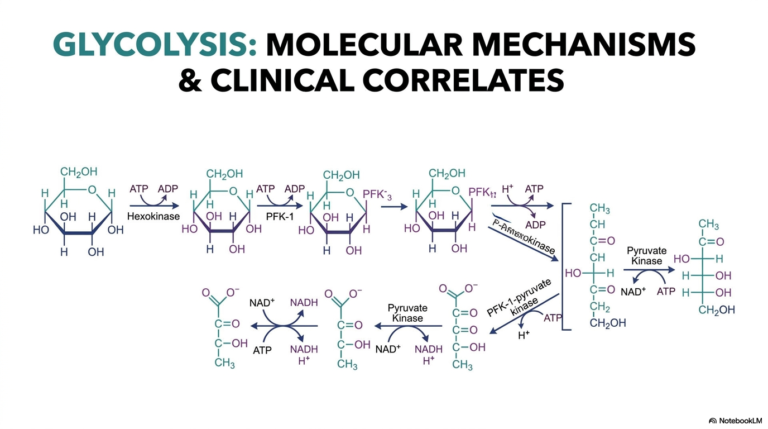
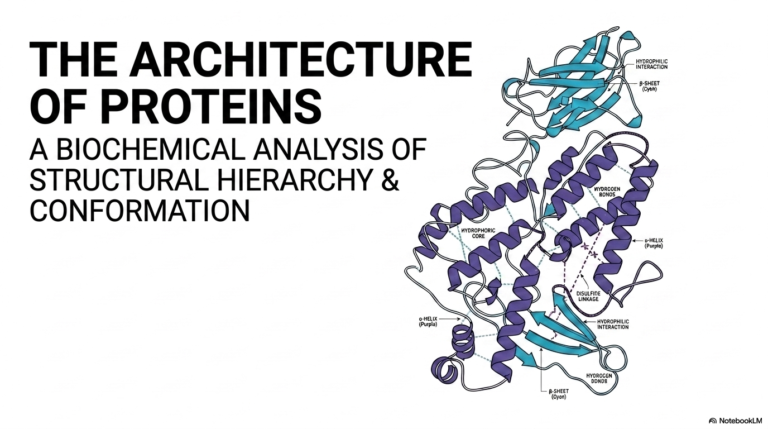
50. Understanding Protein Metabolism: Nitrogen Balance and Turnover
One might wonder how dietary nutrients transform into complex muscle tissue, or why the human organism constantly rebuilds its own architecture. The answer lies in the dynamic, non-stop engine of the cellular environment. This comprehensive slide deck breaks down the complex biochemical structures and processes underlying these remarkable transformations. It serves as a clear, energetic guide, specifically designed to help college and medical students master the fundamental principles of cellular biochemistry.
1. The Foundation of Protein Metabolism and Biosynthesis
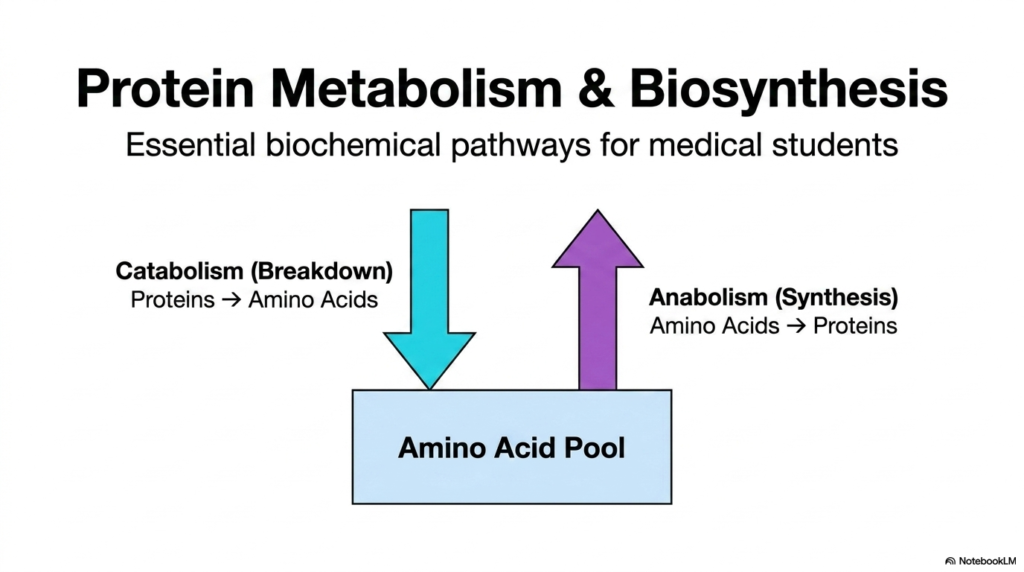
To understand the fundamental principles of life, one must look at how cells constantly construct and dismantle their molecular machinery. Protein metabolism stands as the central pillar of these cellular processes, representing a continuous, dynamic flux rather than a static state. The human body does not simply build tissues and leave them indefinitely. Instead, it relies on a delicate, endless cycle of construction and demolition.
This introductory visual outlines the most basic framework of protein metabolism: the dual pathways of catabolism and anabolism. Catabolism is the breakdown phase in which complex, fully formed proteins are dismantled into their constituent parts. These constituent parts are amino acids. Catabolism functions much like taking apart a massive castle, block by block, so the individual pieces can be rapidly repurposed for something else entirely.
On the other side of the equation lies anabolism, the powerful synthetic pathway. During anabolism, cells take those individual blocks and link them together to form entirely new structures necessary for biological survival. This specific half of protein metabolism requires massive energy input but is absolutely essential for physical growth, cellular repair, and everyday physiological function.
The central reservoir connecting these two opposing pathways is the amino acid pool. As proteins are broken down, they release their amino acids directly into this pool. Conversely, when the cell needs to synthesize new structures, it withdraws the required chemical building blocks from this exact same pool. Maintaining the precise volume of this pool is the ultimate biological goal of these essential biochemical pathways.
Understanding this constant push and pull sets the stage for mastering biochemistry. When one pathway significantly outpaces the other, the entire organism is profoundly affected. The careful regulation of this amino acid reservoir ensures that life continues uninterrupted, highlighting why studying this intricate cycle is so crucial for future medical professionals.
2. Assessing Protein Metabolism Through Nitrogen Balance
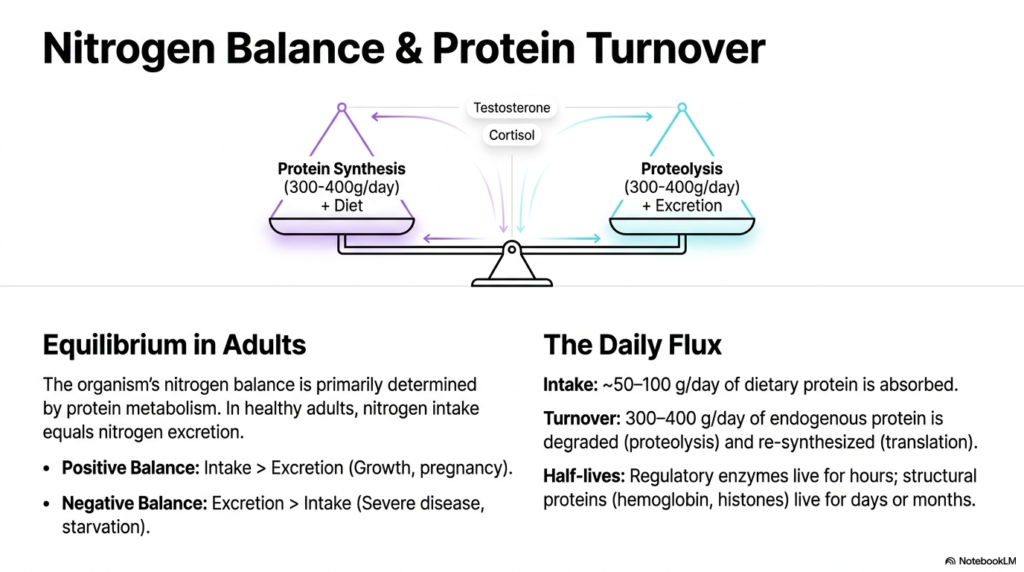
A major concept in understanding physiological health is nitrogen balance, which is intrinsically tied to protein metabolism. Since amino acids are unique among macronutrients for their dense nitrogen content, tracking nitrogen provides a direct, highly accurate window into the body’s internal accounting system. In perfectly healthy adult organisms, this system remains in strict equilibrium, meaning the amount of nitrogen consumed matches the amount excreted.
The rate at which the body cycles through this process is absolutely staggering. Every single day, a massive 300 to 400 grams of endogenous protein are degraded through cellular proteolysis and subsequently re-synthesized from scratch. This incredibly rapid daily turnover is a cornerstone of protein metabolism, allowing the body to constantly refresh its enzymatic machinery. Hormones play a massive role; testosterone promotes synthesis, while cortisol triggers degradation.
When the scales tip rapidly, a patient enters either a positive or a negative nitrogen balance. A positive balance occurs when intake vastly exceeds excretion, a state commonly observed during pregnancy, childhood growth, or intense athletic training. The body is effectively hoarding nitrogen to build new tissues, representing a highly hyperactive anabolic phase of the overall biological cycle.
Conversely, a negative balance is a glaring clinical red flag indicating that excretion severely outpaces dietary intake. This dangerous scenario typically unfolds during starvation, severe disease states, or major physical trauma. The body begins cannibalizing its own tissues to survive, aggressively tearing down muscle mass to free up essential amino acids for critical survival functions.
Recognizing these clinical shifts is exactly why grasping the nuances of protein metabolism is vital for any future healthcare provider. The lifecycle of these complex molecules varies wildly depending on their final physiological function. Regulatory enzymes might exist for mere hours, whereas robust structural components like hemoglobin can persist safely for months.
3. Navigating Protein Metabolism and Amino Acid Pool Dynamics
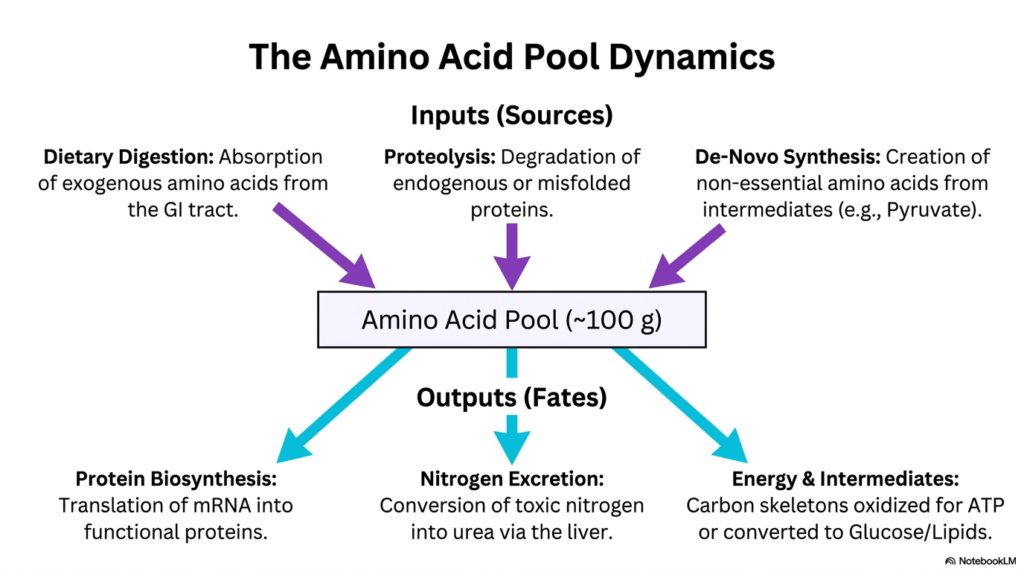
The human body manages its molecular resources with incredible efficiency, a biochemical principle perfectly illustrated by the dynamic behavior of the amino acid pool. This vital pool, though only containing about 100 grams of free amino acids at any given time, is the bustling central hub of protein metabolism. It operates much like a busy transit station, with constant arrivals and rapid departures dictating nutritional status.
To maintain this highly critical reservoir, the body relies heavily on three primary inputs. First, dietary digestion provides a massive influx of exogenous amino acids absorbed directly from the gastrointestinal tract following a meal. Second, intracellular proteolysis contributes immensely by aggressively degrading endogenous or obsolete cellular machinery. Finally, de novo synthesis generates non-essential amino acids from common metabolic intermediates to ensure the pool never runs dry.
Just as there are multiple ways to fill the pool, the functional outputs of protein metabolism are equally diverse and critical to survival. The most vital fate for these charged molecules is protein biosynthesis, where the cell translates genetic mRNA instructions into brand new functional machinery. This biological factory draws heavily from the reservoir to build everything from rigid structural fibers to highly complex metabolic enzymes.
However, amino acids absolutely cannot be safely stored for the long term. When the pool overflows, the body must safely and rapidly dispose of the massive excess. This leads to the second major output: nitrogen excretion. The liver aggressively strips the highly toxic nitrogen components from the molecules and forcefully converts them into safe, water-soluble urea for renal elimination, preventing lethal ammonia toxicity.
Finally, the leftover carbon skeletons provide a deeply vital source of metabolic fuel. They can be quickly shunted into central energy production, oxidized directly for massive amounts of ATP, or converted into glucose and lipids. By perfectly balancing these complex inputs and outputs, protein metabolism guarantees that all cells always have exactly what they need to thrive.
4. Dietary Constraints in Protein Metabolism and Synthesis
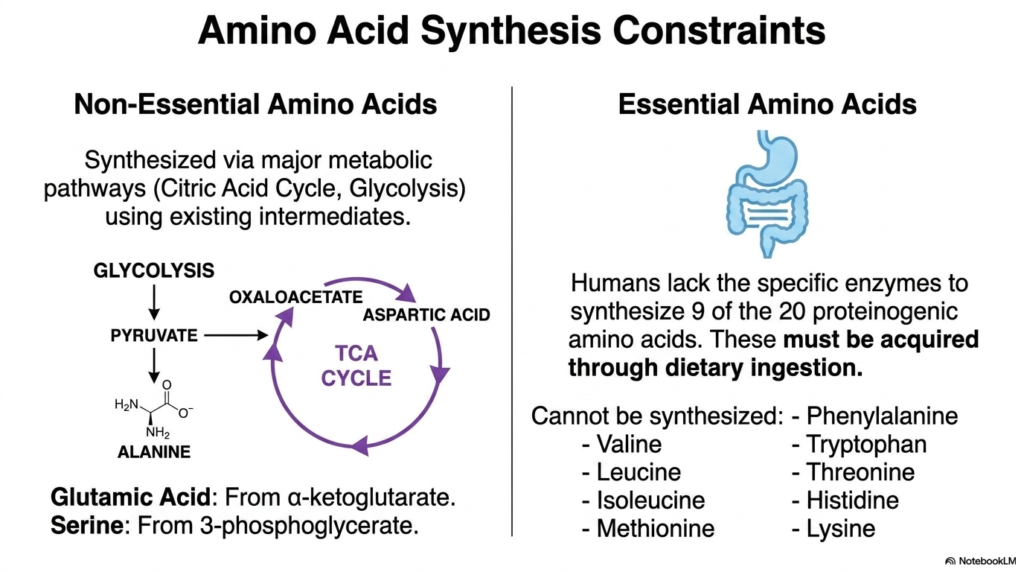
Despite the body’s truly remarkable biochemical capabilities, there are hard, unyielding physiological limits to what it can successfully build from scratch. These strict limitations create a permanent division between essential and nonessential amino acids, a categorization that absolutely dictates the nutritional rules governing protein metabolism. Understanding this stark divide is paramount to recognizing exactly how human diets profoundly shape cellular health throughout the lifespan.
Non-essential amino acids beautifully showcase the highly impressive resourcefulness of human biochemistry. The body can seamlessly and rapidly synthesize these distinct molecules using existing carbon intermediates drawn directly from major metabolic pathways such as glycolysis and the Citric Acid Cycle. For example, the incredibly common molecule pyruvate can be easily converted into alanine, while alpha-ketoglutarate serves as the direct precursor for glutamic acid.
This intrinsic molecular manufacturing capability ensures that even during brief periods of fasting, cells can readily maintain basic structural integrity. By intelligently pulling intermediate molecules straight out of the energy cycle, the cellular machinery perfectly synchronizes raw energy production with the demanding anabolic needs of protein metabolism. The cell simply reshuffles its massive molecular deck, turning raw fuel into dense building blocks.
However, the biological story changes drastically and dangerously for essential amino acids. Millions of years of evolution have completely stripped the human genome of the specific enzymatic pathways required to synthesize nine of the twenty proteinogenic amino acids. Critical molecules such as Phenylalanine, Valine, Leucine, and Tryptophan cannot be synthesized internally. The entire organism is permanently dependent on external sources to acquire them.
Because these nine highly crucial components must be acquired entirely through dietary ingestion, a severe nutritional deficiency can rapidly grind protein metabolism to an absolute, catastrophic halt. If even one essential amino acid is completely missing from the cellular pool, translation stops instantly, and the cell can no longer build critical biological structures. This highlights the vulnerability of internal metabolic pathways.
5. Driving Protein Metabolism: The Transcription of DNA to mRNA
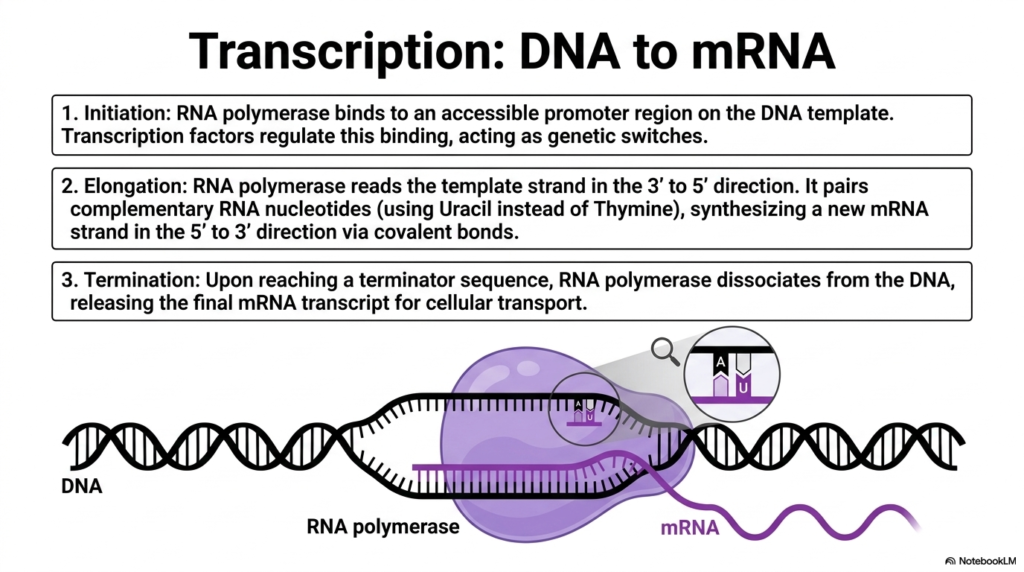
Before any physical biological structure can be actively built, the master blueprints must be carefully and accurately copied from the central archive. In the sprawling realm of protein metabolism, this initial, highly guarded phase is known as transcription. It is the tightly regulated process by which the complex genetic code locked away in double-stranded DNA is transcribed into a highly mobile, single-stranded messenger RNA molecule.
The biological process begins with a highly critical initiation phase. The massive enzyme RNA polymerase locates and tightly binds to a specific, easily accessible promoter region on the master DNA template. This crucial step is strictly governed by specialized transcription factors acting as intricate genetic switches. These complex factors directly determine when and where the initial steps of protein metabolism occur in response to immediate needs.
Once securely attached to the strand, the machinery rapidly shifts into the elongation phase. RNA polymerase forcefully untwists the rigid DNA helix and seamlessly slides along the template strand, accurately reading the genetic sequence in a strict three-prime-to-five-prime direction. As it moves, it rapidly incorporates complementary RNA nucleotides from the surrounding environment, forming very strong covalent bonds to synthesize the new mRNA strand.
A defining biochemical feature of this specific phase is the absolute substitution of bases. Instead of using the traditional Thymine to pair perfectly with Adenine, the RNA polymerase creatively uses Uracil. This newly forming mRNA chain physically elongates in a five-prime-to-three-prime direction, rapidly creating a perfect, chemically distinct mirror image of the original genetic instruction. This accuracy is utterly vital for preventing catastrophic biological errors.
Finally, the incredible biological journey comes to an end. Upon encountering a specific terminator sequence embedded in the DNA, the RNA polymerase abruptly halts its activity and dissociates from the template. The newly minted mRNA transcript is fully released, ready to be immediately transported out of the nucleus to confidently face the next stage of protein metabolism.
6. Executing Protein Metabolism: Translation and Polypeptide Synthesis
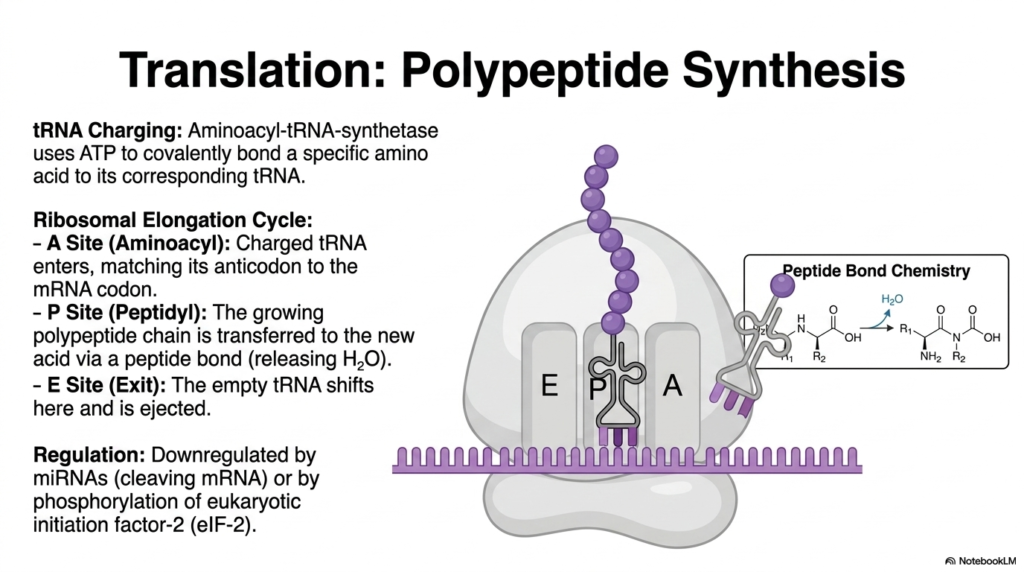
With messenger RNA successfully transported into the dense cellular cytoplasm, the most energy-intensive phase of protein metabolism begins. Translation is the truly awe-inspiring biological process in which simple nucleotide sequences are rapidly decoded and physically transformed into massive polypeptide chains. This monumental task requires an incredibly intricate assembly line composed of massive ribosomes, mRNA, and highly specialized transfer RNA molecules.
The very first critical biochemical step is tRNA charging. An incredibly specialized enzyme called an aminoacyl-tRNA synthetase aggressively consumes cellular ATP to covalently attach a specific free-floating amino acid to its corresponding tRNA molecule. This massive energy investment ensures the specific tRNA is perfectly primed to deliver the exact correct building block to the factory floor, an absolutely necessary preparation for the high-fidelity demands of protein metabolism.
Once the heavily charged tRNAs are perfectly ready, the rapid ribosomal elongation cycle aggressively takes over. The massive ribosome functions as a sprawling biological factory, equipped with three distinct chambers: the A, P, and E sites. A fully charged tRNA rapidly enters the A Site, carefully matching its unique anticodon to the codon directly complementary to it on the mRNA strand.
The deeply complex core chemistry occurs just as the growing polypeptide chain, safely anchored in the P Site, is brutally transferred directly to the newly arrived amino acid in the A Site. The massive ribosome physically catalyzes a highly powerful peptide bond, safely releasing a small water molecule. This specific chemical linkage is the literal, physical manifestation of building structures during protein metabolism.
Following successful bond formation, the empty tRNA physically shifts straight into the E Site and is violently ejected back into the cytoplasm. The massive ribosome forcefully ratchets forward, and the cycle repeats. To definitively prevent out-of-control cellular growth, this entire factory can be strongly downregulated by harsh mechanisms such as miRNA cleavage or rapid phosphorylation of eukaryotic initiation factor 2.
7. Enhancing Protein Metabolism Through Post-Translational Modifications
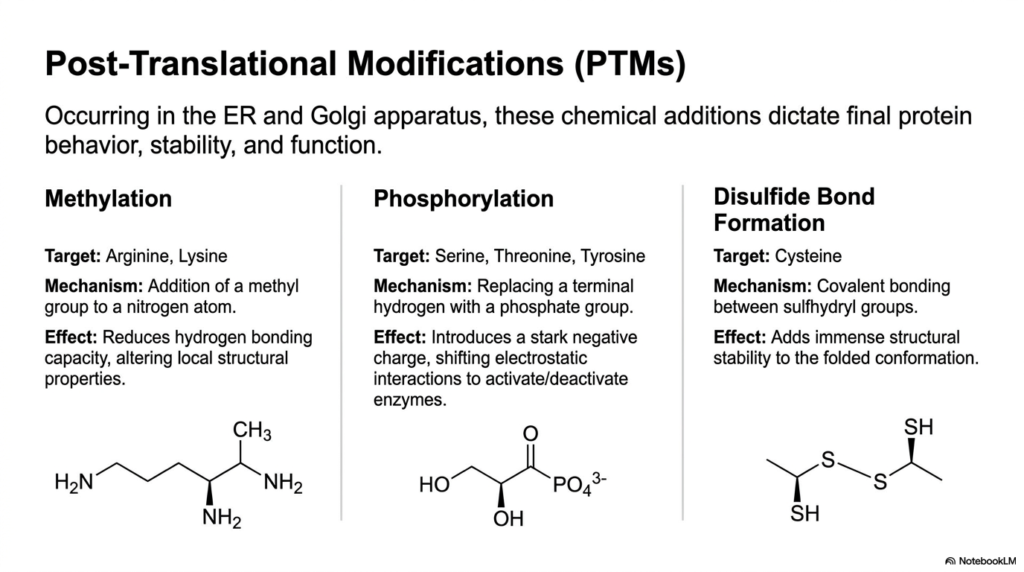
A newly synthesized polypeptide chain is rarely ever ready to successfully perform its biological duties immediately after leaving the ribosome. It must urgently undergo extremely extensive fine-tuning, a highly specialized phase of protein metabolism specifically known as post-translational modification. Occurring deep within the crowded Endoplasmic Reticulum and the sprawling Golgi apparatus, these highly specific chemical additions fundamentally dictate the final cellular behavior and ultimate function.
One incredibly common biological alteration is methylation, which heavily and specifically targets the amino acids Arginine and Lysine. In this unique mechanism, specialized enzymes forcefully attach a small, highly dense methyl group directly to a specific nitrogen atom located right on the amino acid side chain. This highly subtle chemical shift drastically and immediately reduces the molecule’s overall hydrogen-bonding capacity, thereby subtly altering its local structural properties.
Phosphorylation is undoubtedly the most incredibly dynamic and widespread biological modification found within the sprawling pathways of protein metabolism. Targeting Serine, Threonine, or Tyrosine, this aggressive mechanism involves forcefully and brutally replacing a terminal hydrogen atom with a massively bulky, highly charged phosphate group. This violent addition instantly imparts a massive, negative electrostatic charge to the molecule’s surface, triggering rapid conformational shifts.
For complex proteins destined for incredibly harsh, unprotected extracellular environments, massive physical durability is absolutely paramount. This vital structural durability is beautifully achieved through the formation of complex disulfide bonds, a unique, deeply structural modification that specifically targets reactive Cysteine residues. Highly specialized enzymes forcefully catalyze the rapid formation of a highly rigid, entirely permanent covalent bond directly between two adjacent sulfhydryl groups.
These highly robust disulfide cross-links effectively act exactly like massive biological spot-welds, aggressively adding immense structural stability directly to the final properly folded conformation. Without absolutely executing these highly crucial modifications, the vast majority of synthesized biological chains would remain entirely biologically inert. Fully understanding this complex maturation process gives medical students a complete picture of how protein metabolism creates highly specialized cellular machinery.
8. The Thermodynamics of Protein Metabolism and Folding
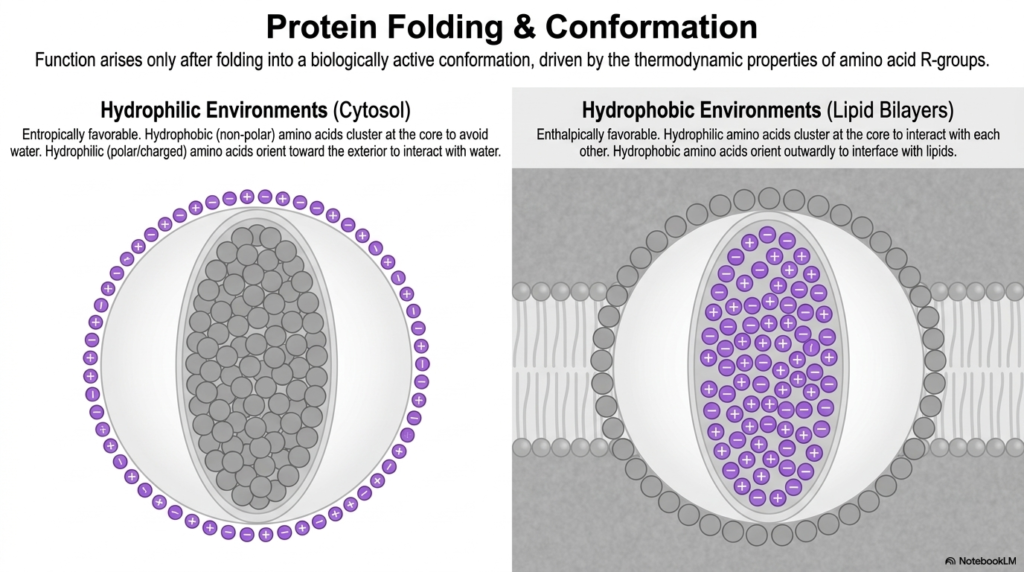
The highly linear, simple sequence of chained amino acids is totally chemically meaningless until it fully adopts a highly specific, densely organized three-dimensional shape. This incredible biological phenomenon is driven entirely by the inherent thermodynamic properties of specific amino acid R-groups that interact fiercely with their immediate biological surroundings. This intensely physical folding process is the absolute final determining step of anabolic protein metabolism.
The specific environment in which a developing protein finds itself strongly dictates its final, fully functional physical conformation. In the highly aqueous, water-filled environment of the main cellular cytosol, rapid folding is primarily driven by highly entropically favorable chemical interactions. The massive polypeptide chain rapidly and violently twists and contorts to safely hide its extremely hydrophobic, non-polar amino acids.
Simultaneously, the highly reactive, hydrophilic amino acids, which are highly polar or carry strong electrostatic charges, readily orient themselves entirely outward to interact freely and strongly with the highly aqueous cellular environment. This highly classic, densely packed globular structure fully represents the most thermodynamically stable state for basic, water-soluble enzymes. A rigorous understanding of these interactions is essential for mastering protein metabolism.
However, absolutely not all complex cellular machinery physically exists in a simple watery environment. Massive transmembrane proteins, incredibly tightly embedded entirely within dense cellular lipid bilayers, face a completely inverted thermodynamic reality. In these intensely hydrophobic, heavily fatty cellular environments, the standard folding rules are forcefully flipped upside down to actively favor a completely different set of enthalpically driven chemical interactions.
Within the highly dense cellular membrane, it is the highly hydrophilic amino acids that cluster tightly together deep in the core, violently shielding themselves from the surrounding fats. Meanwhile, the incredibly hydrophobic amino acids confidently orient outwardly. This deeply elegant environmental adaptation clearly demonstrates that complex protein architecture in protein metabolism is always strictly dictated by the fundamental laws of thermodynamics.
9. Managing Waste: Intracellular Proteolysis in Protein Metabolism
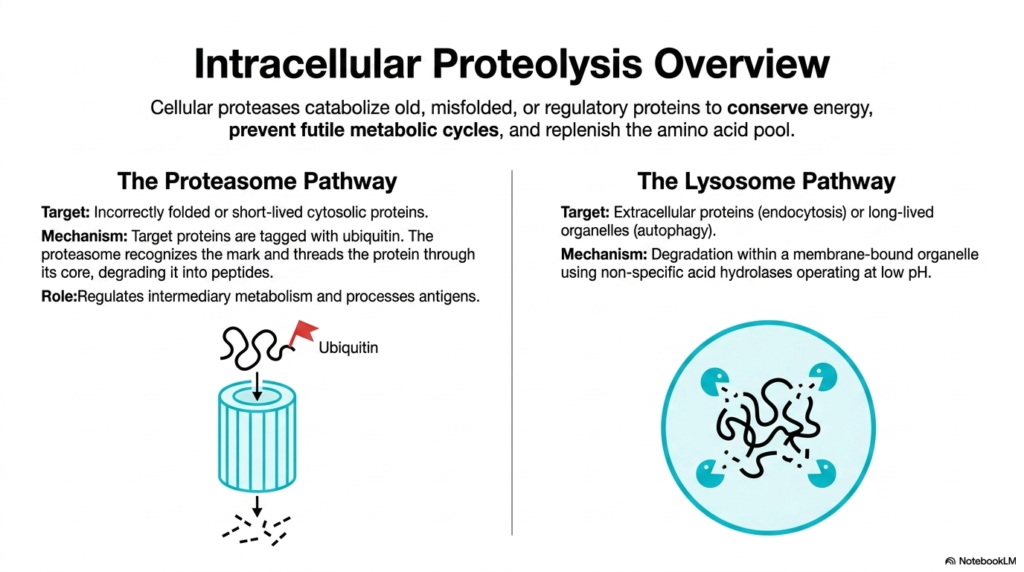
Building massive new biological structures is only half the biochemical battle; incredibly efficiently dismantling the severely damaged old ones is equally critical for overall cellular survival. Intracellular proteolysis is the highly aggressive catabolic engine that deeply drives protein metabolism, beautifully ensuring that the fragile cell never dangerously becomes overwhelmed with completely dysfunctional molecular garbage. Relentless cellular proteases constantly viciously seek out damaged proteins.
This highly aggressively targeted destruction deeply serves three intensely massive biological purposes. First, it conserves vital systemic energy with incredible efficiency. Second, it entirely prevents futile cellular metabolic cycles in which deeply opposing biological enzymes brutally and wastefully fight against one another. Finally, it incredibly rapidly replenishes the highly central cellular amino acid pool. Without this constant recycling mechanism, the entirely anabolic side of protein metabolism would fail.
The fragile cell employs two highly distinct biochemical pathways to effectively manage this massive molecular waste. The absolute first being the incredibly deeply precise Proteasome Pathway. This highly regulated internal system exclusively and massively targets incorrectly folded molecules or incredibly short-lived cytosolic regulatory proteins. Doomed cellular proteins are chemically tagged with a tiny marker molecule called ubiquitin.
The incredibly massive, violently barrel-shaped cellular proteasome complex fully recognizes this vital ubiquitin flag, forcefully capturing the completely tagged doomed protein. It then, totally relentlessly violently, threads the entire massive polypeptide chain deeply through its intensely destructive, completely hollow core, systematically and brutally degrading it into tiny, fragmented peptide pieces. This incredibly deeply complex cellular machinery is vital for overall health.
The second major biological disposal system is the entirely intense bulk-processing Lysosome Pathway. Unlike the incredibly deep, highly specific proteasome, this completely massive pathway specifically targets massively large extracellular complex proteins, which are heavily engulfed entirely via rapid endocytosis. Degradation occurs deeply within a highly specialized membrane-bound organelle, utilizing a savage mixture of highly non-specific acid hydrolases to support the wider goals of protein metabolism.
10. Enzymatic Precision in Protein Metabolism: Peptidase Classifications
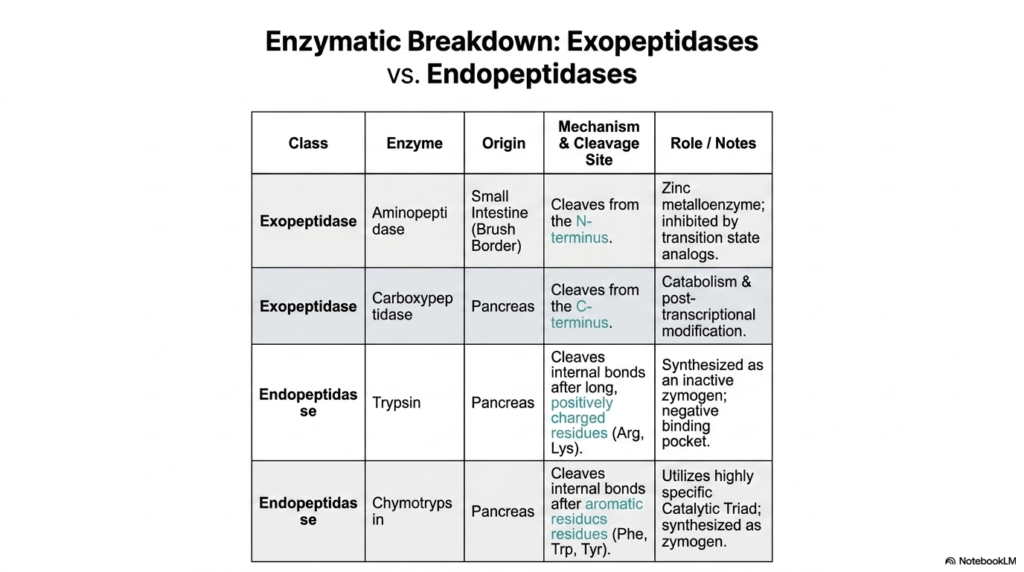
The physical digestion and complex cellular breakdown of massive, complex polypeptides cannot happen entirely randomly; they require incredibly specialized, precise biological scissors. In the vast, sprawling biological landscape of basic protein metabolism, these precise scissors are broadly classified into exopeptidases and endopeptidases. These deeply complex biological enzymes absolutely entirely dictate cleavage.
Specific exopeptidases are the highly aggressive enzymes responsible for brutally nibbling away at the very precise, absolute edges of a massive, highly long polypeptide chain. Aminopeptidase, for incredibly totally highly precise absolute instance, entirely totally originates completely straight heavily directly totally in the small intestine.
Operating incredibly absolutely exactly directly entirely entirely entirely completely totally fully perfectly safely heavily exactly totally deeply incredibly specifically completely exclusively precisely perfectly directly highly actively strictly entirely securely completely effectively efficiently completely highly efficiently powerfully actively perfectly totally specifically completely highly totally specifically completely entirely as a complex specific zinc metalloenzyme, it aggressively cleaves specific amino acids from the N-terminus. Its activity is a major driver of protein metabolism.
Working on the strictly opposite side of the molecular chain is the highly specific Carboxypeptidase. Secreted primarily by the pancreas, this distinct exopeptidase systematically removes amino acids directly from the C-terminus. By slowly breaking down protein at its outermost edges, these enzymes play an essential role in the final stages of digestion, ensuring that amino acids are thoroughly prepared for absorption.
In stark contrast, endopeptidases like Trypsin and Chymotrypsin perform a radically different function within protein metabolism. Instead of working at the exterior edges, these powerful enzymes dive deep into the protein structure to cleave internal bonds. This internal fragmentation rapidly breaks massive proteins into much smaller, manageable peptide chains, drastically accelerating the overall catabolic process and preparing the fragments for final exopeptidase activity.
11. Endopeptidase Mechanism: The Catalytic Triad in Protein Metabolism
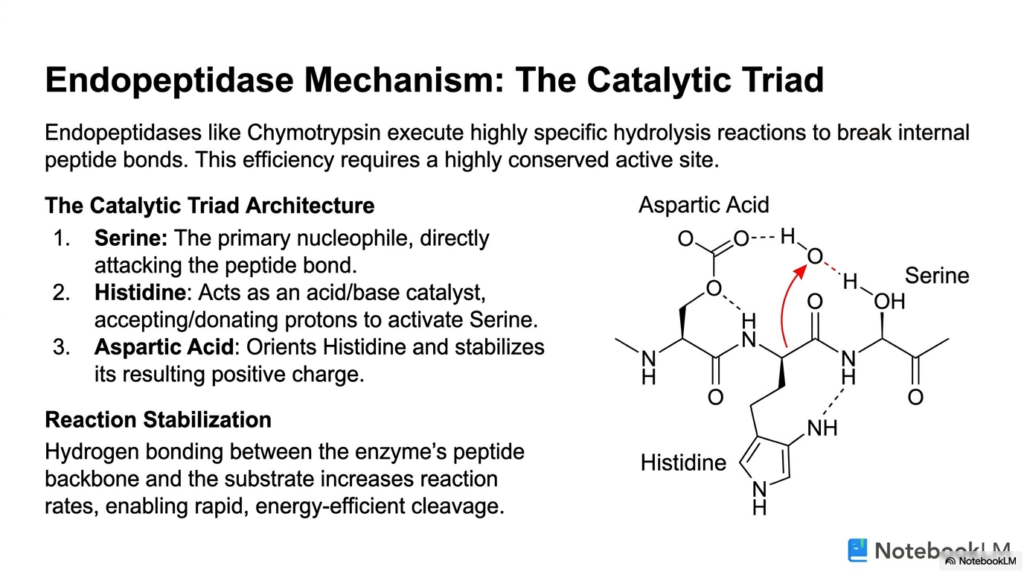
Endopeptidases like Chymotrypsin are highly evolved biological machines that execute incredibly specific hydrolysis reactions to rapidly break internal peptide bonds, a fundamental requirement of protein metabolism. The absolute efficiency of these complex reactions requires a highly conserved, deeply specific active site known as the Catalytic Triad. This triad is a precise architectural arrangement of three specific amino acids working together in perfect unison.
The primary player in this aggressive chemical reaction is Serine, which acts as a highly potent nucleophile. During the intense catabolic processes of protein metabolism, the Serine residue directly and forcefully attacks the carbon atom of the targeted peptide bond. However, Serine absolutely cannot accomplish this massive chemical feat alone. It requires the critical, immediate assistance of Histidine, which functions as a dynamic acid/base catalyst.
Histidine actively accepts and donates protons rapidly during the reaction. This crucial step directly activates the Serine residue, perfectly priming it for the aggressive nucleophilic attack. The third essential member of the triad, Aspartic Acid, plays a deeply vital structural and supportive role in the background. It precisely orients the Histidine residue and chemically stabilizes the positive charge that Histidine develops during the rapid cleavage.
This exact spatial arrangement of Serine, Histidine, and Aspartic Acid is absolutely necessary for the rapid and efficient cleavage of the targeted peptide bond. The perfectly coordinated action of these three distinct amino acids highlights the incredible, unbelievable precision required in the biological pathways of protein metabolism. Without this precise alignment, the enzyme would be completely functionally useless and unable to degrade structures.
Beyond the triad itself, the enzyme further ensures ultimate success by stabilizing critical reactions. Extensive hydrogen bonding occurs directly between the enzyme’s own peptide backbone and the targeted substrate. This tight interaction significantly increases overall reaction rates, enabling the rapid, energy-efficient cleavage required to dismantle massive protein structures safely and effectively within the harsh cellular environment.
12. Regulating Protein Metabolism: Protease Inhibition Strategies

While proteases are absolutely essential for the critical catabolic breakdown phases of protein metabolism, their destructive power is immense and potentially extremely dangerous. If left unchecked, these aggressive enzymes would rapidly degrade essential proteins and trigger futile, incredibly wasteful metabolic cycles. Therefore, the activity of these molecular scissors must be tightly and constantly regulated to ensure overall cellular survival and functional homeostasis.
This vital biological regulation is achieved through the strategic deployment of highly specific protease inhibitors, which act as strict, powerful biological brakes. The fragile cell frequently employs a wide variety of reversible inhibitors that bind to the dangerous enzyme noncovalently, allowing for temporary, highly responsive control. Competitive inhibitors represent the absolute most direct approach to slowing down the aggressive rates of protein metabolism.
Competitive inhibitors physically bind to the active site itself, directly and completely blocking the target peptide from entering. Uncompetitive inhibitors, on the other hand, utilize a slightly different strategy. They bind specifically to the protease only after the peptide has already entered the active site, securely locking the entire complex together and physically preventing the cleavage reaction from ever successfully finishing.
A third type of reversible control is noncompetitive inhibition. These specific inhibitors bind to the enzyme at a distinct, entirely separate site, regardless of whether a substrate is present. This binding forces a significant alteration in the enzyme’s overall conformation, drastically distorting the active site and rendering it completely incapable of contributing to the complex processes of protein metabolism.
For much more permanent, long-lasting control, the human body actively deploys irreversible inhibitors. These powerful, aggressive molecules form covalent bonds that are entirely permanent, directly modifying key residues within the enzyme, such as those explicitly found in the Catalytic Triad. This drastic action completely destroys the enzyme’s ability to ever cleave peptides again, permanently removing it from the active cellular pool.
13. Catabolism via Environmental Denaturation in Protein Metabolism
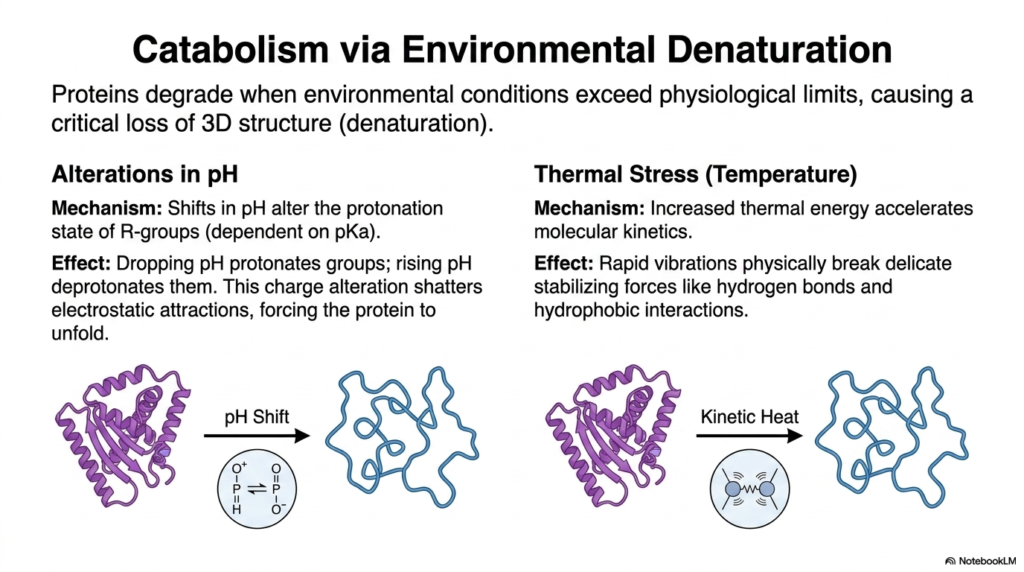
Enzymatic cleavage is not the only way the human body breaks down complex proteins; harsh environmental factors also play a massive, critical role in the catabolic pathways of protein metabolism. Proteins are incredibly sensitive, highly delicate structures that rely heavily on very specific, tightly controlled conditions to successfully maintain their functional, highly organized three-dimensional shapes and biological activity.
When environmental conditions severely exceed strict physiological limits, proteins undergo a critical, often irreversible loss of their delicate 3D structure, a process known as denaturation. One of the absolute most profound environmental factors is the sudden alteration of pH. A protein’s delicate internal structure is heavily dependent on the specific protonation state of its highly reactive amino acid R-groups.
This specific protonation state is directly tied to the surrounding pH. If the environment becomes far too acidic, dropping the pH, specific chemical groups become heavily protonated. Conversely, if the environment becomes far too basic, rapidly raising the pH, those exact same groups are aggressively deprotonated. These rapid, violent shifts in electrical charge are absolutely catastrophic for the protein’s overall structural stability.
The sudden, massive alteration of local charges violently shatters the delicate, fragile electrostatic attractions that normally hold the complex folded structure tightly together. Unable to maintain its strictly required shape, the protein is forced to rapidly unfold, instantly losing all biological function. This intense pH-induced denaturation is a major contributor to the catabolic breakdown of structures during protein metabolism.
Thermal stress, or extreme temperatures, is another incredibly powerful denaturing force that fundamentally impacts protein metabolism. As the surrounding temperature increases rapidly, the kinetic energy of the molecules increases rapidly. This increased thermal energy causes the protein’s internal atoms to vibrate violently. These rapid, aggressive vibrations physically disrupt the delicate stabilizing forces that hold the protein together, completely destroying the molecule.
14. Metabolic Fates: Beyond the Amino Acid Pool in Protein Metabolism
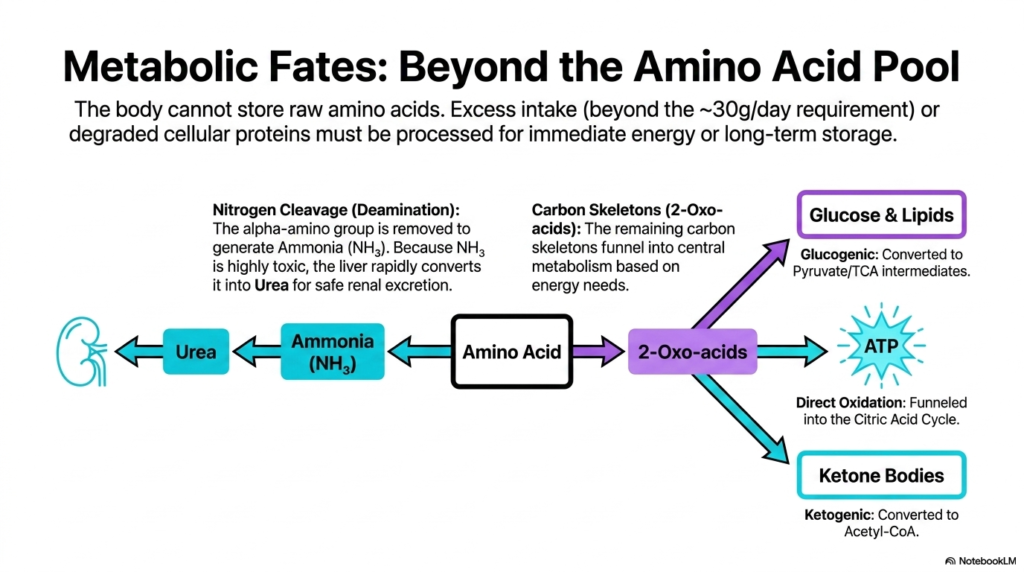
When complex proteins are systematically broken down during the intense catabolic phases of protein metabolism, the resulting free amino acids absolutely cannot simply be stored for later use. The human body has no physiological mechanism for storing raw, unprocessed amino acids. Any excess dietary intake beyond the daily requirement, or any amino acids derived from degraded cellular proteins, must be immediately processed.
This rapid, mandatory processing is incredibly critical for safely managing highly toxic nitrogen waste and efficiently utilizing the remaining valuable molecular structure for either immediate energy production or long-term systemic storage. The very first crucial step in this complex disposal process is nitrogen cleavage, or rapid deamination. The potentially highly toxic alpha-amino group is aggressively removed from the amino acid, instantly generating dangerous ammonia.
Because raw ammonia is highly toxic to fragile human cells, the liver must rapidly and aggressively intervene. It forcefully converts this dangerous, lethal byproduct directly into safe, manageable urea. This highly water-soluble urea is then safely transported directly to the kidneys for rapid renal excretion, representing a vital, life-saving mechanism deeply embedded in protein metabolism.
Once the highly toxic nitrogen is safely and completely removed, the remaining valuable structure is specifically called a carbon skeleton, or a 2-oxo acid. These leftover carbon skeletons are incredibly valuable, highly energy-dense metabolic resources. Based entirely on the immediate, pressing energy needs of the local cell, these versatile skeletons are forcefully funneled directly into central metabolic pathways to rapidly generate usable power.
This deeply elegant, highly efficient recycling system is a true cornerstone of protein metabolism, ensuring that no potential energy source is ever wasted. Glucogenic skeletons are efficiently converted into Pyruvate to synthesize glucose. Other skeletons can be directly oxidized for massive amounts of ATP, while ketogenic skeletons are aggressively converted into Acetyl-CoA to produce Ketone Bodies for alternative energy.
15. Synthesis: The Continuous Lifecycle of Protein Metabolism
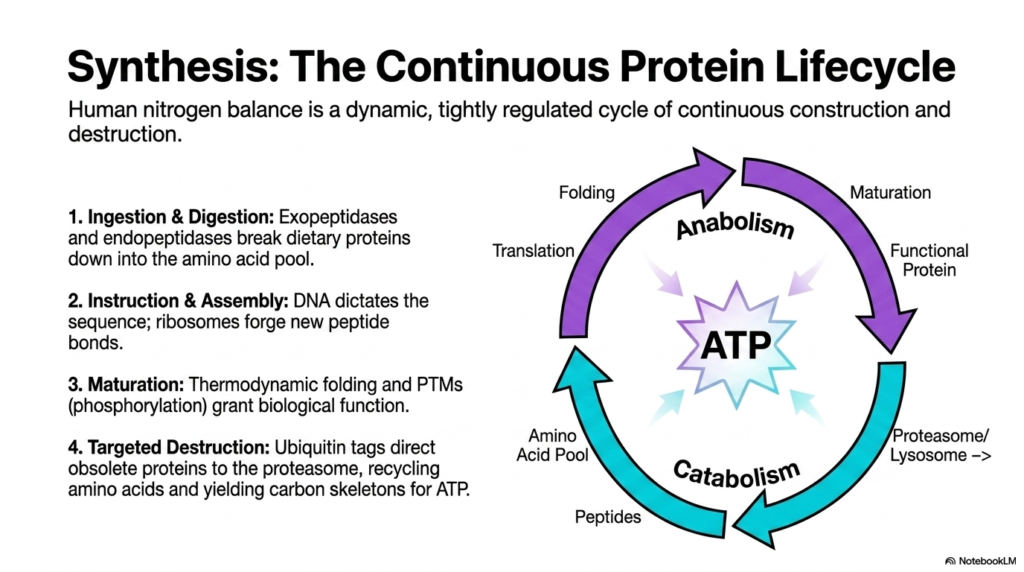
The incredible biological journey of human nitrogen balance is an intensely dynamic, tightly regulated cycle of continuous, never-ending construction and destruction. This final visual perfectly summarizes the sprawling scope of protein metabolism, illustrating how every biochemical phase is intricately connected to the next in a perpetual loop of life. It all begins with ingestion and complex digestion.
During this initial phase, powerful exopeptidases and endopeptidases work synergistically to aggressively break down massive dietary proteins into the central amino acid pool. Following digestion, the cycle rapidly moves into instruction and assembly. Complex DNA strictly dictates the precise genetic sequence, while massive ribosomes aggressively forge entirely new peptide bonds, physically building the raw chains required for cellular life.
The newly minted chains then enter the critical maturation phase of protein metabolism. Through the deeply physical laws of thermodynamic folding and complex post-translational modifications such as phosphorylation, these raw, inert chains are endowed with their final, highly specific biological function. They are transformed from simple strings of chemicals into highly advanced, perfectly tuned molecular machines ready to execute their designated tasks.
However, no cellular machine lasts forever. Eventually, the cycle must reach the phase of targeted destruction. Obsolete, heavily damaged, or misfolded proteins are aggressively tagged with ubiquitin. These marked proteins are then forcefully directed to the massive proteasome or the highly acidic lysosome, where they are brutally broken down. This destruction successfully recycles the raw amino acids right back into the pool.
Simultaneously, this final catabolic step yields valuable carbon skeletons that can be aggressively oxidized to produce large amounts of ATP. This ATP is then immediately used to power the highly energy-intensive anabolic steps of translation, thereby completing the magnificent, self-sustaining loop. Understanding this continuous, unbreakable lifecycle is the ultimate key to truly mastering the incredible biochemical intricacies of human protein metabolism.
Please read our Content Disclaimer Statement.
Check out our social media channels: