63. Cell Fractionation: Unlocking the Cell’s Functional Compartments
Imagine trying to understand how a massive, complex city operates by only looking at it from orbit. To truly grasp its functions, one must examine its individual factories and power plants up close. This same fundamental principle applies to biochemistry. The core purpose of this slide deck is to explore how scientists meticulously dismantle tissues to study cellular inner workings. By analyzing these steps, students will discover exactly how we isolate and understand the fundamental factories of life.
Slide 1: Introduction to Cell Fractionation Methods
The foundational step in any advanced biochemical analysis begins with a robust understanding of cell fractionation. When researchers need to study a specific mitochondrial enzyme or an endoplasmic reticulum receptor in isolation, they cannot simply examine an intact tissue sample. They must deliberately tear the biological structure apart to isolate the individual functioning compartments. The underlying concept of cell fractionation is the critical key to unlocking these tiny, complex cellular structures for deep scientific investigation.
By systematically breaking down outer cellular boundaries, scientists can successfully isolate specific organelles for highly detailed biochemical analysis. The striking visual representation of a gradient in a laboratory test tube beautifully captures the ultimate goal of cell fractionation. The distinct colored bands represent separate biological layers, each containing pure, isolated populations of organelles cleanly separated from the chaotic mixture of the whole organism.
This introductory slide clearly establishes the core objective of cell fractionation: physical isolation and molecular enrichment. Without this essential laboratory technique, modern biochemistry, pharmacology, and molecular biology would be impossible to perform. It allows researchers to study isolated enzymatic reactions without interference from competing cellular pathways present in the intact organism.
Understanding how to physically pull apart the complex cellular matrix helps medical and college students grasp how specific pharmaceutical drugs target precise, localized functions. By mastering the theory behind this organelle separation, future scientists and physicians gain a powerful framework for exploring cellular pathology. They learn precisely how isolating these components paves the way for understanding metabolism, genetic transcription, and energy production at the most fundamental level.
Slide 2: Physical Principles Governing Cell Fractionation
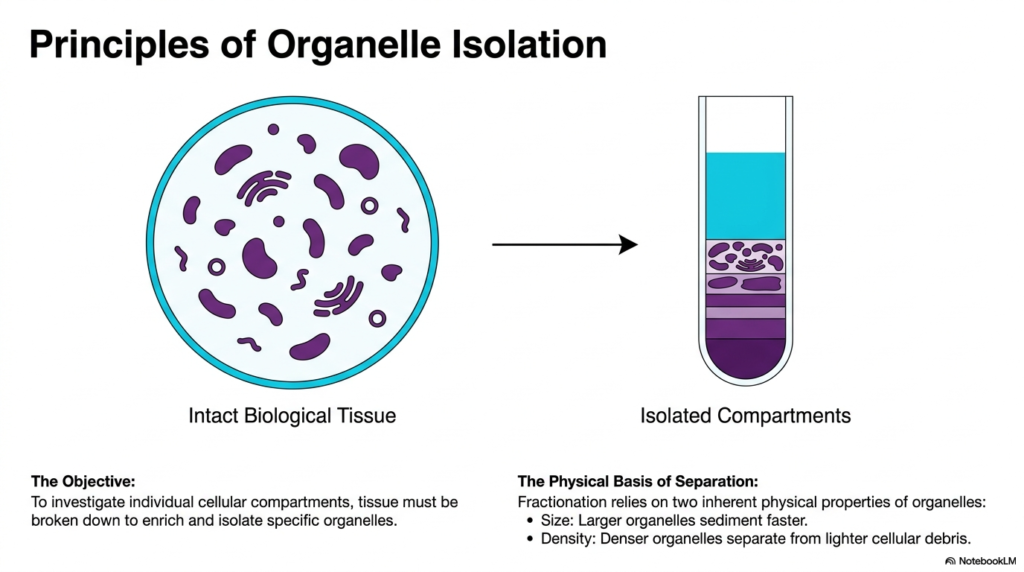
To successfully separate microscopic components, one must understand the inherent physical properties that drive cell fractionation. Intact biological tissue is a remarkably dense and complex matrix filled with various functioning compartments. The primary objective is to investigate these individual cellular compartments by breaking down the tissue to enrich for and isolate specific organelles. Cell fractionation provides the necessary mechanical approach to achieve this targeted enrichment.
The visual transition from intact biological tissue to isolated compartments illustrates the dramatic transformation that occurs during a proper cell fractionation protocol. Researchers move from a chaotic, densely populated environment to a highly organized state in which different organelles are stratified by their unique physical characteristics. This stratification is the definitive hallmark of a successful cell fractionation procedure.
The physical basis of this separation relies primarily on two inherent properties of the organelles: size and density. Larger organelles naturally have greater mass, causing them to sediment much faster under physical forces. Understanding this size differential is crucial for biochemistry students trying to predict which cellular structures will separate first during laboratory experiments.
Furthermore, the density of the organelles dictates how they separate from lighter cellular debris. Denser organelles will move more rapidly toward the bottom of the test tube, creating distinct, isolated layers. By exploiting these simple yet fundamental principles of physics, biochemists can reliably and predictably sort the microscopic components of life.
Every successful isolation heavily depends on exploiting the slight physical variations between these tiny biological machines. Grasping these foundational rules allows students to understand the underlying physics that make complex biochemical diagnostics possible.
Slide 3: Disruption Strategies in Cell Fractionation

Before any sorting can occur, researchers must first breach the tough exterior boundaries of the biological sample, a crucial early phase of cell fractionation. This process, known as homogenization and lysis, involves breaking open the protective outer barriers while carefully preserving the delicate structures hidden inside. Choosing the appropriate disruption method is essential to proper cell fractionation design.
Mechanical disruption offers several distinct pathways for tearing biological tissues apart. The Potter Homogenizer is particularly notable for using a precisely fitted rotating Teflon pestle housed within a glass tube to generate gentle friction. This method provides gentle mechanical disruption, making it an ideal choice for isolating highly fragile internal structures without compromising their biochemical function.
Other mechanical methods used in cell fractionation include aggressive grinding of frozen tissue or shearing the sample with high-pressure rotating knives. Alternatively, researchers may rely on enzymatic and osmotic techniques. Enzymatic lysis involves the targeted degradation of the rigid cell wall, while osmotic shock ruptures cells by applying extreme hypotonic pressure, causing them to swell and burst.
Repeated thermal cycling, known as freeze/thaw lysis, is another highly effective method for breaking down complex barriers. Following these initial disruption steps, coarse filtration is absolutely mandatory. The raw biological suspension must be carefully filtered through gauze to remove any surviving intact whole cells and tough connective tissue fragments before proceeding to the centrifugation stages.
Without this critical filtration step, the entire separation process would become severely compromised by unwanted macroscopic debris. Mastering these varied lysis techniques provides aspiring medical professionals and research scientists with the foundational skills needed to safely extract vital biological machinery from virtually any tissue type.
Slide 4: Optimizing the Cell Fractionation Buffer

Maintaining the biological activity of delicate organelles requires incredibly strict environmental control throughout the entire cell fractionation process. The homogenization buffer is a specialized liquid designed to protect these microscopic structures when they are forcibly removed from their natural environment. Without a properly formulated buffer, cell fractionation would yield nothing but dead, non-functional cellular debris.
Isotonicity is the first crucial condition for a successful homogenization buffer. The osmotic value of the buffer must perfectly match the internal intracellular osmotic value of the original living tissue. This careful biochemical balancing act prevents isolated organelles from taking up excessive water and bursting in hypotonic conditions, or catastrophically shrinking in hypertonic conditions, during cell fractionation.
Temperature control is another non-negotiable factor in any professional cell fractionation protocol. Researchers must perform all reactions on ice, strictly maintaining temperatures between 0 and 5 degrees Celsius. This precise thermal regulation significantly slows destructive enzymatic degradation reactions catalyzed by highly reactive cytosolic enzymes inevitably released when the tissue is initially disrupted.
Finally, chemical protectors must be meticulously integrated into the buffer solution. The strategic addition of specific thiols and metal-chelating agents serves a profoundly important biochemical purpose. These protective chemicals shield functional sulfhydryl groups located on vital proteins from suffering damaging oxidation. Proper buffer preparation enables structures such as isolated mitochondria to actively consume substrates and reliably produce ATP via oxidative phosphorylation for hours.
This remarkable preservation of functional biological activity is the ultimate proof of a perfectly executed preparatory phase. For students studying cellular metabolism, understanding the complex chemistry of the homogenization buffer highlights just how delicate and carefully orchestrated life’s internal machinery truly is.
Slide 5: The Stages of Cell Fractionation Centrifugation

With the biological tissue properly broken down and safely stabilized, the core physical separation of cell fractionation can finally commence. Differential centrifugation is a powerful sequential technique that separates particles entirely based on their distinct sedimentation rates. This crucial mechanism drives the central sorting phase of modern cell fractionation experiments.
The procedure begins with a low-acceleration step using standard benchtop centrifuges. This initial application of force effectively separates the largest, densest biological particles, driving them to the bottom to form a solid sediment called a pellet. The remaining liquid above the pellet, containing smaller structures, is known as the supernatant and must be carefully preserved for the next phase.
In the second step of this cell fractionation method, researchers perform step-by-step acceleration. The saved supernatant is carefully decanted into a fresh tube and subsequently subjected to significantly higher gravitational forces. Using advanced high-speed and ultracentrifuges, scientists apply intense centrifugal force to force the next size class of organelles out of the suspension.
The final stage involves successive sedimentation, an iterative process of gradually increasing the applied gravitational force. Because particles that are much smaller and less dense stubbornly remain suspended in the liquid, they require exponentially higher g-forces to be forced out of the solution. This strategic, stepwise acceleration isolates increasingly microscopic biological components with remarkable precision.
Understanding this sequential gravitational sorting allows college biochemistry students to conceptually visualize how complex mixtures are methodically simplified. It perfectly demonstrates how manipulating basic physics in the laboratory can cleanly resolve the chaotic inner space of biology into neat, highly organized, and easily analyzable discrete packages.
Slide 6: The Low-Speed Cell Fractionation Step

The first major isolated product of the differential centrifugation process is the low-speed fraction, a crucial milestone in complete cell fractionation. By applying a relatively gentle gravitational force of 600 times Earth’s gravity for exactly 10 minutes, researchers can easily pull down the heaviest cellular components. This initial step in cell fractionation relies on readily attainable accelerations using standard laboratory benchtop centrifuges.
The primary organelle isolated during this first low-speed spin is the nucleus. Because the nucleus houses the organism’s entire genetic blueprint, it is naturally the largest and densest functional compartment in a standard eukaryotic cell. Consequently, it sediments rapidly, forming a distinct dark pellet at the bottom of the centrifuge tube while leaving lighter materials suspended in the fluid above.
However, achieving a perfectly pure sample is incredibly difficult during the early stages of cell fractionation. The low-speed fraction often contains notable contaminants and co-sediments that naturally fall at similar physical rates. These unwanted impurities typically include structural cytoskeletal fragments, intact cells that survived the initial filtration process, and dense pieces of connective tissue.
To definitively verify that the isolated pellet genuinely contains the targeted nuclear material, biochemists rely on specific molecular identifiers. The presence of DNA serves as the definitive biochemical marker for this step, as it is localized exclusively within this primary low-speed fraction. Testing for this specific nucleic acid allows students and scientists to confidently confirm the success of their initial separation.
Recognizing these inherent contaminants and understanding how to definitively validate the isolated material are essential critical-thinking skills for any developing medical researcher. It underscores the vital importance of combining physical separation techniques with rigorous molecular verification to ensure complete scientific accuracy.
Slide 7: The Medium-Speed Cell Fractionation Step

After successfully removing the heavy nuclei, researchers accelerate the remaining supernatant to isolate the next layer of complexity in the cell fractionation process. The medium-speed fraction requires spinning the liquid at a much stronger force of 15,000 times gravity for fifteen continuous minutes. This essential intermediate step of cell fractionation isolates highly active metabolic centers.
The primary organelles yielded from this specific cell fractionation stage are the mitochondria, lysosomes, and peroxisomes. In plant tissue samples, this exact gravitational threshold is also famously responsible for cleanly isolating the green chloroplasts. By carefully decanting the supernatant left over from the initial nuclear centrifugation, biochemists isolate the engines and recycling centers of the biological system.
Because this physical layer contains multiple distinct organelles that co-sediment due to similar densities, definitive biochemical markers are absolutely critical for distinguishing them. To verify the presence of mitochondria, scientists test for the activity of succinate dehydrogenase or cytochrome-c oxidase. These complex enzymes are deeply involved in energy production and serve as ideal molecular signatures of the mitochondrial fraction.
Similarly, unique enzymes help identify the other major compartments isolated in this spin. Lysosomes are accurately identified by detecting specific degradative enzymes such as beta-N-acetylhexosaminidase and beta-galactosidase. Meanwhile, peroxisomes are confirmed by the strong presence of catalase, an enzyme that breaks down dangerous hydrogen peroxide.
Understanding this diverse molecular checklist allows advanced biology students to accurately map metabolic functions to their precise physical locations. It highlights how biochemical identification becomes completely necessary when physical separation alone cannot perfectly distinguish between similarly sized cellular factories.
Slide 8: The High-Speed Cell Fractionation Step

To reach the truly microscopic structural elements of the biological system, researchers must push the limits of cell fractionation by applying extreme physical forces. The high-speed fraction subjects the previously saved supernatant to a staggering 100,000-times-gravity force for a full sixty minutes. This intensive phase of cell fractionation successfully strips out the delicate, highly complex internal membrane networks.
The primary compartments isolated at this high-speed threshold include the endoplasmic reticulum and the vital plasma membrane. In scientific literature, this specific cell fractionation layer is frequently referred to as the microsomal fraction or simply as membrane vesicles. The extreme physical forces naturally cause the delicate ER, the Golgi apparatus, and the outer cellular membranes to fragment and then spontaneously reseal into tiny, empty spherical structures.
Identifying these tiny, generic-looking vesicles requires testing for highly specific, membrane-bound biochemical markers. To confidently prove the presence of the original plasma membrane, scientists specifically assay for the vital Na+/K+ ATPase pump or the specific enzyme phosphodiesterase I. Finding these active proteins confirms that the outer boundary layer has been successfully isolated.
Validating the internal membrane networks requires completely different enzymatic signatures. The endoplasmic reticulum is definitively marked by the presence of glucose 6-phosphatase, a critical enzyme in sugar metabolism. Furthermore, the presence of the Golgi complex can be reliably confirmed by testing the sample for active alpha-mannosidase II.
Understanding the nature of these resealed vesicles is fundamental for medical students studying pharmacology, as these membranes contain the vital receptors targeted by modern therapeutic drugs. Isolating them provides a clean testing ground for new biochemical treatments.
Slide 9: The Ultra-Speed Cell Fractionation Step

Reaching the absolute smallest physical components of life requires the most demanding and rigorous stage of the entire cell fractionation protocol. The ultra-speed fraction forces the remaining liquid through an astonishing 300,000-times-gravity field for a prolonged 120 minutes. This specialized cell fractionation step relies heavily on advanced ultracentrifuges spinning at near-impossible speeds within a complete vacuum.
The primary components isolated during this final particulate phase of cell fractionation are the tiny ribosomes and massive free-floating macromolecules. These structures represent the smallest isolable particulate components within the living cell. Forcing these incredibly light, microscopic protein-assembly machines out of their stable suspension requires immense, prolonged gravitational manipulation.
Despite the extreme forces applied, this delicate fraction is still highly susceptible to microscopic contamination. The most common unwanted co-sediments found at this profound depth include infectious viruses and abnormally large free macromolecules that mimic ribosomal densities. Researchers must remain highly vigilant, as viral contamination can severely disrupt sensitive biochemical sequencing experiments.
To accurately confirm the presence and purity of the isolated ribosomal factories, scientists utilize a very specific biochemical marker. Because ribosomes are primarily composed of tightly packed structural nucleic acids, the definitive presence of ribosomal RNA (rRNA) serves as the ultimate verification.
Mastering this final physical extraction demonstrates how deeply we can probe the fundamental building blocks of biology. For students focusing on genetics and molecular biology, isolating pristine ribosomes is the crucial first step toward fully understanding complex protein synthesis and viral pathology.
Slide 10: The Cytosol and Cell Fractionation

After subjecting the original sample to multiple exhausting rounds of extreme gravitational force, the particulate portion of the cell fractionation process officially concludes. Following the massive 300,000-times-gravity centrifugation step, all particulate and membranous matter has been successfully removed from the suspension. The perfectly clear, remaining liquid is the final supernatant, representing the very end of the physical cell fractionation journey.
This remaining soluble fraction is universally known as the cytosol. It consists entirely of the original specialized homogenization buffer that was expertly mixed at the beginning and thoroughly combined with the cell’s naturally soluble molecular components. This highly complex liquid contains thousands of free-floating metabolic enzymes, simple sugars, and dissolved raw materials that are completely non-membrane-bound.
Because the cytosol lacks any physical structure or identifiable shapes, visual confirmation of this cell fractionation stage is completely impossible. Instead, biochemists must rely on a highly reliable, universally present soluble biochemical marker. The enzyme L-Lactate dehydrogenase, which freely floats within the cellular fluid to assist in rapid anaerobic energy production, serves as the perfect definitive molecular signature for this liquid fraction.
Understanding the true biochemical nature of the cytosol is incredibly important for medical and science students. It proves that a significant portion of vital biological activity does not occur within specialized, walled-off factories, but rather freely dissolved in the chaotic, watery matrix that physically connects every single internal organelle.
Slide 11: Validating Cell Fractionation Results

Performing the intense physical separation is only half the battle; ensuring the scientific accuracy of the cell fractionation is equally important. Isolated cell organelles rapidly lose their inherent biological activity, and physical fractions are often contaminated by co-sedimenting cellular debris. The fundamental problem facing every cell fractionation experiment is determining how to definitively prove the exact contents hidden within the test tube.
The universally accepted solution to this complex problem is to actively analyze each fraction for highly specific Marker Molecules. These specialized markers are distinct, measurable enzymes or unique nucleic acids that occur exclusively or predominantly in one very specific type of cellular compartment. By hunting for these rare biological signatures, researchers can cut through the visual ambiguity of cell fractionation.
The final result of this rigorous molecular testing provides absolute scientific confidence. When researchers measure the precise enzymatic activity of these organelle-specific marker enzymes, it strongly confirms the precise biological compartmentation. Furthermore, testing these markers ensures the overall biochemical purity of the specifically isolated cellular fraction, thereby confirming that the experiment was completely successful.
Grasping the critical importance of marker validation protects young scientists from publishing wildly inaccurate data derived from heavily contaminated laboratory samples. It reinforces the rigid academic principle that visual or physical separation is essentially meaningless in advanced biochemistry without strictly verified, quantifiable molecular evidence backing it up.
Slide 12: The Cell Fractionation Marker Matrix

To streamline the rigorous validation process, biochemists rely heavily on a standardized diagnostic matrix that details crucial cell-fractionation markers. This comprehensive table efficiently maps every major cellular compartment directly to its corresponding highly specific biochemical marker. This matrix serves as an essential diagnostic tool for anyone performing advanced cell fractionation in a modern laboratory.
The matrix clearly outlines the distinct chemical signatures required to validate a successful cell fractionation experiment. For instance, the heavy nucleus is definitively marked by the presence of vital DNA, categorized as a nucleic acid. The plasma membrane is confidently identified by tracking the active enzyme Na+/K+ ATPase, which possesses the specific Enzyme Commission number 3.1.4.1, detailing its exact biochemical classification.
As the matrix continues, it reveals the enzymatic fingerprints for the deeper intracellular structures. The delicate endoplasmic reticulum relies on Glucose 6-phosphatase, while the nearby Golgi complex is validated using alpha-Mannosidase II. Lysosomes are uniquely proven by beta-N-Acetylhexosaminidase and beta-Galactosidase. The energy-producing mitochondrion requires the presence of either Succinate dehydrogenase or cytochrome c oxidase.
Finally, the matrix covers the smallest biological elements. Endosomes are identified by measurable uptake of peroxidase, while isolated ribosomes are confirmed by rRNA structure. The completely soluble cytosol is demonstrated by L-lactate dehydrogenase. Memorizing these vital molecular pairings is a fundamental requirement for any serious medical student looking to master complex cellular diagnostics.
Slide 13: The Complete Cell Fractionation Pathway

Synthesizing all the complex individual steps reveals the elegant, overarching design of the complete cell fractionation pathway. The entire rigorous procedure flows logically, beginning forcefully with intact tissue and ending smoothly with isolated molecular components. This comprehensive cell fractionation roadmap illustrates exactly how targeted physical disruption and sequential gravity systematically dismantle biological complexity.
The pathway begins by aggressively converting intact tissue into a raw, whole-cell homogenate using a specialized Potter Homogenizer, a protective buffer, and coarse-gauge filtration. The first relatively gentle spin at 600× gravity isolates Pellet 1, which contains only the heavy nucleus marked by DNA. The remaining liquid supernatant moves downward, driving the next phase of the cell fractionation separation process.
The sequential cascade continues as the supernatant is centrifuged at 15,000 × g to yield Pellet 2, which contains mitochondria validated by succinate dehydrogenase activity. A dramatically faster 100,000-times-gravity spin isolates Pellet 3, containing the microsomal fragments of the ER and plasma membrane, marked by glucose 6-phosphatase. The final extreme ultra-spin at 300,000 times gravity pulls down Pellet 4, the tiny ribosomes marked purely by rRNA.
What remains completely suspended is the final, crystal-clear supernatant containing the soluble cytosol, verified by L-Lactate dehydrogenase. Ultimately, this complete fractionation pathway yields beautifully enriched, biochemically distinct cellular compartments that are fully stabilized and ready for highly specialized laboratory assays.
Please read our Content Disclaimer Statement.
Check out our social media channels: