72. Plasma Membrane Transport Processes: Active and Passive Transport
Consider a medieval castle without doors, gates, or guards; it would quickly fall to invaders or starve from a lack of supplies. The biological cell faces a similar fundamental challenge, relying entirely on the intricate machinery of the lipid bilayer to survive and thrive. This slide deck explores the foundational mechanisms of cellular permeability, detailing how molecules move into and out of the cell. Understanding these physiological processes is critical for grasping how cells maintain their delicate internal balance amidst a constantly changing external environment.
Slide 1: Introduction to Plasma Membrane Transport: The Cell’s Gatekeeping System
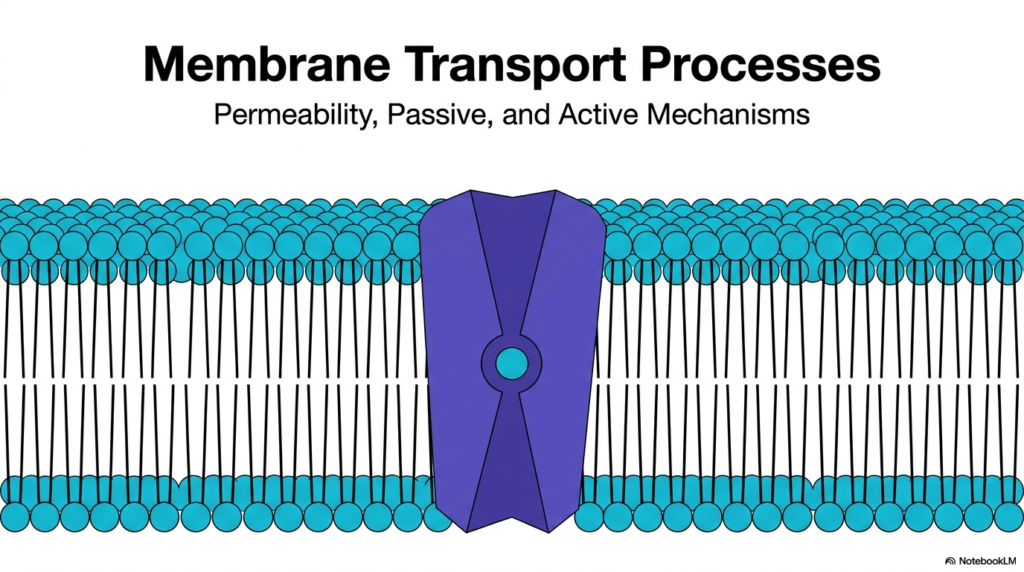
The overarching theme of these slides is Plasma Membrane Transport, a fundamental concept in biochemistry and cellular physiology. Plasma Membrane Transport dictates exactly which substances can cross the cellular barrier, acting as the ultimate gatekeeper for the organism. By controlling cellular permeability, the cell ensures that essential nutrients enter, toxic waste products exit, and vital ion gradients are preserved. Without a highly regulated system for Plasma Membrane Transport, the cell’s internal environment would rapidly equilibrate with its surroundings, leading to immediate cellular death.
The featured image showcases the classic phospholipid bilayer punctuated by a massive purple transport protein. This visual perfectly encapsulates the dual nature of Plasma Membrane Transport. The lipid bilayer provides a formidable hydrophobic barrier, while the embedded proteins act as selective channels and pumps. Throughout this educational journey, medical and biology students will discover the diverse passive and active mechanisms that govern the transport of these molecules. Recognizing how these structures function offers profound insights into pharmacology, human physiology, and the molecular basis of disease.
As students delve deeper into this deck, the complexity of Plasma Membrane Transport will become clear. The cell does not rely on a single, uniform method to move varied cargo. Instead, a sophisticated array of mechanisms exists, ranging from simple diffusion to energy-demanding active transport. By mastering the dynamics of Plasma Membrane Transport, students will build a strong foundation for understanding complex biochemical pathways. This knowledge is essential for future medical professionals who will manipulate these pathways to treat metabolic disorders.
Slide 2: Rules of Permeability in Plasma Membrane Transport: Who Gets In?
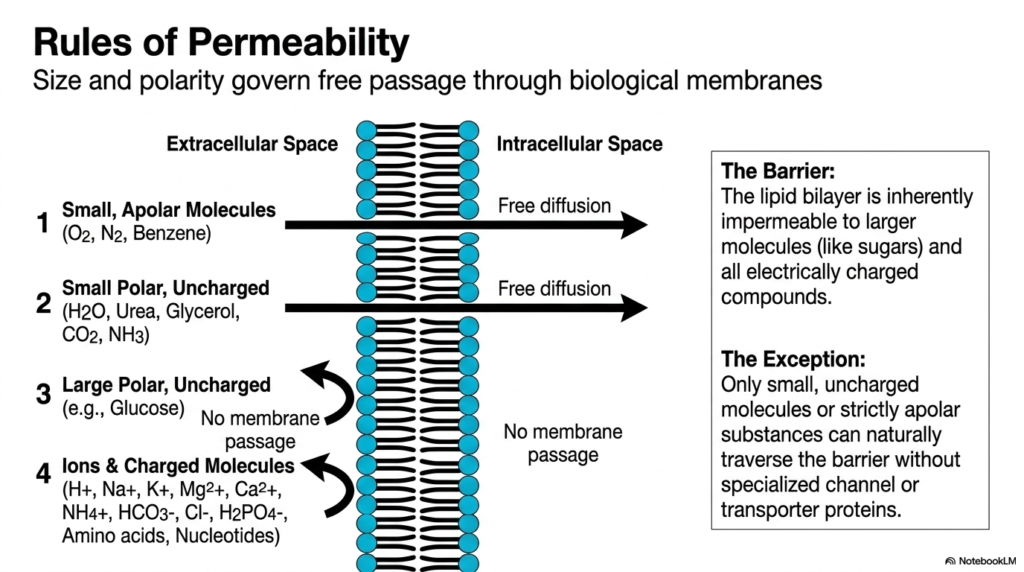
When mixing oil and water, a classic chemical resistance keeps the substances apart. This same physical resistance keeps biological cells intact. This slide breaks down the strict physical and chemical rules governing cellular permeability. The primary purpose is to establish why certain molecules can slip through the cellular boundary effortlessly while others are completely blocked. Understanding these basic permeability rules is essential for predicting how different pharmaceutical drugs, environmental nutrients, and respiratory gases will behave when they encounter the cell’s rigid outer boundary.
The fundamental rule of Plasma Membrane Transport relies on two chemical properties: molecular size and polarity. Small, apolar molecules like oxygen, nitrogen, and benzene possess the ultimate biological pass. Because they share the hydrophobic core of the lipid bilayer, they diffuse freely. Similarly, small, uncharged polar molecules, such as water, urea, and carbon dioxide, can cross the barrier. In these specific instances, Plasma Membrane Transport occurs spontaneously without the need for any specialized protein assistance or metabolic energy input.
However, the barrier is notoriously unforgiving to other biological substances. Large polar uncharged molecules, like glucose, simply cannot pass through the lipid bilayer on their own. Their size prevents them from squeezing between the densely packed lipid tails. Furthermore, all ions and electrically charged molecules are completely rejected. Ions like sodium, potassium, and calcium possess strong hydration shells and electrical charges that render simple Plasma Membrane Transport impossible. For these vital molecules, the structural membrane is inherently and strictly impermeable.
This is where the exceptions to the rules of Plasma Membrane Transport become biologically fascinating. Because the cell desperately needs glucose for energy and ions for signaling, it must employ specialized channel and transporter proteins. These integral proteins bypass the strict lipid barrier, creating selective pathways for large or charged cargo. By memorizing these fundamental rules of Plasma Membrane Transport, biochemistry students can easily deduce which biological molecules require protein assistance and which can cross biological membranes independently.
Slide 3: Free Diffusion in Plasma Membrane Transport: The Path of Least Resistance
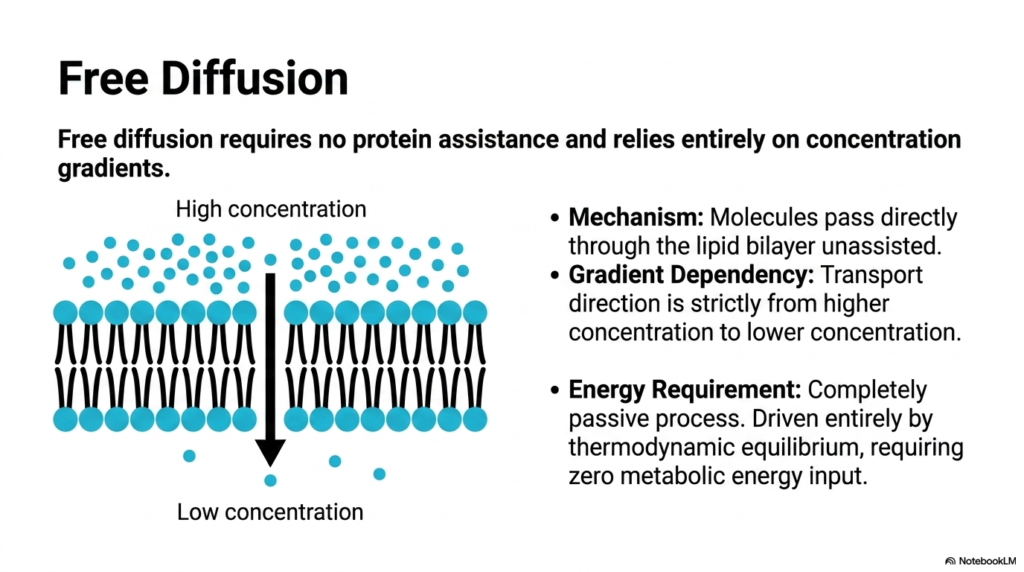
Observe a crowded room where individuals naturally spread out into empty spaces to find breathing room. Molecules in a solution behave exactly the same way. This slide focuses on free diffusion, the absolute simplest form of cellular movement. The core purpose is to illustrate how concentration gradients independently drive the movement of compatible molecules across the cell barrier. Grasping this basic thermodynamic principle is absolutely crucial before diving into more complex, protein-mediated cellular mechanisms that require metabolic energy.
In the context of Plasma Membrane Transport, free diffusion is a passive process that requires no metabolic energy. Molecules move directly through the lipid bilayer unassisted, bypassing the need for transport proteins. The sole driving force behind this specific form of Plasma Membrane Transport is the concentration gradient. Cargo transport is strictly directional, flowing downhill from an area of higher concentration to an area of lower concentration. It is a biological system driven entirely by the relentless pursuit of thermodynamic equilibrium.
Because it lacks protein mediators, free diffusion is unique among Plasma Membrane Transport mechanisms. The rate of this specific diffusion is directly proportional to the concentration gradient and the relative hydrophobicity of the moving molecule. Oxygen entering a muscle cell or carbon dioxide exiting a red blood cell is a classic example. As long as a gradient exists, this passive Plasma Membrane Transport will continue relentlessly until the chemical concentrations on both sides of the cellular barrier are perfectly equalized.
Understanding this passive form of Plasma Membrane Transport provides a strong baseline for general cellular physiology. Cells rely heavily on free diffusion for rapid gas exchange during cellular respiration. Without this instantaneous, unassisted movement, human tissues would quickly suffocate. However, because free diffusion is limited only to small, uncharged, or nonpolar molecules, it highlights the cell’s ongoing need for more sophisticated machinery. This limitation sets the perfect stage for exploring the diverse protein-assisted systems that sustain complex biological life.
Slide 4: The Electrochemical Gradient in Plasma Membrane Transport: The Dual-Force Engine
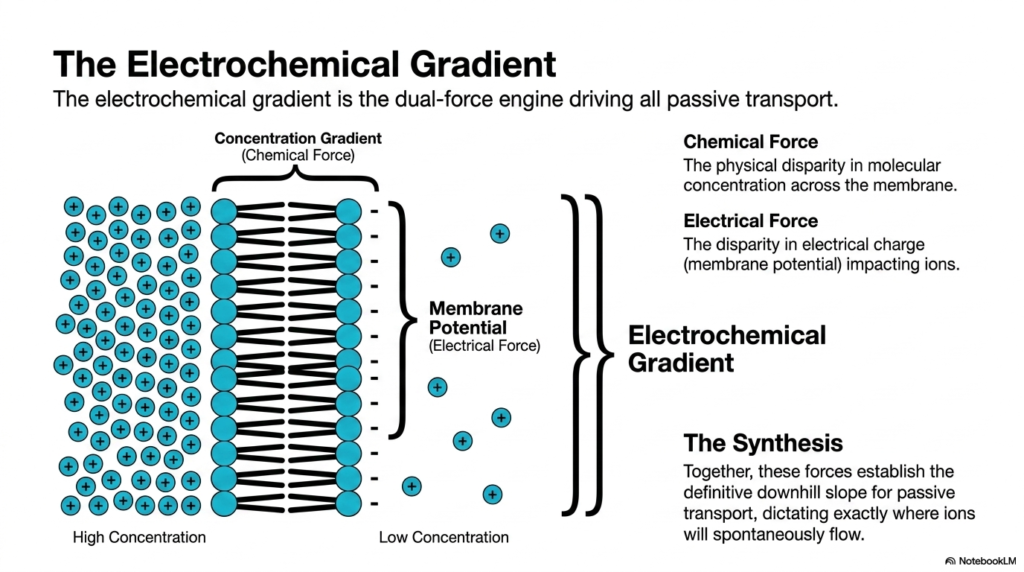
Think of water flowing down a mountain; gravity forces it downward, while the shape of the riverbed directs its path. In cellular biology, ions experience a similar irresistible physical pull. This slide explains the electrochemical gradient, the vital dual-force engine driving all passive cellular movement. The specific purpose of this slide is to dissect the distinct chemical and electrical forces that combine to dictate exactly where and how fast ionic molecules will spontaneously flow across the cellular boundary.
When discussing the movement of ions, Plasma Membrane Transport cannot rely on simple concentration gradients alone. Instead, it is governed by the electrochemical gradient, which consists of two distinct components. The first is the chemical force, which represents the physical disparity in molecular concentration across the membrane. The second is the electrical force, or membrane potential, which represents the significant disparity in electrical charge. In passive Plasma Membrane Transport, these two fundamental forces interact to determine the net movement.
The synthesis of these dual forces establishes the definitive downhill slope for passive Plasma Membrane Transport. If a cell has a negative internal charge, positive ions will be electrically pulled inward, even if their internal concentration is already high. Conversely, the concentration gradient might push an ion outward while the electrical gradient pulls it inward. The true direction of Plasma Membrane Transport is determined by whichever physical force is mathematically stronger, creating a highly dynamic cellular environment.
Mastering the electrochemical gradient is non-negotiable for students studying Plasma Membrane Transport. This dual-force engine is directly responsible for critical physiological events, including the rapid firing of action potentials in neurons and the contraction of muscle fibers. By manipulating these gradients, cells can store vital potential energy, functioning much like a chemical battery. This stored energy is eventually harnessed to drive other essential biological processes, proving that Plasma Membrane Transport regulates fundamental cellular bioenergetics and human physiological functions.
Slide 5: Facilitated Diffusion in Plasma Membrane Transport: The Cellular Escorts
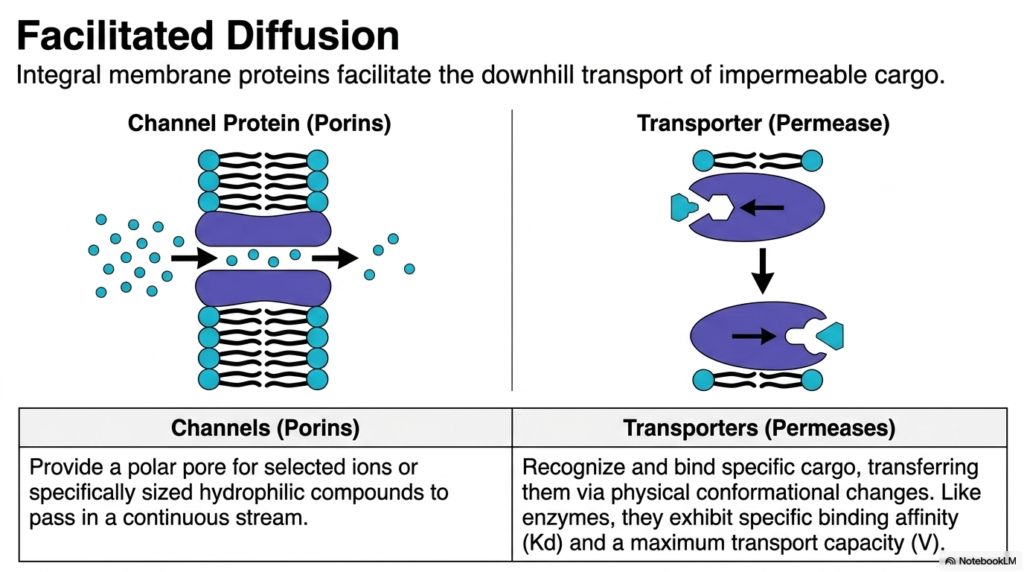
Consider an exclusive building; without a security badge, the heavy doors remain shut, but a designated escort can walk a guest right through. Cells use similar escorts to move challenging biological molecules. This slide introduces facilitated diffusion, highlighting the integral membrane proteins that enable the downhill transport of impermeable cargo. The objective is to differentiate between distinct channel proteins and transporters, illustrating how they selectively guide essential nutrients and ions safely across the dense cellular boundary.
When molecules are too large or charged to cross the plasma membrane independently, Plasma Membrane Transport relies on facilitated diffusion. This process uses specific integral proteins to transport molecules down their concentration gradients. Because it follows the downhill flow, this form of Plasma Membrane Transport remains passive and requires no cellular energy. The proteins involved in facilitated Plasma Membrane Transport are broadly categorized into two distinct structural classes: open channels, also known as porins, and shifting transporters, frequently referred to as permeases.
Channels provide a specialized polar pore straight through the lipid bilayer. They are designed for selected ions or specific hydrophilic compounds, allowing them to pass through in a rapid, continuous stream. In contrast, transporters execute Plasma Membrane Transport through physical conformational changes. A transporter must physically recognize and bind its specific chemical cargo. Once bound, the membrane protein shifts its shape, carrying the molecule to the opposite side. This mechanism ensures that only the correct biological molecules gain entry.
Transporters involved in Plasma Membrane Transport function remarkably like metabolic enzymes. They exhibit specific binding affinities and possess a strict maximum transport capacity. Once all the transporter proteins are fully occupied, the rate of Plasma Membrane Transport plateaus, regardless of how steep the concentration gradient becomes. This saturation point is a critical biochemical concept that separates facilitated transport from simple free diffusion. Understanding these protein dynamics is absolutely essential for designing targeted pharmaceutical drugs that modulate cellular uptake.
Slide 6: Active Transport in Plasma Membrane Transport: Pushing Uphill
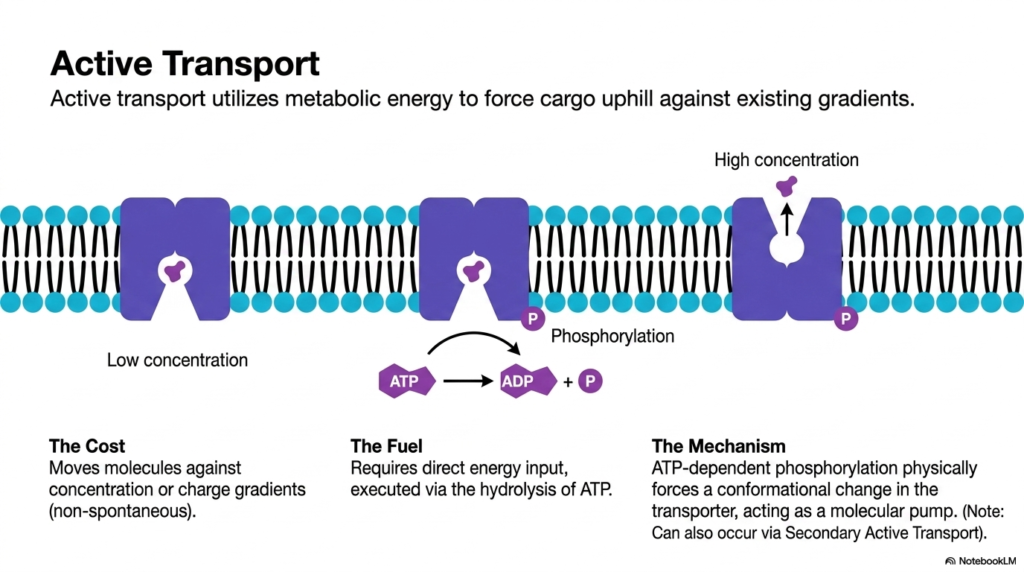
Pushing a heavy boulder up a steep hill requires a massive amount of physical exertion and energy. Pushing biological molecules against their natural gradients requires the same level of commitment from the cell. This slide defines active transport and demonstrates how metabolic energy is used to force cargo uphill. The core purpose is to explain the mechanical role of ATP in driving non-spontaneous molecular movement, a biochemical process absolutely critical for establishing the vital ion gradients necessary for complex biological life.
Unlike passive mechanisms, active Plasma Membrane Transport moves molecules against existing concentration or charge gradients. Because this cellular movement is thermodynamically non-spontaneous, the cell must pay a steep energetic cost. The standard fuel for primary active Plasma Membrane Transport is adenosine triphosphate (ATP). This direct energy input allows cells to stockpile specific nutrients and expel toxic cellular waste products. Without active Plasma Membrane Transport, essential internal gradients would vanish, severely crippling basic cellular function.
The mechanism behind this energetically demanding Plasma Membrane Transport relies on a biochemical process called phosphorylation. An ATP molecule is hydrolyzed into ADP and an inorganic phosphate group. This specific phosphate group binds directly to the transport protein. The addition of this bulky, negatively charged group forces a dramatic conformational change within the integral transporter. Acting as a powerful molecular pump, this shifting protein mechanically pushes the bound cargo across the barrier, executing active Plasma Membrane Transport.
This ATP-dependent form of Plasma Membrane Transport is responsible for maintaining the defining characteristics of a living biological cell. For example, sodium-potassium pumps work constantly to keep sodium levels low inside the cell while keeping potassium levels high. This uphill battle consumes a massive portion of the human body’s daily metabolic energy. By studying active Plasma Membrane Transport, medical students gain crucial insights into how cellular energy failure, such as during oxygen deprivation, rapidly leads to catastrophic tissue damage.
Slide 7: Transport Configurations in Plasma Membrane Transport: Organizing Cellular Traffic
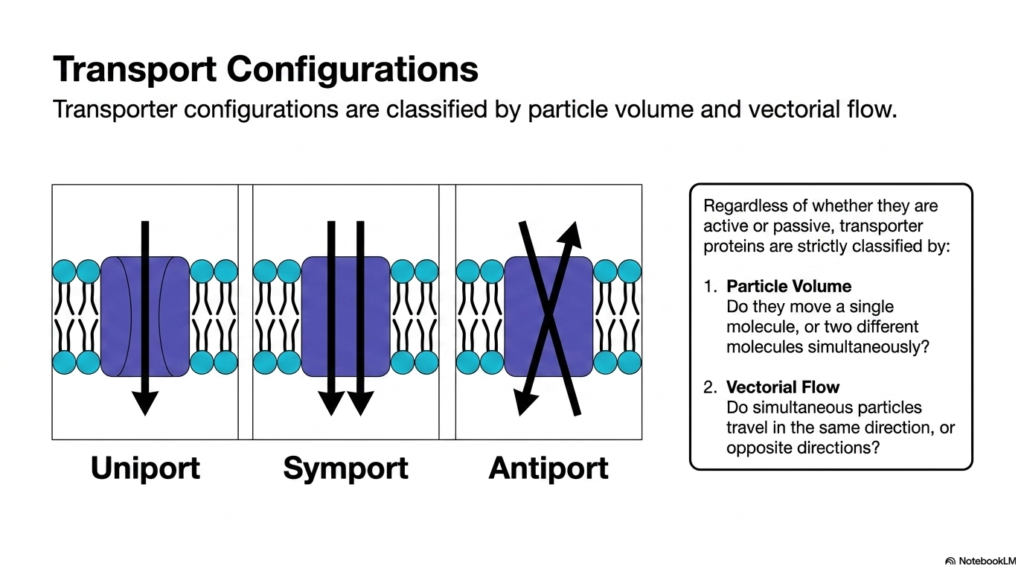
Managing a bustling city intersection requires clear rules about which cars can turn, go straight, or move together. The cell manages its molecular traffic with similar precision. This slide outlines the primary transport configurations used by biological membranes. The main goal is to classify transporter proteins by particle volume and vectorial flow, providing biochemistry students with a standardized framework for understanding how diverse molecules share cellular pathways without causing widespread biochemical congestion or systemic physiological failure.
Regardless of whether they are active or passive, proteins involved in Plasma Membrane Transport are strictly classified by two operational metrics. The first metric is particle volume, which asks a simple biological question: does the transport protein move a single molecular species, or does it move two different molecules simultaneously? The second metric governing Plasma Membrane Transport is vectorial flow. When multiple particles are involved, do they travel in the same direction, or do they move in entirely opposite directions?
By applying these two defining parameters, biochemists categorize Plasma Membrane Transport into three distinct structural systems: uniport, symport, and antiport. Uniport systems handle one specific molecule at a time. Symport systems couple two different molecules that move in the same direction. Antiport systems couple two distinct molecules moving in opposite directions. These classifications of Plasma Membrane Transport are universally recognized, appearing in organisms from simple bacteria to complex human tissues. They represent the fundamental logistical strategies that cells use daily.
Understanding these specific configurations of Plasma Membrane Transport is vital for making sense of complex metabolic pathways. In many physiological scenarios, the movement of one essential cellular nutrient is intricately linked to the movement of an abundant ion. By studying these linked Plasma Membrane Transport configurations, students can accurately map out vast cellular networks. Recognizing the functional difference between a symporter and an antiporter is the essential first step in diagnosing specific genetic transport defects and renal diseases.
Slide 8: Uniport Mechanisms in Plasma Membrane Transport: The Solo Travelers
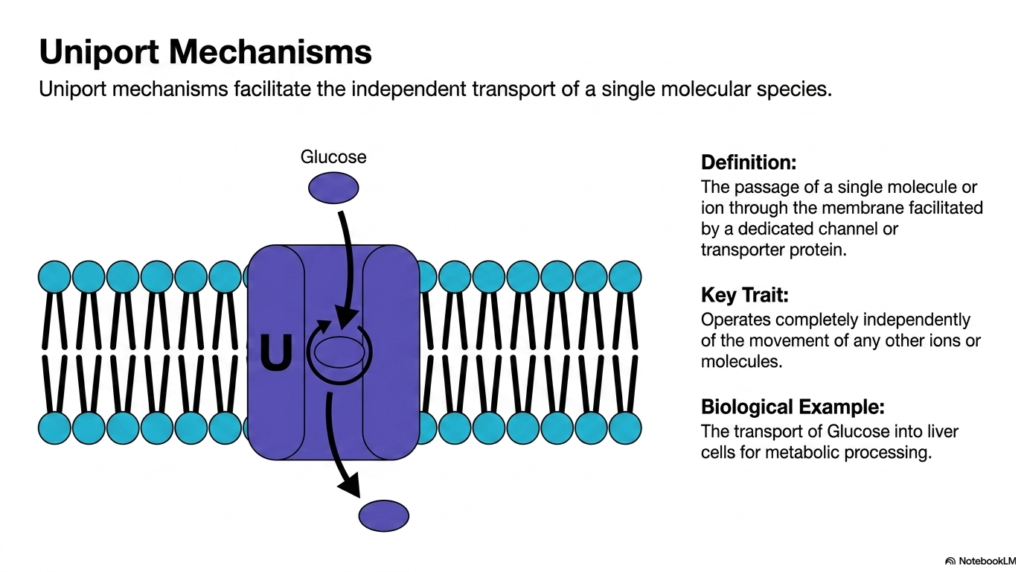
Think of an exclusive single-occupancy elevator moving passengers one at a time between floors. Uniport mechanisms operate with the exact same singular focus. This slide explores the basic uniport configuration, detailing the independent transport of a single molecular species. The clear objective is to define this straightforward biological mechanism and highlight its unique operational independence compared to more complex, coupled transport systems. Understanding uniport systems provides a clear foundation for analyzing targeted cellular nutrient acquisition and basic metabolic sustenance.
A specific uniport mechanism is defined as the passage of a single molecule or ion facilitated by a dedicated channel or transporter protein. In the vast biological landscape of Plasma Membrane Transport, uniport systems are the dedicated solo travelers. The defining key trait of this specific Plasma Membrane Transport configuration is its strict operational independence. The movement of the primary cellular cargo occurs without any reliance whatsoever on the simultaneous movement of other biological ions across the cellular boundary.
Because it operates completely independently, uniport Plasma Membrane Transport is highly specialized and precise. The dedicated transport protein undergoes conformational changes triggered solely by its specific chemical substrate. A classic biological example of this Plasma Membrane Transport is the vital movement of glucose into human liver cells. When systemic blood sugar levels rise, specialized uniport proteins bind to the abundant extracellular glucose and transport it into the cell for metabolic processing, relying entirely on the existing glucose concentration gradient.
Studying uniport systems provides an excellent physiological baseline for understanding more complex forms of Plasma Membrane Transport. These independent biological transporters are crucial for maintaining basal metabolic rates and ensuring cells receive steady supplies of basic energy substrates. When a uniport Plasma Membrane Transport system fails due to a genetic mutation, the specific cell loses access to that molecule, leading to targeted metabolic starvation. Furthermore, uniport proteins remain frequent targets for modern pharmacological intervention and sophisticated drug design.
Slide 9: Symport Mechanisms in Plasma Membrane Transport: The Carpool System
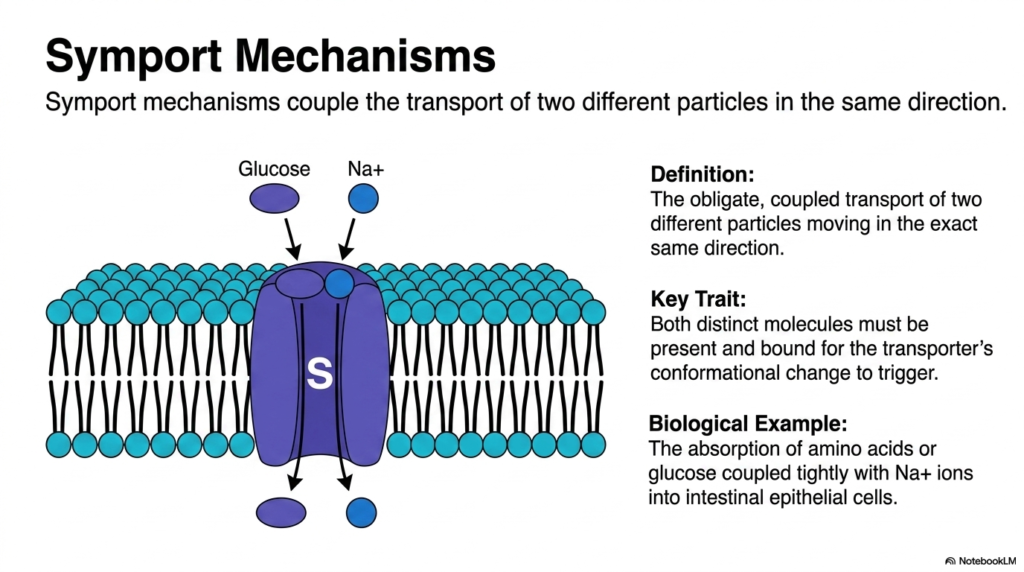
Consider a buddy system in which two individuals must hold hands to enter a restricted event; if one is missing, neither gets in. Symport mechanisms enforce a similar strict coupling. This slide breaks down symport systems, illustrating how dedicated transporters couple the movement of two different particles in the same direction. The purpose is to explain the significant biological advantages of linking molecular movements and how cells ingeniously exploit one strong gradient to move another reluctant molecule against its will.
Symport mechanisms represent a highly coordinated and efficient form of Plasma Membrane Transport. The standard biochemical definition describes the obligate, coupled transport of two different biological particles moving in the exact same direction. The key trait of this specific Plasma Membrane Transport is absolute structural dependency. Both distinct chemical molecules must be present and correctly bound to the transporter protein before any conformational change is triggered. If either cargo is absent, this form of Plasma Membrane Transport halts entirely.
This type of coupled Plasma Membrane Transport is biologically ingenious and incredibly efficient. Cells frequently use the strong, downhill concentration gradient of an ion to drag a different molecule uphill against its own natural gradient. A prime biological example of this Plasma Membrane Transport is the critical absorption of vital nutrients in the human gut. The transport of dietary amino acids or glucose is tightly coupled with sodium ions moving aggressively into intestinal epithelial cells, providing an essential driving force.
By utilizing dedicated symporters, cells maximize their energetic efficiency during intricate Plasma Membrane Transport. Instead of spending precious cellular ATP to move dietary glucose uphill, the cell simply piggybacks the sugar onto a pre-existing strong sodium gradient. Grasping this specific Plasma Membrane Transport strategy is undeniably essential for medical students. It perfectly explains the physiological basis of oral rehydration therapies, in which providing patients with both sodium and glucose simultaneously exploits these symporters to rapidly restore life-saving electrolyte balance.
Slide 10: Antiport Mechanisms in Plasma Membrane Transport: The Revolving Door
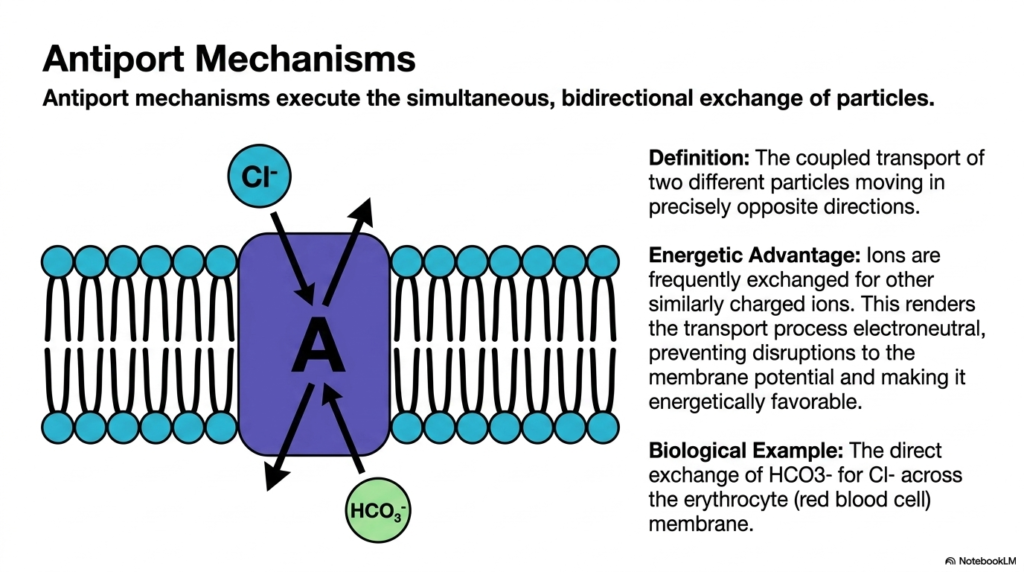
Consider a revolving door at a busy hotel; as one guest enters, another is simultaneously pushed outside. Antiport mechanisms operate as the exact biological equivalent of this exchange. This slide examines the antiport configuration, focusing on the simultaneous, bidirectional exchange of cellular particles. The main physiological goal is to highlight the specific energetic advantages of swapping complex ions and how this highly specific exchange system prevents dangerous disruptions to the biological cell’s extremely delicate electrical membrane balance.
Antiport mechanisms execute a fascinating and remarkably complex form of Plasma Membrane Transport. The standard definition of an antiporter involves the obligate coupled transport of two different biological particles moving in precisely opposite directions. Like its symport counterpart, this specific type of Plasma Membrane Transport is strictly dependent on both molecules. The outward cellular movement of one specific molecule is mechanically linked to the inward movement of another. This bidirectional Plasma Membrane Transport ensures a balanced exchange of resources.
There is a distinct, measurable energetic advantage to using these antiport systems for Plasma Membrane Transport. Charged ions are frequently exchanged for other similarly charged biological ions. This strategic cellular swap renders the overall transport process functionally electroneutral. By preventing massive shifts in cellular electrical charge, electroneutral Plasma Membrane Transport prevents dangerous disruptions to the resting membrane potential. Consequently, the exchange process is highly energetically favorable, allowing cells to regulate their internal chemistry without wasting excessive metabolic energy.
A classic biological example of this Plasma Membrane Transport is the precise exchange of bicarbonate for chloride across the erythrocyte membrane. This specific chemical exchange allows red blood cells to transport human carbon dioxide efficiently without altering their internal electrical charge. Understanding this bidirectional Plasma Membrane Transport is absolutely crucial for comprehending acid-base balance and systemic respiratory physiology. For medical students, mastering antiport systems unlocks the mechanical reasoning behind how the human body maintains a strictly maintained systemic blood pH.
Slide 11: The Diagnostic Matrix of Plasma Membrane Transport: Synthesizing Cellular Logistics
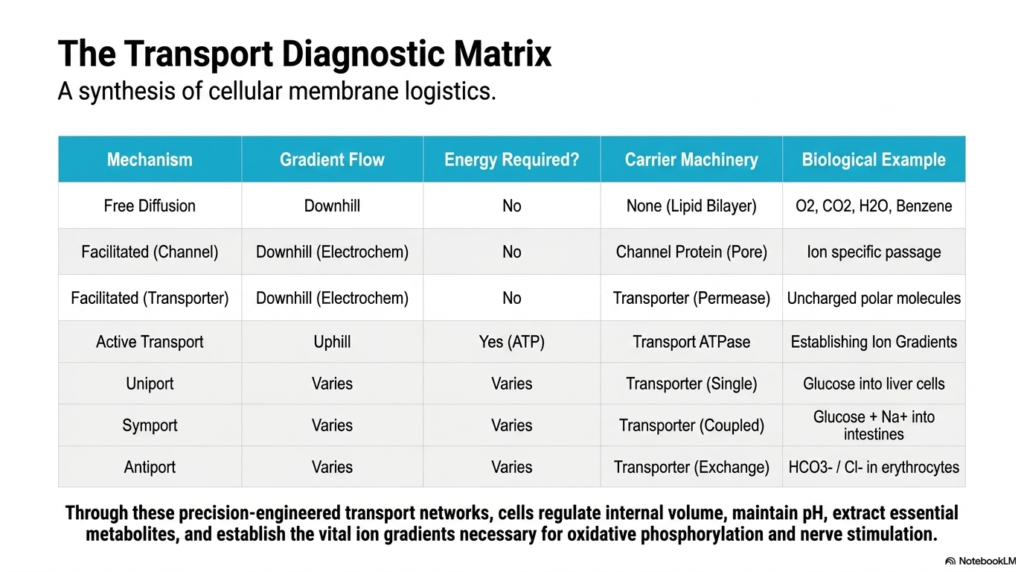
Navigating a complex subway map requires a clear legend summarizing all the different routes and rules. The final slide provides that exact legend for complicated cellular movement. This slide presents the Transport Diagnostic Matrix, a comprehensive synthesis of cellular membrane logistics. The core purpose is to consolidate all the previous physiological transport mechanisms into a single, easily digestible academic framework, allowing biochemistry students to rapidly compare gradient flows, chemical energy requirements, and structural carrier machinery at a single glance.
This diagnostic matrix serves as the ultimate study guide for mastering complex Plasma Membrane Transport. It summarizes how simple free diffusion functions as a downhill process that requires no metabolic energy and no carrier machinery. In sharp contrast, it details how facilitated Plasma Membrane Transport utilizes specific integral channel and transporter proteins to move polar or uncharged molecules downhill. Furthermore, the detailed table clearly distinguishes active Plasma Membrane Transport, emphasizing its unique requirement for ATP to push molecules uphill.
The matrix also outlines the distinct protein carrier configurations essential to Plasma Membrane Transport. It meticulously details the operational differences between the single-molecule uniport, the co-directional symport, and the bidirectional antiport systems. By categorizing these various forms of Plasma Membrane Transport, the summary matrix provides a structural overview of overall cellular logistics. Students can use this tool to rapidly identify how specific biological molecules, such as oxygen, glucose, or bicarbonate, use distinct transport pathways to cross the lipid bilayer.
Through these precision-engineered networks of Plasma Membrane Transport, biological cells achieve true systemic homeostasis. They continuously regulate their internal cellular volume, maintain optimal internal pH levels, extract essential metabolic dietary nutrients, and establish the vital chemical ion gradients necessary for nerve stimulation and muscle contraction. A deep understanding of Plasma Membrane Transport is the undisputed bedrock of modern human biochemistry. By integrating the concepts in this matrix, future scientists will be well equipped to tackle complex physiological and pathological challenges.
Please read our Content Disclaimer Statement.
Check out our social media channels: