66. Eukaryotic Cytoskeleton: Structure, Components and Functions
Imagine a bustling city entirely devoid of roads, support scaffolding, or transport vehicles; absolutely nothing would function. Similarly, biological cells require a dynamic internal framework to maintain their precise shape, move actively, and safely transport essential cargo. This vital framework is the eukaryotic cytoskeleton. The core purpose of this comprehensive slide deck is to explore the intricate molecular architecture and biochemical dynamics of the eukaryotic cytoskeleton. By carefully examining its diverse protein systems, students will gain a thorough understanding of the fundamental mechanisms that drive cellular life.
Slide 1: Eukaryotic Cytoskeleton: Molecular Architecture and Biochemical Dynamics

The foundation of cellular biology begins with understanding the eukaryotic cytoskeleton, the intricate internal scaffolding that dictates cellular shape and integrity. This dynamic network of interlacing protein polymers extends throughout the cytoplasm, providing essential structural support and organization for a myriad of biochemical processes. Without the eukaryotic cytoskeleton, cells would simply collapse into disorganized, amorphous spheres, entirely unable to coordinate the complex, life-sustaining reactions required for survival. The cover slide visually captures this profound concept by depicting the interlaced nature of distinct structural components. This imagery highlights the interconnected, highly organized nature of these vital protein networks.
The visual representation on this introductory slide serves as a powerful conceptual anchor for studying the eukaryotic cytoskeleton. It illustrates how different filament types crosslink and interact to form a cohesive structural matrix. This matrix is not merely static; it is a highly responsive system that continually builds and rebuilds itself to meet the cell’s physiological demands. Understanding this visual metaphor is crucial for medical and college students, as it sets the stage for exploring the specific biochemical interactions that govern cellular stability and movement.
As this presentation unfolds, it will delve into the molecular architecture that makes the eukaryotic cytoskeleton such an extraordinary biological phenomenon. The focus will remain on the detailed biochemical dynamics that allow these protein structures to function as both rigid supports and flexible transport highways. By dissecting the composition and polymerization processes of these systems, students will gain a comprehensive understanding of cellular mechanics. This foundational knowledge is absolutely critical for recognizing how cellular dysfunction leads to various human pathologies and diseases.
The journey through this cellular scaffolding reveals the beauty of biochemical engineering. Every protein subunit and energy-dependent motor action demonstrates the precision of biological systems. Grasping the overall scope of this architectural marvel prepares the learner for the intricate details ahead, moving from broad structural concepts to the specific, molecular mechanisms that power cellular life.
Slide 2: Eukaryotic Cytoskeleton: The Three Pillars of Cellular Scaffolding
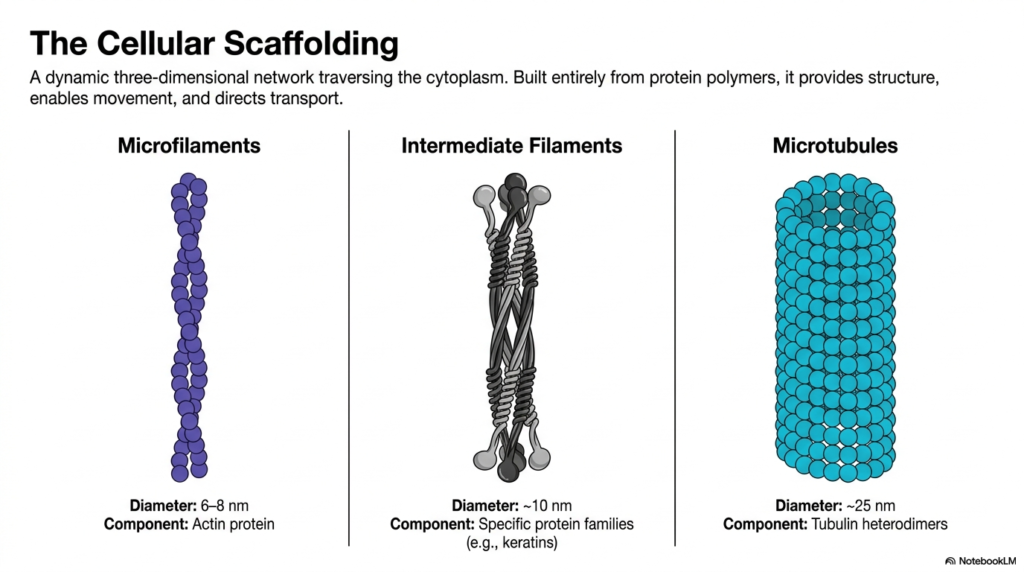
The eukaryotic cytoskeleton is a dynamic, three-dimensional network that completely traverses the cytoplasm. This robust framework is built entirely from diverse protein polymers, each serving distinct yet overlapping physiological roles. The three primary pillars of the eukaryotic cytoskeleton are microfilaments, intermediate filaments, and microtubules. Together, they provide essential structure, enable intricate cellular movement, and direct the transport of intracellular cargo. Understanding the physical dimensions and biochemical composition of these three components provides a crucial baseline for further exploration into cellular mechanics.
Microfilaments, the thinnest components, measure approximately six to eight nanometers in diameter and are composed entirely of actin protein subunits. These flexible structures often concentrate near the cell periphery, dictating cell shape and driving local membrane movements. In contrast, intermediate filaments offer a middle ground with a diameter of about ten nanometers. Constructed from specific protein families such as keratins, they act as the cell’s resilient shock absorbers, providing unparalleled mechanical strength and stability against shear stress and physical deformation.
The largest structures within the eukaryotic cytoskeleton are microtubules, which form hollow cylinders measuring roughly twenty-five nanometers in diameter. Built from tubulin heterodimers, these robust tubes act as the primary structural struts and intracellular transport highways. By comparing the diameters, structural subunits, and general functions of microfilaments, intermediate filaments, and microtubules, students can appreciate the specialized design of this cellular scaffolding. This tripartite system perfectly illustrates how biological forms are precisely tailored to execute complex physiological functions within the microscopic world of the cell.
The synergistic interaction between these three filament systems ensures that the cell remains a highly organized, functional unit rather than a chaotic soup of organelles. Their ability to rapidly assemble, disassemble, and reorganize is a testament to the sophisticated biochemical regulation occurring constantly within living organisms. Grasping this overarching architecture is the first step toward mastering the complexities of molecular biology and cellular physiology.
Slide 3: Eukaryotic Cytoskeleton: The Biochemical Assembly of Actin Microfilaments
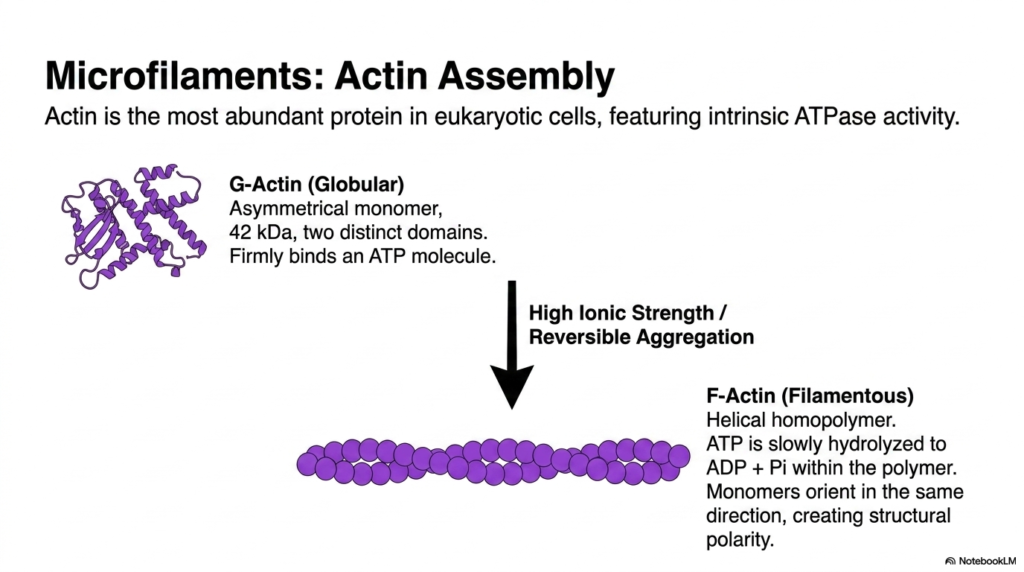
Actin stands out as the most abundant protein in the eukaryotic cytoskeleton across many cell types, exhibiting remarkable intrinsic ATPase activity. The fundamental building block is G-actin, a globular, asymmetrical monomer weighing forty-two kilodaltons. This monomer features two distinct domains and firmly binds a single molecule of ATP. The asymmetrical nature of G-actin is a critical biochemical feature, as it underpins the directional growth and overall polarity of the assembled filament, a concept essential for cellular movement.
The transformation from individual monomers into a functional component of the eukaryotic cytoskeleton requires specific biochemical conditions. Under high ionic strength, G-actin undergoes reversible aggregation, polymerizing to form F-actin, or filamentous actin. This resulting structure is a helical homopolymer that twists into the familiar microfilament shape. During this polymerization process, the bound ATP is slowly hydrolyzed to ADP and inorganic phosphate within the polymer matrix. This hydrolysis fundamentally alters the binding affinity between subunits, driving the dynamic turnover of the filament structure.
A defining characteristic of F-actin within the eukaryotic cytoskeleton is its structural polarity. Because every G-actin monomer orients in the same direction during assembly, the resulting filament has distinct ends with markedly different biochemical properties. This uniform orientation is not merely a structural anomaly; it is a vital functional requirement. The polarity allows motor proteins to move directionally along the filament and regulates the precise rates of polymerization and depolymerization, ultimately granting the cell control over its shape and motility.
The ability of actin to rapidly transition between a soluble globular state and a rigid filamentous state perfectly demonstrates the highly dynamic nature of cellular architecture. By tightly regulating this assembly process, cells can project membranes, engulf nutrients, and divide with astonishing precision. This regulated assembly forms the mechanical basis for countless physiological processes across all domains of life.
Slide 4: Eukaryotic Cytoskeleton: Treadmilling Dynamics and Pharmacological Control of Actin
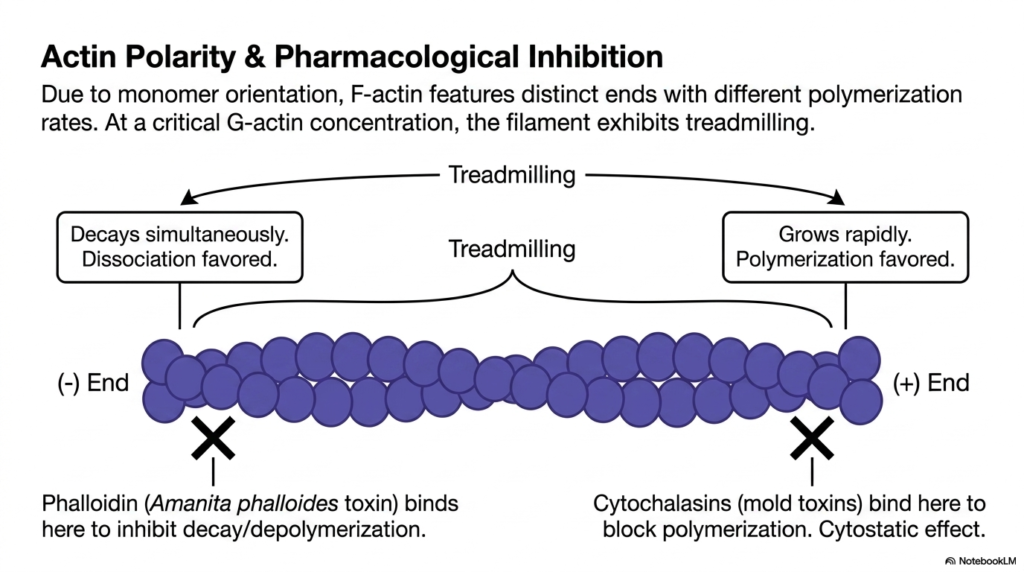
Because of the uniform orientation of its monomers, F-actin within the eukaryotic cytoskeleton features highly distinct ends that exhibit vastly different polymerization rates. The plus end grows rapidly, strongly favoring the addition of new ATP-bound G-actin monomers. Conversely, the minus end favors dissociation, where older, ADP-bound actin subunits are lost. When the concentration of available G-actin reaches a specific critical threshold, the filament enters a fascinating state known as treadmilling. During treadmilling, the filament grows at one end while simultaneously decaying at the other.
This treadmilling phenomenon is a cornerstone of movement within the eukaryotic cytoskeleton. Even though the overall length of the microfilament may remain completely constant, the individual actin subunits are continually moving through the polymer like a biological conveyor belt. This continuous flux allows the cell to rapidly remodel its internal framework and push the plasma membrane forward during cell migration. Understanding this dynamic instability is crucial for comprehending how cells can physically propel themselves through their surrounding environment.
The critical importance of the eukaryotic cytoskeleton is further highlighted by the mechanisms of various potent natural toxins. For example, phalloidin, a lethal toxin derived from the death cap mushroom, binds specifically to the minus end of F-actin, aggressively inhibiting depolymerization and freezing the filament in place. Alternatively, cytochalasins, which are potent mold toxins, bind directly to the plus end, completely blocking new polymerization and exerting a profound cytostatic effect that halts cellular division and migration.
By utilizing these naturally occurring pharmacological inhibitors, researchers have unraveled the profound complexities of actin dynamics. These toxins demonstrate that the continuous turnover of actin filaments is absolutely essential for cell survival. Disrupting this delicate balance, either by preventing growth or halting decay, leads to immediate cellular paralysis and eventual death, highlighting the exquisite sensitivity of these microscopic structural systems.
Slide 5: Eukaryotic Cytoskeleton: Regulatory Proteins and Actin Network Modulation
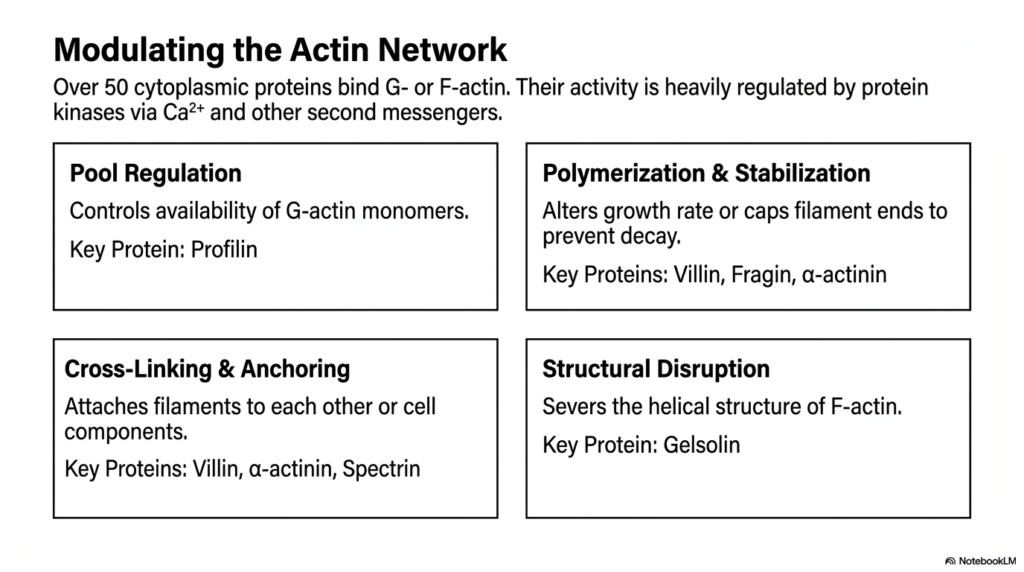
The eukaryotic cytoskeleton is not a static construct; it is a highly regulated network controlled by over fifty specialized cytoplasmic proteins that bind directly to G-actin or F-actin. The activity of these vital accessory proteins is tightly regulated by protein kinases in response to calcium ions and other second messengers. This intricate regulatory cascade ensures that the cell can precisely dictate when, where, and how actin filaments assemble or disassemble. One primary mechanism of control is pool regulation, which governs the availability of free G-actin monomers in the cytoplasm.
Profilin is a key protein involved in pool regulation, preventing premature polymerization and ensuring a ready supply of subunits for the eukaryotic cytoskeleton. Beyond simply controlling the monomer pool, cells must also regulate the polymerization and stabilization of existing filaments. Key proteins like villin, fragin, and alpha-actinin can alter the filament growth rate or completely cap the ends to prevent rapid decay. This capping mechanism provides structural longevity to filaments that need to maintain their architectural integrity over extended periods.
To form a functional tissue matrix, the eukaryotic cytoskeleton must also engage in extensive cross-linking and anchoring. Proteins such as spectrin and alpha-actinin physically attach individual actin filaments to one another or anchor them firmly to other cellular components, such as the plasma membrane. This cross-linking creates a robust, three-dimensional web that provides formidable mechanical strength. Conversely, when rapid remodeling is required, structural disruption proteins like gelsolin actively sever the helical structure of F-actin, instantly liquefying the local cytoskeletal network.
This complex interplay of polymerizing, stabilizing, cross-linking, and severing proteins provides the cell with an unparalleled degree of architectural control. By simply modulating the activity of these accessory proteins through localized calcium signaling, a cell can instantly transform its cytoplasm from a rigid gel to a fluid state, facilitating immediate structural adaptation.
Slide 6: Eukaryotic Cytoskeleton: Intermediate Filaments and Structural Resilience

Unlike the highly dynamic nature of actin and tubulin, intermediate filaments form the highly stable, permanent scaffolds of the eukaryotic cytoskeleton. These robust polymers lack the structural polarity seen in microfilaments and microtubules, meaning they do not possess distinct growing or shrinking ends. Furthermore, free intermediate filament monomers rarely exist floating in the cytoplasm; they are almost exclusively found integrated within the polymerized network. This permanence provides cells with an incredibly durable mechanical framework that withstands immense physical stress and shear forces.
The biochemical composition of intermediate filaments within the eukaryotic cytoskeleton is remarkably diverse, consisting of five primary protein families that are highly specific to particular cell types. For instance, cytokeratins are the hallmark intermediate filaments found in epithelial cells, providing the tough outer barrier of the skin. Desmin is specifically utilized by muscle cells to maintain structural integrity during forceful contractions, while vimentin provides support within mesenchymal cells. This tissue-specific distribution highlights the evolutionary customization of the cellular scaffold.
In the nervous system, the eukaryotic cytoskeleton relies on Glial Fibrillary Acidic Protein, or GFAP, to support glial cells, while neurons utilize specialized neurofilaments to maintain the long, delicate structures of their axons. Additionally, laminins are a unique class of intermediate filament proteins that form a protective meshwork that exclusively supports the inner nuclear membrane. This incredible diversity allows different tissues to tailor their internal mechanical properties to meet their specific physiological and environmental demands.
Understanding the tissue-specific nature of these five protein families is exceptionally important in modern medical diagnostics. Because these filaments remain stable even when cells become cancerous, pathologists routinely use intermediate filament typing to precisely identify the cellular origins of metastatic tumors. This biochemical signature turns these structural proteins into invaluable clinical biomarkers for disease diagnosis and targeted treatment strategies.
Slide 7: Eukaryotic Cytoskeleton: The Hierarchical Assembly of Intermediate Filaments
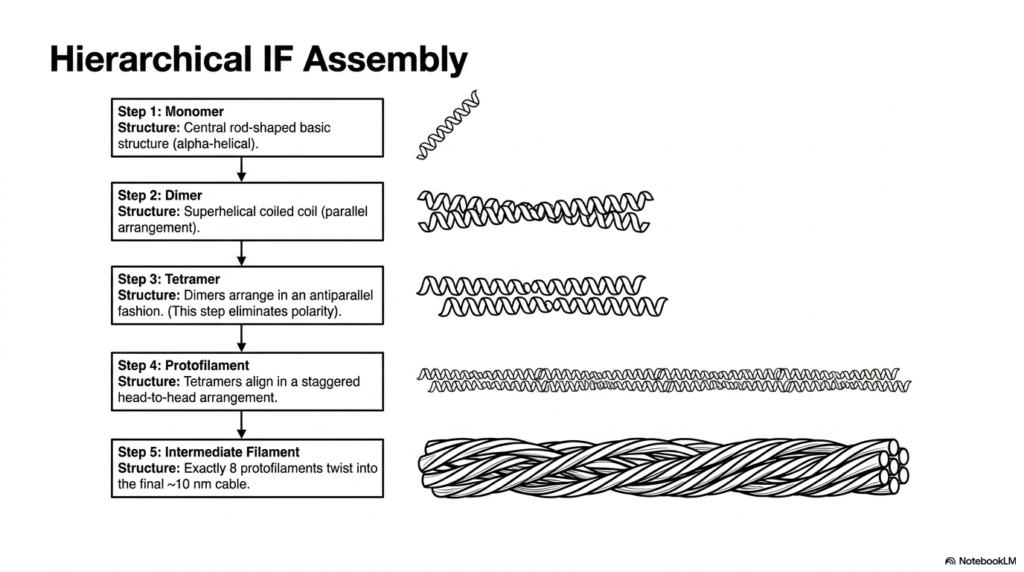
The construction of intermediate filaments within the eukaryotic cytoskeleton is a marvel of hierarchical biochemical assembly. The process begins with a single monomer featuring a central, rod-shaped basic structure that is distinctly alpha-helical. Unlike the globular subunits of actin or tubulin, this elongated fibrous shape is perfectly designed for lateral association and tensile strength. In the second step, two of these monomers align in parallel, twisting together to form a highly stable superhelical coiled-coil dimer.
The assembly of the eukaryotic cytoskeleton continues as two dimers arrange themselves in a staggered, antiparallel fashion to form a tetramer. This antiparallel arrangement is biochemically crucial because it completely eliminates the structural polarity characteristic of the individual monomers. Consequently, the resulting tetramer and all subsequent structures built from it lack the distinct plus and minus ends seen in other cytoskeletal systems. This lack of polarity is precisely why intermediate filaments do not participate in directional motor transport.
In the fourth step of building this component of the eukaryotic cytoskeleton, these non-polar tetramers align linearly in a staggered, head-to-head arrangement to create a long protofilament. Finally, the ultimate structural unit is formed when exactly eight of these protofilaments bundle and twist together. They intertwine to form the final rope-like cable, measuring approximately 10 nanometers in diameter. This multi-layered, twisted-cable architecture provides intermediate filaments with their legendary resistance to stretching and bending.
The resulting ten-nanometer cable behaves much like a microscopic steel wire rope, capable of withstanding substantial physical trauma and cellular deformation without snapping. This incredibly robust hierarchical assembly ensures that tissues subjected to constant mechanical stress, such as the outer layers of the skin or the continually beating muscle cells of the heart, strictly maintain their cellular integrity under extreme physiological pressure, thereby preventing catastrophic tissue rupture and cellular death.
Slide 8: Eukaryotic Cytoskeleton: The Cylindrical Architecture of Microtubules
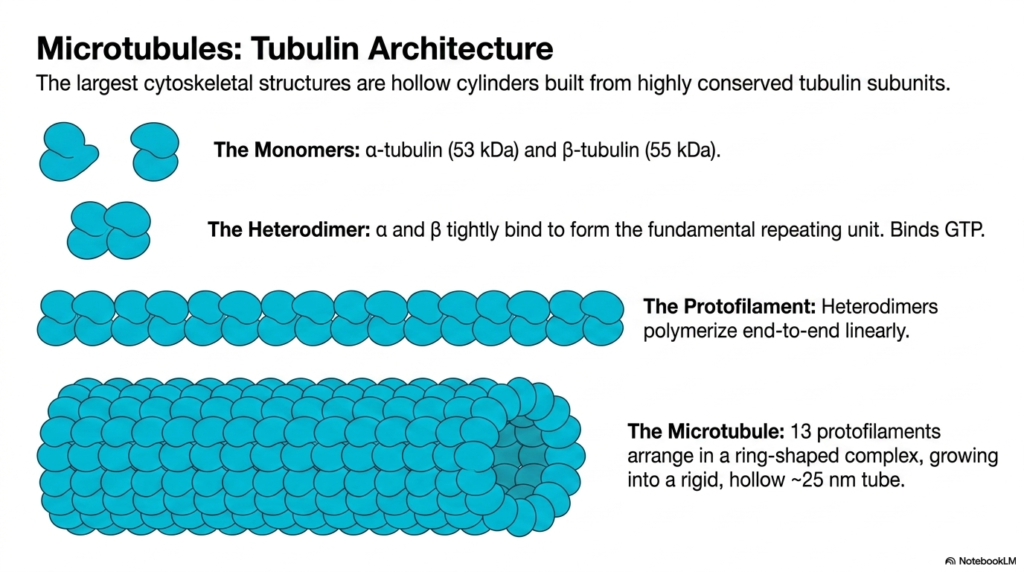
Microtubules are universally recognized as the largest structural elements within the eukaryotic cytoskeleton. Unlike the solid ropes of intermediate filaments or the thin twists of actin, microtubules are massive, hollow cylinders built from highly conserved tubulin subunits. The fundamental building blocks are two distinct globular monomers: alpha-tubulin, weighing fifty-three kilodaltons, and beta-tubulin, weighing fifty-five kilodaltons. These two monomers tightly bind to one another to form the fundamental repeating unit, the tubulin heterodimer, which actively binds guanosine triphosphate, or GTP.
The assembly of this colossal component of the eukaryotic cytoskeleton begins when these tubulin heterodimers polymerize end-to-end in a strictly linear fashion to form a long string known as a protofilament. Because the heterodimers always align in the exact same orientation, alternating alpha and beta subunits, the resulting protofilament possesses inherent structural polarity. This polarity is a defining feature of the microtubule network, establishing a clear biochemical directionality that is absolutely essential for the movement of intracellular motor proteins.
To form the final functional tube of the eukaryotic cytoskeleton, precisely 13 individual protofilaments arrange themselves side by side in a circular, ring-shaped complex. This lateral association causes the structure to fold into a rigid, hollow tube measuring approximately twenty-five nanometers in external diameter. The hollow, cylindrical geometry of the microtubule provides exceptional stiffness and resistance to compressive forces, allowing these structures to span vast cellular distances without buckling or collapsing under the weight of the cytoplasm.
This unique tubular architecture makes microtubules the premier structural struts of the microscopic cellular world. Their impressive biochemical rigidity allows them to dictate the overall spatial layout of cellular organelles, form the highly resilient mitotic spindle required for accurate chromosome segregation during cell division, and construct the complex internal structural cores of motile cellular appendages like cilia and flagella.
Slide 9: Eukaryotic Cytoskeleton: Microtubule Dynamic Instability and Toxin Interdiction
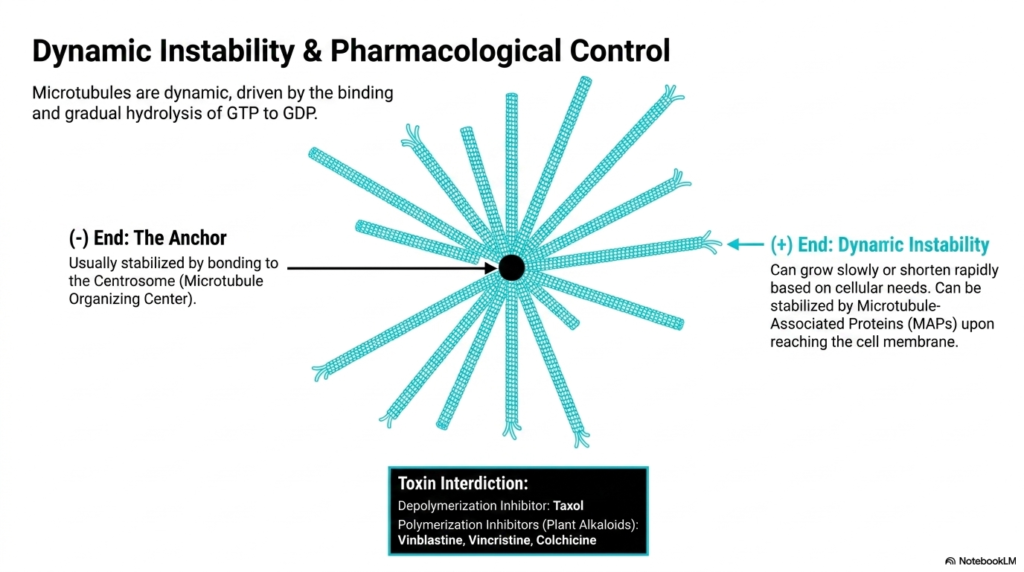
Microtubules within the eukaryotic cytoskeleton are incredibly dynamic structures, constantly shifting between phases of rapid growth and catastrophic shrinkage. This remarkable behavior, termed dynamic instability, is entirely driven by the binding and gradual hydrolysis of GTP to GDP within the beta-tubulin subunit. The minus end of the microtubule acts as the stable anchor, usually firmly attached to and stabilized by the Centrosome, which functions as the primary Microtubule Organizing Center located near the cell’s nucleus.
In stark contrast to the anchored minus end, the plus end of this eukaryotic cytoskeleton component radiates outward toward the cell periphery and exhibits profound dynamic instability. It can grow slowly as new GTP-bound heterodimers are added, or it can shorten rapidly in a dramatic event called a catastrophe if the GTP cap is lost. However, this volatile plus end can be therapeutically stabilized by specific Microtubule-Associated Proteins (MAPs) once it reaches its target near the cell membrane.
The rapid turnover of the eukaryotic cytoskeleton makes it a prime target for pharmacological intervention, particularly in the treatment of aggressive cancers. Several potent toxins directly interfere with microtubule dynamics. Taxol, for instance, acts as a powerful depolymerization inhibitor, binding tightly to microtubules and freezing them in a polymerized state. This artificial stabilization completely halts the dynamic turnover required for the mitotic spindle to function, effectively blocking cell division and leading to cellular apoptosis.
Conversely, plant alkaloids such as vinblastine, vincristine, and colchicine function as profound polymerization inhibitors. By binding to free tubulin heterodimers, they prevent the assembly of new microtubules, causing the existing network to rapidly dissolve. Whether freezing the network or dissolving it, these chemical agents prove that the constant, dynamic instability of microtubules is an absolute prerequisite for successful cellular proliferation and basic survival.
Slide 10: Eukaryotic Cytoskeleton: Executing Major Physiological Tasks
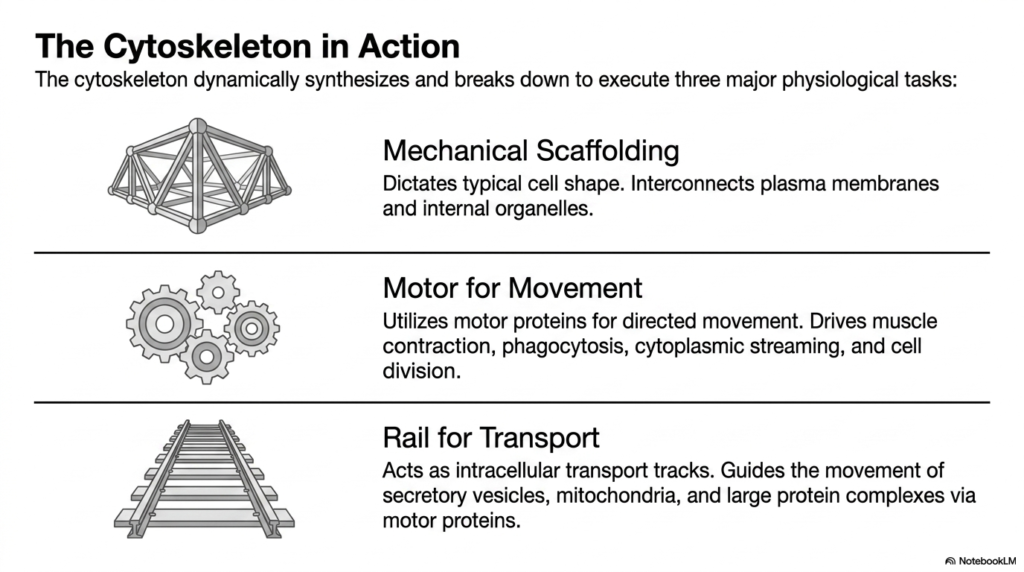
The true brilliance of the eukaryotic cytoskeleton lies in its ability to dynamically assemble and disassemble its networks to execute three major physiological tasks essential for cellular life. The first critical function is providing mechanical scaffolding. The intricate web of filaments fundamentally determines the cell’s typical shape, preventing it from collapsing under external pressure. Furthermore, this scaffold physically interconnects the plasma membrane with internal organelles, anchoring them in optimal spatial orientations to maximize biochemical efficiency.
Beyond static support, the eukaryotic cytoskeleton serves as an indispensable motor for movement. By using specialized motor proteins that convert chemical energy into mechanical force, the cytoskeletal network enables dramatic changes in cellular shape. This directed movement is the fundamental driving force behind major physiological events, including the powerful contraction of muscle fibers, the engulfment of foreign particles during phagocytosis, the cytoplasmic streaming that churns, and the physical separation of cells during active cell division.
Finally, the eukaryotic cytoskeleton operates as an elaborate railway system for intracellular transport. Microtubules and microfilaments act as precise, high-speed transport tracks traversing the vast expanse of the cytoplasm. This network guides the rapid, directional movement of essential secretory vesicles, massive organelles like mitochondria, and large protein complexes. Specialized motor proteins haul this crucial cargo along the cytoskeletal rails, ensuring that biological materials reach their exact cellular destinations precisely when they are needed most.
Without this highly dynamic and deeply multifunctional system, a living cell would be entirely immobilized and completely incapable of distributing vital nutrients, removing toxic metabolic waste, or interacting with its environment. The profound ability to simultaneously act as a rigid architectural support beam, an active contractile biological engine, and a directed molecular highway makes this integrated protein network the most versatile, complex, and essential structural component discovered within modern cellular biology.
Slide 11: Eukaryotic Cytoskeleton: Architectural Study of the Microvillus
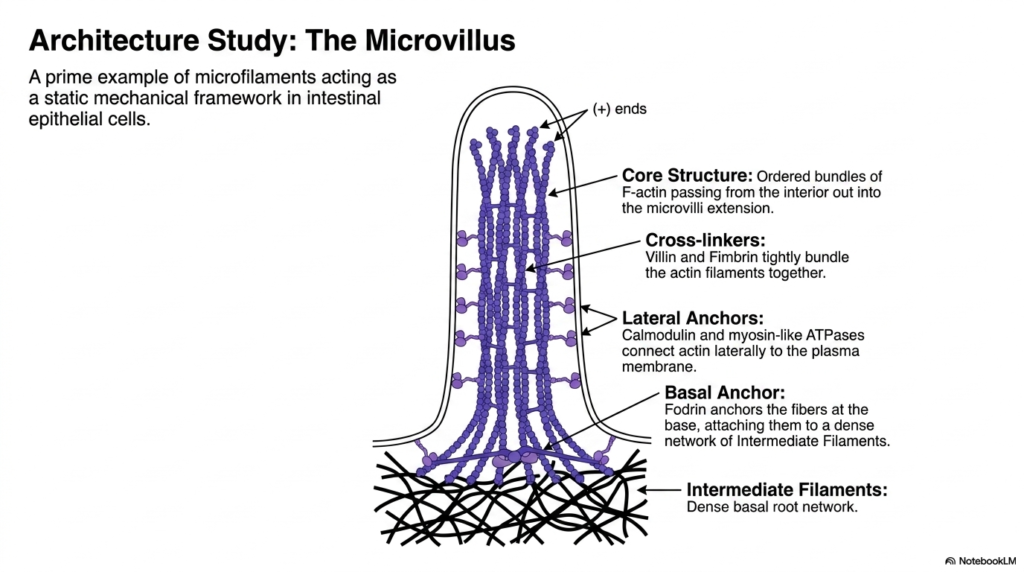
To truly grasp the functional capacity of the eukaryotic cytoskeleton, one must examine specific architectural studies, such as the structure of the microvillus. Microvilli are a prime example of microfilaments forming a highly stable, static mechanical framework. Found extensively on the apical surface of intestinal epithelial cells, these tiny cellular protrusions exponentially increase the surface area available for nutrient absorption. The structural integrity of these vital extensions relies entirely on the precise organization of the internal actin network.
The core structure of the microvillus within the eukaryotic cytoskeleton consists of highly ordered bundles of F-actin that extend straight from the cell’s interior into the microvillar extension. The plus ends of these actin filaments are strictly oriented toward the very tip of the protrusion. To maintain this tight, parallel configuration, specific cross-linking proteins, namely villin and fimbrin, tightly bundle the actin filaments together, preventing them from splaying apart under the mechanical stress of digestion.
The stability of this eukaryotic cytoskeleton extension is further reinforced by complex lateral and basal anchoring systems. Calmodulin and specialized myosin-like ATPases act as lateral anchors, physically connecting the central actin bundle to the microvillus’s surrounding plasma membrane. At the base of the protrusion, a protein called fodrin firmly anchors the actin fibers, heavily attaching them to a dense, underlying terminal web of intermediate filaments, which provides a massive, stable basal root network.
This intricate architectural masterpiece clearly demonstrates how distinctly different cytoskeletal elements collaborate perfectly to create highly resilient, specialized cellular appendages. The internal actin core provides the rigid, projecting shape; the specific cross-linkers maintain the strict structural bundle organization; and the deep intermediate filaments securely lock the entire structure into the cellular cortex. This comprehensive study perfectly illustrates the sheer mechanical ingenuity and structural beauty of biological design functioning within human physiology.
Slide 12: Eukaryotic Cytoskeleton: Guiding Intracellular Transport
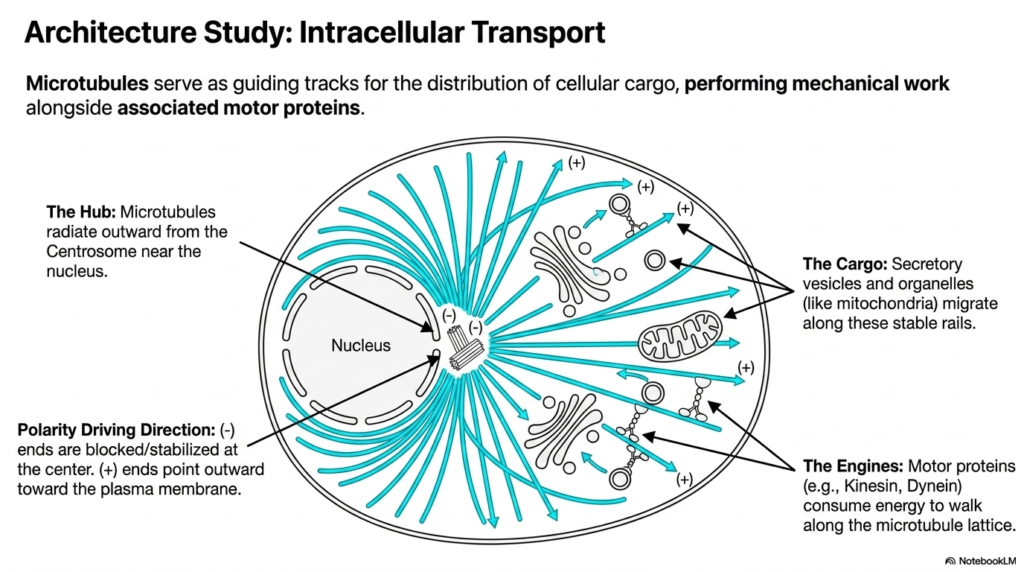
Moving beyond static structures, this architectural study highlights the eukaryotic cytoskeleton as an incredibly active transport highway. Microtubules serve as rigid, guiding tracks for the transport of cellular cargo, performing immense mechanical work directly alongside associated motor proteins. The hub of this transportation network is the centrosome, located centrally near the cell nucleus. From this central organizing center, individual microtubules radiate outward in all directions, establishing a comprehensive structural grid across the cytoplasm.
The structural polarity of the eukaryotic cytoskeleton absolutely dictates the driving direction of all intracellular transport. The minus ends of the microtubules are structurally blocked and firmly stabilized at the central centrosome hub. Conversely, the fast-growing plus ends constantly point outward toward the distant plasma membrane. This distinct polarization creates a highly predictable, one-way biochemical road system that allows motor proteins to perfectly navigate the complex, crowded internal environment of the living cell without losing their way.
Navigating this network within the eukaryotic cytoskeleton requires specialized molecular engines. Motor proteins, most notably kinesin and dynein, actively consume chemical energy derived from ATP hydrolysis to physically walk along the rigid microtubule lattice. These remarkable proteins haul massive molecular cargo, including vital secretory vesicles and entire organelles like mitochondria, migrating them safely along the stable rails. Kinesin generally drives cargo outward toward the plus ends, while dynein pulls cargo inward toward the minus ends.
This incredibly complex logistics network meticulously ensures that freshly synthesized biological proteins rapidly reach the cellular membrane for active secretion, and that massive, energy-producing mitochondria are strategically positioned exactly where cellular ATP demand is highest. Without this highly directed, motor-driven microtubular transport system, large complex cells would completely fail to distribute vital resources efficiently, inevitably leading to rapid metabolic collapse, severe physiological dysfunction, and ultimate cellular death.
Slide 13: Eukaryotic Cytoskeleton: Visualizing Complex Density In Vivo
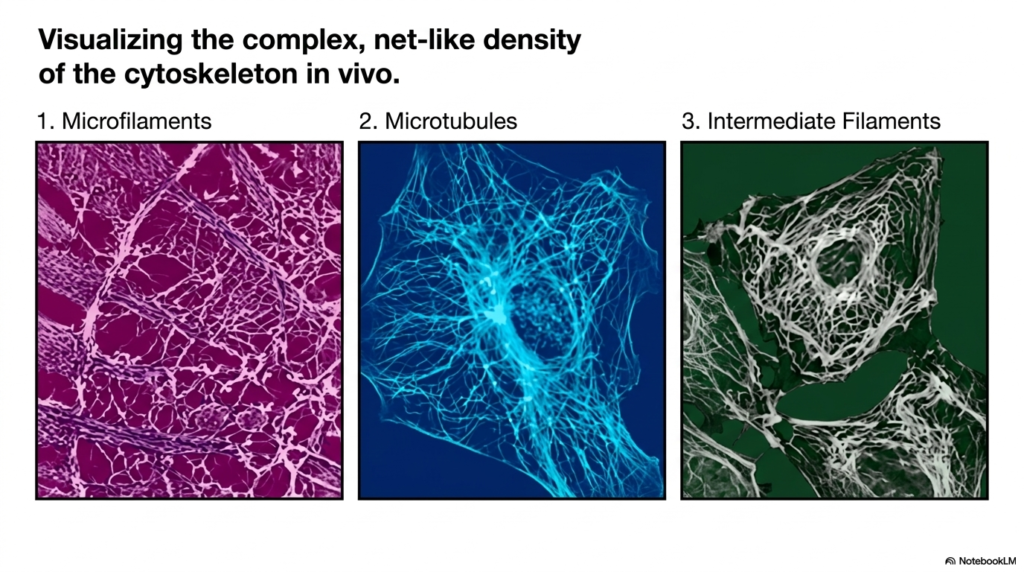
Beyond schematic diagrams, it is incredibly important to visualize the eukaryotic cytoskeleton as it exists in living cells. Modern fluorescence microscopy and advanced histological staining techniques allow scientists to vividly visualize the complex, net-like density of the cytoskeleton directly in vivo. These imaging modalities reveal that the cytoplasm is not a watery void, but rather a densely packed, highly structured jungle of interacting protein fibers that completely fills the intracellular space, defining the cell’s physical boundaries.
When specifically isolating the microfilaments of the eukaryotic cytoskeleton, one observes a dense, intricate webbing usually concentrated just beneath the plasma membrane. This cortical actin network forms a tight, protective mesh that maintains cell shape and drives surface motility. In stark contrast, visualizing the microtubule network reveals a stunning, star-like array of long, sweeping fibers radiating outward from the deep perinuclear region, creating a vast, organized grid designed explicitly for long-distance intracellular communication and heavy cargo transport.
Finally, imaging the intermediate filaments of the eukaryotic cytoskeleton unveils a robust, resilient meshwork that often surrounds the fragile nucleus and extends powerfully out to the cell periphery. This specific network looks like a thick, twisted biological wire system designed to resist massive physical stretching and cellular tearing. By comparing these three distinct in vivo images side by side, students can fully appreciate how these overlapping systems coexist within the same confined cytoplasmic space.
Viewing these breathtaking real-world biological images perfectly bridges the difficult gap between abstract biochemical theory and tangible, observable physiological reality. It deeply underscores the profound structural complexity and immense physical density of the intracellular environment, serving as a powerful visual reminder to students that modern cellular biology is essentially a study of highly organized, mechanically robust structural engineering occurring constantly at the absolute microscopic, molecular level.
Slide 14: Eukaryotic Cytoskeleton: The Biochemical Synthesis Matrix
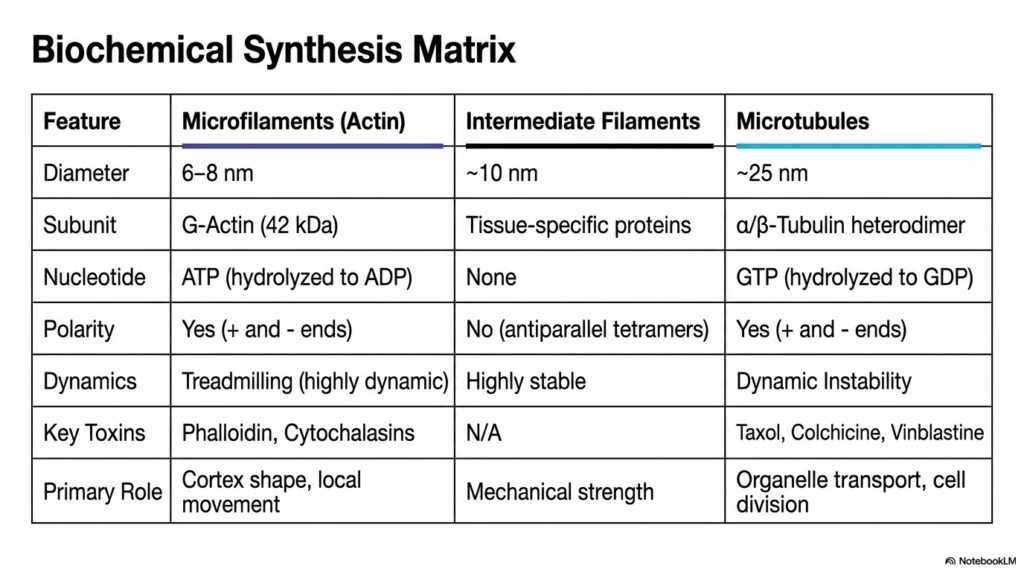
The final slide presents a comprehensive biochemical synthesis matrix that beautifully summarizes the defining properties of the eukaryotic cytoskeleton. This detailed comparative table serves as an ultimate review tool, systematically breaking down the complex features of microfilaments, intermediate filaments, and microtubules side by side. By directly comparing their distinct physical diameters, specific protein subunits, and required nucleotides, students can quickly synthesize the extensive biochemical data presented throughout the detailed structural slide deck.
A crucial comparative element within this eukaryotic cytoskeleton matrix is the concept of structural polarity and dynamics. The table explicitly highlights that while actin microfilaments and tubulin microtubules both possess distinct polarities and utilize high-energy nucleotides such as ATP and GTP to drive dynamic instability and treadmilling, intermediate filaments lack both polarity and nucleotide dependence, resulting in a highly stable, static network. This core biochemical distinction fundamentally dictates their entirely separate physiological roles within the cell.
Furthermore, the matrix outlines the extensive pharmacological vulnerabilities of the eukaryotic cytoskeleton by listing key toxins that disrupt each filament system. From the actin-binding properties of phalloidin to the microtubule-freezing action of Taxol, these chemical interactions reinforce the critical importance of cytoskeletal dynamics in maintaining cellular life. Finally, the matrix powerfully summarizes the primary physiological role of each system, directly linking molecular architecture to macroscopic biological function, such as maintaining cortex shape, mechanical strength, or organelle transport.
Mastering the dense comparative details carefully housed within this final synthesis matrix is absolutely essential for any serious academic student aspiring to a deep understanding of advanced human cellular biology, medical pathology, or clinical pharmacology. It brilliantly and concisely encapsulates the intricate biochemical diversity, the physical structural resilience, and the profound physiological synergy that ultimately defines the dynamic internal scaffolding system of all higher eukaryotic life forms currently found on Earth.
Please read our Content Disclaimer Statement.
Check out our social media channels: