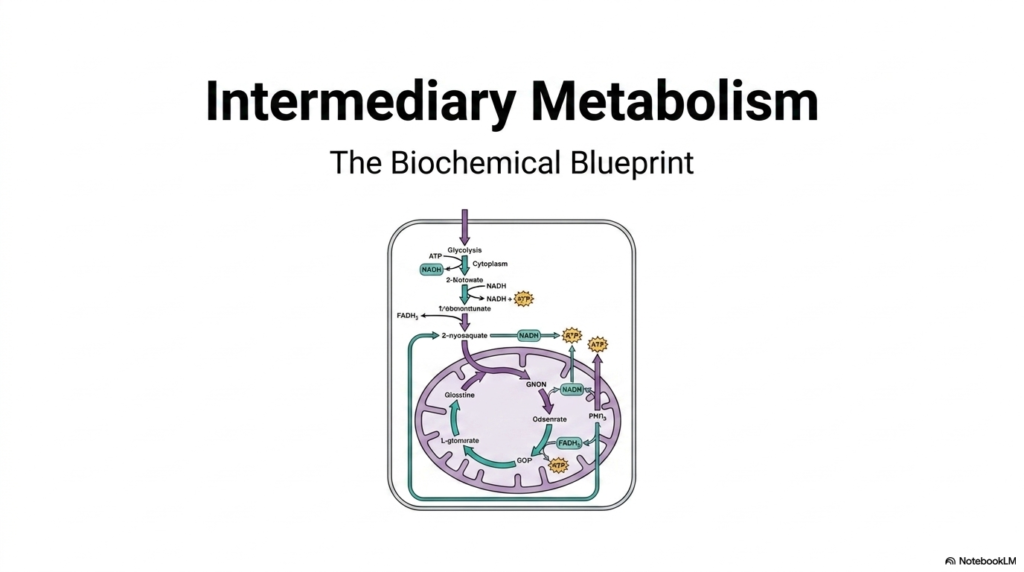
The journey into the cellular universe begins with a macroscopic view of intermediary metabolism. This overarching concept represents the complete biochemical blueprint of how biological systems process fuels. It is the intricate web of chemical reactions that sustains life, carefully balancing the breakdown of nutrients with the creation of new cellular materials. Understanding intermediary metabolism is crucial for any medical or biological sciences student, as it provides the foundation for understanding human physiology, pathology, and pharmacology.
The featured diagram provides a spectacular visual map of these essential pathways. At the very top, one can observe glycolysis taking place within the cytoplasm of the cell. This initial step of intermediary metabolism takes glucose and systematically splits it, extracting a small but vital amount of immediate energy in the form of ATP and NADH. This cytosolic process is just the opening act, preparing the resulting molecules to dive deeper into the cellular powerhouse known as the mitochondrion.
As metabolic flow enters the depicted mitochondrial structures, the true complexity of intermediary metabolism reveals itself. The carbon skeletons derived from glucose enter the mitochondrial matrix, a highly specialized environment designed for maximal energy extraction. Here, cycles churn continuously, and electron carriers like NADH and FADH2 are loaded with high-energy electrons. The visual represents how these interconnected loops never truly stop; they continuously feed into one another to maintain the delicate energetic balance required by the cell.
Ultimately, this introductory slide serves as the perfect roadmap. It shows that intermediary metabolism is not just a random assortment of enzymes, but a highly coordinated, integrated system. The arrows flowing from the cytoplasm into the mitochondria illustrate a beautiful biological assembly line. By mastering this blueprint, students can anticipate how a disruption in a specific enzyme might ripple through the entire cellular ecosystem, causing metabolic diseases and severe energy failures.
Slide 2: The Core Dichotomy of Intermediary Metabolism
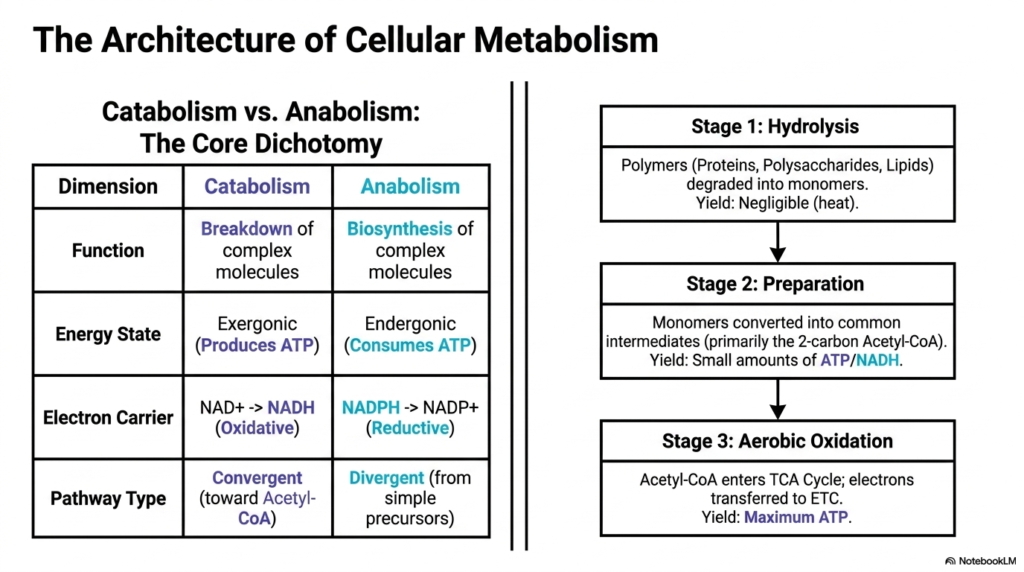
When diving into intermediary metabolism, one must first grasp its fundamental architecture. This architecture is defined by a beautiful dichotomy: the opposing yet complementary forces of catabolism and anabolism. Catabolism is the destructive phase that breaks down complex molecules into simpler ones, an exergonic process that produces much-needed ATP. Conversely, anabolism is the constructive phase, an endergonic process that consumes ATP to build complex tissues and cellular machinery. These twin pillars govern all energy flux in the human body.
Within the grand scheme of intermediary metabolism, catabolic pathways are generally oxidative and convergent. They strip electrons from fuel molecules, transferring them to carriers like NAD+ to form NADH, and they funnel diverse nutrients down into a common denominator: Acetyl-CoA. On the other hand, anabolic pathways are reductive and divergent, often using the reducing power of NADPH to build a vast array of complex molecules from simple precursors. This careful balancing act prevents energetic waste.
To further organize this complexity, intermediary metabolism breaks the catabolic breakdown of nutrients into three distinct stages. Stage one involves the simple hydrolysis of large polymers. Proteins, polysaccharides, and lipids are degraded into their respective monomers, such as amino acids and glucose. This initial stage yields negligible energy but is absolutely necessary to prepare the raw materials for cellular entry and further processing. It is the digestive prelude to cellular respiration.
Stages two and three represent the metabolic heavy lifting. In stage two, those monomers are converted into common intermediates, primarily the two-carbon powerhouse molecule, Acetyl-CoA, yielding a small amount of ATP and NADH. Finally, stage three is aerobic oxidation. The Acetyl-CoA enters the TCA cycle, transferring its remaining electrons to the electron transport chain. This final stage is where the vast majority of cellular ATP is synthesized, completing the energy extraction process.
Slide 3: The Energy Transducers of Intermediary Metabolism
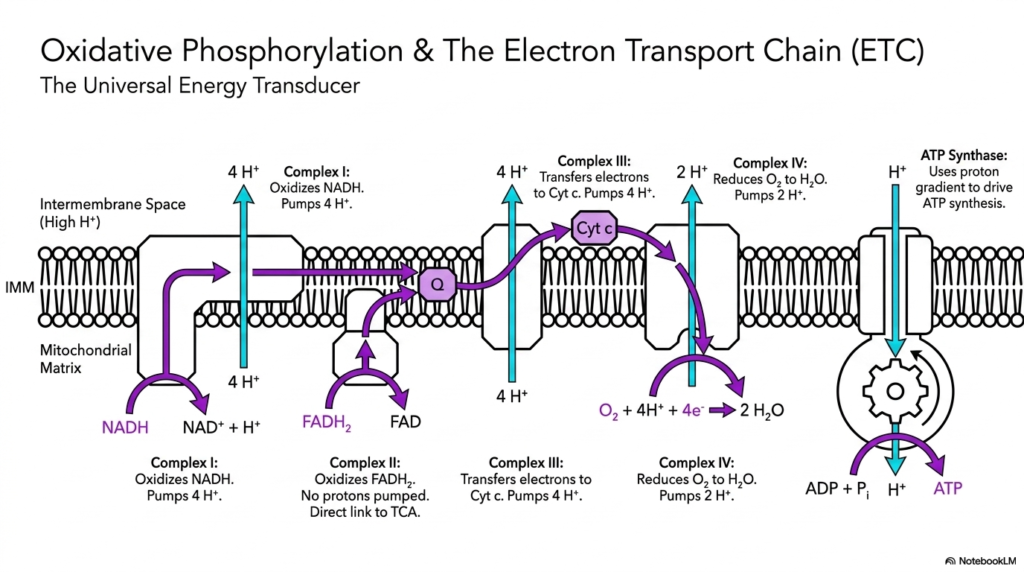
The climax of cellular energy production resides in the inner mitochondrial membrane, a process formally known as oxidative phosphorylation. This is arguably the most crucial phase of intermediary metabolism, serving as the cell’s universal energy transducer. Here, the electron transport chain uses the reducing equivalents NADH and FADH2, which were painstakingly generated during earlier metabolic cycles. These electron carriers deliver their high-energy cargo to a series of protein complexes embedded in the lipid membrane.
Complex I starts the sequence by oxidizing NADH, while Complex II uniquely oxidizes FADH2 and provides a direct physical link to the TCA cycle. As electrons flow through these complexes, releasing energy step by step, that energy is not simply lost as heat. Instead, intermediary metabolism brilliantly harnesses this electron flow to pump protons from the mitochondrial matrix into the intermembrane space. Complex I pumps four protons, Complex III pumps another four, and Complex IV pumps two, creating a massive electrochemical gradient.
This proton gradient is essentially a biological battery, heavily charged and ready to do vital cellular work. At Complex IV, the flowing electrons finally meet their ultimate acceptor: molecular oxygen. The oxygen organisms breathe is reduced to harmless water, a vital step that keeps the entire electron transport chain from backing up. If oxygen is deprived, this entire segment of intermediary metabolism grinds to a halt, leading to rapid cellular death due to a profound lack of ATP production.
The grand finale of this process features a remarkable molecular machine called ATP Synthase. As protons naturally attempt to flow back into the matrix to relieve the gradient’s immense pressure, they are forced to travel through the ATP Synthase channel. This downward flow physically spins the enzyme’s internal rotor, mechanically driving the synthesis of ATP from ADP and inorganic phosphate. It is a stunning display of molecular machinery that provides the bulk of the energy necessary for human life.
Slide 4: Glycolysis as the Foundation of Intermediary Metabolism
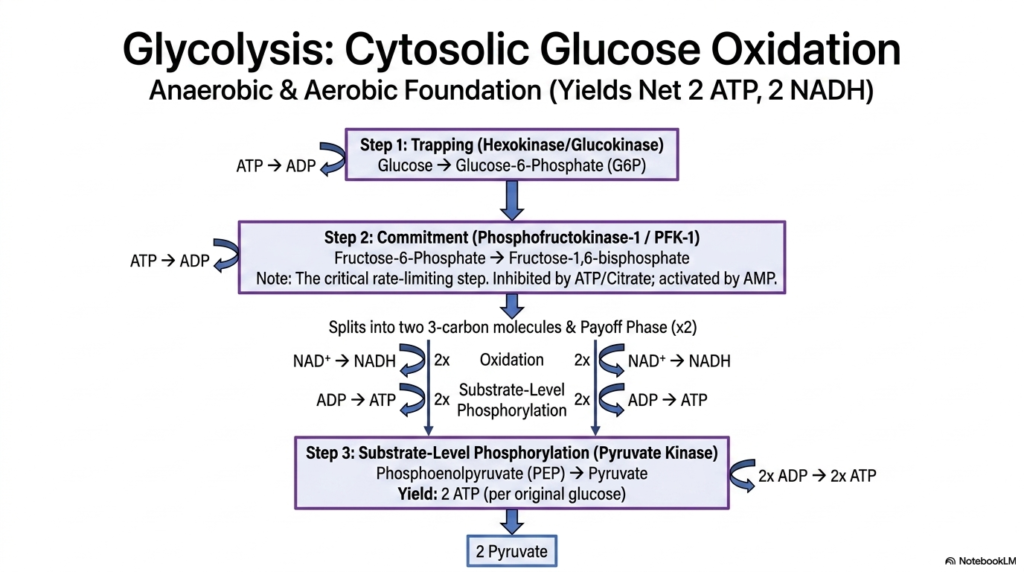
The oxidation of glucose begins in the cellular cytosol through a highly conserved ten-step pathway known as glycolysis. This pathway is a cornerstone of intermediary metabolism, serving as the primary energy-yielding process in both aerobic and anaerobic cellular environments. Because it does not strictly require oxygen, glycolysis is an ancient metabolic pathway that provides a rapid, albeit modest, supply of ATP. The process meticulously breaks down a six-carbon glucose molecule into two distinct three-carbon pyruvate molecules.
Glycolysis is carefully regulated by key enzymes, starting with the initial trapping phase. Hexokinase, or Glucokinase in the liver, phosphorylates glucose into Glucose-6-Phosphate using an ATP molecule. This addition of a negative charge permanently traps the sugar inside the cell. The most critical regulatory checkpoint in this branch of intermediary metabolism is the commitment step catalyzed by Phosphofructokinase-1 (PFK-1). PFK-1 commits the sugar to the glycolytic pathway and is highly sensitive to the cell’s energy state, inhibited by high ATP and activated by AMP.
Following this irreversible commitment, the six-carbon molecule is cleaved into two three-carbon halves, marking the start of the crucial payoff phase. It is here that intermediary metabolism finally begins to yield a positive return on its initial energy investment. Through the concurrent processes of oxidation and substrate-level phosphorylation, the pathway generates energy-rich molecules. Substrate-level phosphorylation directly transfers a high-energy phosphate group from a substrate to ADP to form ATP, occurring independently of the electron transport chain.
The final step of the pathway is catalyzed by Pyruvate Kinase, which converts Phosphoenolpyruvate into the end-product, Pyruvate, while generating yet another molecule of ATP. By the end of glycolysis, the net yield from one single glucose molecule is exactly two ATP and two NADH molecules. While this energetic yield is small compared to oxidative phosphorylation, it remains absolutely vital for cells like erythrocytes that lack mitochondria entirely, and for muscle tissues experiencing rapid bursts of high-intensity exertion.
Slide 5: Gluconeogenesis in Intermediary Metabolism
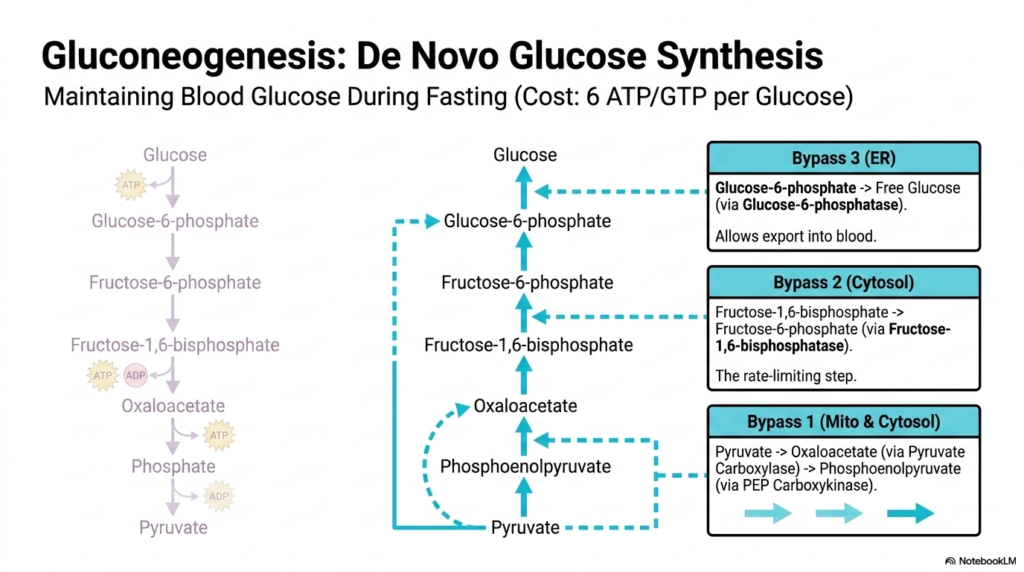
While glycolysis efficiently breaks down glucose, the body must also have a mechanism to synthesize it from scratch during prolonged fasting or intense exercise. This critical process is called gluconeogenesis, a vital, life-saving component of intermediary metabolism primarily orchestrated by the liver. Maintaining adequate blood glucose levels is absolutely essential, as specialized tissues such as the brain and red blood cells rely almost exclusively on a constant supply of this sugar. However, synthesizing glucose de novo is a highly energy-intensive endeavor.
Gluconeogenesis is deliberately not just glycolysis operating in reverse. While glycolysis shares several reversible enzymatic steps, there are three distinct irreversible hurdles that must be explicitly bypassed. This clever biochemical rerouting is a hallmark of intermediary metabolism, ensuring that neither pathway is simultaneously active, which would create a futile, energy-wasting cycle. Creating a single glucose molecule requires a hefty six ATP or GTP equivalents, underscoring the body’s commitment to maintaining systemic blood sugar levels.
The first major bypass occurs between the mitochondria and cytosol, converting Pyruvate back into Phosphoenolpyruvate. This requires two specialized enzymes working in tandem: Pyruvate Carboxylase and PEP Carboxykinase, detouring through a critical oxaloacetate intermediate. The second bypass is the major rate-limiting step in this pathway, occurring primarily in the cytosol. Here, the enzyme Fructose-1,6-bisphosphatase removes a phosphate group to form Fructose-6-phosphate, an enzyme heavily regulated by the same hormonal signals that control glycolysis but in the exact opposite direction.
The third and final bypass is localized strictly to the endoplasmic reticulum, utilizing the specialized enzyme Glucose-6-phosphatase to convert Glucose-6-phosphate into free, unphosphorylated glucose. This crucial step in intermediary metabolism successfully removes the trapping phosphate group, allowing the newly synthesized glucose to be exported from liver cells and released into the systemic bloodstream. This dynamic interplay showcases how the body dynamically shifts gears to seamlessly adapt to a fed or fasted physiological state.
Slide 6: The Amphibolic Core of Intermediary Metabolism

Deep within the mitochondrial matrix lies the Citric Acid Cycle, widely known as the TCA or Krebs cycle. This spinning wheel of biochemical reactions is truly the central amphibolic core of intermediary metabolism. It is uniquely classified as ‘amphibolic’ because it serves crucial dual purposes: it acts as the final common pathway for the catabolic oxidation of all macronutrients and provides critical precursors for anabolic biosynthesis. It is the central roundabout where multiple metabolic pathways intersect and freely exchange materials.
The cycle officially begins when the two-carbon Acetyl-CoA condenses with the four-carbon oxaloacetate to form the distinct six-carbon molecule, citrate. Through a series of elegant decarboxylations and progressive oxidations, the cycle systematically extracts high-energy electrons from the carbon skeleton. For every single turn of the cycle—representing the processing of one Acetyl-CoA molecule—the energetic yield is highly impressive. The cycle directly produces three molecules of NADH, one molecule of FADH2, and one GTP, while releasing two molecules of carbon dioxide as waste.
However, the TCA cycle is far more than just a biochemical furnace for burning cellular fuel. It is deeply embedded in the complex synthetic aspects of intermediary metabolism. Several intermediate molecules are continuously siphoned off from the cycle to build other vital cellular components. For instance, citrate can readily exit the mitochondria and enter the cytosol to serve as a crucial precursor for fatty acid synthesis, thereby converting excess carbohydrate calories into long-term lipid storage.
Other cycle intermediates are equally vital for the synthesis of structural and functional biomolecules. Alpha-ketoglutarate serves as a direct precursor for the synthesis of glutamate and other essential amino acids. Succinyl-CoA is a mandatory building block for synthesizing heme, the crucial iron-carrying component of hemoglobin. Finally, oxaloacetate is not only the cycle’s starting material but also a primary substrate for gluconeogenesis. This profound interconnectedness within intermediary metabolism highlights why the TCA cycle is the master regulator of cellular function.
Slide 7: Lipid Breakdown in Intermediary Metabolism
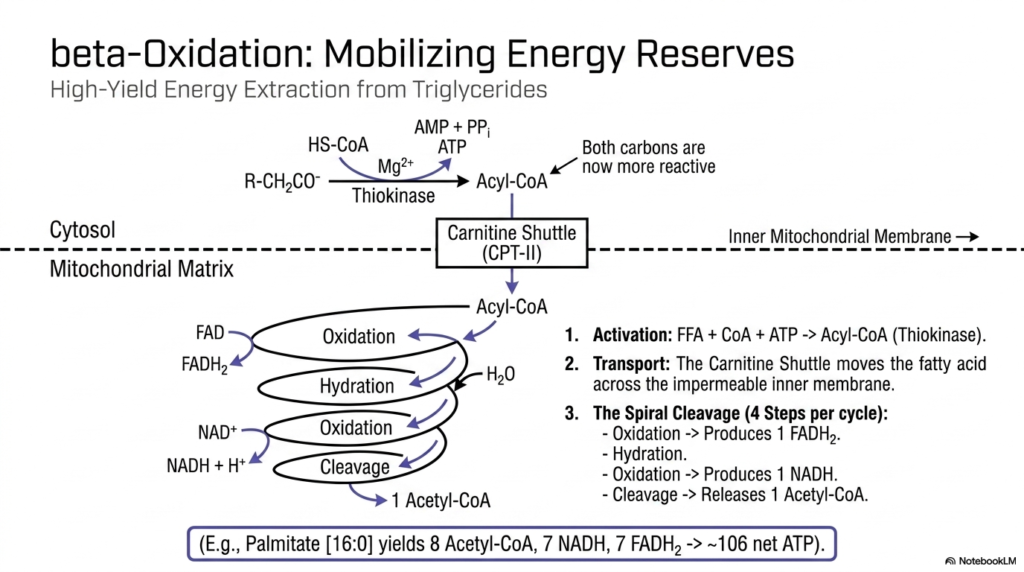
When carbohydrate stores become depleted during fasting, the body brilliantly shifts its metabolic reliance to its most concentrated form of stored energy: triglycerides. The breakdown of these fats into usable cellular energy is a highly efficient segment of intermediary metabolism, formally known as beta-oxidation. Occurring entirely within the protected confines of the mitochondrial matrix, beta-oxidation is a cyclical, spiral process that progressively chops down long-chain fatty acids into two-carbon Acetyl-CoA units, extracting substantial amounts of reducing power.
Before the actual breakdown sequence can even begin, the free fatty acid must be activated in the cytosol by attaching to Coenzyme A, an ATP-dependent process mediated by the enzyme thiokinase. However, this activated Acyl-CoA cannot cross the inner mitochondrial membrane on its own. Intermediary metabolism elegantly overcomes this physical barrier via the Carnitine Shuttle system. The enzyme CPT-I attaches carnitine to the fatty acid, allowing it to be ferried across the impermeable membrane, where CPT-II then rapidly reattaches the CoA inside the matrix.
Once safely positioned inside the mitochondrial matrix, the actual spiral cleavage process begins. Each individual cycle consists of four distinct and highly conserved steps: oxidation, hydration, a second oxidation, and finally cleavage. The first oxidation generates one FADH2 molecule, while the second oxidation yields one NADH molecule. The final cleavage step releases one two-carbon Acetyl-CoA unit, leaving the original fatty acid chain exactly two carbons shorter, ready to undergo the four-step cycle all over again until the entire chain is fully processed.
The overall energy yield from complete beta-oxidation is simply staggering, clearly showcasing the incredibly high-yield nature of lipid intermediary metabolism. For example, the complete oxidation of a single 16-carbon palmitate molecule requires exactly seven passes through the enzymatic spiral. This generates 8 Acetyl-CoA molecules, 7 NADH, and 7 FADH2. When fully processed through the TCA cycle and electron transport chain, this equates to roughly 106 net ATP, far surpassing the energy extracted from a single molecule of glucose.
Slide 8: Biosynthesis within Intermediary Metabolism
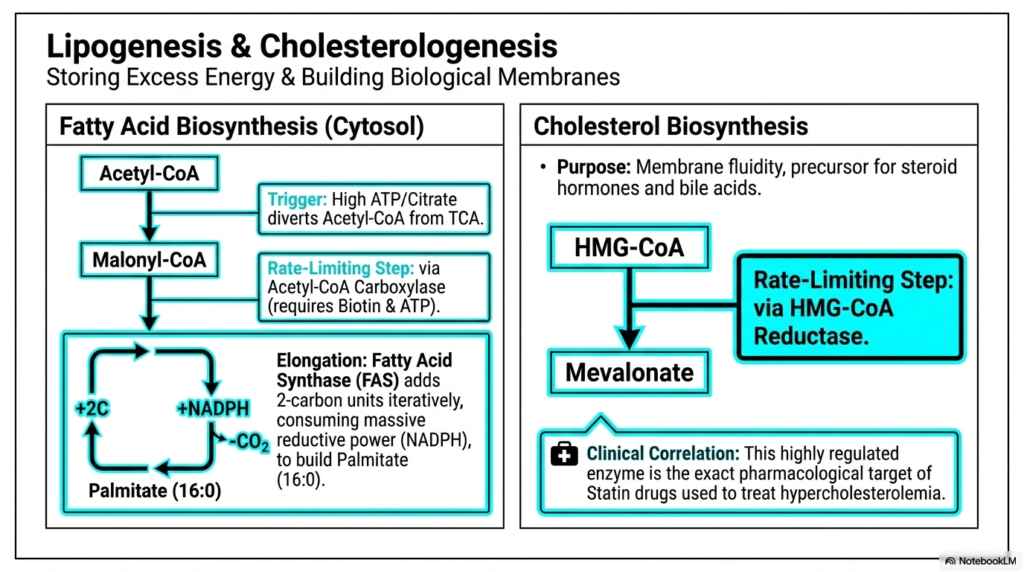
While beta-oxidation efficiently handles lipid breakdown, the cell must also possess robust mechanisms for storing excess dietary energy and synthesizing complex structural lipids. Lipogenesis and cholesterologenesis constitute the primary anabolic arm of lipid metabolism within intermediary metabolism. These crucial pathways occur directly in the cytosol and are highly active when cellular ATP and citrate levels are abundant. This abundance signals that the cell has easily met its immediate energy demands and should logically divert excess Acetyl-CoA toward long-term storage.
Fatty acid biosynthesis relies heavily on the crucial, rate-limiting enzyme Acetyl-CoA Carboxylase (ACC). ACC strictly uses biotin and ATP to convert starting Acetyl-CoA into Malonyl-CoA. From there, the massive multi-enzyme complex known as Fatty Acid Synthase (FAS) takes complete control. FAS acts as an incredible molecular assembly line within intermediary metabolism, iteratively adding two-carbon units to a constantly growing fatty acid chain. This process consumes a massive amount of reductive power in the form of NADPH to eventually build a complete 16-carbon palmitate molecule.
Functioning in strict parallel with fatty acid synthesis is the complex synthesis of cholesterol, a crucial molecule that maintains membrane fluidity and serves as an essential precursor for steroid hormones and bile acids. The cholesterol synthesis pathway shares early precursors with ketogenesis but diverges permanently after the formation of HMG-CoA. This highly specialized pathway highlights the sheer biological versatility of intermediary metabolism to construct widely different biological structures from the exact same simple two-carbon starting material.
The rate-limiting step in cholesterol biosynthesis is catalyzed by the enzyme HMG-CoA Reductase, which irreversibly converts HMG-CoA into mevalonate. This specific regulatory enzyme holds immense clinical and pharmacological significance. It is the exact molecular target of statin drugs, a widespread medication class universally used to treat hypercholesterolemia. By competitively inhibiting this singular step of intermediary metabolism, statins effectively bottleneck the entire pathway, drastically lowering systemic cholesterol levels and reducing cardiovascular disease risk.
Slide 9: Nitrogen Management and Intermediary Metabolism
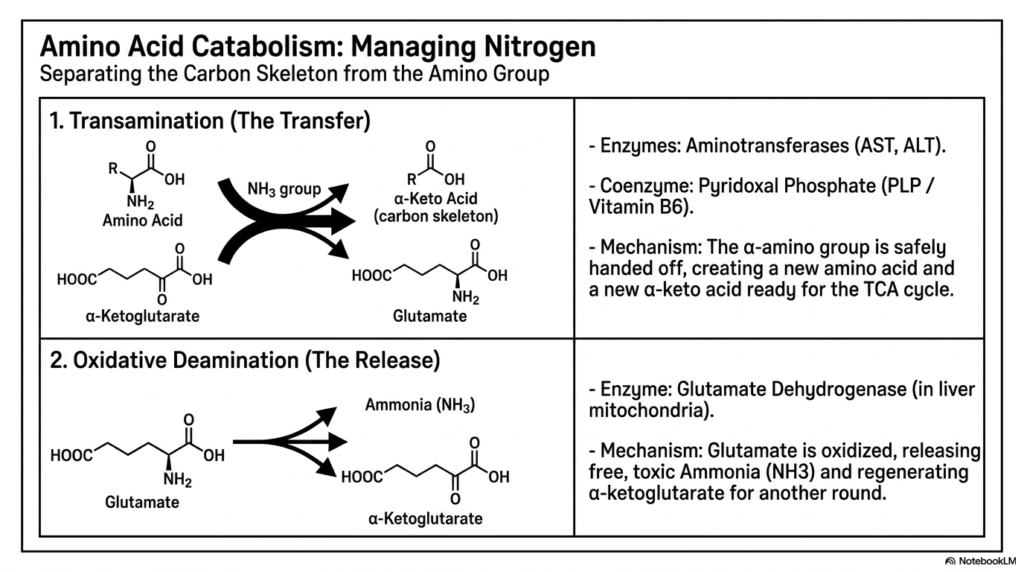
Unlike carbohydrates and lipids, which are composed primarily of carbon, hydrogen, and oxygen, cellular proteins present a unique metabolic challenge due to their high nitrogen content. Amino acid catabolism requires highly dedicated, specialized pathways within intermediary metabolism to safely separate the valuable carbon skeleton from the potentially toxic amino group. This careful separation ensures that the carbon backbone can be utilized for energy production or glucose synthesis, while the nitrogen is safely and efficiently packaged for physiological excretion.
The first step in this delicate metabolic processing is known as transamination. This represents the safe, temporary transfer of the alpha-amino group from an amino acid to an alpha-keto acid, typically alpha-ketoglutarate. Catalyzed by key aminotransferases such as AST and ALT, this reaction fundamentally swaps the functional groups. It relies entirely on the essential coenzyme Pyridoxal Phosphate, derived directly from Vitamin B6. This initial step of intermediary metabolism essentially hands off the nitrogen, creating a new amino acid and leaving behind a bare carbon skeleton ready for the TCA cycle.
Once the cellular nitrogen is temporarily parked on glutamate, the second major phase, oxidative deamination, begins. This phase is predominantly executed in the liver mitochondria by the highly active enzyme Glutamate Dehydrogenase. During this specific process, glutamate is aggressively oxidized, smoothly regenerating the alpha-ketoglutarate needed to sustain further upstream transamination reactions. However, this absolutely critical step of intermediary metabolism also directly releases the amino group into the cellular environment as free ammonia (NH3).
The biological generation of free ammonia is an incredibly high-stakes event. Ammonia is intensely toxic, particularly to the delicate central nervous system, and its unchecked accumulation can quickly lead to lethal encephalopathy. Therefore, the oxidative deamination step must be tightly and permanently coupled with immediate downstream detoxification pathways. The body’s elegant solution to this toxic byproduct seamlessly connects the breakdown of proteins directly into the subsequent specialized cycle designed specifically for rapid nitrogen disposal.
Slide 10: The Urea Cycle in Intermediary Metabolism
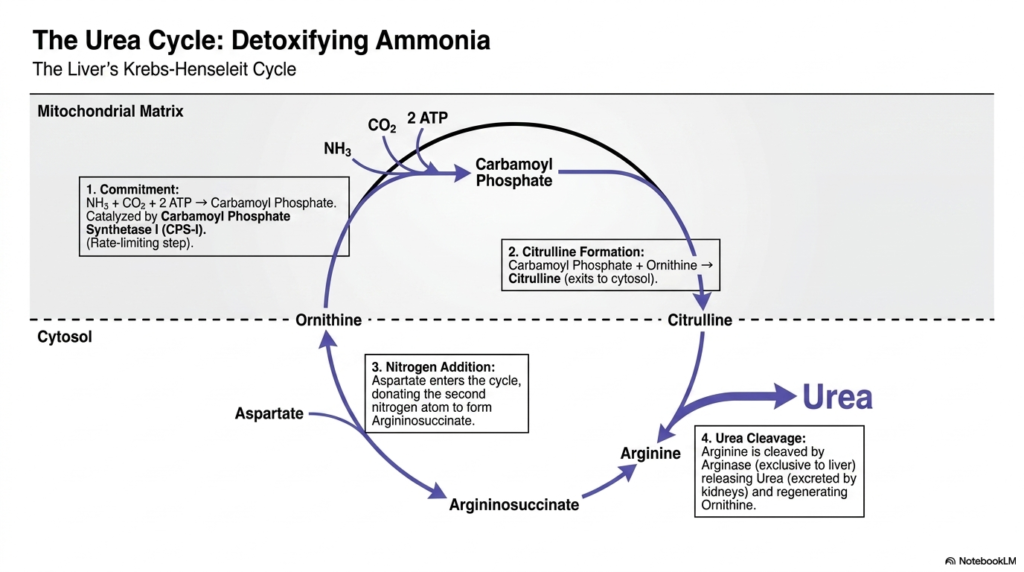
To effectively handle the highly toxic ammonia generated by amino acid breakdown, the liver utilizes a brilliant biochemical cycle known as the Urea Cycle, or the Krebs-Henseleit cycle. This intricate cycle is a quintessential example of highly organ-specific intermediary metabolism, as the liver is uniquely equipped with the full complement of required enzymes to execute it. The cycle’s primary biological objective is to convert highly toxic, highly soluble ammonia into urea, a relatively benign, easily excretable compound that can be safely transported to the kidneys.
The cycle is aggressively triggered deep within the mitochondrial matrix by its primary commitment step. Free ammonia is combined with ambient carbon dioxide and two ATP molecules to rapidly form Carbamoyl Phosphate. This crucial, rate-limiting chemical reaction is catalyzed by Carbamoyl Phosphate Synthetase I (CPS-I). Following this, carbamoyl phosphate donates its moiety directly to ornithine, creating citrulline. This step brilliantly highlights the physical compartmentalization often seen in intermediary metabolism, as citrulline must then be exported from the mitochondria into the cytosol to continue the cycle.
Once residing in the cytosol, a second nitrogen atom is strategically incorporated into the cycle. This second nitrogen is donated directly by the amino acid aspartate, which then combines with citrulline to form the complex molecule argininosuccinate. This massive molecule is subsequently cleaved to form arginine. This intricate shuffling of atoms perfectly demonstrates the profound resourcefulness of intermediary metabolism, seamlessly utilizing multiple distinct molecular sources to gather the exact two nitrogen atoms required to assemble a single molecule of urea.
The final enzymatic step of the cycle relies entirely on arginase, an enzyme active exclusively in the liver. Arginase forcefully cleaves arginine, releasing the final urea molecule directly into the bloodstream to be cleared by the kidneys, while simultaneously regenerating ornithine to be shuttled back into the mitochondria for another round. A genetic failure at any point in this loop of intermediary metabolism rapidly leads to hyperammonemia, thereby strongly reinforcing the critical, life-sustaining nature of these nitrogen-disposal pathways.
Slide 11: Nucleotide Synthesis in Intermediary Metabolism
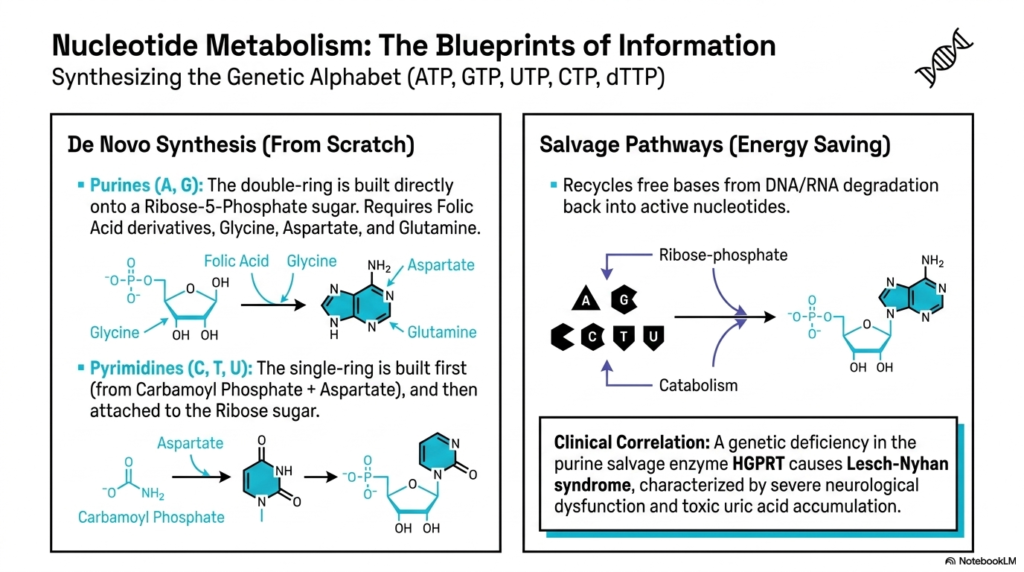
Beyond mere energy extraction and macronutrient storage, intermediary metabolism is also responsible for fabricating the very biological blueprints of cellular information: nucleotides. The constant synthesis of ATP, GTP, UTP, CTP, and dTTP is absolutely essential for vital functions such as DNA replication, RNA transcription, and the maintenance of the primary cellular energy currency. The human body uses two distinct strategies to acquire these genetic alphabets: building them entirely from scratch via de novo synthesis or efficiently recycling them through energy-saving salvage pathways.
De novo synthesis is an incredibly complex process and requires significant, sustained energy investments. For purines like Adenine and Guanine, the massive double-ring structure is built directly onto a foundational Ribose-5-Phosphate sugar base. This demanding pathway draws heavily on resources from various corners of intermediary metabolism, strictly requiring inputs such as folic acid derivatives, glycine, aspartate, and glutamine. Conversely, pyrimidines are constructed differently; their single-ring structure is cleanly assembled first from carbamoyl phosphate and aspartate, and only later structurally attached to the necessary ribose sugar.
Because de novo synthesis is so metabolically demanding, cells heavily prefer to utilize nucleotide salvage pathways whenever possible. As DNA and RNA naturally degrade over time, free nucleotide bases are constantly released into the environment. Salvage pathways rapidly reattach these free bases to a ribose-phosphate backbone, smoothly returning them directly into the active nucleotide pool. This elegant recycling system is a prime example of how intermediary metabolism actively optimizes resource management, drastically reducing the cellular ATP expenditure required to maintain functional genetic materials.
The absolutely critical importance of these specific salvage pathways is starkly illustrated by clinical pathology. A severe genetic deficiency in the purine salvage enzyme HGPRT directly leads to Lesch-Nyhan syndrome. Without this specific enzyme, the salvage pathway fails completely, forcing purines to degrade into highly toxic levels of uric acid while simultaneously driving excessive, unchecked de novo synthesis. This tragic, permanent disruption in intermediary metabolism results in severe neurological dysfunction, highlighting the incredibly delicate biochemical balance required for normal human physiological function.
Slide 12: The Grand Integration of Intermediary Metabolism
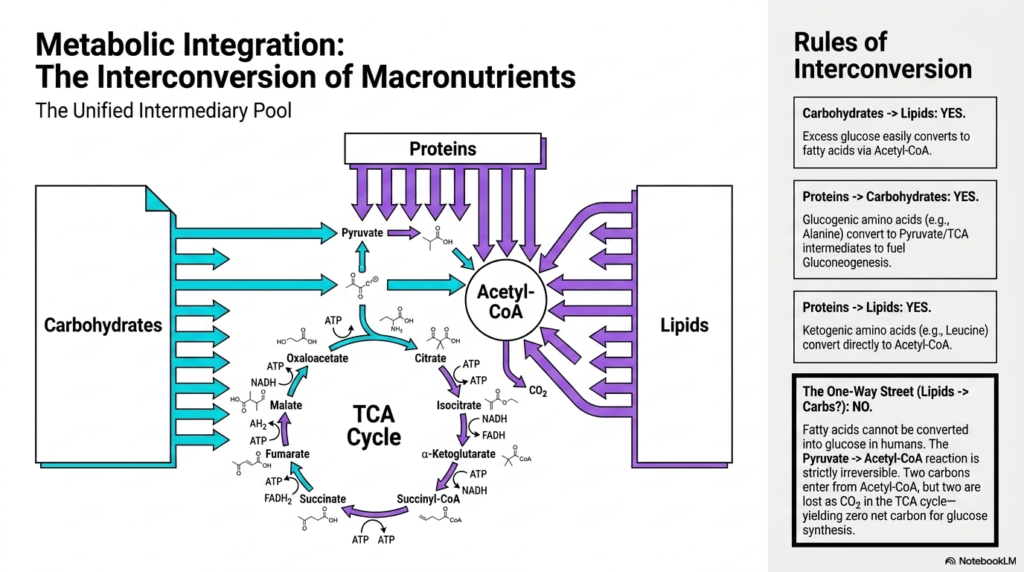
To truly master biochemistry, one must move far beyond memorizing isolated pathways and fully appreciate the sheer scale of metabolic integration. This final slide beautifully maps the unified pool of intermediary metabolism, clearly demonstrating how the major macronutrients—carbohydrates, proteins, and lipids—constantly interconvert to dynamically meet the body’s changing needs. The central hub for all this biochemical traffic is the TCA cycle and its immediate precursors, like Pyruvate and Acetyl-CoA, which essentially serve as the universal biological crossroads.
The biological rules of this interconversion are truly fascinating. Carbohydrates can very easily be converted into complex lipids. When systemic glucose is highly abundant, it is rapidly broken down into Acetyl-CoA, which, instead of burning in the TCA cycle, is heavily diverted into fatty acid synthesis. Proteins are similarly incredibly versatile within intermediary metabolism. Glucogenic amino acids can be neatly converted into pyruvate or TCA intermediates to fuel gluconeogenesis, while ketogenic amino acids can convert directly into Acetyl-CoA to synthesize body fats or vital ketone bodies.
However, there is one exceptionally strict biological one-way street: in humans, lipids absolutely cannot be converted back into carbohydrates. Fatty acids strictly break down exclusively into Acetyl-CoA units. The specific enzymatic reaction converting Pyruvate to Acetyl-CoA is strictly and completely irreversible. Furthermore, when the two carbons of Acetyl-CoA enter the spinning TCA cycle, exactly two carbons are lost as exhaled carbon dioxide before oxaloacetate is finally regenerated. Therefore, this unbending rule of intermediary metabolism dictates that fatty acids always yield a net of zero carbon for de novo glucose synthesis.
This final integration slide perfectly encapsulates the biochemical machine’s ultimate elegance and incredible precision. By strictly maintaining these biological rules and highly interconnected pathways, intermediary metabolism ensures that the brain gets a constant supply of glucose, excess dietary energy is safely stored as fat for the long term, and proteins can act as emergency fuel when desperately needed. It is a perfectly balanced, beautifully automated biological system that gracefully sustains the sheer complexity of human life.
Please read our Content Disclaimer Statement.
Check out our social media channels: