68. Mitochondria Explained: The Cell’s Biochemical Powerhouse
Imagine trying to power a bustling, modern metropolis with a single household battery. It simply would not work. Eukaryotic cells face an immense energy demand, which is exactly why they rely on specialized biological powerhouses to sustain life. This presentation details the intricate architecture and metabolic machinery of these dynamic organelles. The core purpose of this guide is to break down complex biochemical structures and processes into digestible, fundamental concepts, providing college and medical students with a clear, energetic understanding of cellular energy dynamics.
Slide 1: General Structure and Biochemistry of Mitochondria
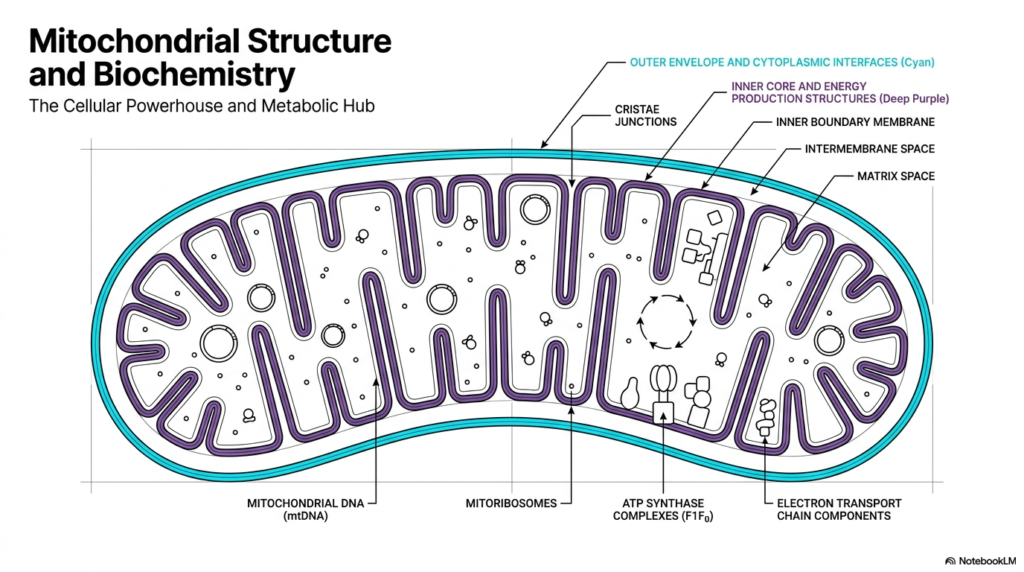
The first crucial concept to understand is the general structural blueprint of mitochondria. These organelles are characterized by a highly specialized double-membrane architecture that creates distinct internal compartments. This structural division is the physical foundation that allows mitochondria to function as the primary metabolic hubs of eukaryotic cells. By separating the internal environment from the rest of the cytoplasm, the organelle creates specialized zones for different biochemical reactions. The outer envelope serves as the interface with the cytoplasm, while the deeper layers handle intensive energy production.
Moving inward, the inner boundary membrane folds extensively to form distinct structures known as cristae. These intricate folds are joined at cristae junctions and dramatically increase the available surface area within mitochondria. This expanded surface is vital because it provides ample physical space to house the extensive enzymatic machinery required for energy conversion. Without this structural adaptation, mitochondria would lack the physical real estate needed to generate sufficient cellular power. It is a perfect example of form following function in cellular biology.
Deep within the innermost compartment lies the matrix space. This dense aqueous environment contains the unique genetic material of mitochondria, known as mitochondrial DNA. Unlike the linear chromosomes found in the cell nucleus, this genome is typically circular. Furthermore, the matrix houses dedicated mitoribosomes, which allow the organelle to synthesize a specific subset of its own proteins locally. The presence of these unique genetic and translation components highlights the distinct evolutionary history of mitochondria and their relative autonomy within the cell.
Finally, the inner membrane itself is packed with critical protein complexes. Students will notice electron transport chain components and large ATP synthase complexes embedded throughout this lipid bilayer. These structures work in a highly coordinated sequence to drive the biochemical reactions necessary for life. The strategic placement of these complexes along the cristae ensures that mitochondria can efficiently transfer electrons and synthesize chemical energy. This spatial organization is absolutely essential for maintaining the high metabolic output required by dynamic living tissues.
Slide 2: Anatomy, Cellular Distribution, and Plasticity of Mitochondria
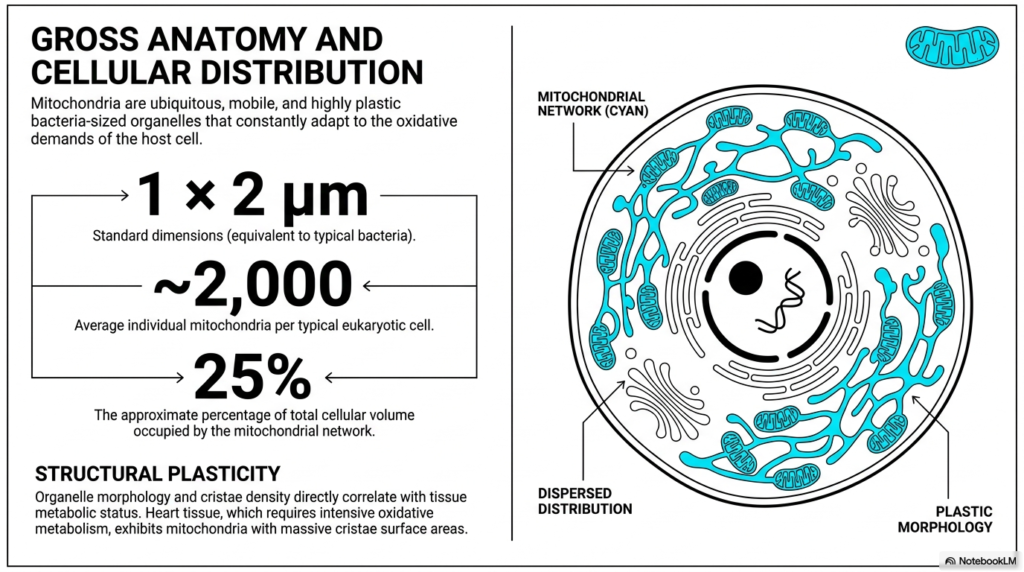
When examining the gross anatomy, it is fascinating to note that mitochondria share physical dimensions remarkably similar to those of typical bacteria. Measuring approximately one by two micrometers, these standard dimensions provide a baseline for understanding their scale. However, a single cell does not just rely on one or two of these powerhouses. An average eukaryotic cell contains approximately 2,000 mitochondria to meet its energy needs. This massive accumulation means the extensive mitochondrial network can occupy up to 25% of the total cellular volume.
The distribution of mitochondria within the cell is far from static or random. Instead of remaining anchored in one place, they form highly mobile networks that dynamically disperse throughout the cytoplasm. They continuously move to cellular regions where energy demand is the highest. This constant mobility ensures that ATP is delivered precisely where it is needed for cellular processes. By functioning as a dispersed, adaptable network rather than isolated units, mitochondria efficiently support the complex spatial energy requirements of large eukaryotic cells.
Perhaps the most remarkable physical trait of mitochondria is their structural plasticity. These organelles are not rigid structures; they actively change their internal morphology in response to the immediate needs of the host cell. The density of the internal cristae directly correlates with the metabolic status of the surrounding tissue. Tissues that undergo intensive oxidative metabolism require a much higher energy output and therefore trigger structural adaptations within the organelle to accommodate this demand.
A prime example of this structural adaptation is found in heart tissue. Because cardiac muscle requires continuous, relentless energy to maintain a heartbeat, the mitochondria within these cells exhibit a massive expansion of their cristae surface areas. This physical plasticity allows the organelle to pack in significantly more energy-producing protein complexes. Ultimately, mitochondria’s ability to physically adapt their internal architecture ensures they can continually meet fluctuating oxidative demands across different biological tissues.
Slide 3: Evolutionary Origins and Endosymbiont Theory of Mitochondria
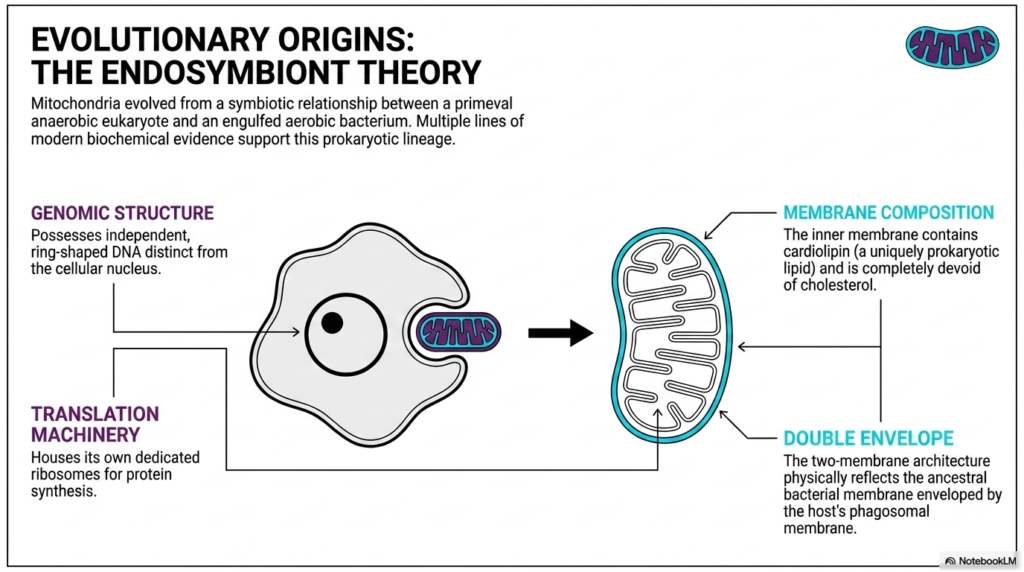
The evolutionary history of mitochondria is one of the most captivating stories in biology, perfectly explained by the endosymbiont theory. This scientific model proposes that these powerhouses evolved from a symbiotic relationship formed billions of years ago. A primeval anaerobic eukaryote engulfed a free-living aerobic bacterium, but instead of digesting it, the two formed a mutually beneficial partnership. Over countless generations, this engulfed bacterium became fully integrated into the host cell. Today, multiple lines of modern biochemical evidence strongly support the prokaryotic lineage of mitochondria.
One major piece of evidence lies in the genomic structure. Mitochondria possess an independent, ring-shaped DNA structure that is entirely distinct from the linear DNA stored inside the cellular nucleus. This circular genome is a classic hallmark of bacterial biology. Additionally, mitochondria house their own dedicated translation machinery, complete with specialized ribosomes for local protein synthesis. The fact that these organelles retain the ability to autonomously synthesize proteins strongly reflects the independent functionality of their ancient bacterial ancestors.
The membrane composition provides another fascinating clue to their ancient origins. The inner membrane of mitochondria contains cardiolipin, a uniquely prokaryotic lipid molecule. Crucially, this inner membrane is completely devoid of cholesterol, which is standard in eukaryotic membranes but absent in bacteria. This unique biochemical signature in the lipid profile strongly points back to a distinct bacterial heritage. It shows that while mitochondria have adapted to their host, they have preserved ancient biochemical remnants.
Finally, the defining physical architecture of mitochondria points directly to an endosymbiotic past. The distinct two-membrane double envelope physically reflects this ancient cellular merger. The inner membrane is derived from the ancestral bacterial membrane, while the outer envelope represents the host cell’s phagosomal membrane from the original engulfment event. Together, these combined biochemical and structural features make a compelling, undeniable case for how mitochondria transformed from independent microbes into vital cellular components.
Slide 4: The Genome and Genetic Streamlining of Mitochondria
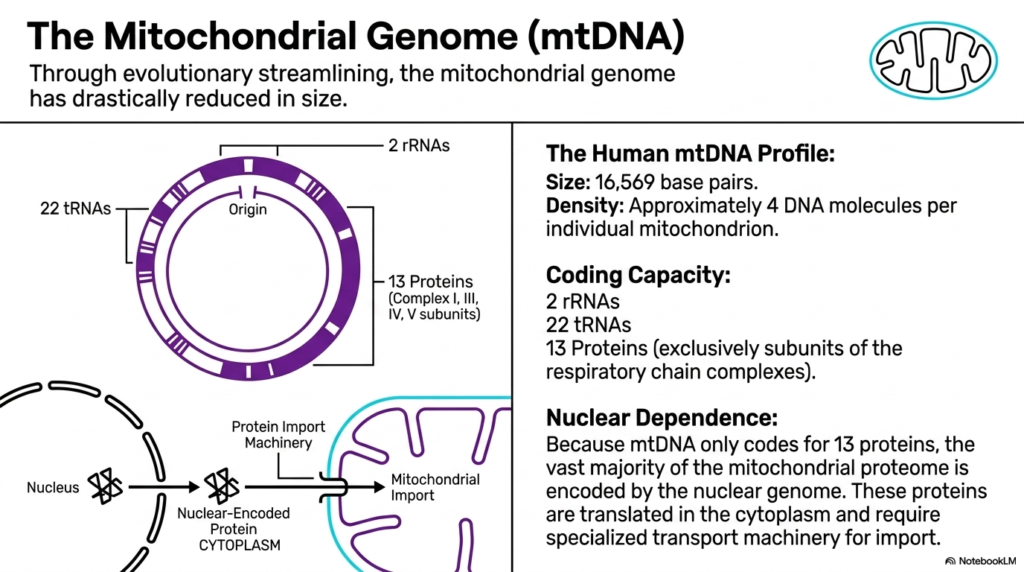
While these organelles contain their own genetic material, mitochondrial genomes have undergone dramatic evolutionary streamlining over millions of years. This process of genetic reduction means the mitochondrial genome is now incredibly compact compared to the massive genetic code housed in the nucleus. Through evolutionary history, many original bacterial genes were either lost or permanently transferred to the host cell’s nuclear DNA. Today, the genetic blueprint retained within mitochondria represents only the most essential components needed for localized cellular energy production.
To understand the specifics, scientists closely analyze the human mitochondrial DNA profile. This uniquely compact genome consists of precisely 16,569 base pairs arranged in a continuous loop. Despite this small size, there is a relatively high density of genetic material, with approximately 4 distinct DNA molecules per mitochondrion. This localized clustering ensures that mitochondria always have immediate access to the instructions required for synthesizing their most critical internal machinery.
The functional coding capacity of this streamlined genome is remarkably specialized. The DNA housed within mitochondria codes for only two ribosomal RNAs and twenty-two transfer RNAs. Most importantly, it encodes exactly thirteen specific proteins. Interestingly, all thirteen of these encoded proteins function exclusively as critical subunits for the respiratory chain complexes. This means the mitochondrial genome is entirely dedicated to supporting the physical structures that directly drive oxidative phosphorylation and essential energy generation.
Because this genome encodes only 13 proteins, mitochondria exhibit an enormous degree of nuclear dependence. The vast majority of the functional mitochondrial proteome is actually encoded by the cellular nuclear genome. These essential proteins must first be translated in the host cell’s cytoplasm. Afterward, they require highly specialized, dedicated transport machinery to cross the membranes and enter the mitochondria. This intricate interplay demonstrates how deeply integrated the organelle has become with its host.
Slide 5: Differential Permeability in the Membranes of Mitochondria
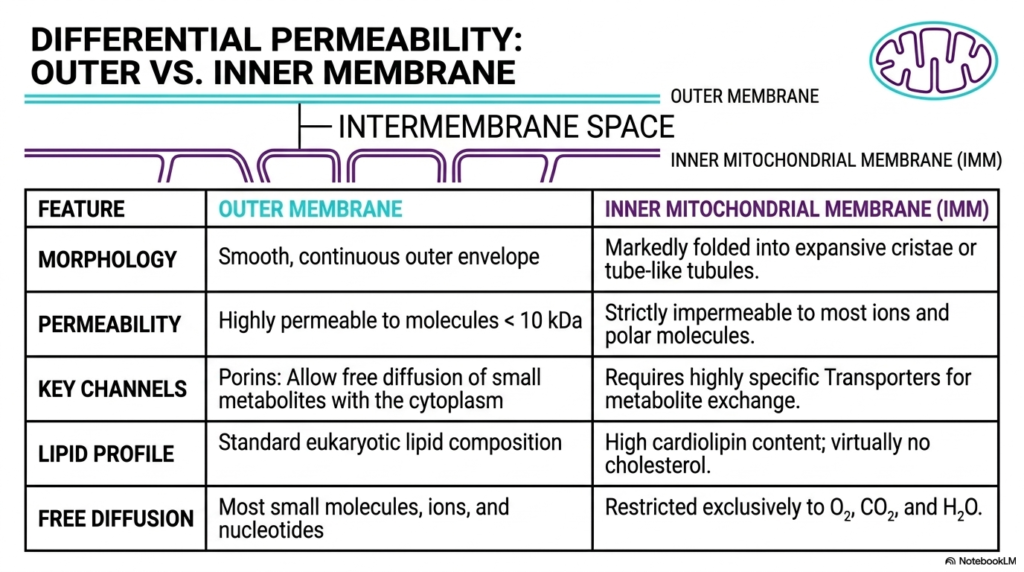
A core biochemical principle of mitochondria is the differential permeability of their two distinct boundary layers. The outer membrane and the inner mitochondrial membrane possess entirely different physical and chemical properties. This stark contrast allows the intermembrane space to serve as a functional intermediate zone. By carefully controlling what molecules can pass through which barrier, mitochondria can precisely regulate their internal environment and establish the essential biochemical gradients needed for cellular life.
The outer membrane functions as the initial protective envelope and features a smooth, continuous morphology. Biochemically, it shares a standard eukaryotic lipid composition, making it quite similar to other cellular structures. A defining feature is its high permeability to molecules smaller than ten kilodaltons. This is made possible by abundant pore-forming proteins called porins. These channels allow the free diffusion of small metabolites, ions, and nucleotides, meaning the outermost layer of mitochondria interacts very openly with the surrounding cytoplasm.
In sharp contrast, the inner mitochondrial membrane serves as an incredibly strict barrier. Morphologically, it is markedly folded into expansive cristae or tube-like tubules. This inner boundary is strictly impermeable to most ions and polar molecules. Because simple diffusion is blocked, the inner layer requires highly specific carrier proteins and transporters for any required metabolite exchange. This strict impermeability is the fundamental reason mitochondria can successfully trap protons and build a massive proton gradient.
The unique lipid profile of the inner membrane further enforces this differential permeability. As mentioned previously, it boasts a high cardiolipin content and is virtually devoid of cholesterol. Because of this specialized lipid composition, free diffusion across the inner boundary is restricted to essential, tiny molecules such as oxygen, carbon dioxide, and water. This extreme barrier function ensures that the complex energetic reactions occurring deep within mitochondria remain perfectly isolated and highly controlled at all times.
Slide 6: Functional Topography of the Inner Membrane in Mitochondria
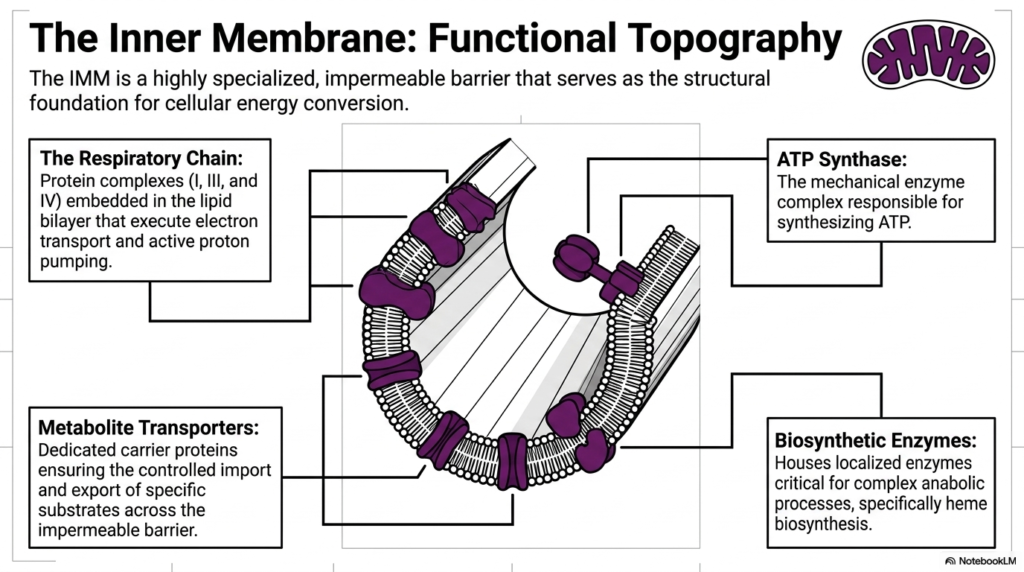
The functional topography of the inner mitochondrial membrane is arguably one of the most critical structural concepts in biochemistry. This highly specialized, completely impermeable barrier serves as the absolute structural foundation for cellular energy conversion. Because it blocks free diffusion, every single protein complex embedded within it serves a specific, vital role. The precise arrangement of these embedded components ensures that mitochondria operate with maximum biochemical efficiency, acting as microscopic molecular factories.
The most prominent feature of this membrane is the respiratory chain. This consists of massive protein assemblies, specifically Complexes I, III, and IV, firmly embedded within the lipid bilayer. These specific complexes are responsible for carrying out the electron transport process while simultaneously driving active proton pumping. By carefully passing electrons from one complex to the next, the inner membrane of mitochondria safely manages massive amounts of chemical energy without damaging the surrounding cellular environment.
Working directly downstream from the respiratory chain is the remarkable ATP Synthase complex. This structure functions as a literal mechanical enzyme that finally synthesizes cellular ATP. It acts as a molecular turbine, harvesting the energy gradient established by the other complexes. The strategic localization of ATP Synthase directly alongside the respiratory chain on the inner membrane allows mitochondria to instantly convert the generated physical gradients into usable, storable chemical energy for the cell.
Beyond direct energy generation, this highly specialized barrier houses other crucial functional components. Dedicated metabolite transporters act as specific carrier proteins, ensuring the carefully controlled import and export of specific substrates across the otherwise impermeable boundary. Additionally, the inner membrane contains localized biosynthetic enzymes that are absolutely critical for complex anabolic processes, particularly the intricate pathways of heme biosynthesis. Together, these elements make the inner membrane of mitochondria a dynamic and versatile biochemical hub.
Slide 7: The Matrix Space as the Metabolic Hub of Mitochondria
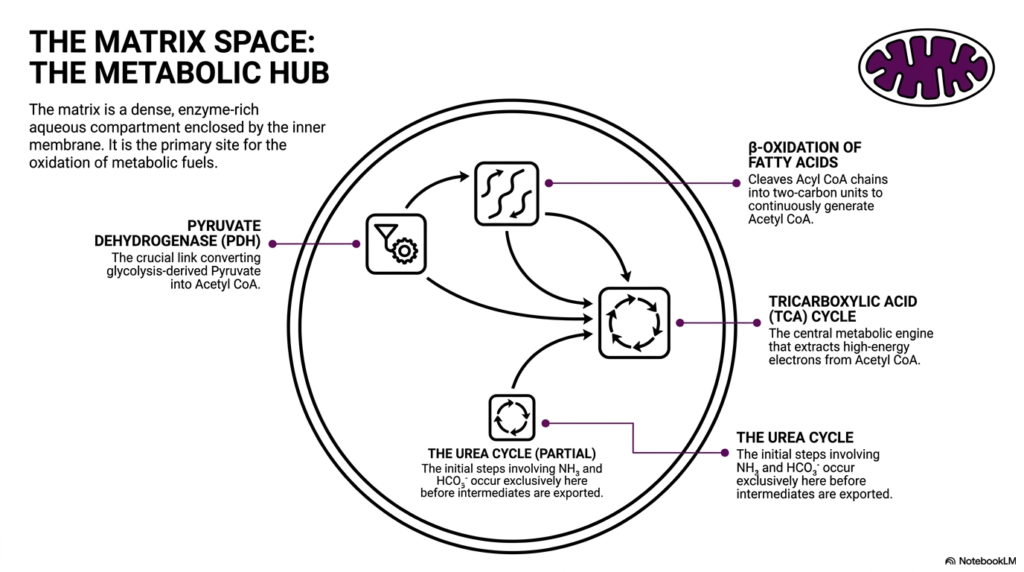
Deep inside the protective inner membrane lies the matrix space, which serves as the central metabolic hub of mitochondria. This internal region is an incredibly dense, enzyme-rich aqueous compartment that houses an array of critical biochemical pathways. It is the primary, dedicated site for the complete oxidation of incoming metabolic fuels. By centralizing these complex reactions within a single enclosed environment, mitochondria can rapidly process nutrients and extract maximum energy with remarkable efficiency.
A crucial player in this enclosed space is the pyruvate dehydrogenase complex. This massive enzyme serves as the absolutely crucial link connecting cytoplasmic glycolysis to mitochondrial energy production. It operates by converting glycolysis-derived pyruvate molecules directly into acetyl-CoA. This vital conversion step ensures that carbon substrates are properly prepared before they are fed into the deeper oxidative engines of mitochondria. Without this initial gateway step, cellular energy production would grind to a halt.
Once Acetyl CoA is generated, it directly enters the tricarboxylic acid cycle, widely known as the TCA cycle. This cycle is the true central metabolic engine operating inside mitochondria. Through a series of carefully orchestrated enzymatic reactions, the TCA cycle strips high-energy electrons away from acetyl-CoA. Simultaneously, another process called beta-oxidation continuously cleaves long fatty acid chains into two-carbon units, feeding even more Acetyl CoA into the cycle to ensure the engine never stops running.
Finally, the matrix space is not solely dedicated to energy extraction; it also plays a role in cellular waste management. The initial, vital steps of the urea cycle occur exclusively here before intermediate molecules are safely exported to the cytoplasm. These early steps involve safely managing ammonia and bicarbonate ions. By hosting these diverse yet interconnected pathways, the dense matrix space demonstrates why mitochondria are considered the incredibly versatile metabolic centers of all complex cellular life.
Slide 8: Oxidative Phosphorylation and Gradient Formation in Mitochondria
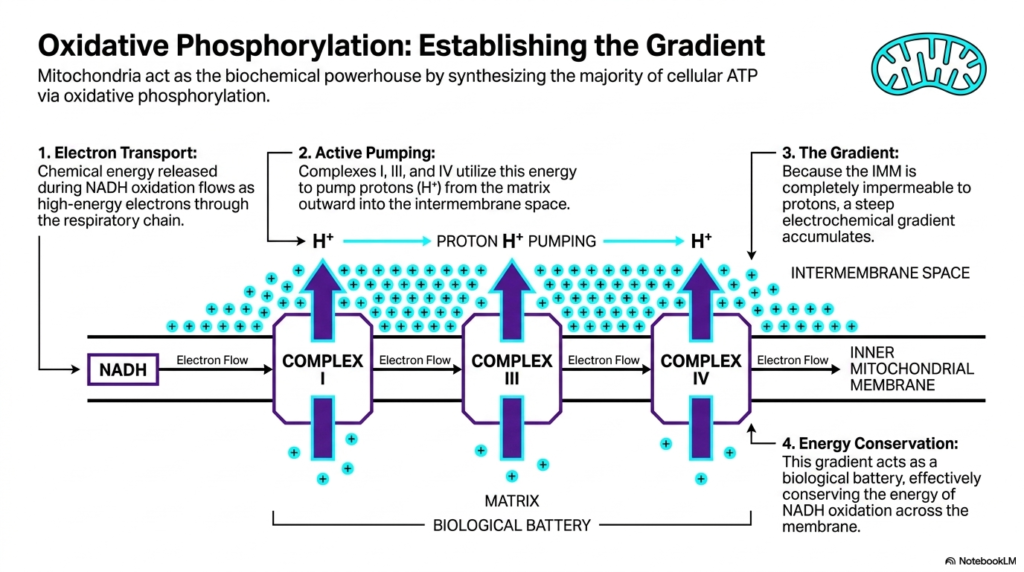
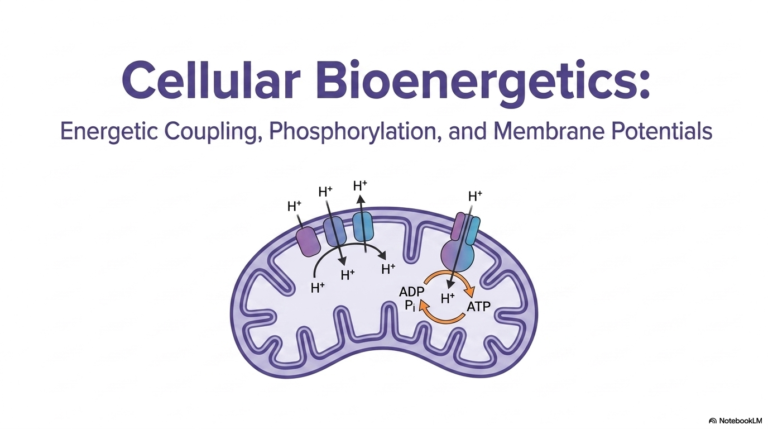
The primary mechanism establishing these organelles as the true biochemical powerhouse is oxidative phosphorylation. Mitochondria use this continuous, multi-step process to produce the vast majority of cellular ATP. This process begins with electron transport. Chemical energy, initially released during the oxidation of molecules like NADH, flows as a stream of high-energy electrons directly through the respiratory chain. This controlled flow of electrons acts as the necessary spark that ignites the entire energy conversion process.
As these high-energy electrons flow down the chain, they interact heavily with specific transmembrane proteins. Complexes I, III, and IV actively utilize this flowing electron energy to physically pump protons outward from the matrix. These positively charged hydrogen ions are actively pumped directly into the narrow intermembrane space. This continuous active pumping by the respiratory components of mitochondria creates a massive accumulation of protons on one side of the barrier, radically altering the local chemical environment.
Because the inner mitochondrial membrane is completely impermeable to protons, these pumped ions cannot simply diffuse back into the matrix space. As a direct result of this strict barrier, a tremendous, steep electrochemical gradient rapidly accumulates within the intermembrane space. This massive concentration of positively charged protons represents a huge reservoir of stored potential energy. The ability to successfully establish and maintain this steep gradient is the defining biochemical triumph of all functional mitochondria.
This trapped electrochemical gradient effectively acts as an organic biological battery. It is a brilliant method of energy conservation, physically conserving the rapidly released chemical energy of NADH oxidation by storing it directly across the inner membrane. Just as a physical battery stores electrical potential energy for later use, mitochondria store chemical potential energy in this proton gradient. This stored potential is now perfectly poised and ready to drive the final, crucial step of cellular energy synthesis.
Slide 9: Harvesting Gradient Energy via ATP Synthase in Mitochondria
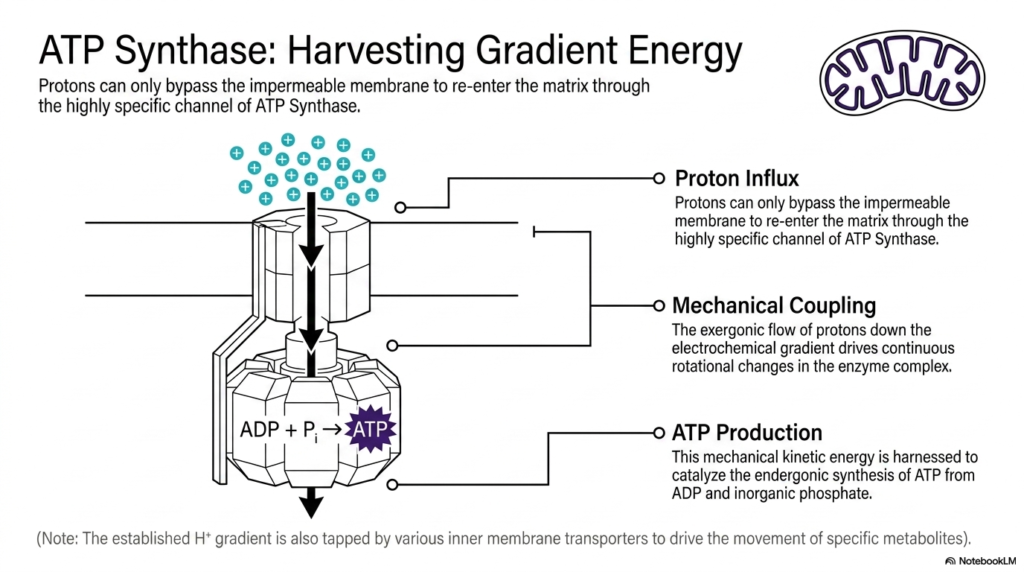
Having successfully established a massive biological battery, the next step is to harvest the stored gradient energy. Because the accumulated protons are desperate to return to the lower-concentration matrix, they seek a pathway across the barrier. However, these protons can bypass the strictly impermeable inner membrane of mitochondria only through a specific exit route. That singular, highly regulated channel is provided by the massive ATP Synthase enzyme complex, which spans the lipid bilayer.
As the positive protons rush through this specific channel to re-enter the matrix, an incredible physical phenomenon occurs. This massive proton influx results in direct mechanical coupling. The exergonic, downhill flow of protons rushing down their steep electrochemical gradient physically drives continuous, microscopic rotational changes within the enzyme complex itself. ATP Synthase literally spins like a microscopic cellular turbine, perfectly demonstrating how mitochondria transform raw chemical gradients directly into dynamic, kinetic mechanical energy.
This spinning mechanical kinetic energy is immediately put to crucial biochemical use. The rotating internal components of ATP Synthase physically force its active sites to change shape. This rapid physical movement is seamlessly harnessed to actively catalyze the endergonic synthesis of cellular ATP. By forcibly combining depleted ADP molecules with free inorganic phosphate, mitochondria continuously churn out fresh, usable energy molecules, effectively reloading the cell’s energy currency second by second.
It is also important to note that ATP production is not the sole use for this massive gradient. The established proton gradient is incredibly versatile and is also frequently tapped by various other inner membrane transporters. These secondary transporters use stored energy to actively transport specific metabolites into and out of the matrix space. This ensures that mitochondria can simultaneously power the cell while actively maintaining their own internal chemical balance.
Slide 10: Signaling and Cellular Regulation Roles of Mitochondria
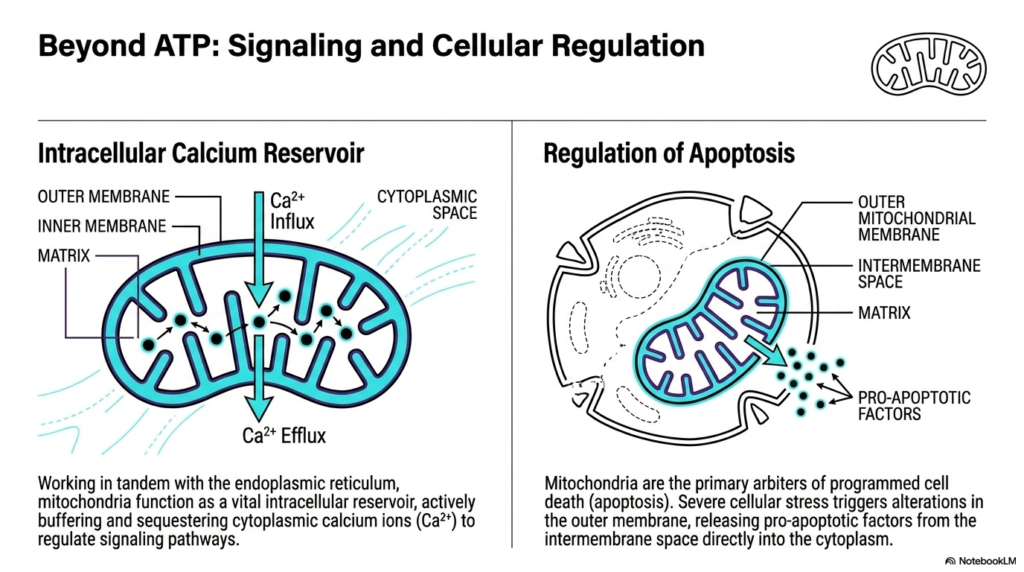
While famously known for energy production, the functional repertoire of these organelles extends far beyond simply making ATP. Mitochondria also play massive, vital roles in complex intracellular signaling and overall cellular regulation. One of their most critical secondary functions is acting as a dynamic intracellular calcium reservoir. By working in close tandem with the endoplasmic reticulum, mitochondria actively buffer the cytoplasm, carefully sequestering and releasing calcium ions to precisely regulate delicate cellular signaling pathways.
This precise calcium regulation requires constant, dynamic movement across the organelle’s membranes. Calcium ions undergo active influx from the cytoplasm, moving directly across the outer and inner membranes to be safely stored deep within the matrix. When the cell requires a specific signaling cascade, rapid calcium efflux occurs, releasing the ions back into the cellular environment. This buffering ability ensures that mitochondria act as crucial stabilizing forces, preventing toxic calcium overload within the living cell.
Furthermore, these dynamic organelles act as the primary, ultimate arbiters of programmed cell death, a critical biological process known as apoptosis. When a cell experiences severe, irreversible stress or massive damage, it must be safely dismantled to protect the surrounding healthy tissue. Severe cellular stress directly triggers highly specific structural alterations in the outer membrane of mitochondria. These precise structural changes are the absolute biological point of no return for a damaged cell.
Following these severe structural alterations, mitochondria initiate the final stages of the apoptotic cascade. They actively release deadly pro-apoptotic factors that were previously safely trapped within the intermembrane space into the open cytoplasm. The sudden rush of these factors rapidly activates the cellular demolition machinery. By carefully controlling this dramatic, lethal release, mitochondria ensure that highly damaged or dangerously mutated cells are swiftly and safely removed from the biological system.
Slide 11: Master Biochemical Localization Map of Mitochondria
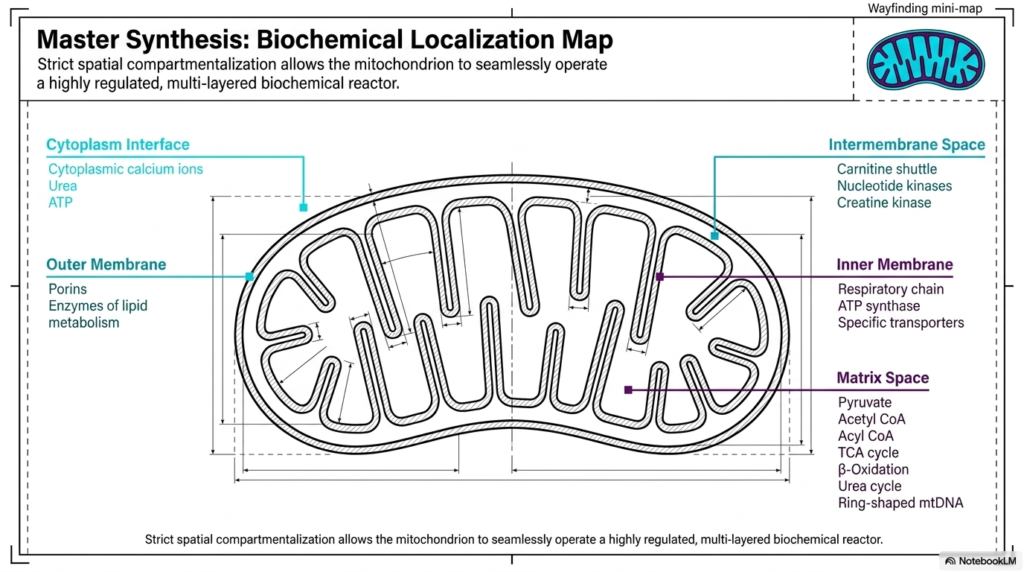
To fully appreciate the complexity of these organelles, it is essential to review a master biochemical localization map. This comprehensive overview highlights how strict spatial compartmentalization enables mitochondria to operate seamlessly as a highly regulated, multi-layered biochemical reactor. By permanently isolating distinct reactions into very specific physical zones, the organelle prevents chemical interference and maximizes total metabolic efficiency. Every metabolic process has a specific, assigned geographic location within the complex mitochondrial architecture.
Starting at the outermost layer, the cytoplasmic interface constantly exchanges critical elements such as cytoplasmic calcium ions, urea, and freshly synthesized ATP. Moving inward, the outer membrane physically houses the porins, along with specific enzymes dedicated to localized lipid metabolism. Just inside this boundary, the fluid intermembrane space holds specialized shuttles, including the carnitine shuttle, as well as crucial nucleotide kinases and creatine kinase. This specific layering allows mitochondria to effectively buffer incoming and outgoing chemical traffic.
Deeper still, the inner membrane serves as the heavily fortified core of energy production. This strict barrier permanently houses the mighty respiratory chain, the mechanical ATP synthase turbines, and countless highly specific transporters. Finally, the innermost matrix space operates as the ultimate chemical furnace. This dense zone contains exclusively pyruvate, acetyl-CoA, the entire TCA cycle, beta-oxidation enzymes, the initial steps of the urea cycle, and the specialized ring-shaped genetic material of mitochondria.
In conclusion, this master synthesis perfectly illustrates the sheer biochemical elegance of the organelle. The strict, dedicated spatial compartmentalization ensures that numerous distinct, massive chemical pathways can occur simultaneously without causing internal cellular chaos. By acting as highly regulated, seamlessly operating biological reactors, mitochondria provide the constant, massive energy foundation required to sustain the complex, dynamic nature of all advanced eukaryotic life.
Please read our Content Disclaimer Statement.
Check out our social media channels: