40. The Pentose Phosphate Pathway: Biochemical Structures and Concepts
Imagine a bustling city that needs both construction materials to build towering skyscrapers and a robust defense system to ward off relentless invaders. In the human body, cells face a remarkably similar dual challenge. They must continuously synthesize essential building blocks while constantly battling against toxic oxidative stress. This slide deck explores the molecular intricacies of cellular survival, focusing on highly specialized metabolic routes. The core purpose of this presentation is to provide a comprehensive, step-by-step molecular atlas of these biochemical shunts for medical and college students, unraveling exactly how cells maintain equilibrium.
1. Introduction to the Pentose Phosphate Pathway
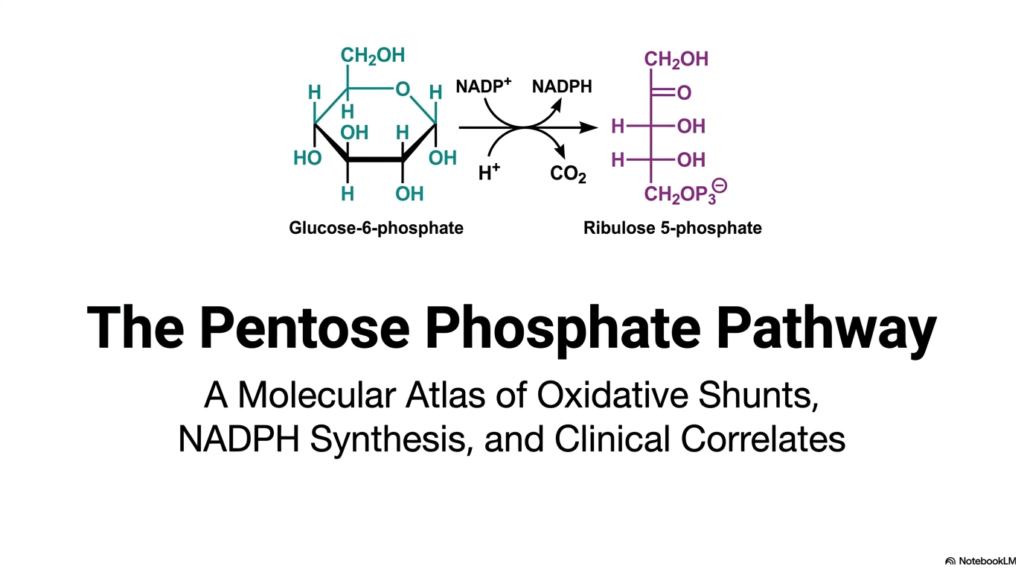
The opening slide introduces the Pentose Phosphate Pathway, a fundamental metabolic cascade that operates exclusively in the cellular cytosol. This molecular atlas provides a sweeping overview of vital oxidative shunts, the synthesis of nicotinamide adenine dinucleotide phosphate (NADPH), and essential clinical correlates. By displaying the initial transformation of glucose-6-phosphate into ribulose 5-phosphate, the graphic instantly establishes the core biochemical conversion that drives the entire sequence. The observer can clearly trace the atomic rearrangements that define this unique metabolic offshoot.
For college and medical students, mastering the Pentose Phosphate Pathway is absolutely paramount. It represents a vital cellular mechanism that intentionally diverges from standard glycolysis. Instead of generating adenosine triphosphate (ATP) for cellular energy, this alternative route serves as the principal source of reducing equivalents. The visual transformation depicted perfectly highlights the release of carbon dioxide and the simultaneous reduction of NADP+ to NADPH, illustrating the pathway’s dual role in maintaining robust metabolic health.
The Pentose Phosphate Pathway ultimately serves as the metabolic backbone for both anabolism and oxidative defense. The molecular structures shown on the slide—specifically the cyclic glucose derivative transforming into a linear five-carbon sugar—demonstrate the exact atomic rearrangements required for cellular survival. This introductory schematic lays the groundwork for understanding how specific tissues harness these precise chemical reactions to synthesize complex lipids, build genetic nucleotides, and vigorously protect themselves against free-radical damage.
2. Overview of the Hexose Monophosphate Shunt in the Pentose Phosphate Pathway
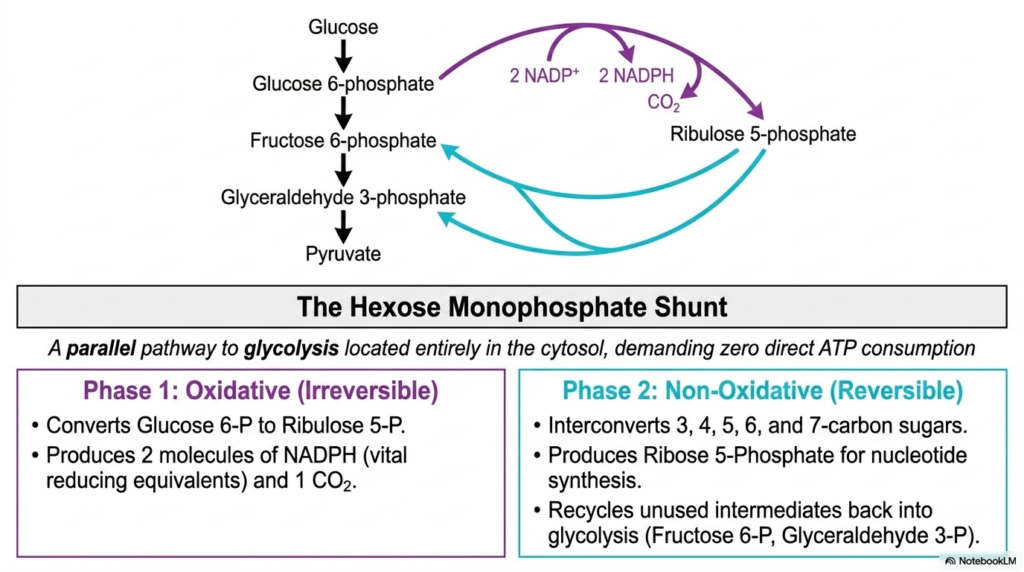
The second slide delves into the overall architecture of the Pentose Phosphate Pathway, also commonly referred to in the medical literature as the hexose monophosphate shunt. This schematic brilliantly illustrates how the cascade functions as a parallel metabolic route to glycolysis. Both biochemical sequences share glucose as a starting point and operate entirely within the cellular cytosol. A striking feature highlighted here is that this shunt requires absolutely no direct ATP consumption, distinguishing it significantly from other energy-intensive cellular processes.
The diagram clearly demarcates the two major phases of the Pentose Phosphate Pathway, beginning with the irreversible oxidative phase. In this initial stage, glucose 6-phosphate undergoes a permanent conversion into ribulose 5-phosphate. This transformation is biologically invaluable because it produces two distinct molecules of NADPH alongside one molecule of carbon dioxide. NADPH serves as a vital reducing equivalent, supplying the necessary electron power for various anabolic reactions and essential cellular defense mechanisms that dictate tissue survival.
Following the oxidative stage, the Pentose Phosphate Pathway gracefully transitions into the reversible non-oxidative phase. This highly dynamic segment is responsible for the complex interconversion of three-, four-, five-, six-, and seven-carbon sugars. Most notably, it generates ribose 5-phosphate, the indispensable foundational sugar required for DNA and RNA nucleotide synthesis. Furthermore, these non-oxidative reactions efficiently recycle unused intermediates, channeling them back into the main glycolytic pathway.
3. The Oxidative Cascade of the Pentose Phosphate Pathway
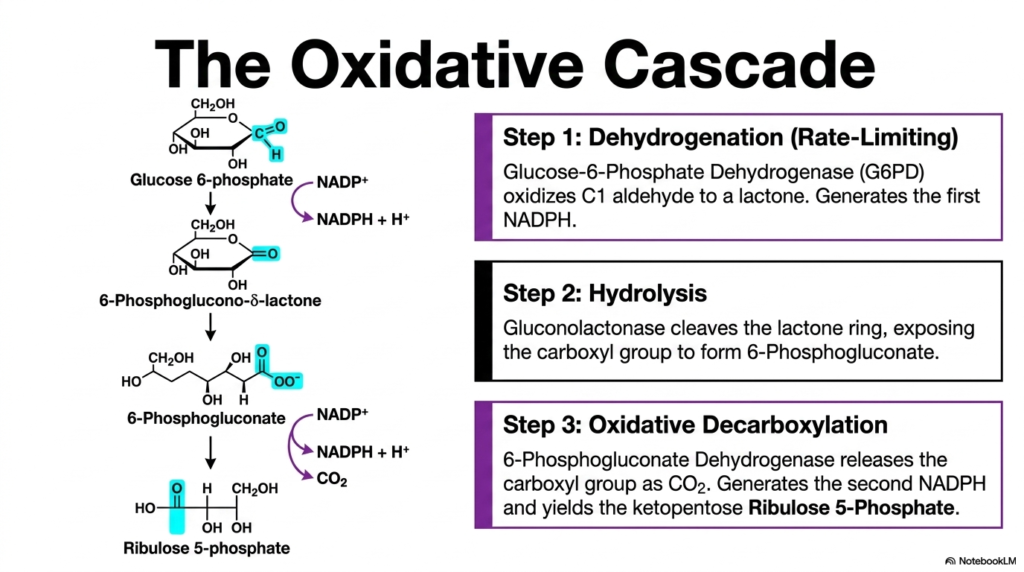
Moving deeper into the molecular specifics, the third slide maps out the irreversible oxidative cascade of the Pentose Phosphate Pathway. The graphic breaks down the initial sequence into three distinct, highly regulated enzymatic steps. The very first step is dehydrogenation, which is notably the strictly rate-limiting reaction of the entire sequence. Here, the enzyme glucose-6-phosphate dehydrogenase oxidizes the C1 aldehyde of the sugar to form a lactone, simultaneously generating the very first molecule of NADPH for the cell.
The second phase of this oxidative portion of the Pentose Phosphate Pathway involves a critical hydrolysis reaction. The enzyme gluconolactonase expertly cleaves the newly formed lactone ring. This spontaneous ring-opening event exposes the carboxyl group, resulting in the rapid formation of 6-phosphogluconate. The visual representation of molecular structures helps biochemistry students clearly trace the movement of atoms and the subtle shifts in molecular geometry that meticulously prepare the substrate for its final oxidative transformation.
The concluding step in the oxidative branch of the Pentose Phosphate Pathway is an oxidative decarboxylation event. The specialized enzyme 6-phosphogluconate dehydrogenase acts on the substrate, permanently releasing the exposed carboxyl group as carbon dioxide. This critical chemical reaction not only yields the ketopentose ribulose 5-phosphate but also, triumphantly, generates the second molecule of NADPH. Through these three coordinated enzymatic maneuvers, the cell successfully secures immense reducing power.
4. G6PD: The Gatekeeper of the Pentose Phosphate Pathway
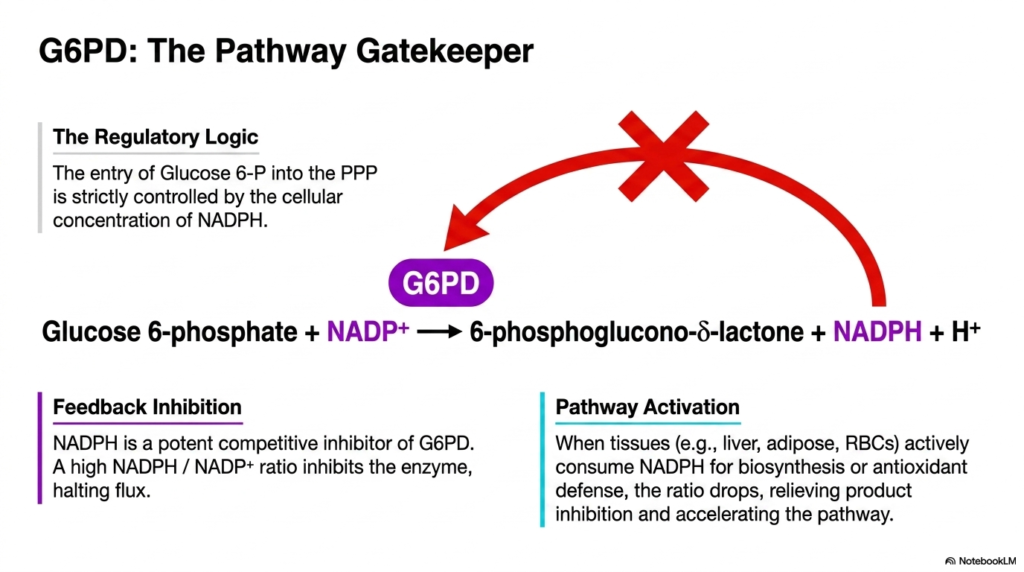
The fourth slide introduces the strict regulatory logic governing the Pentose Phosphate Pathway. At the center of this complex regulation is glucose-6-phosphate dehydrogenase (G6PD), appropriately titled the pathway gatekeeper. The entry of glucose 6-phosphate into this specific metabolic route is not random; it is meticulously controlled by the cellular concentration of NADPH. The large red restrictive arrow in the diagram emphasizes how flux through this enzymatic checkpoint is tightly modulated to meet the organism’s exact biochemical demands.
Feedback inhibition is the primary physiological mechanism controlling this initial step of the Pentose Phosphate Pathway. NADPH acts as a highly potent competitive inhibitor of the G6PD enzyme. When the cellular ratio of NADPH to NADP+ remains high, the abundant NADPH binds directly to the enzyme, effectively halting further metabolic flux. This precise feedback loop ensures that the cell does not waste valuable glucose resources producing reducing equivalents when the existing supply is already fully sufficient for immediate biochemical activities.
Conversely, pathway activation within the Pentose Phosphate Pathway occurs dynamically when tissues actively consume their NADPH reserves. In organs like the liver and adipose tissue, which heavily rely on reductive biosynthesis, or in red blood cells that require constant antioxidant defense, the cellular NADPH ratio frequently plummets. This sudden depletion relieves the product inhibition on G6PD, instantly accelerating the enzyme’s catalytic activity and restoring the crucial flow of metabolites through the cascade to rapidly replenish the lost reducing power.
5. Phase 2 Initiators in the Pentose Phosphate Pathway
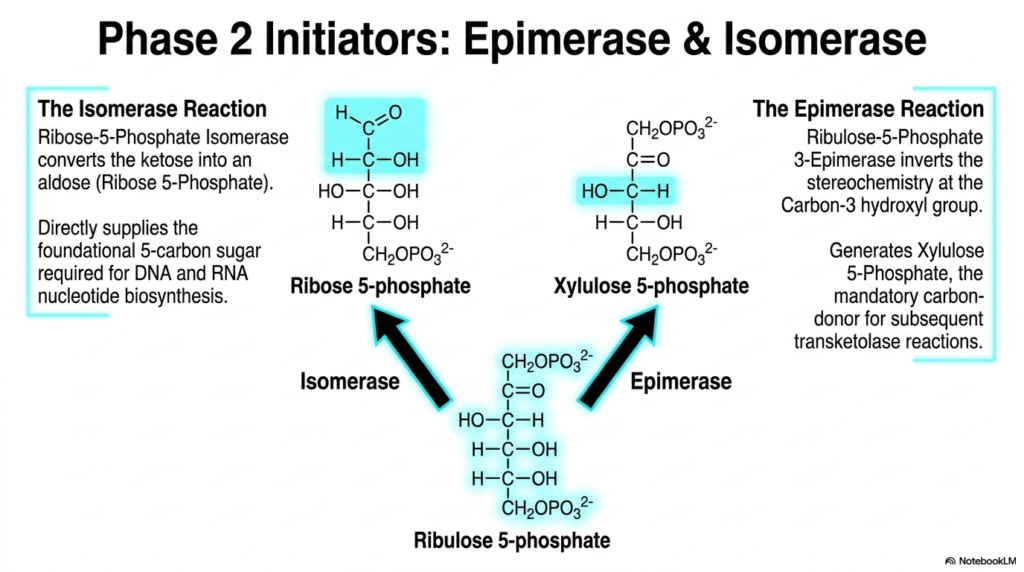
The fifth slide transitions directly into the reversible, non-oxidative branch of the Pentose Phosphate Pathway by deeply examining its two primary initiating enzymes: an epimerase and an isomerase. Ribulose 5-phosphate, the end product of the preceding oxidative phase, serves as the common foundational substrate for both of these critical reactions. The slide visually splits the metabolic pathway into two distinct directions, illustrating how a single precursor molecule can be structurally rearranged to fulfill completely different biochemical requirements within the complex cellular environment.
The isomerase reaction in the Pentose Phosphate Pathway smoothly converts the ketose sugar, ribulose 5-phosphate, into the corresponding aldose sugar, ribose 5-phosphate. This specific structural shift is of paramount biological importance for cellular replication. Ribose 5-phosphate directly supplies the foundational five-carbon sugar backbone fundamentally required for the biosynthesis of both DNA and RNA nucleotides. Without this precise enzymatic isomerization, cellular division, genetic transcription, and overall molecular replication would grind to a halt.
In tandem, the epimerase reaction within the Pentose Phosphate Pathway plays a complementary, yet entirely distinct, structural role. Ribulose-5-phosphate 3-epimerase functions by carefully inverting the stereochemistry specifically at the carbon-3 hydroxyl group. This highly subtle molecular flip generates a new molecule called xylulose 5-phosphate. This new product is not utilized for nucleotides; rather, it acts as the mandatory carbon donor for the subsequent transketolase reactions, ensuring the entire metabolic cycle can continue shifting carbons and ultimately reconnect with glycolysis.
6. The Carbon Shifters of the Pentose Phosphate Pathway
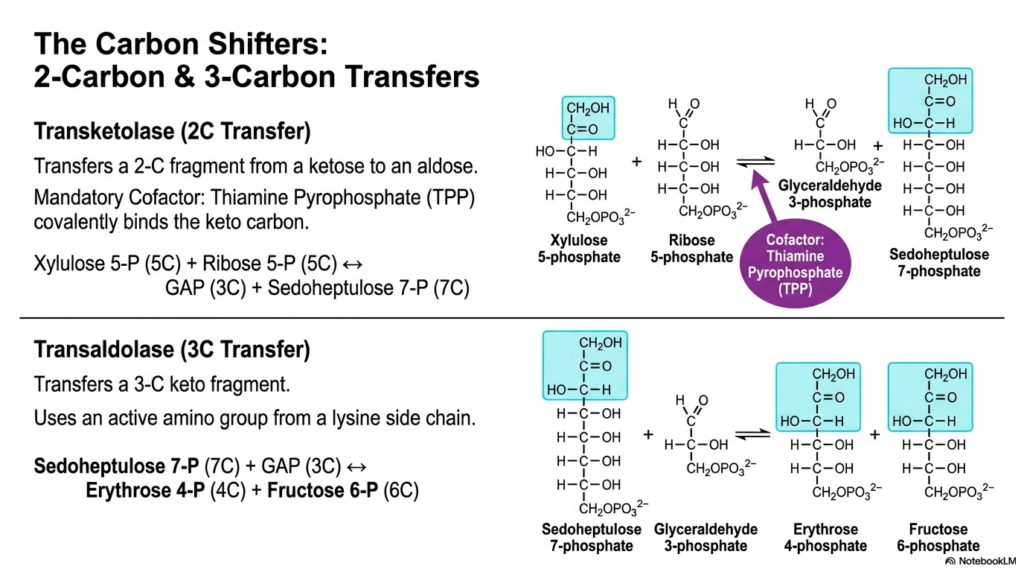
Slide six meticulously details the remarkable enzymatic machinery known as the carbon shifters, functioning entirely within the Pentose Phosphate Pathway. These specialized enzymes, specifically transketolase and transaldolase, are directly responsible for the highly complex reshuffling of carbon atoms between various sugar molecules. The transketolase enzyme specifically transfers a precise two-carbon fragment from a ketose to an aldose. The diagram clearly notes that this unique reaction requires a cofactor, thiamine pyrophosphate (TPP), which covalently binds the keto carbon to facilitate the structural transfer.
The transketolase reaction operating in the Pentose Phosphate Pathway effectively merges xylulose 5-phosphate and ribose 5-phosphate to elegantly create glyceraldehyde 3-phosphate and the unusual seven-carbon sugar, sedoheptulose 7-phosphate. Following this initial transfer, the transaldolase enzyme seamlessly takes over, specializing in the subsequent transfer of a larger three-carbon keto fragment. Unlike transketolase, transaldolase utilizes an amino group derived directly from a lysine side chain within its active site, showcasing a completely different yet equally elegant biochemical mechanism for cleaving and forming bonds.
Through the robust transaldolase reaction, the Pentose Phosphate Pathway seamlessly continues its carbon shuffling by actively reacting sedoheptulose 7-phosphate with glyceraldehyde 3-phosphate. This specific atomic exchange efficiently produces erythrose 4-phosphate and fructose 6-phosphate. By carefully mapping these complex atomic exchanges, medical and biochemistry students can truly appreciate how the cell fluidly manages its carbon resources, transforming unusual five- and seven-carbon intermediates back into highly recognizable metabolic molecules that gracefully integrate into the broader metabolic network.
7. The Net Equation and Mass Balance of the Pentose Phosphate Pathway
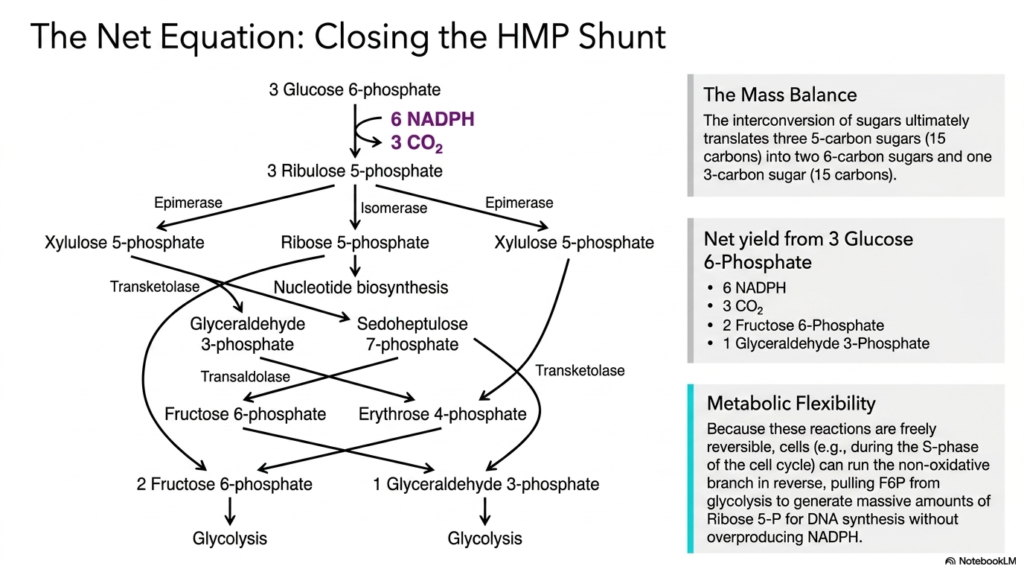
The seventh slide elegantly synthesizes the complex web of interconnected reactions by clearly presenting the net equation and mass balance of the Pentose Phosphate Pathway. When the entire hexose monophosphate shunt is visually closed, the schematic carefully traces the ultimate fate of three distinct molecules of glucose 6-phosphate. The detailed diagram accurately illustrates how the intricate interconversion of sugars ultimately translates three five-carbon sugars, totaling fifteen carbons, into exactly two six-carbon sugars and one three-carbon sugar, flawlessly maintaining the fundamental law of conservation of mass.
The net yield calculated from processing three glucose 6-phosphate molecules straight through the Pentose Phosphate Pathway is incredibly revealing to the biochemistry student. The completed cycle efficiently produces six molecules of NADPH, three molecules of carbon dioxide, two molecules of fructose 6-phosphate, and one molecule of glyceraldehyde 3-phosphate. This precise accounting thoroughly demonstrates the pathway’s extreme efficiency. It maximizes the extraction of highly valuable reducing equivalents while actively ensuring that no viable carbon atoms are wasted by the cell.
A profoundly important concept highlighted here is the extreme metabolic flexibility fundamentally inherent to the Pentose Phosphate Pathway. Because the non-oxidative reactions are freely and completely reversible, living cells can dynamically alter their metabolic flux in response to immediate physiological needs. For example, during the active S-phase of the cell cycle, a rapidly dividing cell can run the non-oxidative branch completely in reverse. This allows the cell to pull fructose 6-phosphate directly from glycolysis to generate massive amounts of ribose 5-phosphate.
8. The Currency of Anabolism: NADPH in the Pentose Phosphate Pathway
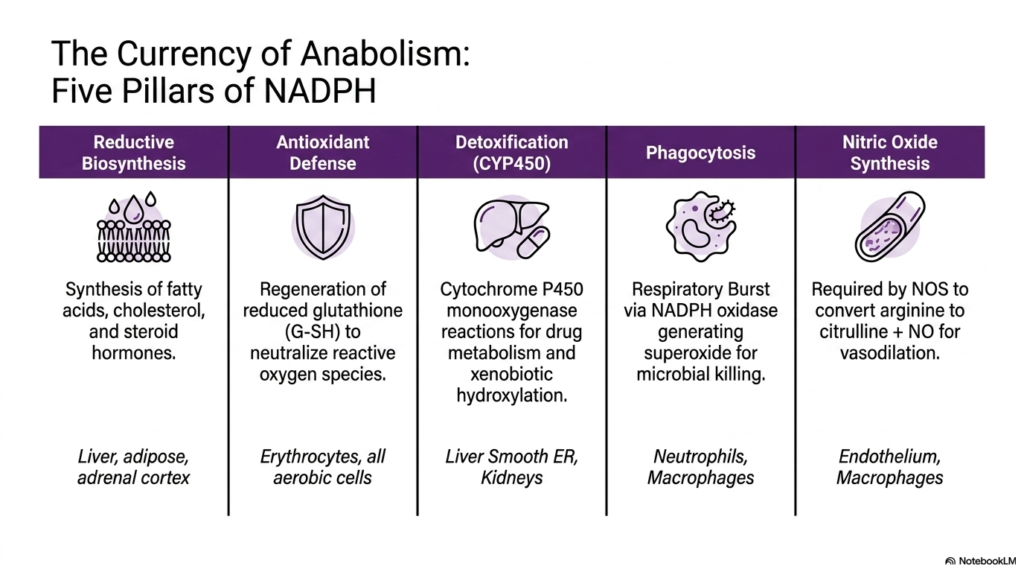
Slide eight delves deeply into the myriad physiological functions of NADPH, rightly labeling it the primary currency of anabolism, generated exclusively by the Pentose Phosphate Pathway. The graphic systematically breaks down the vast utility of this energy-rich molecule into five completely distinct physiological pillars. The first major pillar is reductive biosynthesis, a complex process that relies heavily on NADPH to produce fatty acids, cholesterol, and various steroid hormones. This essential synthetic activity is predominantly localized in specialized tissues such as the liver, adipose tissue, and the adrenal cortex.
The subsequent pillars clearly highlight the critical defensive and functional roles of NADPH produced by the Pentose Phosphate Pathway. The second pillar represents crucial antioxidant defense, in which NADPH is continuously utilized by all aerobic cells, particularly fragile erythrocytes, to regenerate reduced glutathione and neutralize reactive oxygen species. The third pillar heavily involves detoxification in the liver and kidneys, where cytochrome P450 monooxygenases consume large amounts of NADPH to metabolize drugs and safely hydroxylate toxic compounds for excretion.
The final two pillars meticulously demonstrate the highly specialized immune and vascular roles that are closely linked to the Pentose Phosphate Pathway. Phagocytosis strictly relies on NADPH to forcefully power the respiratory burst via NADPH oxidase, rapidly generating superoxide radicals essential for microbial killing in neutrophils and macrophages. Furthermore, endothelial cells and macrophages absolutely require NADPH for nitric oxide synthesis, an essential physiological reaction in which the enzyme nitric oxide synthase efficiently converts arginine to citrulline, producing nitric oxide that brilliantly facilitates critical vascular dilation.
9. Neutralizing ROS: Glutathione and the Pentose Phosphate Pathway
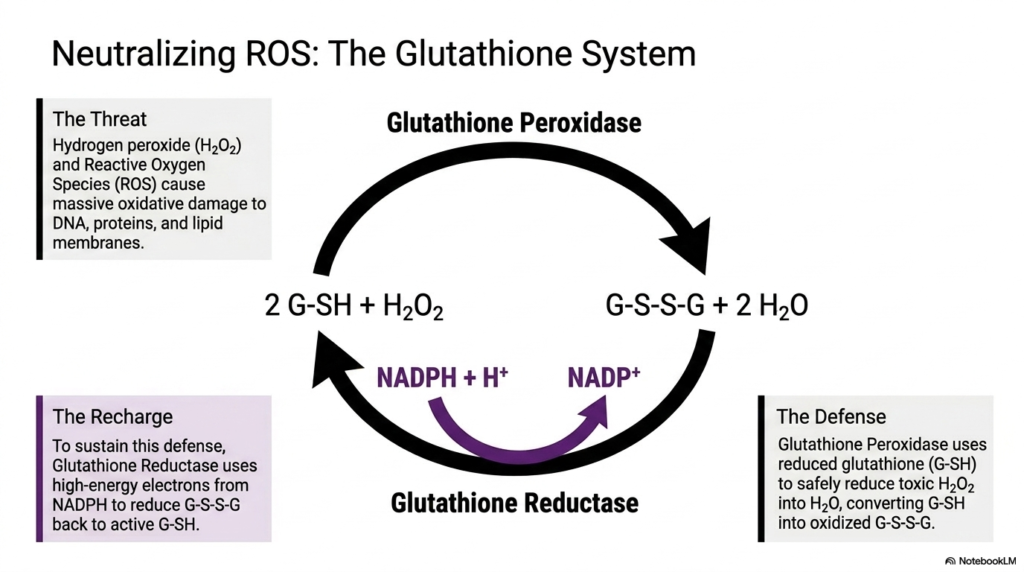
The ninth slide provides a detailed microscopic view of exactly how cells successfully neutralize dangerous reactive oxygen species (ROS), a critical defense mechanism that is entirely dependent on the Pentose Phosphate Pathway. The slide precisely identifies the primary biochemical threats: hydrogen peroxide and various rogue free radicals that cause massive, often entirely irreversible, oxidative damage to DNA, cellular proteins, and lipid membranes. To effectively combat this constant chemical assault, the vulnerable cell employs a sophisticated, cyclical glutathione defense system that essentially acts as a protective molecular shield.
At the very forefront of this biochemical defense is the robust enzyme glutathione peroxidase. This remarkable cellular protein continuously uses reduced glutathione (G-SH) to safely and efficiently reduce highly toxic hydrogen peroxide directly into harmless water molecules. In this ongoing defensive process, the highly protective reduced glutathione is inevitably converted into its oxidized, inactive disulfide form (G-S-S-G). Without a rapidly functioning mechanism to reverse this specific oxidation, the cellular shield would quickly be depleted, leaving the cell completely vulnerable to severe damage.
This is precisely where the absolute vital importance of the Pentose Phosphate Pathway immediately becomes apparent to the observer. To successfully sustain this defense and chemically recharge the entire system, the essential enzyme glutathione reductase continuously steps in. It rapidly uses the high-energy electrons supplied by NADPH—the primary molecular product of the pathway—to fully reduce the oxidized G-S-S-G back to its highly active G-SH state. This ongoing regeneration cycle actively ensures that all aerobic tissues maintain a constant state of cellular readiness.
10. Clinical Correlation: G6PD Deficiency and the Pentose Phosphate Pathway
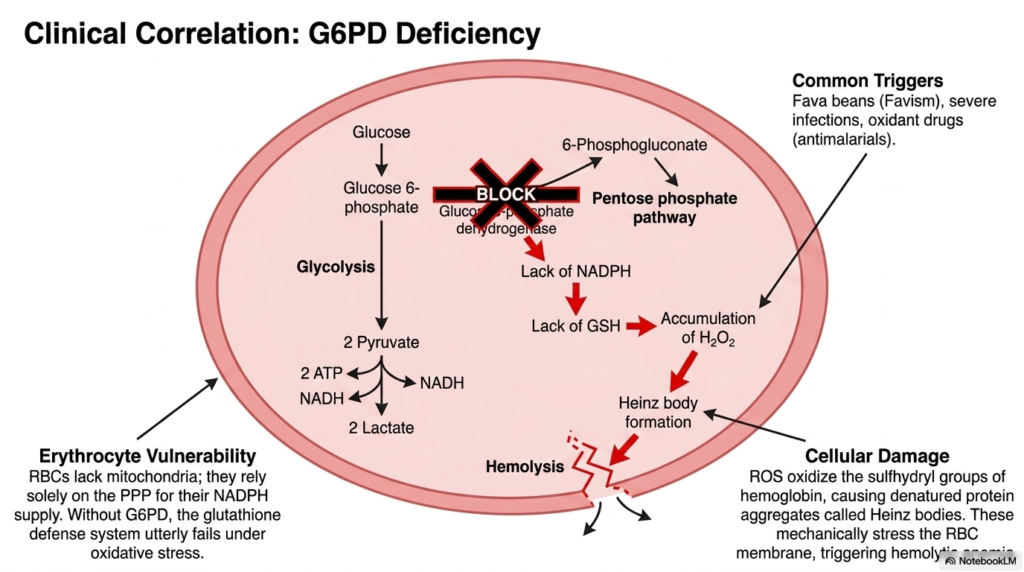
The tenth slide bridges the gap between basic biochemistry and clinical pathology by delving into G6PD deficiency, a direct and dangerous metabolic failure arising from the Pentose Phosphate Pathway. The highly detailed diagram focuses on the erythrocyte, a unique human cell that lacks functional mitochondria. Because they possess no other means of generating NADPH, red blood cells rely solely on this biochemical shunt for their cellular NADPH supply, making them exquisitely vulnerable.
Without a fully functional G6PD enzyme, the Pentose Phosphate Pathway fails to produce sufficient NADPH, directly leading to a dangerously low level of reduced glutathione. When suddenly exposed to common oxidative triggers such as fava beans, severe systemic infections, or common antimalarial drugs, the compromised glutathione defense system utterly and catastrophically fails. The slide vividly illustrates how unchecked hydrogen peroxide rapidly accumulates within the red blood cell, violently shifting the cellular environment into a highly toxic, destructive oxidative state.
This overwhelming oxidative stress directly and severely damages vital cellular components. Reactive oxygen species rapidly and irreversibly oxidize the delicate sulfhydryl groups of hemoglobin, causing the critical proteins to instantly denature and heavily precipitate into rigid aggregates clinically known as Heinz bodies. These microscopic protein clumps impose severe mechanical stress and physical damage on the highly delicate erythrocyte membrane as the compromised cell attempts to navigate narrow capillaries. This severe structural compromise inevitably and rapidly triggers total cellular rupture and massive hemolysis.
11. Diagnostic Matrix of G6PD Variants in the Pentose Phosphate Pathway
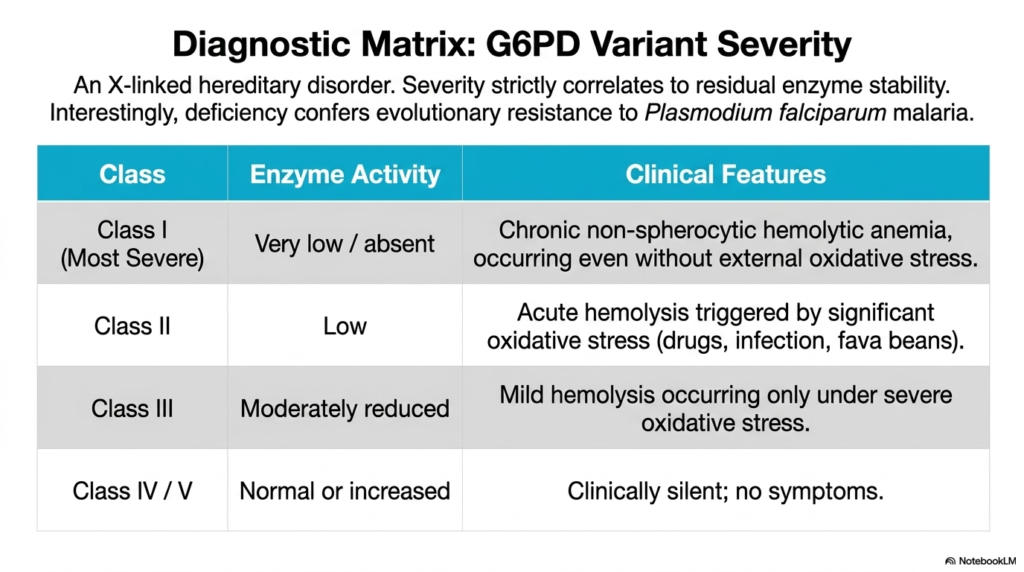
Slide eleven thoroughly expands upon the profound clinical implications of the Pentose Phosphate Pathway by presenting a highly detailed diagnostic matrix of G6PD variant severity. This specific hereditary disorder is strictly X-linked, meaning it predominantly and most severely affects human males. The comprehensive table clearly and systematically delineates how the absolute severity of the clinical phenotype strictly correlates to the precise residual stability and ongoing catalytic activity of the mutated enzyme. This specific spectrum ranges dramatically from entirely asymptomatic individuals to those suffering immensely.
The diagnostic matrix carefully categorizes the structural disruption of the Pentose Phosphate Pathway into highly distinct clinical classes. Class I strictly represents the most incredibly severe manifestation, characterized clinically by very low or completely absent enzymatic activity. Unfortunate patients in this specific category experience severe chronic non-spherocytic hemolytic anemia, which tragically and relentlessly occurs even in the complete and total absence of any external oxidative stress. Class II involves dangerously low enzyme activity, frequently resulting in massive acute hemolytic crises triggered abruptly by specific oxidants.
Interestingly, mild disruptions to the Pentose Phosphate Pathway, precisely as seen in Class III patients, only cause problematic hemolysis during periods of severe external oxidative stress, while Class IV and V genetic variants safely retain normal or even slightly increased baseline activity and remain clinically silent. Fascinatingly, the slide explicitly notes a profoundly intriguing evolutionary twist: this specific enzyme deficiency confers a highly distinct evolutionary biological resistance to Plasmodium falciparum malaria, illustrating a massively complex biological trade-off.
12. Sugar Activation and the Pentose Phosphate Pathway Periphery
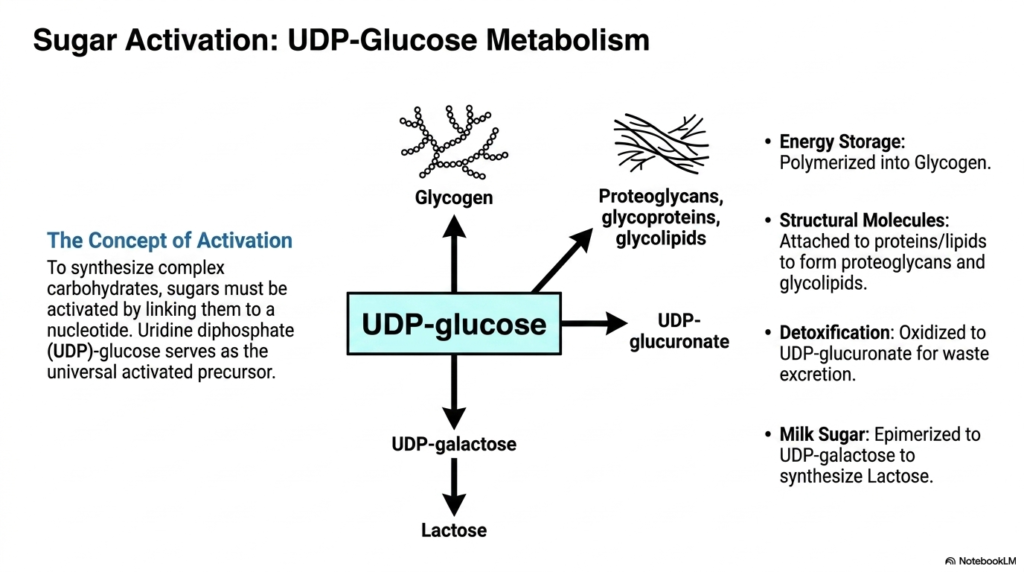
The twelfth slide effectively shifts the cellular focus to peripheral carbohydrate metabolism, which runs directly adjacent to the Pentose Phosphate Pathway, specifically highlighting UDP-glucose metabolism. The central concept initially introduced here is sugar activation. Before basic simple sugars can ever be synthesized into massive, highly complex carbohydrates, they must first be fundamentally biochemically activated by being firmly covalently linked to a nucleotide. Uridine diphosphate (UDP)-glucose serves globally as this universal activated precursor.
The detailed schematic beautifully illustrates the incredible functional versatility of UDP-glucose. For essential long-term energy storage, this highly activated molecule is systematically and efficiently polymerized into dense glycogen granules primarily within the liver and skeletal muscles. To ensure structural integrity, it is routinely attached directly to proteins and lipids, forming vast, intricate networks of proteoglycans, glycoproteins, and glycolipids. While the Pentose Phosphate Pathway heavily provides the essential reducing power for lipid synthesis, this parallel nucleotide-sugar pathway vigorously provides the necessary activated structural backbones.
Furthermore, the complex diagram specifically notes two other highly specialized physiological functions for UDP-glucose. It can be easily epimerized to UDP-galactose, which is absolutely required for the biological synthesis of lactose, commonly known as milk sugar. Additionally, UDP-glucose can be safely oxidized to form UDP-glucuronate. This specific biochemical oxidation step is vitally essential for numerous biological detoxification processes, strongly linking carbohydrate activation completely back to the liver’s broader systemic mandate to safely neutralize and efficiently excrete cellular waste.
13. Glucuronidation: Solubilizing Waste Near the Pentose Phosphate Pathway
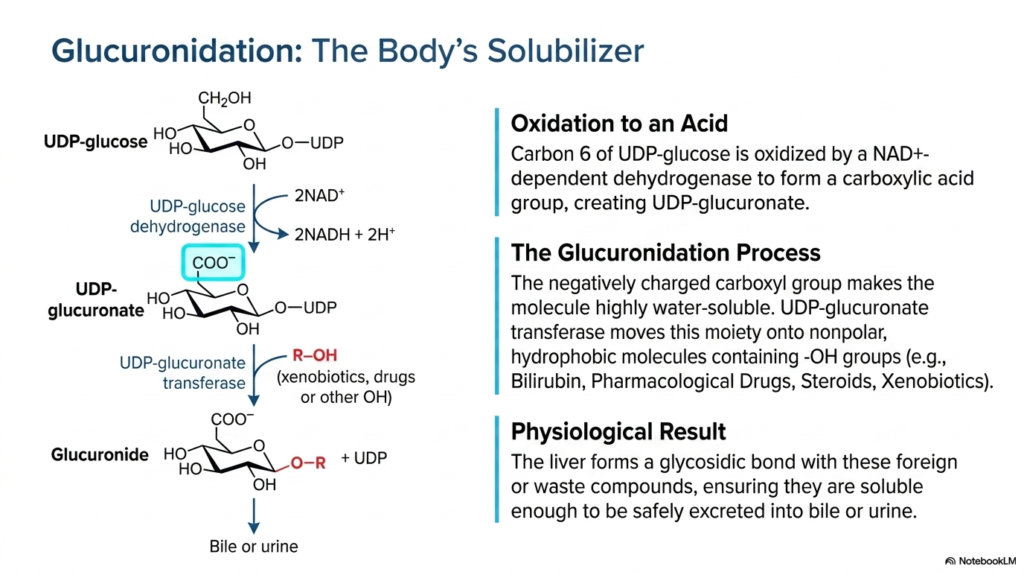
Slide thirteen thoroughly details the complex biochemical process of glucuronidation, a vital cellular detoxification mechanism that occurs robustly in the same hepatic environment as the Pentose Phosphate Pathway. The highly detailed diagram clearly shows the specific enzymatic oxidation of UDP-glucose directly into UDP-glucuronate. A highly specific NAD+-dependent dehydrogenase catalyzes this essential chemical reaction, permanently oxidizing carbon 6 to form a highly polar carboxylic acid group. This newly formed carboxylate actively carries a negative charge at standard physiological pH.
The core purpose of this robust glucuronidation process is to safely serve as the human body’s ultimate solubilizer. The crucial enzyme UDP-glucuronate transferase expertly and efficiently moves this highly negatively charged, intensely water-soluble moiety directly onto numerous nonpolar, highly hydrophobic target molecules that distinctly contain exposed hydroxyl groups. These specific target molecules often include dangerous metabolic waste products such as toxic bilirubin, numerous synthetic pharmacological drugs, naturally circulating steroid hormones, and various potentially harmful environmental xenobiotics encountered by the body.
The final physiological result of this essential chemical transfer is absolutely paramount for maintaining complete bodily homeostasis. The liver successfully and firmly forms a highly strong glycosidic bond directly between the modified glucuronide molecule and the targeted foreign or dangerous waste compound. This newly created biochemical complex is vastly more hydrophilic than the highly dangerous original toxin. This ensures that the neutralized compounds are ultimately sufficiently soluble to be safely and extremely efficiently excreted into the bile or urine, thereby protecting the organism.
14. Glycosyltransferases and Connections to the Pentose Phosphate Pathway
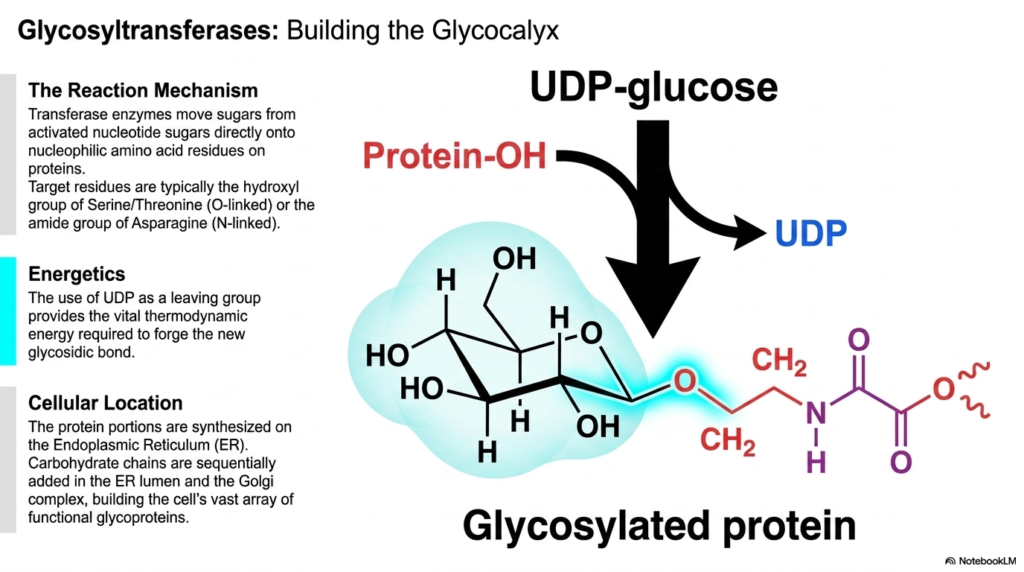
The final slide deeply explores the specific function of glycosyltransferases, the essential cellular enzymes primarily responsible for continuously building the protective glycocalyx. While the Pentose Phosphate Pathway vigorously synthesizes critical structural ribose for intracellular DNA nucleotides, various glycosyltransferases actively build highly complex structural sugars directly on the exterior surface of cellular proteins. These vital transferase enzymes operate constantly by meticulously moving individual sugar molecules directly from highly activated nucleotide sugars, precisely like UDP-glucose, onto specific, highly nucleophilic amino acid residues.
The specific biochemical reaction mechanism is beautifully and elegantly specific. Target amino acid residues typically engage the exposed hydroxyl group of serine or threonine in standard O-linked glycosylation, or the amide group of asparagine in essential N-linked glycosylation. The underlying chemical energetics driving this complex construction are highly thermodynamically favorable. The functional use of UDP as a leaving group reliably provides the thermodynamic energy required to forge the new, highly strong glycosidic bond depicted in the illustration.
Finally, the comprehensive slide accurately locates this intense biochemical activity directly within the complex cellular geography. The unglycosylated protein portions are initially synthesized directly on the rough Endoplasmic Reticulum (ER). The intricate, highly branched carbohydrate chains are then systematically and sequentially added and meticulously modified within the ER lumen and the complex Golgi apparatus. Together with the cytosolic enzymatic machinery of the Pentose Phosphate Pathway, these distinctly separate cellular compartments flawlessly orchestrate a vast, continuous array of biochemical construction.
Please read our Content Disclaimer Statement.
Check out our social media channels: