69. A Guide to Mitochondrial Transport Systems and Metabolic Shuttles
The human body relies on constant energy production, a feat largely carried out by microscopic cellular powerhouses. However, generating this energy requires a complex choreography of molecules moving in and out of highly guarded cellular compartments. The core purpose of this slide deck is to illuminate these intricate mechanisms. By examining specific molecular carriers and shuttles, students will understand how essential metabolites bypass seemingly impenetrable cellular barriers.
Slide 1: Introduction to Mitochondrial Transport Systems
The study of cellular energy production must begin with an examination of Mitochondrial Transport Systems. These systems represent the fundamental infrastructure that allows cells to maintain metabolic homeostasis. The inner cellular compartments do not exist in isolation; they must constantly communicate with the surrounding cytoplasm. This communication happens through highly regulated biochemical exchange and shuttling mechanisms.
Without efficient Mitochondrial Transport Systems, the enzymes housed within the matrix would be starved of their essential substrates. Furthermore, the vital energy currency generated within cannot reach the rest of the cell without these specialized pathways. Understanding Mitochondrial Transport Systems provides the foundation for comprehending both normal physiology and various metabolic diseases. The intricate dance of molecules across these boundaries dictates the organism’s overall metabolic rate.
Medical students and biochemistry scholars must recognize that these barriers are not mere static walls. They are dynamic, selectively permeable interfaces teeming with specialized protein complexes. The mechanisms of biochemical exchange ensure that reducing equivalents, metabolic intermediates, and energy carriers are precisely distributed. This opening overview sets the stage for a deep dive into the specific protein complexes and physical forces that make such precise molecular movement possible.
Slide 2: The Biological Barrier and Mitochondrial Transport Systems
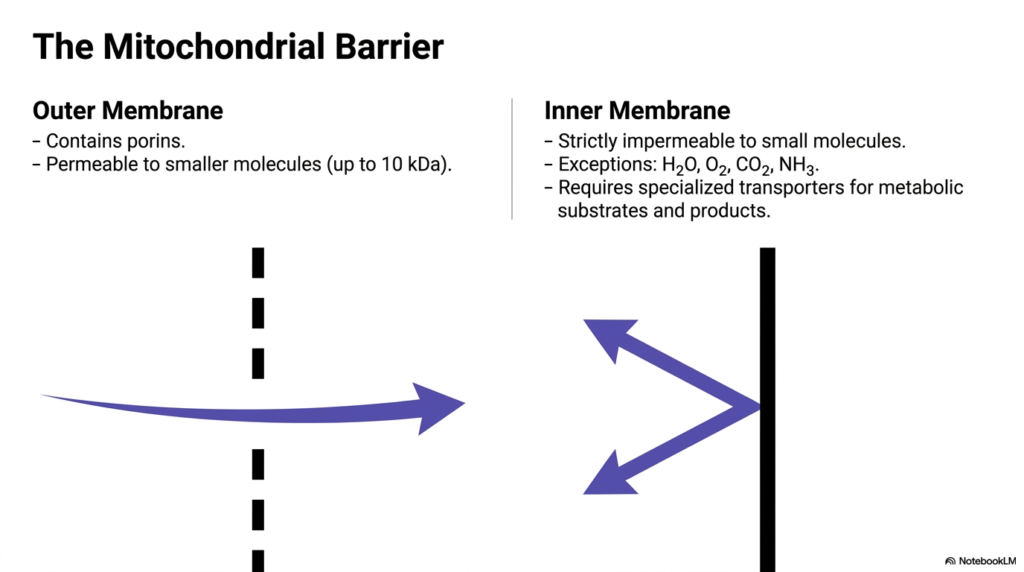
A foundational concept in bioenergetics is the distinct difference between the outer and inner boundaries of this organelle, a contrast that necessitates complex Mitochondrial Transport Systems. The outer membrane acts as a relatively porous sieve. It is populated by specialized channel proteins known as porins. These porins allow the free passage of smaller molecules, typically those weighing up to 10 kilodaltons, including many ions and basic metabolites.
Conversely, the inner membrane presents a formidable, strictly impermeable barrier to most small molecules and ions. There are very few exceptions to this rule; only uncharged, simple molecules like water, oxygen, carbon dioxide, and ammonia can freely diffuse across it. Because of this strict impermeability, the organelle relies heavily on dedicated Mitochondrial Transport Systems to transport metabolic substrates and products. This selective permeability is not a flaw but a crucial evolutionary adaptation required to maintain vital electrochemical gradients.
The contrast between these two membranes highlights the importance of specific carrier proteins. If the inner membrane were as permeable as the outer one, the gradients required for ATP synthesis would instantly dissipate. Therefore, exploring Mitochondrial Transport Systems is essential to understanding how the cell carefully orchestrates metabolite transport without compromising its energy-generating potential. The inner membrane’s strict control mechanisms ensure that only the correct molecules enter or exit at precisely the right moments.
Slide 3: Three Primary Modes within Mitochondrial Transport Systems
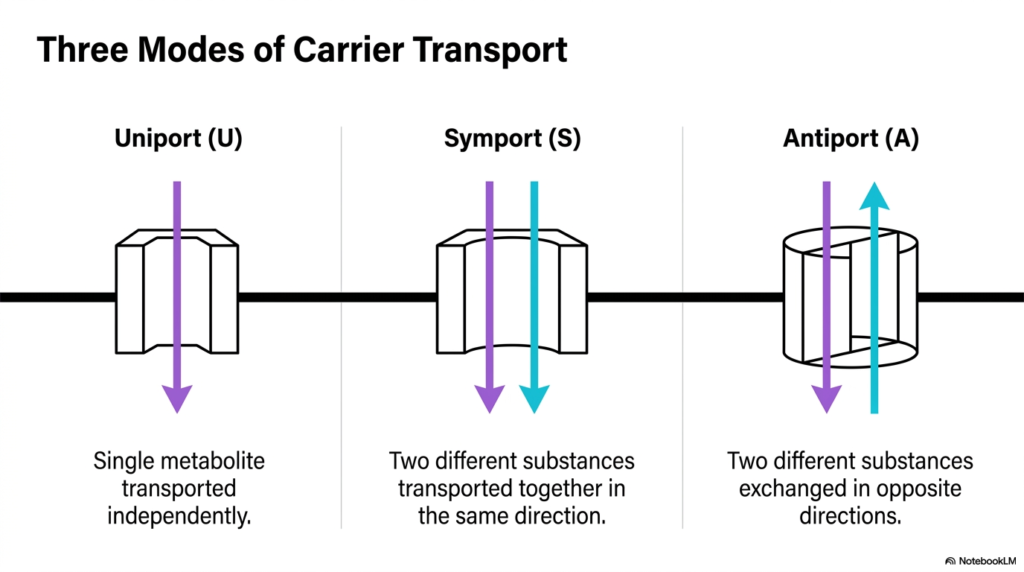
To grasp how molecules navigate these strict barriers, one must categorize the primary mechanisms utilized by Mitochondrial Transport Systems. The transport of metabolites generally falls into three distinct modes: uniport, symport, and antiport. Each mode describes a specific directional flow and the number of substances transported by carrier proteins embedded in the inner membrane.
The uniport mechanism is the simplest form found within Mitochondrial Transport Systems. In a uniport system, a single type of metabolite is transported independently across the membrane, driven primarily by concentration or electrical gradients. There is no coupling to the movement of a secondary molecule. This straightforward transport enables the targeted import or export of specific ions or molecules in response to the cellular environment.
In contrast, symport and antiport systems involve coupled movement. A symport mechanism transports two different substances together in the exact same direction across the membrane. An antiport mechanism, widely used in many Mitochondrial Transport Systems, exchanges two different substances by moving them in opposite directions. This coupled exchange is highly efficient, often using the favorable gradient of one molecule to drive the unfavorable transport of another, thereby maintaining strict metabolic balance across the inner membrane.
Slide 4: Energetics Powering Mitochondrial Transport Systems
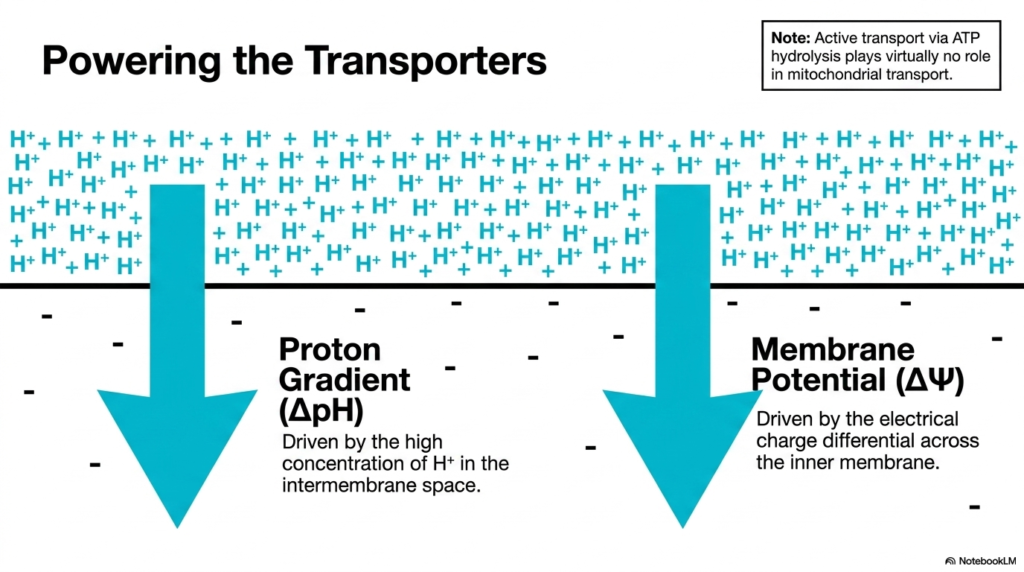
Unlike many transport mechanisms in the general cellular membrane, Mitochondrial Transport Systems rely on a unique energy source. Active transport driven directly by ATP hydrolysis plays virtually no role here. Instead, these carriers are powered by the immense electrochemical forces generated during oxidative phosphorylation. The primary driving force is the proton gradient, often denoted as delta pH.
This proton gradient is created by the electron transport chain’s constant pumping of hydrogen ions into the intermembrane space. The resulting high concentration of protons outside the matrix creates a powerful chemical drive for protons to return inward. Many Mitochondrial Transport Systems harness this exact inward flow. By coupling the transport of specific metabolites to proton movement down their concentration gradient, the organelle efficiently imports necessary substrates without expending valuable ATP.
Additionally, proton movement generates a strong membrane potential, denoted as delta psi. The intermembrane space becomes positively charged relative to the negatively charged matrix. This electrical charge differential strongly influences the movement of charged ions. Consequently, Mitochondrial Transport Systems frequently utilize this membrane potential to drive the import of positively charged molecules or the export of negatively charged ones, ensuring continuous metabolic flux.
Slide 5: Pyruvate and Phosphate Import via Mitochondrial Transport Systems
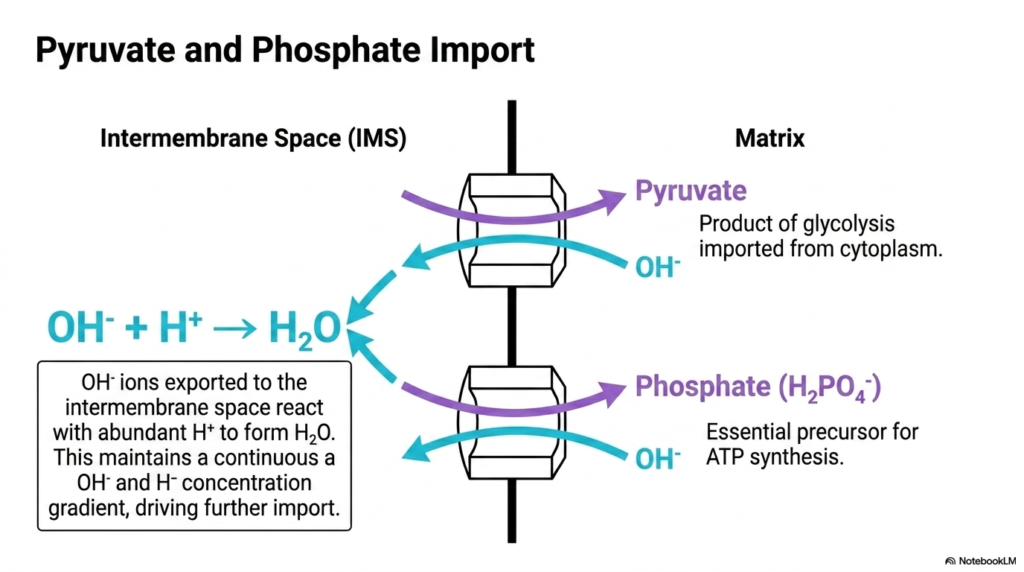
The continuous production of ATP requires a steady supply of specific precursors, which are carefully imported by Mitochondrial Transport Systems. Pyruvate, the end product of cytoplasmic glycolysis, must enter the matrix to fuel the citric acid cycle. Similarly, inorganic phosphate is an absolute requirement for the synthesis of new ATP molecules. The inner membrane manages the import of both these critical substrates through specific antiport mechanisms.
These specific Mitochondrial Transport Systems operate by coupling the inward movement of pyruvate and phosphate with the outward movement of hydroxyl ions. As pyruvate and phosphate enter the negatively charged matrix, hydroxyl ions are exported into the intermembrane space. This mechanism is highly ingenious because it leverages the existing proton gradient established by the electron transport chain.
Once the hydroxyl ions reach the intermembrane space, they immediately react with the abundant hydrogen ions present there to form water. This rapid neutralization prevents the accumulation of exported hydroxyl ions, maintaining a continuous concentration gradient that drives further import. Thus, these Mitochondrial Transport Systems elegantly solve the problem of importing essential, negatively charged substrates against a seemingly unfavorable electrical gradient by utilizing the organelle’s natural acidic exterior.
Slide 6: The Adenine Nucleotide Translocase and Mitochondrial Transport Systems
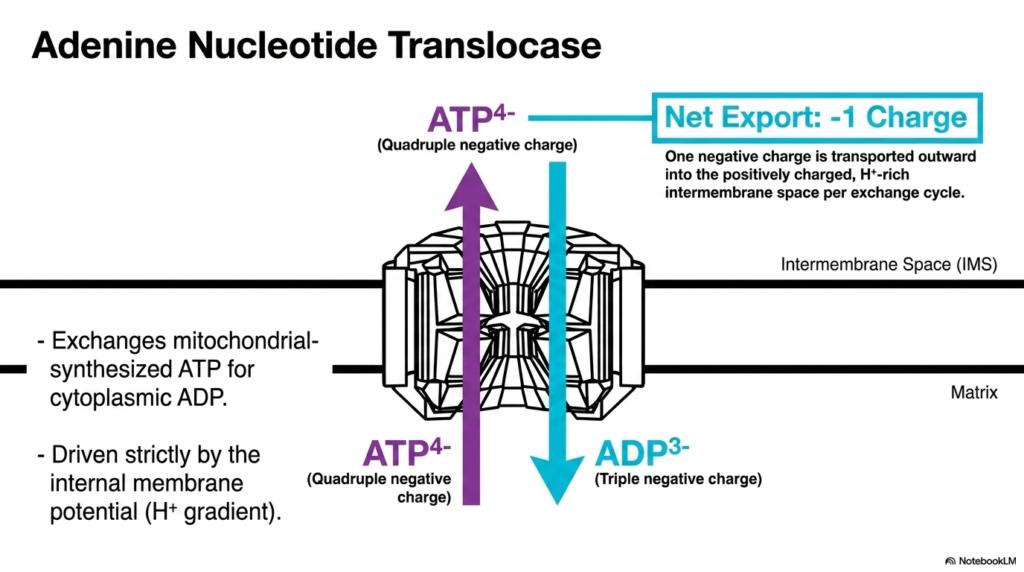
Perhaps one of the most vital components of all Mitochondrial Transport Systems is the adenine nucleotide translocase. This specific carrier protein is responsible for exporting the newly synthesized energy currency, ATP, out of the matrix while simultaneously importing spent ADP from the cytoplasm. This constant exchange is an absolute necessity for cellular survival, as the cytoplasm requires continuous energy while the matrix requires continuous substrate for phosphorylation.
The operation of this complex relies heavily on the electrical properties governed by Mitochondrial Transport Systems. ATP carries a quadruple-negative charge, whereas ADP carries a triple-negative charge. Therefore, every single exchange cycle results in the net export of one negative charge from the matrix into the intermembrane space. This outward movement of a negative charge is strongly favored by the existing electrical environment.
Because the intermembrane space is positively charged due to the high concentration of protons, the extrusion of the more negatively charged ATP molecule is electrochemically advantageous. This means the adenine nucleotide translocase is driven strictly by the internal membrane potential rather than requiring additional energy input. This elegantly highlights how Mitochondrial Transport Systems optimize energy distribution by utilizing the fundamental biophysical gradients already established by cellular respiration.
Slide 7: Malate and Citrate Exchange within Mitochondrial Transport Systems
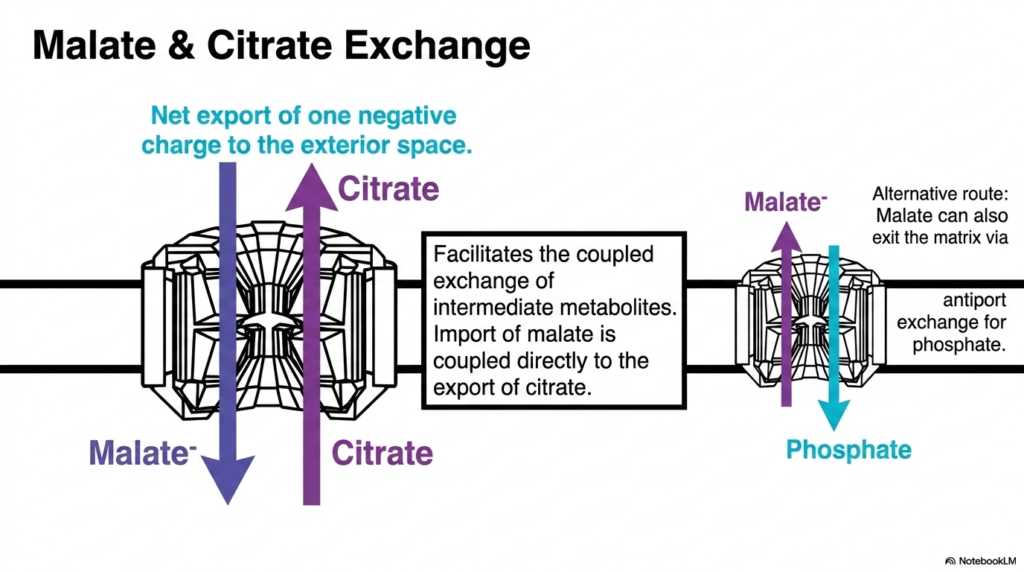
Beyond energy carriers, Mitochondrial Transport Systems must also manage the complex distribution of larger metabolic intermediates. The exchange of malate and citrate is a prime example of this required coordination. The inner membrane features a specific carrier that directly couples the import of malate with the export of citrate. This precise movement allows metabolic intermediates to shift between the matrix and the cytoplasm for various biosynthetic pathways.
This particular exchange process results in the net export of one negative charge to the exterior space. Citrate carries a greater negative charge than malate, so the coordinated swap is gently assisted by the membrane’s electrical potential. Furthermore, Mitochondrial Transport Systems demonstrate remarkable redundancy and flexibility. If malate needs to exit rather than enter, an alternative route exists.
Malate can also exit the matrix via a different antiport mechanism, where it is exchanged for incoming inorganic phosphate. The existence of these interconnected, multifunctional Mitochondrial Transport Systems ensures that the organelle can quickly adapt to the cell’s fluctuating metabolic demands. Whether the cell is engaged in fatty acid synthesis, which requires citrate export, or in gluconeogenesis, which requires malate export, these carriers seamlessly facilitate the necessary biochemical fluxes.
Slide 8: Managing Calcium Ion Flux with Mitochondrial Transport Systems
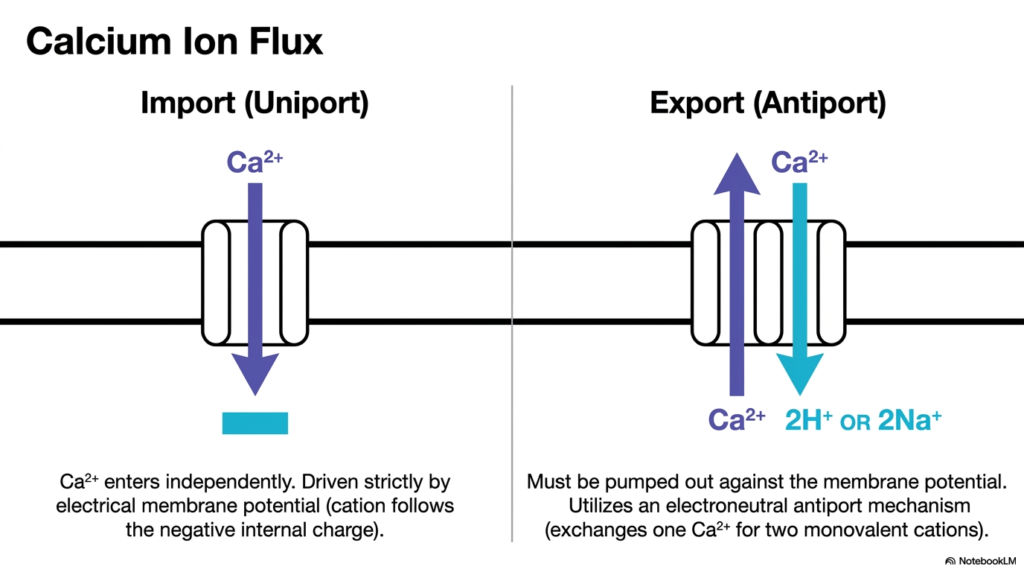
Calcium ions play a critical role in cellular signaling and metabolic regulation, making their movement a key focus of Mitochondrial Transport Systems. The organelle acts as a potent buffer for intracellular calcium, meaning it must rapidly import and export this cation as needed. Calcium is imported via a dedicated uniport mechanism. Because calcium has a strong double-positive charge, it enters the matrix independently, driven fiercely by the negative internal charge.
This electrical membrane potential strongly favors the rapid influx of calcium when cytoplasmic calcium levels rise. However, the subsequent export of calcium presents a significant thermodynamic challenge for Mitochondrial Transport Systems. Because the interior is highly negative, pushing the positively charged calcium ions back out into the intermembrane space means moving them strictly against the strong membrane potential.
To accomplish this difficult task, the organelle utilizes an electroneutral antiport mechanism. The export carrier exchanges one outgoing calcium ion for multiple incoming monovalent cations, specifically two hydrogen ions or two sodium ions. By trading charges in this exact manner, these Mitochondrial Transport Systems balance the electrical equation, allowing the organelle to precisely regulate its internal calcium stores without completely disrupting the vital membrane potential needed for other processes.
Slide 9: Resolving the NADH Problem via Mitochondrial Transport Systems
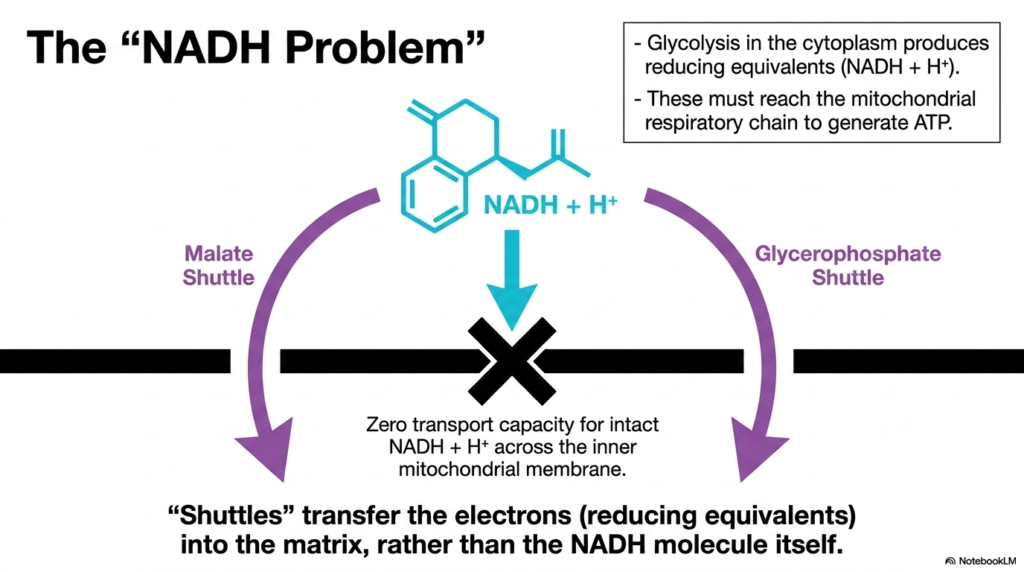
A major biochemical hurdle in cellular respiration is often referred to as the “NADH Problem,” a dilemma ultimately solved by specialized Mitochondrial Transport Systems. Glycolysis, which occurs in the cytoplasm, rapidly breaks down glucose and produces reducing equivalents in the form of NADH and a proton. For the cell to extract maximum ATP from these molecules, they must interact with the respiratory chain located inside the matrix.
However, the intact NADH molecule possesses absolutely zero transport capacity across the inner mitochondrial membrane. There are no pores or carriers capable of moving this large, complex molecule inward. This creates a bottleneck: the valuable electrons trapped in cytoplasmic NADH cannot reach their destination directly. This is where the concept of metabolic shuttles, a brilliant adaptation of Mitochondrial Transport Systems, becomes essential.
Instead of moving the physical NADH molecule itself, these distinct shuttles transfer only the electrons, or reducing equivalents, across the barrier. The electrons are handed off to intermediate molecules that can easily cross the membrane via specific carriers. Once inside, these intermediates donate the electrons to reform NADH or similar carriers within the matrix. These specialized Mitochondrial Transport Systems thus preserve the energy harvested during glycolysis, allowing efficient aerobic respiration to continue uninterrupted.
Slide 10: The Malate Shuttle and Mitochondrial Transport Systems
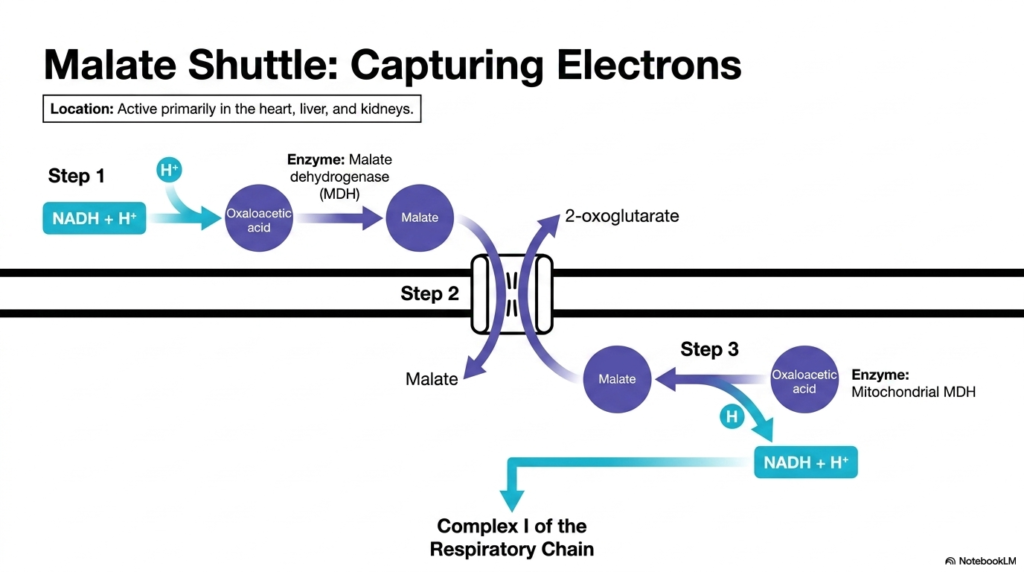
One of the primary solutions to the NADH problem is the malate shuttle, a highly active component of Mitochondrial Transport Systems in tissues with immense energy demands like the heart, liver, and kidneys. This complex multi-step cycle brilliantly bypasses the inner membrane’s impermeability to NADH. The process begins in the cytoplasm, where the enzyme malate dehydrogenase transfers the electrons from cytoplasmic NADH to a molecule called oxaloacetic acid.
This biochemical reduction transforms oxaloacetic acid into malate. Unlike NADH, malate possesses a dedicated carrier protein within the inner membrane. Through these specific Mitochondrial Transport Systems, the newly formed malate successfully crosses the barrier and enters the inner matrix. The electrons from the cytoplasm have now successfully infiltrated the heavily guarded interior of the organelle.
Once inside, step three unfolds. A mitochondrial form of malate dehydrogenase reverses the initial reaction. It strips electrons from malate, transferring them to an internal NAD+ molecule, thereby regenerating NADH in the matrix. The newly formed NADH is now perfectly positioned to donate its electrons directly to Complex I of the respiratory chain. This demonstrates how Mitochondrial Transport Systems efficiently capture and relocate external energy equivalents.
Slide 11: Regenerating the Malate Cycle in Mitochondrial Transport Systems
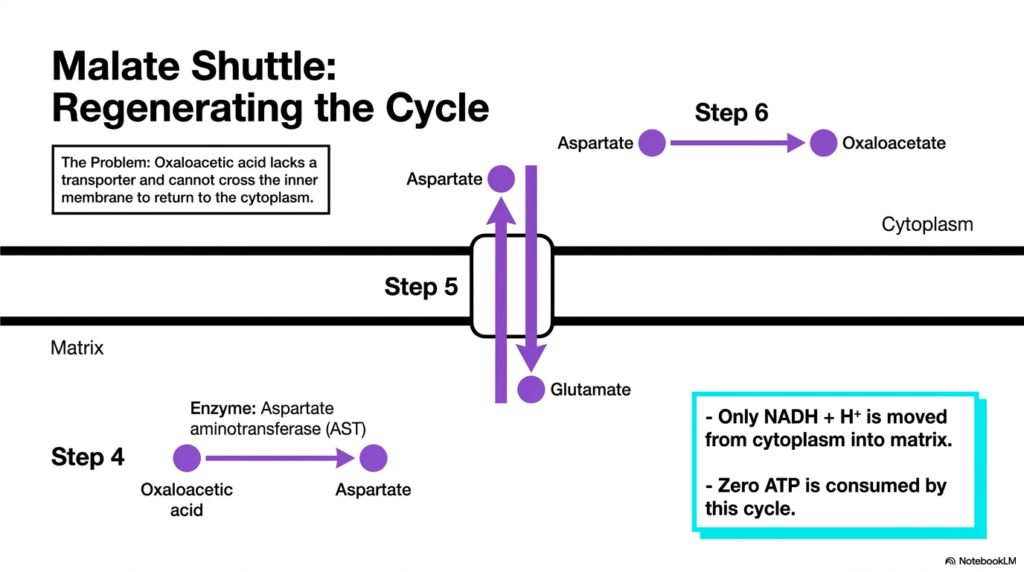
While electron capture is crucial, the malate shuttle must also continuously reset itself to remain an effective component of the mitochondrial transport system. After malate donates its electrons in the matrix, it reverts to oxaloacetic acid. However, a significant new problem arises: oxaloacetic acid entirely lacks a dedicated transporter and cannot cross the inner membrane to return to the cytoplasm. If left unresolved, the entire cycle would instantly halt.
To bypass this, the shuttle relies on a transamination reaction. An enzyme called aspartate aminotransferase converts the trapped oxaloacetic acid into aspartate. Fortunately, aspartate has a specific carrier protein within the Mitochondrial Transport Systems. Aspartate is successfully transported out of the cell into the cytoplasm in direct exchange for incoming glutamate. Once in the cytoplasm, aspartate is converted back into oxaloacetic acid, fully regenerating the cycle for another round.
This intricate biochemical loop highlights the profound efficiency of these specialized Mitochondrial Transport Systems. By moving intermediates back and forth, the shuttle successfully moves only the reducing equivalents from the cytoplasm into the matrix. More importantly, this entire cyclical exchange proceeds smoothly without consuming a single molecule of ATP, making it a highly efficient mechanism for maximizing cellular energy yield.
Slide 12: The Glycerophosphate Shuttle as a Mitochondrial Transport System
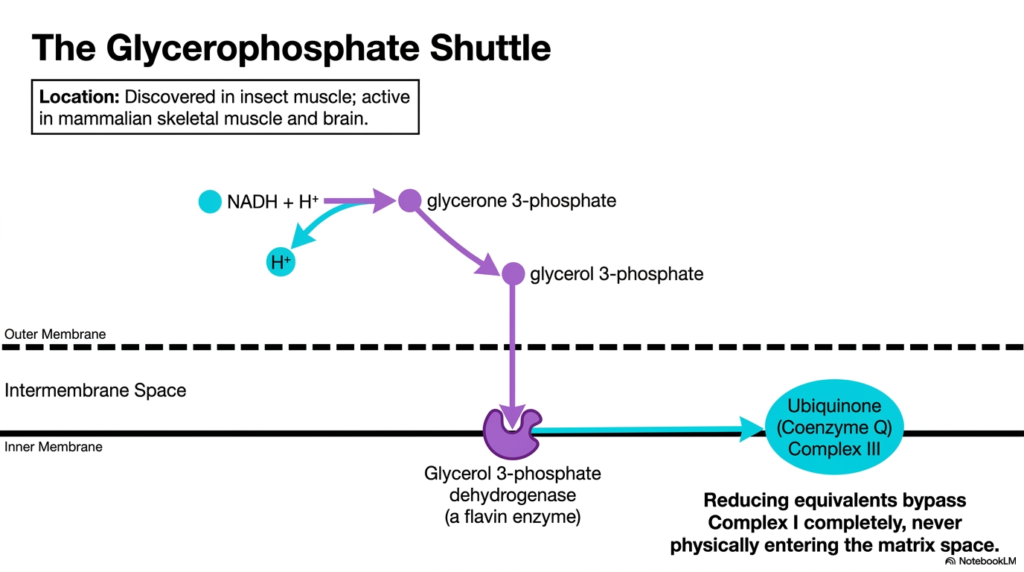
Different tissues require different metabolic strategies, which is why alternative Mitochondrial Transport Systems exist, such as the glycerophosphate shuttle. While less prominent in the liver, this specific shuttle is highly active in mammalian skeletal muscle and the brain, and was originally discovered in insect flight muscle. It offers a faster, albeit slightly less energy-dense, method for transferring cytoplasmic electrons.
In this pathway, the cytoplasmic NADH donates its electrons to glycerone 3-phosphate, converting it into glycerol 3-phosphate. Glycerol 3-phosphate easily diffuses through the porous outer membrane into the intermembrane space. The brilliance of these specific Mitochondrial Transport Systems is that the intermediate molecule never actually has to cross the inner membrane into the matrix.
Instead, an enzyme called glycerol 3-phosphate dehydrogenase is firmly embedded on the outer surface of the inner membrane. This flavin-linked enzyme directly extracts electrons from glycerol 3-phosphate and passes them straight to Ubiquinone (Coenzyme Q), which feeds into Complex III. Because these Mitochondrial Transport Systems completely bypass Complex I, the reducing equivalents enter the respiratory chain slightly further downstream, prioritizing rapid electron transfer over maximum ATP yield.
Slide 13: Diagnostic Matrix for Mitochondrial Transport Systems

To fully synthesize the concepts surrounding the NADH problem, students must compare the two primary shuttles utilized by Mitochondrial Transport Systems. A diagnostic matrix provides a clear, high-level summary of their distinct biochemical characteristics and physiological locations. The malate shuttle operates primarily in the heart, liver, and kidneys, demanding complex antiport carriers to move malate, alpha-ketoglutarate, aspartate, and glutamate.
In stark contrast, the glycerophosphate shuttle dominates in skeletal muscle and brain tissue. It completely sidesteps the need for inner membrane transport carriers, relying solely on outer membrane porins and an enzyme fixed to the intermembrane surface. This distinct structural difference between the two Mitochondrial Transport Systems leads directly to a crucial functional difference in how the electrons join the respiratory cascade.
The malate shuttle delivers electrons directly to Complex I, maximizing the subsequent proton pumping and ATP generation. The glycerophosphate shuttle, prioritizing speed, drops its electrons off at Ubiquinone, yielding slightly less ATP per cytoplasmic NADH. Understanding these tailored Mitochondrial Transport Systems allows clinicians to grasp how different organs optimize cellular respiration in response to their unique, tissue-specific energy demands.
Please read our Content Disclaimer Statement.
Check out our social media channels: