71. Plasma Membrane Functions and Composition
Imagine a bustling city without walls, roads, or traffic control; chaos would reign instantly. The same principle applies to microscopic life. This blog post explores the fundamental structural and operational blueprints of cellular life. The following slide deck breaks down complex biochemical structures and processes into digestible concepts. It provides a comprehensive look at how biological boundaries maintain order, facilitate communication, and drive the essential reactions that make life possible. Let us dive into the intricate world of cellular organization and explore these dynamic systems.
Slide 1: Introduction to Plasma Membrane Functions
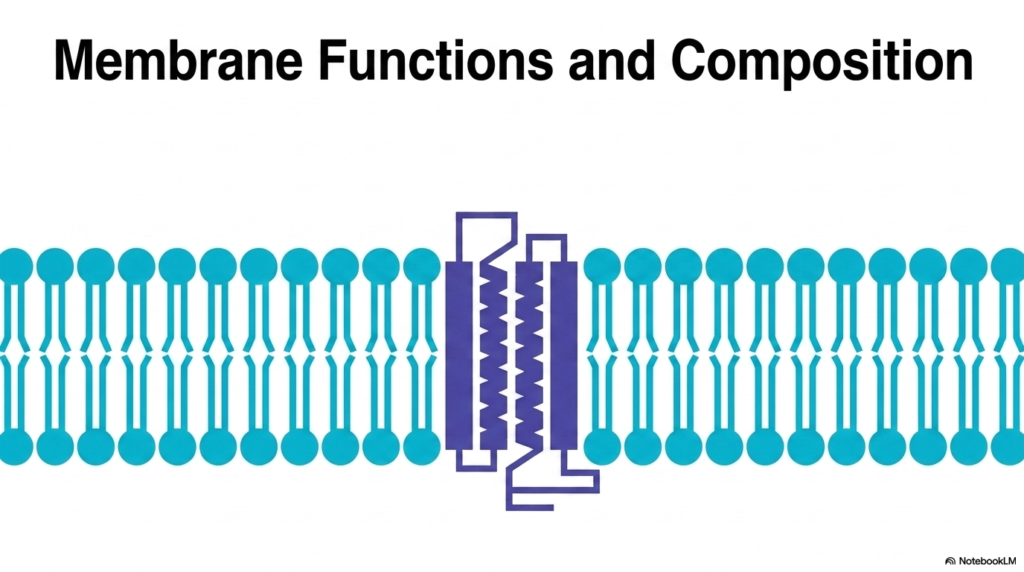
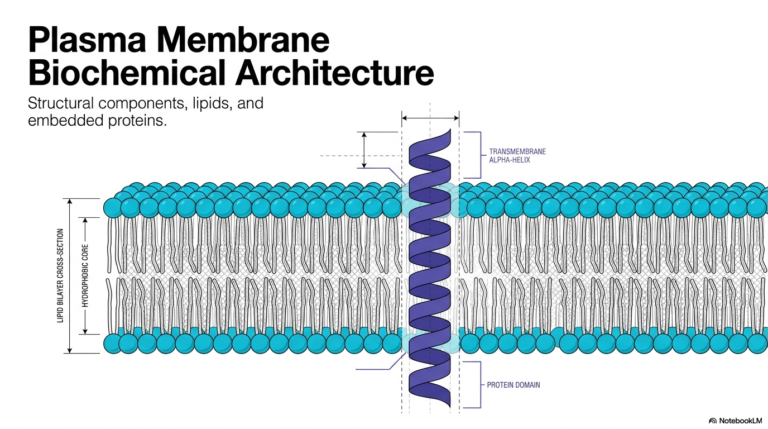
The study of cellular biology begins with an appreciation of boundaries. The visual representation on this slide introduces the foundational architecture that enables all diverse Plasma Membrane Functions. At first glance, one sees a simple lipid bilayer peppered with integral proteins. However, this elegant arrangement is the structural basis for life itself. The amphipathic nature of the lipid molecules creates a stable yet fluid matrix. This matrix separates the internal cellular environment from the unpredictable external world. The embedded proteins act as the active machinery, bridging the gap between inside and outside.
Understanding this basic structure is crucial for grasping the vast array of Plasma Membrane Functions executed by every living cell. The biological boundary is never a static wall. Instead, it is a highly dynamic and responsive interface. The lipids provide the solvent, while the proteins provide the specific action. This combination allows the cell to sense its environment, import nutrients, and export waste products. Without this specific structural arrangement, the cell would simply dissolve into its surroundings, unable to maintain the necessary chemical disequilibrium required for life.
As students of biochemistry, we recognize that the interplay between structure and function is paramount. The diverse Plasma Membrane Functions we will explore depend entirely on this core bilayer template. The transmembrane proteins shown here are just one example of how the boundary is selectively breached. Over the next slides, the precise molecular composition and the sophisticated mechanisms that operate within this microscopic layer will become clear. The biological boundary is an energetic engine, a communication hub, and a protective shield all rolled into one microscopic structure.
The composition of this bilayer is meticulously controlled by the cell. It involves a continuous cycle of synthesis and degradation to adapt to metabolic demands. Every single lipid and protein molecule contributes to the overall stability and functionality of the entire system. Therefore, mastering the basic layout presented here serves as the perfect launchpad. It prepares the observer to understand the complex biochemical realities that govern cellular existence and health.
Slide 2: Structural Asymmetry and Plasma Membrane Functions
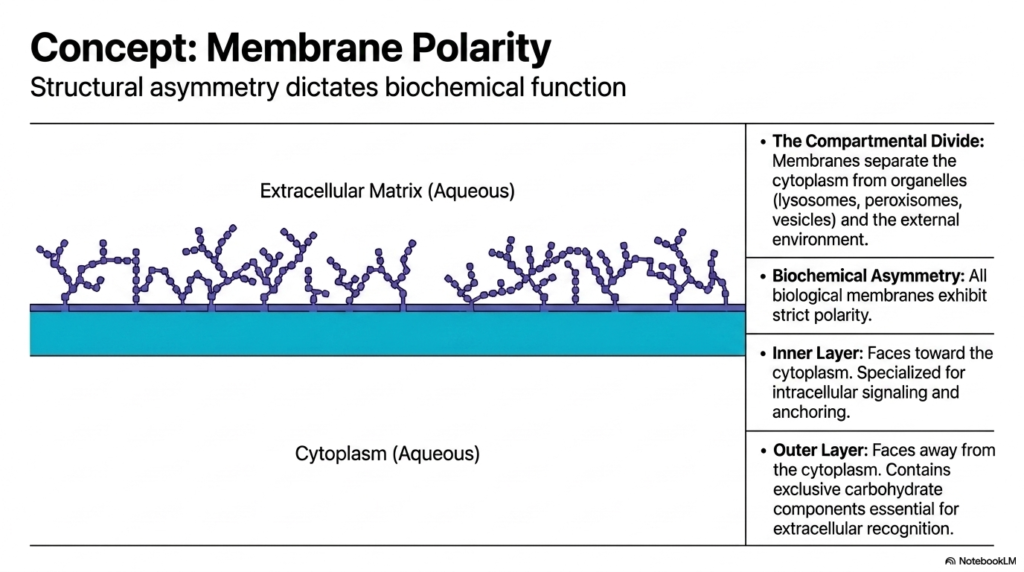
A critical biochemical principle dictating Plasma Membrane Functions is strict structural asymmetry. All biological membranes exhibit inherent polarity, meaning the inner leaflet facing the cytoplasm is biochemically distinct from the outer leaflet facing the extracellular matrix. This compartmental division is essential for separating the internal cytoplasm from distinct organelles, such as lysosomes, and from the unpredictable external environment. Polarity is not a random occurrence but a highly regulated feature established during the synthesis of membrane components in the endoplasmic reticulum. It dictates how the cell interacts with its surroundings.
The inner layer, facing the aqueous cytoplasm, plays a specific role in Plasma Membrane Functions. This cytosolic leaflet is specialized for intracellular signaling and anchoring the cytoskeleton. Specific lipid species are concentrated on this inner side to recruit signaling proteins when the cell receives outside stimuli. This precise spatial arrangement ensures that internal communication cascades trigger rapidly and efficiently. Furthermore, structural proteins bind to this inner surface, giving the cell its distinct shape and physical resilience against mechanical stress.
Conversely, the outer layer presents a completely different biochemical face to the world. It contains exclusive carbohydrate components, such as glycoproteins and glycolipids. These sugar chains project outward, creating a molecular identification code critical for many Plasma Membrane Functions. This exterior coat enables extracellular recognition, immune evasion, and adherence to neighboring cells. The stark contrast between the inner signaling hub and the outer recognition surface highlights how structural asymmetry is the ultimate driver of specialized cellular operations. The boundary is an intelligent, two-faced mediator.
Maintaining this absolute polarity requires significant energy expenditure by the cell. Specialized enzymes continually move specific lipids to their correct leaflet. If this asymmetry is lost, it often signals cellular damage or programmed cell death. Therefore, the strict maintenance of membrane polarity is a fundamental requirement for cellular survival and proper physiological function across all tissue types.
Slide 3: Enclosure, Anchoring, and Plasma Membrane Functions
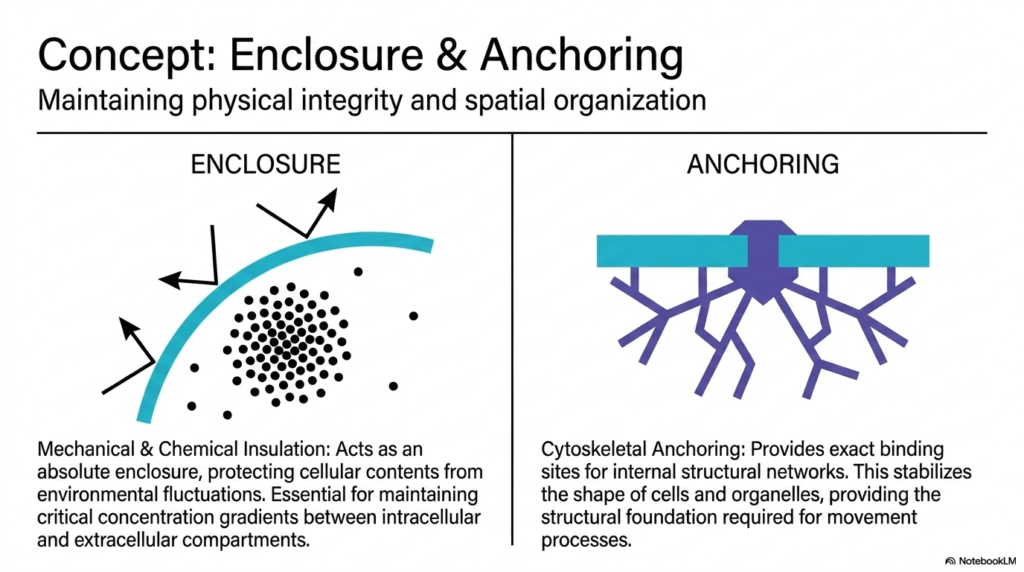
The most immediate and obvious core Plasma Membrane Function is creating a secure physical enclosure. The membrane acts as an absolute mechanical and chemical insulator. It protects sensitive internal cellular contents from harsh environmental fluctuations. This insulating property is critical for maintaining steep concentration gradients between the intracellular and extracellular compartments. For example, cells must maintain low internal sodium levels and high potassium levels. The lipid bilayer prevents these ions from freely diffusing, thereby storing potential energy used for numerous biochemical processes.
Beyond simple enclosure, the membrane provides an essential foundation for structural anchoring. This is another vital aspect of overall Plasma Membrane Functions. The inner surface provides exact binding sites for internal structural networks, primarily the cytoskeleton. By tethering these internal protein filaments to the stable lipid bilayer, the cell establishes its physical integrity. This anchoring mechanism stabilizes the shape of both the entire cell and its internal organelles, preventing them from collapsing under mechanical pressure or osmotic stress.
The synergy between enclosure and anchoring facilitates some of the most dynamic Plasma Membrane Functions observed in biology. Because the internal structural networks are securely bound to the boundary, the cell can coordinate complex movement processes. When the cytoskeleton contracts or expands, it pulls on the membrane, allowing for cellular crawling, division, or the engulfment of external particles. The boundary is therefore not just a passive sack. It is an actively manipulated architectural feature that gives the cell physical agency in its environment.
This dual role of insulation and structural support highlights the multifunctional nature of cellular boundaries. Without the protective enclosure, the complex biochemical reactions of life would be diluted and would halt completely. Simultaneously, without the robust anchoring mechanisms, the cell would lack the organization required to execute directed tasks. Together, these physical properties form the prerequisite conditions for all other advanced metabolic and communicative operations.
Slide 4: Regulated Transport as Key Plasma Membrane Functions
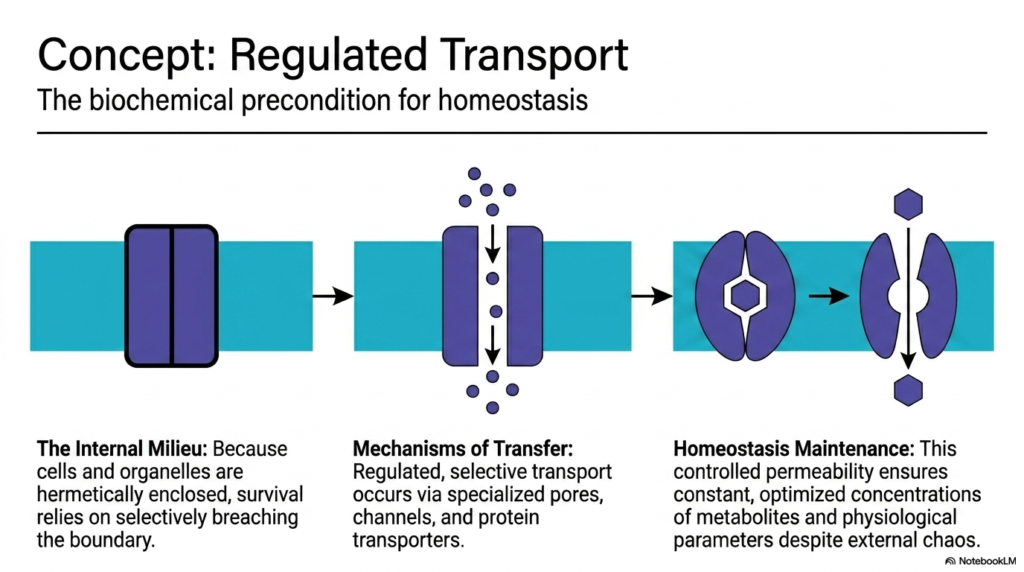
Because cells are hermetically enclosed to protect their internal milieu, their survival depends entirely on selectively breaching that boundary. Regulated transport is perhaps the most heavily studied of all Plasma Membrane Functions. It stands as the absolute biochemical precondition for cellular homeostasis. The lipid bilayer is impermeable to most essential water-soluble molecules, meaning nutrients cannot enter, and metabolic waste cannot exit without assistance. The cell solves this problem by deploying specialized protein structures designed to move specific cargo across the divide.
The mechanisms of transfer involve a fascinating array of specialized pores, channels, and protein transporters. These protein machines are the active agents carrying out transport-related Plasma Membrane Functions. Channels provide gated corridors for rapid ion flow down concentration gradients. Meanwhile, transporters undergo complex conformational changes to physically pump molecules across the boundary, often requiring significant cellular energy in the form of ATP. This level of control allows the cell to dictate exactly what enters and leaves, adapting to shifting metabolic demands in real-time.
This controlled permeability is the cornerstone of homeostasis maintenance, representing the ultimate goal of these Plasma Membrane Functions. By tightly regulating transport, the cell ensures constant, optimized metabolite concentrations, regardless of chaos in the external environment. This creates a stable internal chemistry where sensitive enzymatic reactions can proceed without interruption. The precise balance of ions, sugars, and amino acids is maintained purely through the tireless work of these membrane-embedded transport proteins.
When these transport mechanisms fail, the consequences for the cell are often catastrophic. Many severe human diseases stem directly from defective transport proteins. Therefore, understanding the kinetics and structural biology of these membrane channels is a major focus in modern pharmacology. By targeting these specific transport gateways, medical professionals can effectively modulate cellular behavior and restore physiological balance.
Slide 5: Signaling, Interaction, and Plasma Membrane Functions
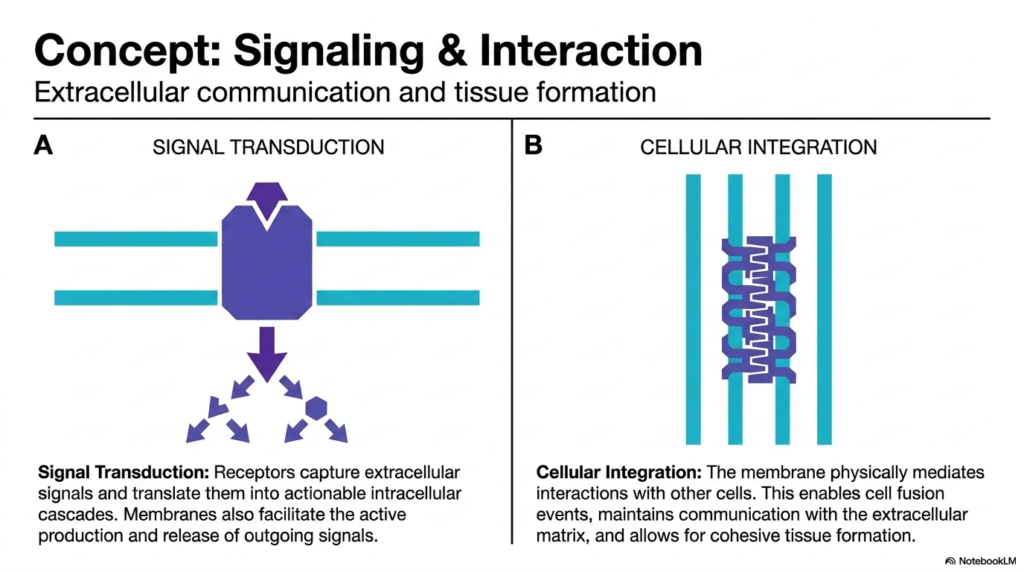
Cells do not exist in isolation; they must constantly interpret their environment and collaborate. Therefore, mediating extracellular communication is one of the most critical Functions of the Plasma Membrane. Signal transduction is the process by which receptors embedded in the cell membrane capture external signals, such as hormones or neurotransmitters. These receptors then undergo structural shifts to translate the external message into actionable intracellular cascades. This allows the cell to alter its gene expression or metabolism without the original signal molecule ever actually entering the cytoplasm.
Furthermore, the membrane facilitates the active production and release of outgoing signals. This makes two-way communication a central pillar of Plasma Membrane Functions. Beyond simple chemical messaging, the boundary physically mediates direct cellular integration. Transmembrane proteins reach out and physically bind to matching proteins on adjacent cells or to the extracellular matrix. This physical linking enables vital biological processes, such as cell fusion, allowing distinct cells to combine their contents when necessary.
This physical and chemical coupling is essential for cohesive tissue formation. It highlights the vast scope of Plasma Membrane Functions in multicellular organisms. Without these interactive capabilities, cells could never organize into the complex tissues and organs required for advanced life. The membrane serves as the ultimate social interface, ensuring that billions of individual cells can operate as a unified, synchronized organism. It is the hardware that runs the cellular internet, coordinating massive biological networks.
The complexity of these signaling hubs cannot be overstated. A single cell may possess thousands of different receptor types, each waiting for a specific molecular key. When triggered, these receptors assemble massive, temporary protein complexes on the inner leaflet of the membrane to amplify the signal. This rapid, targeted response system allows the organism to react to danger, digest food, and maintain overall physiological harmony at incredible speeds.
Slide 6: Interfacial Catalysis within Plasma Membrane Functions
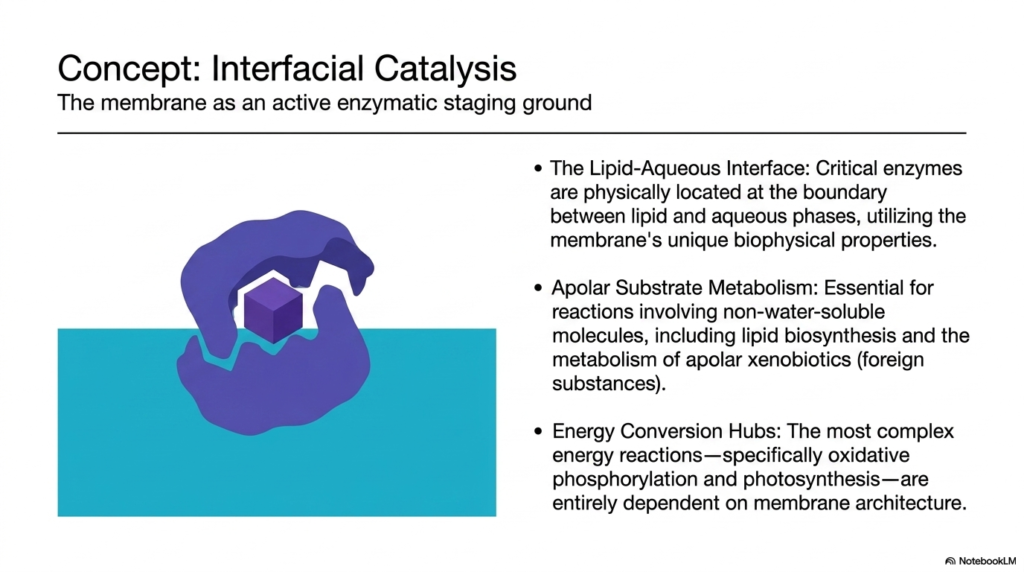
While the cytoplasm is often viewed as the main site for chemical reactions, the boundary itself is an active enzymatic staging ground. Facilitating interfacial catalysis is one of the most specialized Plasma Membrane Functions. The lipid-aqueous interface offers unique biophysical properties not found anywhere else in the cell. Critical enzymes are physically anchored at this boundary between the apolar lipid phase and the watery environment. This strategic positioning allows them to interact simultaneously with water-soluble substrates and lipid-soluble molecules.
This environment is absolutely essential for apolar substrate metabolism. Many vital biochemicals are highly hydrophobic and cannot dissolve in the cytoplasm. Therefore, coordinating the synthesis and modification of these molecules relies entirely on specific Plasma Membrane Functions. Enzymes embedded in the lipid matrix regulate lipid biosynthesis, ensuring the membrane can repair and expand. Furthermore, the metabolism of apolar xenobiotics, which are potentially toxic foreign substances, occurs safely within this specialized interfacial staging area.
Perhaps most importantly, these boundaries serve as the ultimate hubs for energy conversion. The execution of complex energy reactions relies completely on Plasma Membrane Functions and structural architecture. Processes like oxidative phosphorylation in mitochondria and photosynthesis in chloroplasts are entirely dependent on membrane-bound enzyme complexes. These systems utilize the insulating properties of the bilayer to pump protons and create energy gradients. The membrane physically holds the electron transport chain components in perfect spatial alignment, turning the boundary into a biological battery.
Without the stable, two-dimensional platform provided by the lipid bilayer, these massive enzymatic assembly lines would drift apart and cease functioning. The ability to concentrate reactants within a restricted two-dimensional plane massively increases reaction rates compared to three-dimensional diffusion in the cytoplasm. This demonstrates how the physical architecture of the cellular boundary directly drives the thermodynamic viability of life itself.
Slide 7: Composition Underpinning Plasma Membrane Functions
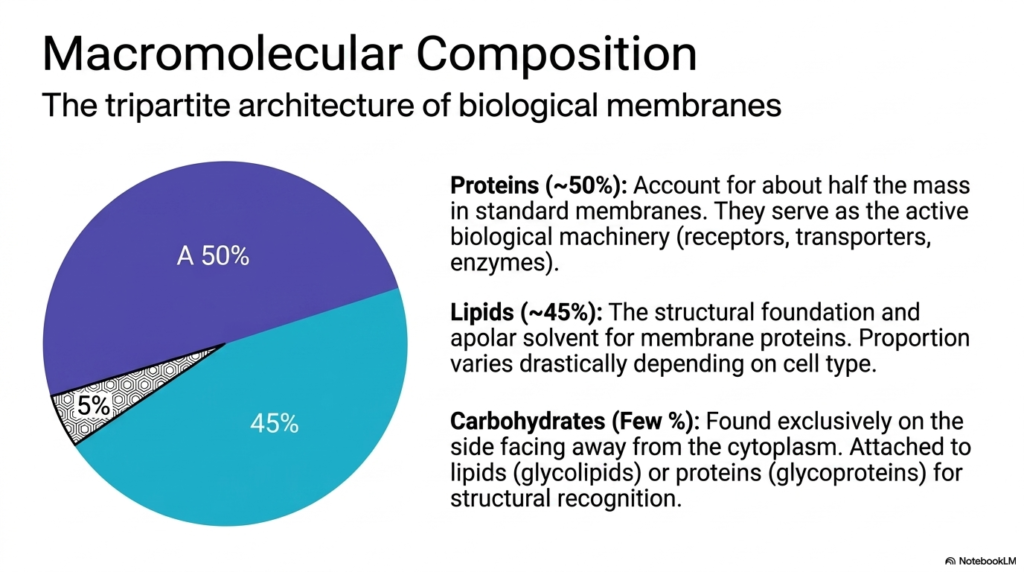
To fully grasp how a boundary executes its tasks, one must examine its tripartite architecture. The macromolecular composition directly determines the scope and efficiency of all Plasma Membrane Functions. A standard biological boundary is roughly divided into lipids, proteins, and a small fraction of carbohydrates. Proteins account for approximately fifty percent of the mass in standard biological boundaries. These proteins serve as the active biological machinery, functioning as the receptors, transporters, and enzymes discussed previously. They perform the heavy lifting.
Lipids make up about forty-five percent of the mass and provide the essential structural foundation. They act as the apolar solvent that houses and supports the active membrane proteins, enabling all protein-driven Plasma Membrane Functions. The lipid matrix is not merely a passive structural element; its fluidity and thickness directly modulate protein activity. The proportion of lipids to proteins varies drastically depending on the specific cell type and its unique physiological requirements. This ratio is highly adaptable.
Finally, carbohydrates account for only a few percent of the total mass, yet they are vital for specific Plasma Membrane Functions. They are found exclusively on the side facing away from the cytoplasm. These sugar chains are attached to lipids, forming glycolipids, or attached to proteins, forming glycoproteins. This carbohydrate coating provides structural recognition, allowing cells to identify one another and sort themselves correctly within complex tissues. It acts as a molecular fingerprint.
This tripartite mixture is never static. The cell constantly synthesizes new components and degrades old ones to adjust the boundary’s physical properties. By subtly shifting the ratio of lipids, proteins, and carbohydrates, the cell adapts to temperature changes, metabolic stress, and developmental signals. Understanding this proportional distribution is the key to predicting how a specific cell will interact with its immediate environment and perform its required physiological duties.
Slide 8: Compositional Extremes and Plasma Membrane Functions
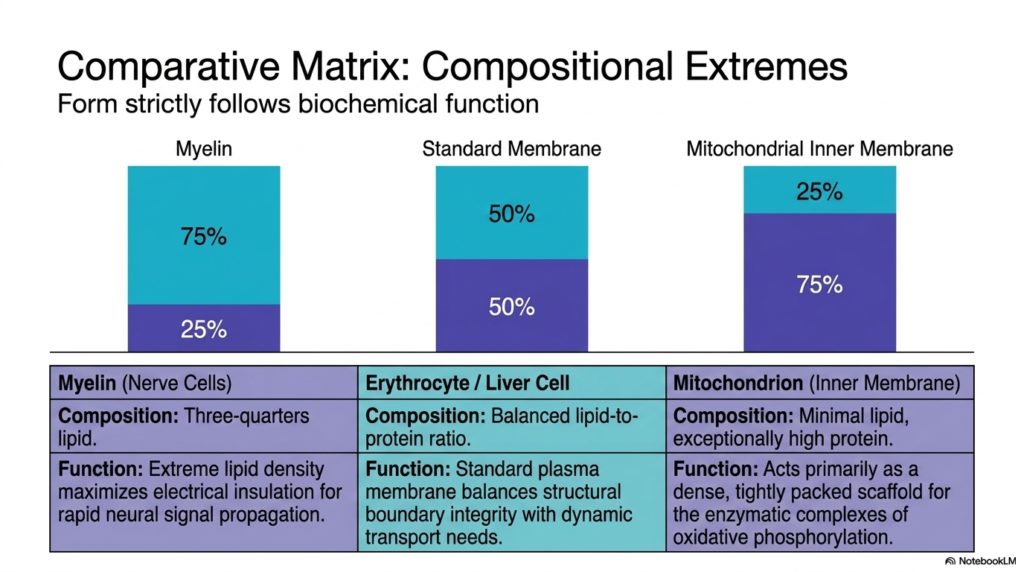
In biochemistry, form strictly follows function. This concept is vividly illustrated by examining cells with extreme compositional ratios to understand specialized Plasma Membrane Functions. This slide contrasts three vastly different biological boundaries. First, myelin forms the protective sheath around nerve cells. Its composition is heavily skewed, consisting of roughly 75% lipid and only 25% protein. This extreme lipid density maximizes electrical insulation, which is a highly specialized requirement for rapid neural signal propagation.
On the other end of the spectrum is the inner mitochondrial membrane. Here, the requirements for Plasma Membrane Functions demand a completely inverted ratio. This boundary contains minimal lipid content and is exceptionally high in protein, at roughly seventy-five percent. The inner mitochondrial membrane serves primarily as a dense, tightly packed scaffold for the enzymatic complexes that drive oxidative phosphorylation. It requires massive amounts of active machinery packed into a small area to meet the huge energy demands of the cell.
Between these extremes sits the standard erythrocyte or liver cell. This represents the baseline for typical Plasma Membrane Functions. These cells maintain a balanced fifty-fifty lipid-to-protein ratio. This balanced standard membrane perfectly harmonizes structural boundary integrity with dynamic transport needs and signaling capabilities. By observing these extremes, students can clearly see that the cell customizes its boundary architecture. It synthesizes specific ratios to perfectly match its designated physiological role.
This incredible plasticity in composition demonstrates the evolutionary adaptability of cellular life. A nerve cell would fail to conduct impulses if it possessed the protein-dense structure of a mitochondrion. Conversely, a mitochondrion could never generate sufficient ATP if its boundaries were insulated heavily like a myelin sheath. Therefore, analyzing the precise lipid-to-protein mass ratio provides immediate, critical insight into the primary metabolic function and evolutionary history of any given biological tissue.
Slide 9: Lipid Diversity Driving Plasma Membrane Functions
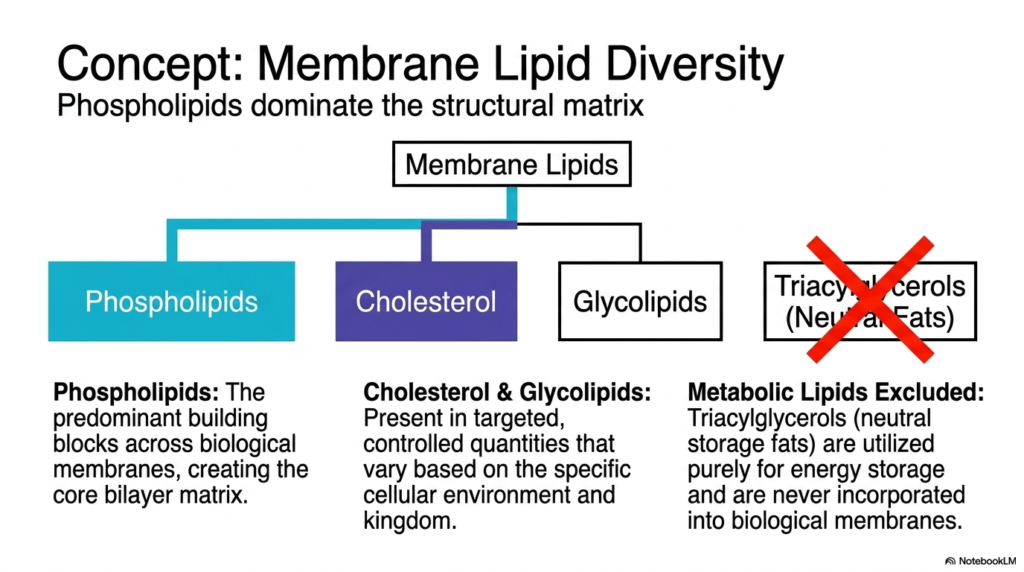
The structural matrix of the cellular boundary is not monolithic; it relies on immense chemical variety. Understanding membrane lipid diversity is crucial for explaining how different Plasma Membrane Functions are regulated. The primary building blocks across all biological boundaries are phospholipids. These amphipathic molecules possess hydrophilic heads and hydrophobic tails, naturally forming the bilayer core in aqueous environments. Phospholipids dominate the structural landscape and provide the foundational solvent required for protein integration.
Beyond the core phospholipids, other lipid species are deployed to fine-tune specific Plasma Membrane Functions. Cholesterol and glycolipids are present in targeted, carefully controlled quantities. These molecules vary significantly based on the specific cellular environment and biological kingdom. Cholesterol inserts itself between phospholipids, acting as a fluidity buffer. It prevents the boundary from becoming too rigid in the cold or too liquid in the heat. Glycolipids, as discussed, are strictly reserved for the outer leaflet to facilitate external recognition.
It is equally important to note what is excluded from these structures to understand the Functions of the Plasma Membrane. Metabolic lipids, specifically triacylglycerols or neutral fats, are strictly excluded from biological boundaries. Triacylglycerols are highly hydrophobic and lack the polar head group required to form a stable interface with water. They are used solely for energy storage within intracellular droplets. They are never incorporated into the boundary’s functional architecture.
The precise mixture of these lipid species dictates the thickness, curvature, and fluidity of the localized boundary area. Cells actively cluster specific lipids together to form specialized microdomains, often called lipid rafts. These rafts float within the larger liquid matrix and serve to concentrate specific signaling proteins into functional hubs. Therefore, lipid diversity is not just a structural curiosity; it is a highly active mechanism for organizing and optimizing cellular signaling pathways.
Slide 10: Evolutionary Biomarkers and Plasma Membrane Functions
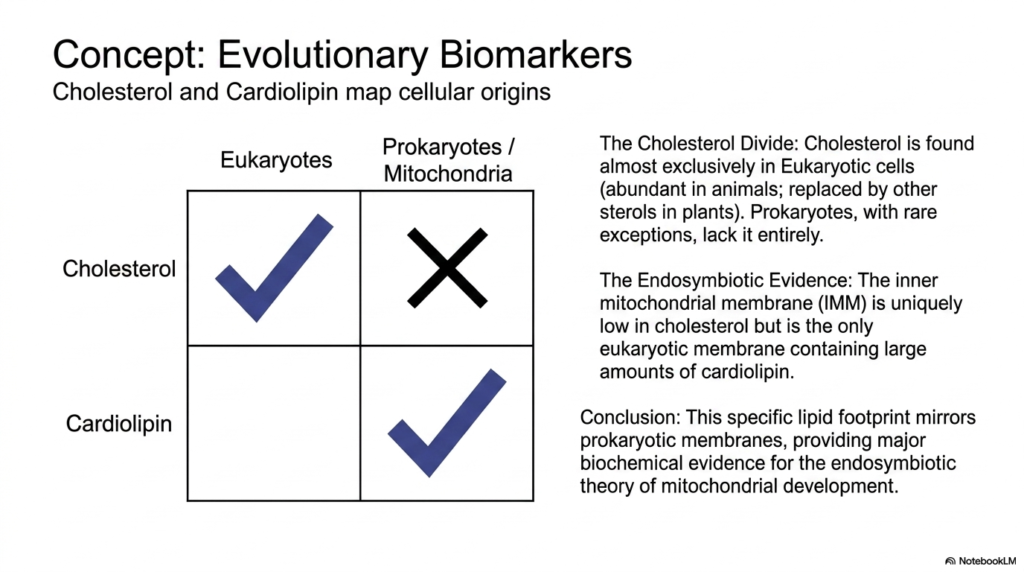
The specific lipid composition of a cell serves as a historical record, revealing deep evolutionary relationships. Certain lipid species act as biomarkers, providing insights into the origins of diverse Plasma Membrane Functions. A striking example is the distribution of cholesterol. Cholesterol is found almost exclusively in eukaryotic cells, is highly abundant in animal tissues, and is replaced by similar sterols in plants. Prokaryotes, with rare exceptions, lack cholesterol entirely in their membranes.
Conversely, cardiolipin is another vital biomarker associated with specific Plasma Membrane Functions. Cardiolipin is a unique, complex lipid found abundantly in the membranes of prokaryotic bacteria. Interestingly, the inner mitochondrial membrane is uniquely low in cholesterol but is the only eukaryotic location containing large amounts of cardiolipin. This stark compositional difference between the cell’s outer boundary and its internal power plant is a crucial biochemical footprint.
This specific lipid footprint perfectly mirrors prokaryotic membranes, validating our understanding of how modern Plasma Membrane Functions evolved. It provides major biochemical evidence for the endosymbiotic theory of mitochondrial development. The presence of cardiolipin strongly supports the concept that mitochondria were once free-living bacteria absorbed by an ancient host cell. The ancient bacterial boundary was preserved and adapted to become the highly specialized energy-converting inner membrane.
By mapping these lipid signatures, biochemists can trace the lineage of cellular life back billions of years. The distinct separation of cholesterol and cardiolipin demonstrates that fundamental biochemical mechanisms are highly conserved across evolutionary time. This historical context enriches our understanding of cellular biology. It proves that the sophisticated boundaries surrounding modern human cells are the result of ancient microbial mergers and relentless evolutionary refinement over countless generations.
These ancient lipid species are not merely relics; they are strictly required for survival. For instance, the unique physical structure of cardiolipin is essential for stabilizing the massive protein complexes responsible for ATP production in mitochondria. Without this specific ancient lipid, the entire energy grid of the modern eukaryotic cell would catastrophically fail. Thus, evolutionary history directly dictates modern metabolic capability.
Slide 11: Synthesis of Plasma Membrane Functions
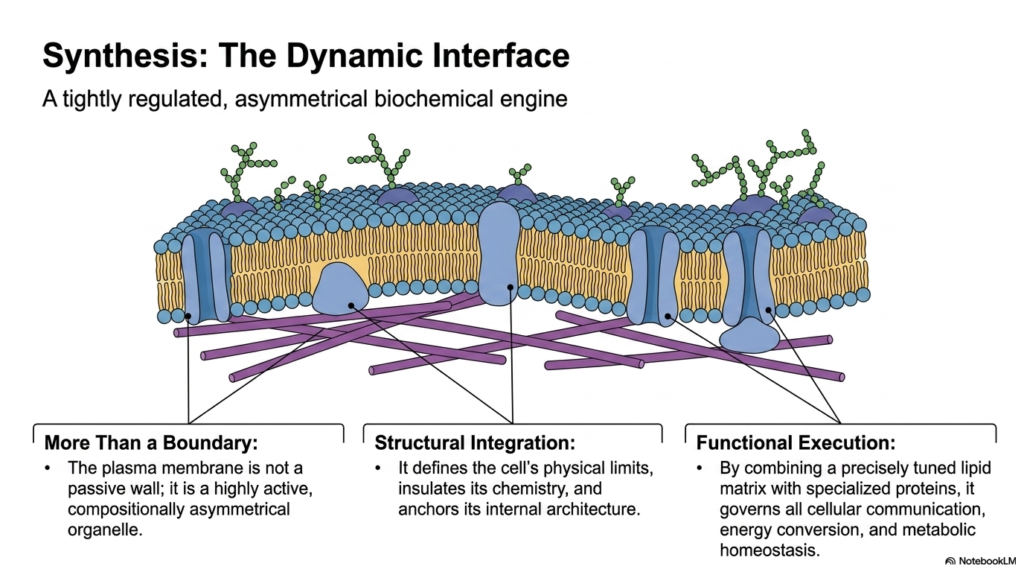
Bringing all these concepts together, the biological boundary emerges as a tightly regulated, asymmetrical biochemical engine. The grand synthesis of all Plasma Membrane Functions reveals that the boundary is far more than a simple wall. It is a highly active, compositionally asymmetrical organelle in its own right. It dictates the cell’s physical limits while simultaneously insulating its delicate internal chemistry from external chaos. It physically anchors the internal architecture, giving the cell form and stability.
This structural integration is seamlessly paired with complex functional execution, which is the hallmark of advanced Plasma Membrane Functions. By combining a precisely tuned lipid matrix with specialized, highly evolved proteins, the boundary governs all critical interactions. It manages cellular communication networks, oversees vital energy conversion processes, and ensures strict metabolic homeostasis. The lipid solvent and the protein machinery work in absolute concert to keep the cell alive and responsive.
Understanding the full scope of Plasma Membrane Functions is the ultimate goal for students of molecular biology and medicine. This dynamic interface is the primary target for countless pharmaceutical interventions. By manipulating how receptors signal, how channels transport, or how lipids organize, modern medicine can correct cellular dysfunction. The boundary is the frontline of cellular defense and the primary conduit for interaction with the surrounding universe.
Ultimately, the cellular boundary is a masterpiece of biochemical engineering. It perfectly balances the contradictory needs for isolation and connection. It must remain structurally robust to prevent the leakage of essential metabolites, yet fluid enough to allow protein machines to shift, bind, and transport materials. This exquisite balance of biophysical properties ensures that every single cell can operate autonomously while remaining fully integrated into the larger, complex organism.
Mastering these concepts provides a profound appreciation for the microscopic mechanisms that sustain life. Every heartbeat, every thought, and every metabolic adjustment relies completely on the rapid, flawless execution of the boundary mechanisms detailed in this presentation. The study of these cellular interfaces remains one of the most exciting and medically relevant frontiers in modern biochemical research.
Please read our Content Disclaimer Statement.
Check out our social media channels: