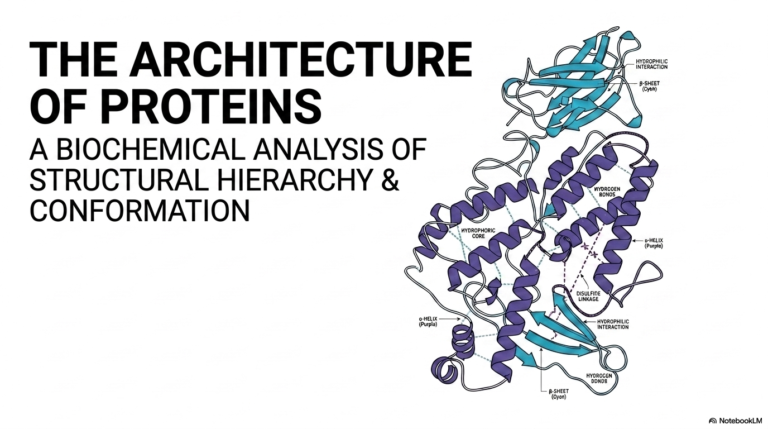
30. Coenzymes Explained: Functions, Types, and Enzyme Catalysis
Have you ever wondered how a simple carbohydrate physically transforms into the massive kinetic energy that completely powers a beating human heart? The incredible secret lies not just strictly in giant cellular proteins, but deeply within their essential, incredibly tiny chemical partners. This comprehensive academic slide deck systematically breaks down the complex foundational biochemistry of non-protein helpers, offering students a highly visual, extremely mechanistic guide to molecular catalysis. Understanding these highly dynamic, incredibly vital structures reveals exactly how human cells survive, adapt, and generate life-sustaining energy.
Slide 1: Defining the Biochemistry of Coenzymes
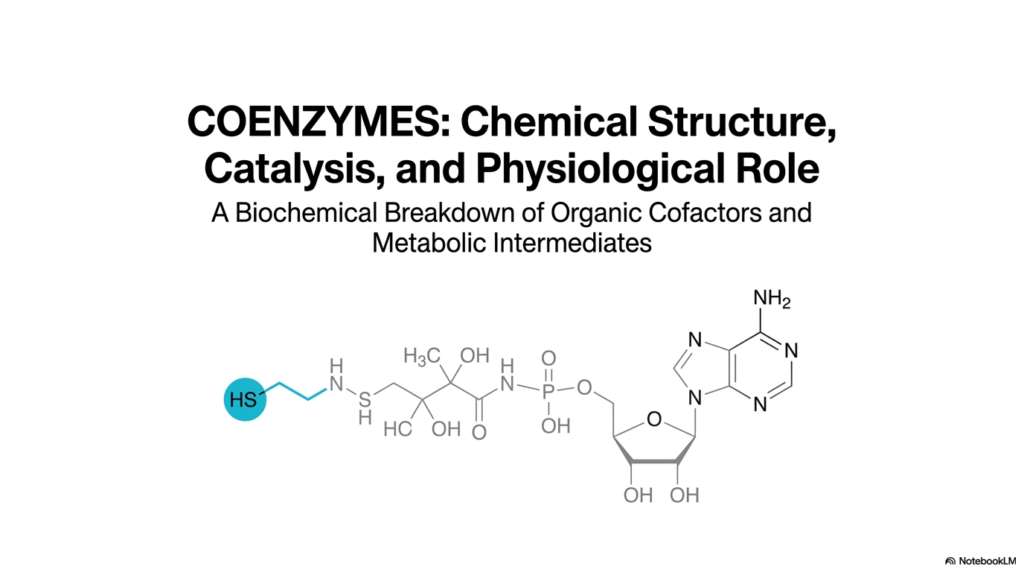
The foundational exploration of molecular catalysis begins here by deeply examining the absolutely vital chemical partners of cellular proteins. In advanced biochemistry, understanding exactly how massive biological enzymes actively accelerate chemical reactions is strictly only half the academic battle. The other crucial half strictly involves a deep exploration of the non-protein molecules that make these extremely complex reactions thermodynamically possible. This very first slide directly visually introduces the overarching academic theme of this entire series: the complex chemical and physical structure, detailed catalytic mechanisms, and broad physiological and cellular roles of coenzymes.
To truly master this highly complex subject, college students must quickly recognize that coenzymes are strictly not merely passive structural components. Instead, they are highly active, extremely dynamic chemical cellular reagents that aggressively chemically participate in vital metabolic reactions. The beautiful cover slide visually emphasizes this core concept by prominently featuring the intricate chemical structure of a specific molecule, highlighting its distinct functional reactive groups. Notice the dense nucleotide chemical handle, the complex vitamin-derived structural linker, and the highly reactive terminal group.
Furthermore, exploring these highly intricate structures provides a crucial bridge between basic microscopic chemical reactions and complex human physiology. The complex organic molecular structures prominently featured in this opening slide are essentially the active, highly functional chemical forms of the daily essential vitamins routinely ingested through a strictly normal, healthy human diet. Therefore, a deeply thorough, highly mechanistic academic understanding of these exact chemical molecular structures is absolutely necessary for accurately identifying and successfully comprehensively robustly cleanly treating complex metabolic diseases.
Slide 2: The Architecture of Catalysis and Coenzymes
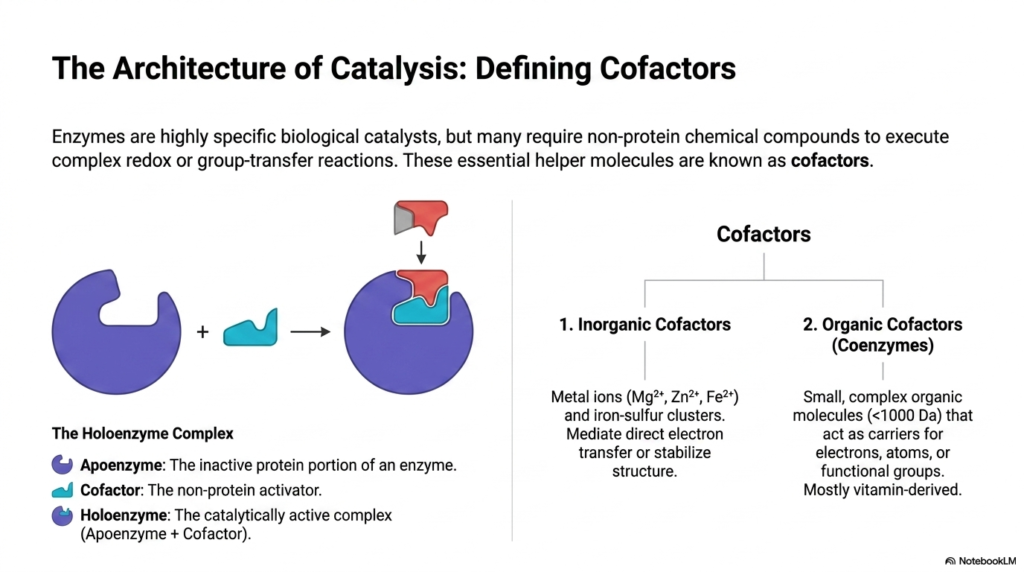
Enzymes are universally renowned for their incredible substrate specificity and extreme catalytic efficiency across biological systems. However, many of these massive biological catalysts are chemically inadequate on their own. They heavily require the physical assistance of non-protein chemical compounds to execute complex redox or challenging group-transfer reactions. This slide introduces the fundamental hierarchy of these essential helper molecules. At the top foundational level exists the strictly inactive protein portion known as the apoenzyme. When the necessary non-protein activator binds securely to this apoenzyme, it successfully forms the fully functional holoenzyme complex.
These vital non-protein activators are broadly classified into two distinct biochemical categories: simple inorganic cofactors and complex organic cofactors. Inorganic cofactors typically consist of essential metal ions, such as magnesium, zinc, or iron, which mediate direct electron transfer or physically stabilize the active-site structure. On the other hand, organic cofactors are specifically called coenzymes. These are small, highly complex organic molecules that perform remarkably sophisticated chemical duties within the active site. This fundamental distinction is critical for students striving to understand the mechanics of metabolic pathways.
While metal ions provide raw electrostatic pulling power, coenzymes act as specialized, highly dynamic carriers for specific electrons, individual atoms, or entire functional chemical groups. It is structurally fascinating to note that most of these intricate organic molecules are derived directly from essential dietary vitamins. This vital biological connection highlights exactly why specific vitamins are essential for human survival, serving as the molecular building blocks that make the holoenzyme fully functional. Understanding this precise architectural assembly provides deep, mechanistic insight into cellular control.
Slide 3: Binding Dynamics: Soluble Coenzymes vs. Prosthetic Groups
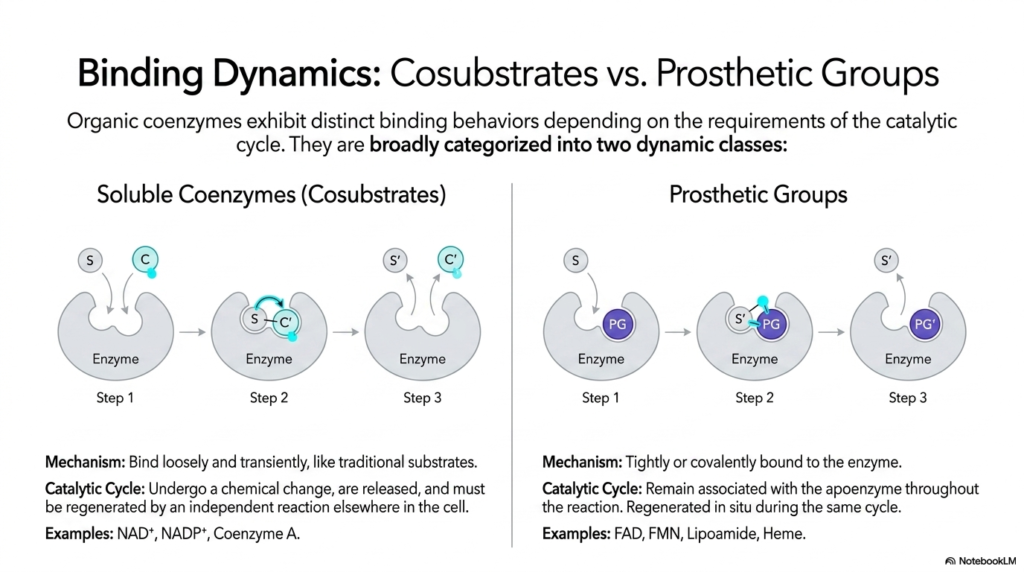
Once the basic architectural definitions are firmly established, the next crucial biochemical concept concerns how these specific molecules interact with their massive enzyme partners. Organic coenzymes exhibit distinctly different binding behaviors depending entirely on the specific chemical requirements of the catalytic cycle. This slide beautifully categorizes them into two highly dynamic functional classes based on their physical affinity and overall permanence within the active site. Understanding this precise structural distinction is vital for accurately tracing exactly how various chemical intermediates flow through different, highly complex metabolic pathways.
The first major class, functionally known as highly soluble coenzymes or transient cosubstrates, behaves very much like traditional enzyme substrates. They bind loosely and transiently to the specific protein’s active site. During the active catalytic cycle, these molecules undergo a highly specific chemical change and are subsequently released entirely from the enzyme. Because they are actively released into the cellular cytosol in an altered chemical state, these molecules must be fully regenerated by an independent chemical reaction elsewhere before they can be reused.
In stark structural contrast, the second dynamic class consists entirely of highly permanent prosthetic groups. These specialized coenzymes are incredibly tightly, and sometimes even covalently, bound directly to the large apoenzyme structure. Unlike highly mobile, transient cosubstrates, rigid prosthetic groups absolutely do not leave the biological active site. They remain permanently and securely associated with the host enzyme throughout the entire, continuous biochemical reaction cycle. Because they are permanently fixed in place, any internal chemical change they undergo must be fully reversed in situ, effectively allowing the massive enzyme to rapidly reset.
Slide 4: Pyridine Nucleotide Coenzymes in Redox Reactions
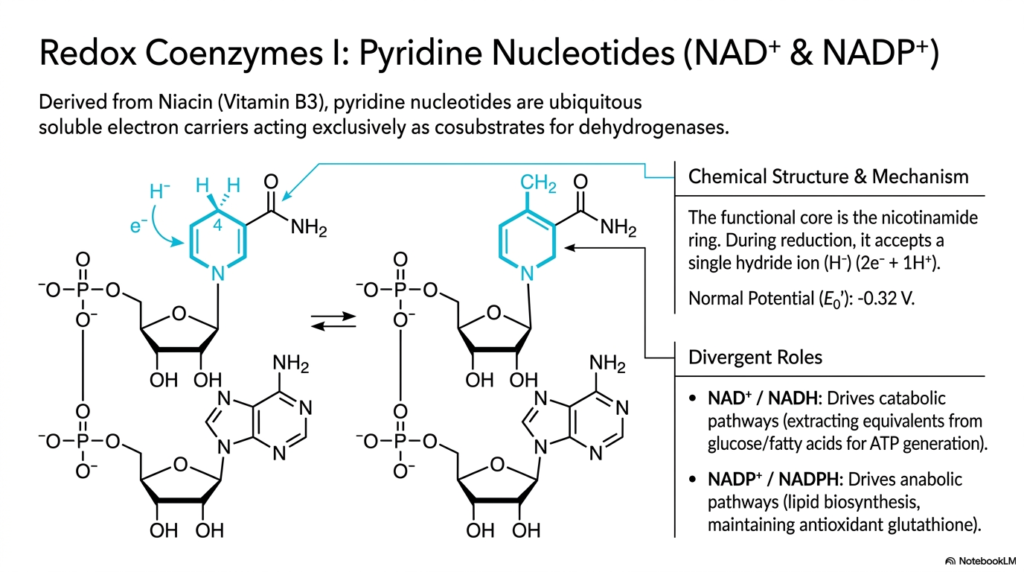
Delving deeply into highly specific molecular families, this detailed slide closely examines the absolutely essential redox coenzymes, commonly known as pyridine nucleotides, highlighting the highly mobile NAD+ and NADP+. Synthesized biochemically straight from Niacin, also known clinically as vital Vitamin B3, these complex molecules are ubiquitous, highly soluble biological electron carriers. They act exclusively as transient cosubstrates strictly for a highly specific, vital class of metabolic enzymes called dehydrogenases. Because they are freely soluble, they readily diffuse through the highly aqueous cellular cytosol, linking entirely different metabolic pathways.
The primary chemical functionality of these ubiquitous coenzymes centers on their unique, chemically reactive nicotinamide ring. The overall molecular structure features a bulky nucleotide base, a standard ribose sugar, and bridging phosphate groups, but the aromatic nicotinamide ring is the actual active reactive center. During a highly targeted cellular reduction reaction, this highly specific ring forcefully accepts a single, highly energetic hydride ion, which consists of two heavy electrons and one proton. This specific chemical mechanism gives pyridine nucleotides a normal standard reduction potential of roughly -0.32 Volts.
Despite their nearly identical, incredibly complex chemical structures, which physically differ only by a single small phosphate group, these two pyridine nucleotide coenzymes serve completely divergent physiological roles. The NAD+ molecular pool is primarily responsible for driving cellular catabolic pathways, physically extracting high-energy electrons directly from the breakdown of glucose to generate large amounts of ATP. Conversely, the heavily phosphorylated NADP+ pool is utilized almost exclusively for vital anabolic pathways, driving massive lipid biosynthesis and actively maintaining critical cellular antioxidant systems to prevent severe oxidative stress.
Slide 5: Flavin Coenzymes and Radical Intermediates
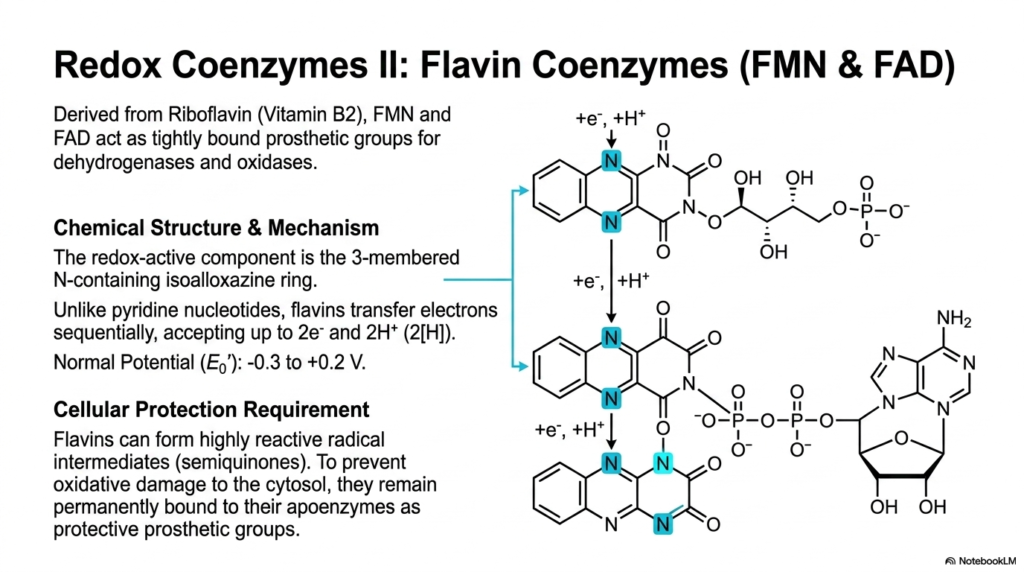
The next major structural class of highly vital redox carriers directly includes the complex flavin coenzymes, specifically highlighting the abundant Flavin Mononucleotide (FMN) and the dense Flavin Adenine Dinucleotide (FAD). These absolutely crucial metabolic molecules are synthesized directly from Riboflavin, commonly known clinically as essential Vitamin B2. Unlike the freely diffusing, highly mobile pyridine nucleotides previously discussed at length, bulky flavins predominantly act as tightly bound, highly permanent prosthetic groups strictly for specific cellular dehydrogenases and oxidases. This fundamental physical difference dictates their highly specific physiological roles as safely localized reactive centers.
The extreme chemical prowess of these particular coenzymes stems directly from their unique, incredibly complex three-membered, nitrogen-containing isoalloxazine aromatic ring. This completely complex, dense ring structure provides a highly significant mechanistic advantage over strictly simpler cellular electron carriers. While highly soluble pyridine nucleotides must forcefully accept a massive full hydride ion instantly, the physically versatile isoalloxazine ring can seamlessly and actively accept heavy electrons sequentially. It can easily and rapidly transfer either a single electron at a time or two heavy electrons simultaneously, giving these molecules a broad, adaptable range of reduction potentials.
This distinct biological ability to seamlessly transfer individual single electrons is exactly what makes flavin coenzymes uniquely strictly essential, but it directly introduces a highly significant and dangerous cellular threat. By actively accepting single heavy electrons, dense flavins can rapidly form highly reactive, completely unstable radical intermediates structurally known as dangerous semiquinones. If these highly specific radical species were ever carelessly allowed to float freely in the aqueous cellular cytosol, they would instantly violently react with dissolved oxygen. Remaining permanently tightly bound directly prevents this massive oxidative damage to the fragile cell.
Slide 6: Lipid and Water-Soluble Redox Coenzymes
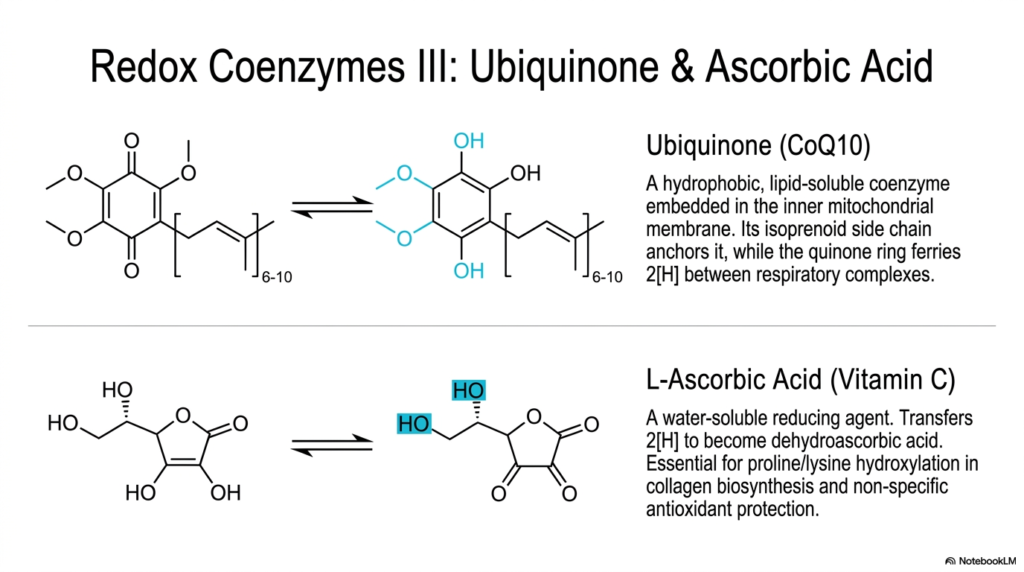
This slide shifts the academic focus to two distinct yet equally vital specialized cellular electron carriers: the highly hydrophobic Ubiquinone and the incredibly hydrophilic Ascorbic Acid. While previous detailed sections focused heavily strictly on complex, bulky nucleotide derivatives, this slide highlights highly structurally unique coenzymes that deliberately operate in vastly entirely distinct biochemical cellular environments. Ubiquinone, frequently strictly referred to clinically as Coenzyme Q10, is a highly hydrophobic, distinctly heavy lipid-soluble molecule. It physically does not exist freely in the cellular cytosol, but remains strictly embedded within the inner membrane.
The precise dense chemical structure of Ubiquinone perfectly matches its highly specific, deeply physically isolated biological function. It possesses a notably long, repeating heavy isoprenoid side chain that physically acts as a highly secure chemical anchor, keeping the massive molecule firmly tethered within the shifting lipid bilayer. Meanwhile, the highly functional, incredibly reactive aromatic quinone ring is solely responsible for rapidly accepting and transferring highly energetic electrons. As vital moving components of cellular respiration, these unique coenzymes act as highly mobile lipid-based junctions between massive protein complexes.
In stark, totally distinct structural contrast, small L-Ascorbic Acid, widely known globally clinically as essential Vitamin C, is an intensely completely water-soluble, highly simple reducing agent. This tiny carbohydrate derivative physically functions quite remarkably differently from its massive lipid-bound counterparts. Ascorbic acid rapidly transfers crucial cellular reducing equivalents, rapidly becoming oxidized to dehydroascorbic acid. The overall vast physiological biochemical roles of these water-soluble coenzymes are highly significant and entirely structurally significant for medical students to master. They are strictly absolutely essential specific cofactors for the specialized enzymes responsible for successfully synthesizing strong human collagen.
Slide 7: Inorganic and Hybrid Redox Coenzymes
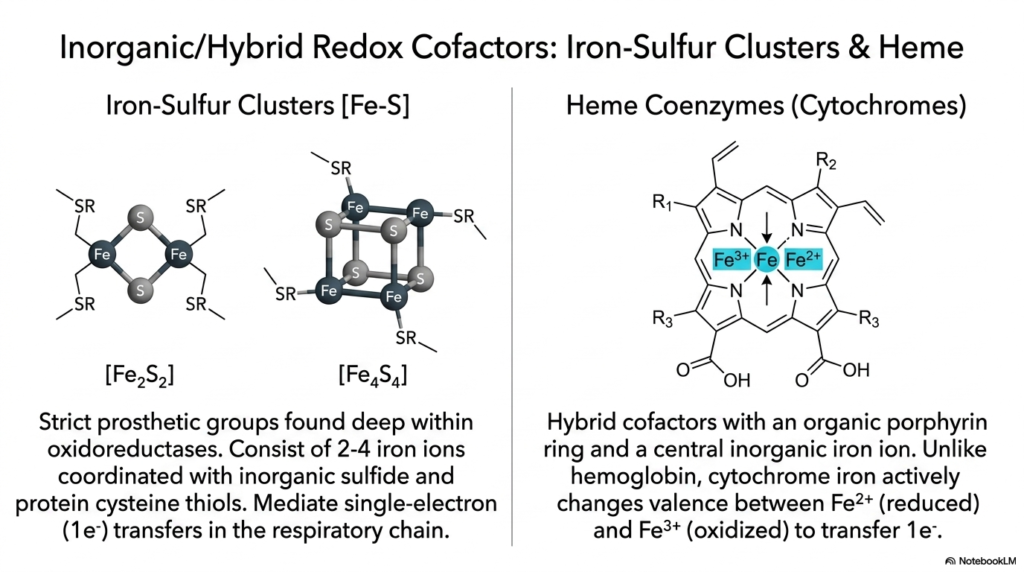
Expanding deeply and structurally well beyond strictly organic molecules, this detailed slide directly and visually introduces highly specialized, complex inorganic and dense, complex hybrid cofactors that are equally crucial for widespread cellular redox biology. First, the intense structural focus shifts directly to incredibly dense Iron-Sulfur clusters, which strictly function in biology as extremely tightly bound, permanently bound prosthetic groups located deep within massive human oxidoreductase enzymes. While strictly inorganic, they function seamlessly alongside traditional organic coenzymes. These remarkable, ancient, dense inorganic structures consist of multiple heavy coordinated iron ions tightly bound together with inorganic sulfide.
Mechanistically, these incredibly dense Iron-Sulfur clusters are highly specialized for their highly specific, physically challenging biological task. Unlike highly complex, fully organic coenzymes that can seamlessly transfer massive full hydrides or entire bulky chemical groups, these extremely rigid inorganic metal clusters explicitly and forcefully mediate only extremely fast, strictly single-electron biological transfers. The heavy individual iron atoms within the highly dense inorganic cluster rapidly and continuously cycle between distinct, highly specific oxidation states to cleanly and safely pass individual electrons straight down a steep energetic gradient.
The detailed molecular breakdown then strictly and heavily transitions directly to completely unique, massive Heme groups, which are chemically best described as incredibly complex hybrid cellular cofactors. Massive Heme molecules directly feature a massive, beautifully dense complex organic porphyrin carbon ring that firmly and securely holds a deeply reactive central heavy inorganic iron ion. These specific, absolutely intricate coenzymes perfectly, completely, seamlessly blur the highly traditional line exactly between purely standard organic molecules and basic, simple inorganic transition metals, completely combining massive structural physical stability with highly potent heavy redox chemistry.
Slide 8: Nucleoside Phosphates as Group-Transfer Coenzymes
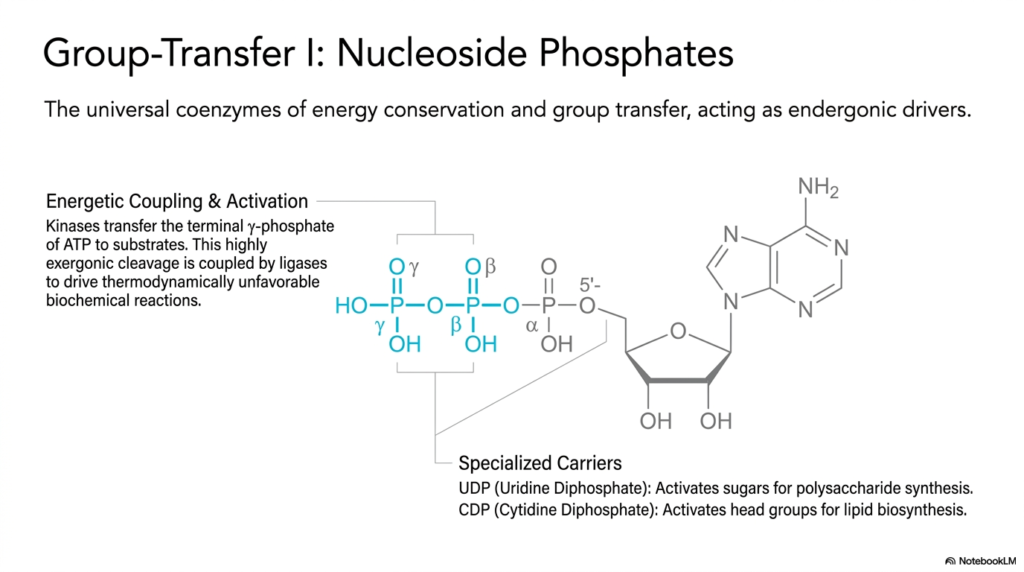
Aggressively shifting deep gears, completely and thoroughly away from strict traditional redox reactions, the detailed structural academic exploration now covers the vast, highly complex biochemical metabolic landscape of essential group-transfer biological reactions. This dense, detailed slide specifically highlights heavy, bulky nucleoside phosphates, which universally serve as the primary foundational biological drivers of immense energy conservation and direct complex molecular physical transfer. While they are frequently simply colloquially referred to as generic cellular energy molecules, they function, physically accurately, entirely mechanistically as incredibly highly specialized, complex coenzymes that forcefully activate targets.
The specific, complex, intense chemical-energetic mechanism of ATP is a breathtaking, absolute masterpiece of direct physical, active, energetic coupling. Highly specific cellular metabolic enzymes called kinases constantly and vigorously catalyze the rapid transfer of the terminal gamma-phosphate strictly from ATP directly onto highly specific waiting target biological substrates. The highly crucial foundational biochemical detail perfectly explored here is that the highly targeted, violent cleavage of these specific heavy coenzymes is intensely exergonic. This successfully provides the massive energetic push required to drive highly physically unfavorable cellular biological machinery.
Beyond standard ubiquitous ATP, other heavy, dense nucleoside phosphates actively, explicitly, and safely act as highly specialized biological carriers strictly for entirely different, highly isolated human metabolic pathways. For a direct, distinct example, Uridine Diphosphate, formally known as UDP, is chemically perfectly tailored to activate simple, small, single sugars for the synthesis of massive, dense complex polysaccharides. Without this highly, incredibly, extremely specific, absolutely, strictly, thoroughly required biological activation rapid step via these highly unique, complex coenzymes, complex human cellular structural carbohydrate massive synthesis would completely, entirely, fatally fail.
Slide 9: The Structural Organization of Coenzyme A
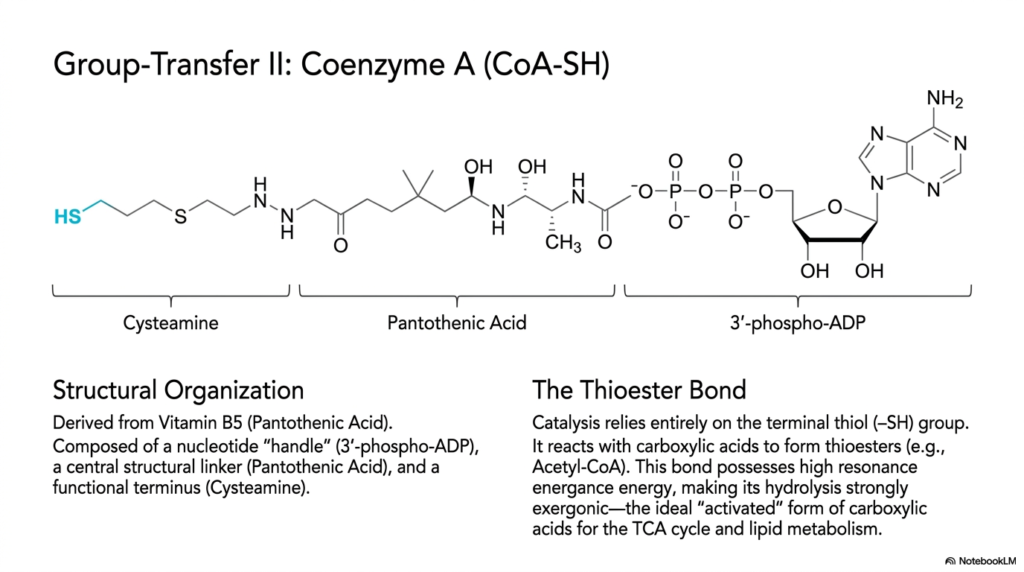
Slide nine deeply and systematically delves into one of the most clinically famous and heavily centralized giant molecules in all of human biochemistry: Coenzyme A, frequently abbreviated as CoA-SH. This incredibly massive, structurally complex molecule is the ultimate, unrivaled biological group-transfer specialist, specifically tasked with safely managing and actively moving highly reactive acyl groups throughout the complex human cell. It directly serves as a massive metabolic structural crossroads for cellular bioenergetics. To fully understand exactly how these specific, giant coenzymes operate, biochemistry students must first carefully examine their unique, beautifully massive tripartite molecular structure.
The overall physical architecture of massive Coenzyme A is brilliantly and logically composed of three fully distinct functional chemical zones. First, it directly features a massive nucleotide handle, specifically a bulky 3′-phospho-ADP molecule. This significantly large molecular handle is primarily used for direct enzyme recognition, enabling various large cellular proteins to firmly grasp the complex molecule. Second, it securely contains a massive central structural vitamin linker derived directly from Pantothenic Acid, also clinically known as Vitamin B5. Finally, the giant organic molecule physically completely terminates in a highly reactive, functional cysteamine terminal end.
The entire massive catalytic energetic purpose of this incredibly giant molecule relies completely on one single, remarkably tiny functional group: the highly reactive terminal thiol, or sulfhydryl (-SH), group located strictly at the extreme distal end of the flexible cysteamine arm. This highly reactive sulfur atom readily and forcefully reacts with simple, small carboxylic acids to form absolutely crucial chemical and biological intermediates, universally known as high-energy thioesters. The high resonance energy specifically found within these giant coenzymes makes rapid hydrolysis violently exergonic, thereby supplying substantial activation energy directly for human lipid metabolism.
Slide 10: Decarboxylation and Transamination Coenzymes
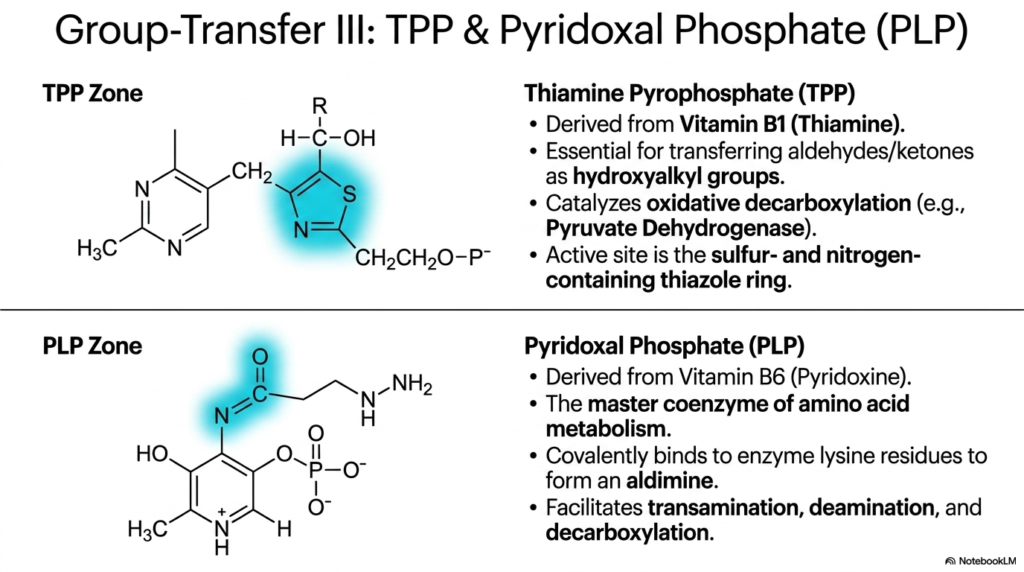
Continuing the deep, highly detailed structural exploration of complex group-transfer biochemistry, this slide directly introduces two highly specialized reactive molecules strictly responsible for safely and forcefully manipulating massive, heavy carbon skeletons: Thiamine Pyrophosphate (TPP) and Pyridoxal Phosphate (PLP). First, the complex physical structure of TPP is thoroughly examined; it is synthesized biologically entirely directly from dietary Thiamine, clinically known globally as the vital Vitamin B1. TPP is an absolute, strict critical chemical requirement specifically for massive biological multienzyme complexes that intensely transfer highly reactive, dangerous aldehydes directly as bulky hydroxyalkyl groups during fundamental cellular metabolism.
The complete chemical mechanistic magic of highly specific TPP occurs exclusively within its highly rigid, uniquely structured biologically active site: the incredibly complex, reactive thiazole ring that directly contains both highly reactive, dense sulfur and heavy nitrogen atoms. The incredibly specific, highly precise, dense spatial arrangement of these uniquely reactive atoms absolutely seamlessly allows the dense ring specifically to successfully, rapidly, forcefully form a highly stabilized, incredibly dangerous reactive carbanion intermediate. These extremely specialized, beautifully rigid coenzymes ultimately enable crucial multienzyme complexes to cleanly rip individual carbon atoms from specific target molecules.
In the distinctly lower visual zone of the slide exists highly reactive Pyridoxal Phosphate, universally abbreviated as crucial PLP, which is synthesized directly from Pyridoxine (Vitamin B6). PLP is universally recognized in clinical practice as the undisputed master coenzyme of complex human amino acid metabolism. Its specific chemical mechanism is beautifully complex: PLP covalently binds deeply to specific rigid lysine residues within its host apoenzyme to form a highly reactive intermediate structure called a Schiff base. By stabilizing highly reactive, unique intermediates, these highly functional, versatile coenzymes effortlessly facilitate a staggering variety of essential biological transamination reactions.
Slide 11: Biotin and Tetrahydrofolate Coenzymes in Carbon Transfer
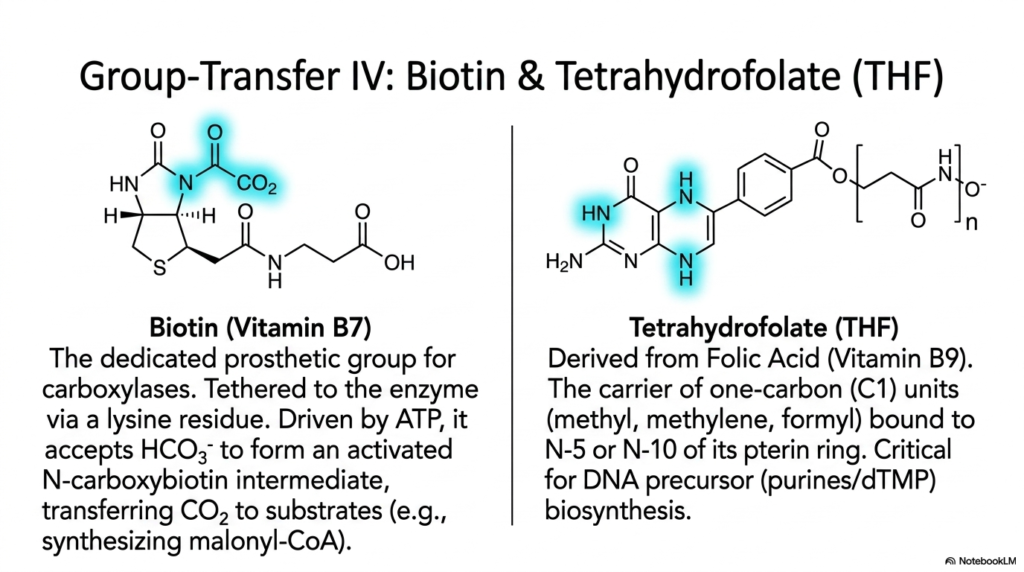
Slide eleven strictly and aggressively tackles the incredibly intricate, physically delicate structural biochemistry of safely transferring incredibly tiny, highly reactive single carbon atoms across cellular spaces. This slide specifically structurally features both Biotin and complex Tetrahydrofolate (THF). Biotin, widely recognized clinically as the essential dietary Vitamin B7, functions chemically as a highly unique, dedicated, permanently tethered prosthetic group for the carboxylase enzymes of humans. Biotin is securely tethered to its massive host protein enzyme via a notably long, highly flexible biological lysine residue, thereby creating a rapidly swinging, massive arm.
Strictly physically and mechanistically, this incredibly specific, dense molecule actively and rapidly accepts a fully oxygenated inorganic bicarbonate ion to forcefully form an incredibly highly activated, heavily negatively charged N-carboxybiotin intermediate. The incredibly long, completely physically flexible biological lysine molecular swinging arm then forcefully and aggressively swings this massive, highly activated carbon dioxide payload directly across the entire giant enzyme protein complex. These incredible swinging-arm coenzymes are strictly, absolutely, fundamentally, and biologically essential for actively, forcefully, physically driving thermodynamically extremely, highly unfavorable, massively complex early fatty acid cellular biosynthesis human pathways rapidly, structurally, forward.
The distinct right visual side of the slide strictly introduces Tetrahydrofolate, universally abbreviated as crucial THF, the highly active, complex derivative of Folic Acid (Vitamin B9). While Biotin strictly handles heavily oxidized, large carbon dioxide molecules, THF is the ultimate, incredibly versatile mobile carrier of various other, incredibly tiny one-carbon units, specifically operating directly in various intermediate redox states, such as the highly reactive methyl group. These highly versatile, mobile-specific coenzymes act as vital cellular single-carbon donors. Without an absolutely sufficient number of complex THF structures physically present, massive cellular DNA replication processes grind to an absolute halt.
Slide 12: Cobalamin Coenzymes and Organometallic Chemistry
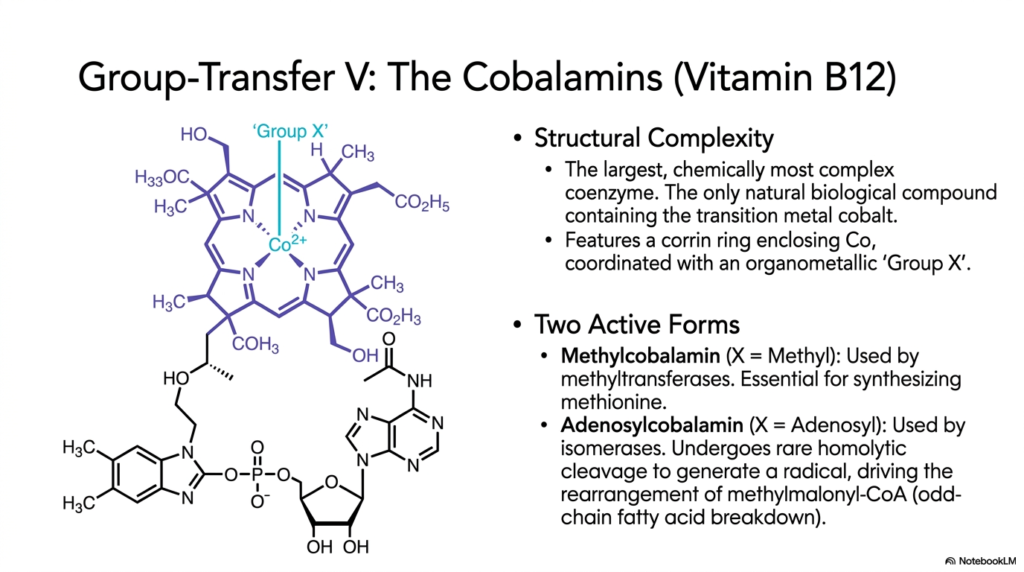
The highly detailed twelfth slide directly and deeply highlights the most incredibly structurally magnificent and complex giant molecule found in the entire human metabolic chemical repertoire: the highly heavy Cobalamins, derived strictly from the vital Vitamin B12. From a strict structural complexity standpoint, these massive biological structures are absolutely the largest and most chemically intricate giant coenzymes found in nature. Remarkably, they are the only naturally occurring biological compounds that strictly contain cobalt. The unique functional chemical identity of the top-attached organometallic group strictly determines the highly specific complex biochemical and physiological function.
The first primary, highly active form strictly utilized by the human body is heavy Methylcobalamin, in which the firmly attached, highly reactive functional top group is a simple, incredibly tiny methyl carbon group. These deeply particular, highly complex, heavy, specific coenzymes are strictly, entirely, absolutely, and exclusively utilized directly by massive, complex methyltransferase metabolic human enzymes. They are absolutely critically required for efficiently synthesizing essential amino acids like methionine. By safely and securely physically handling these tiny, highly reactive methyl radicals, the massive complex molecule completely protects the fragile internal cellular environment from widespread, fatal oxidative chemical damage.
The distinctly second, highly unique active form strictly utilized is absolutely massive Adenosylcobalamin, where a highly bulky, massive adenosyl group firmly acts as the vital active organometallic-attached ligand. Mechanistically, the highly complex enzymatic chemical reaction is aggressively initiated deep within by a profoundly rare, entirely unusual violent homolytic physical cleavage of the strict chemical bond directly between the heavy central cobalt transition-metal atom and carbon. This massive radical chemical mechanism is strictly absolutely essential for actively physically rearranging highly complex methylmalonyl-CoA. Without these incredibly intricate heavy coenzymes, complex odd-chain massive fatty acid metabolism completely fails.
Slide 13: Lipoamide Coenzymes as Dual-Action Prosthetic Groups
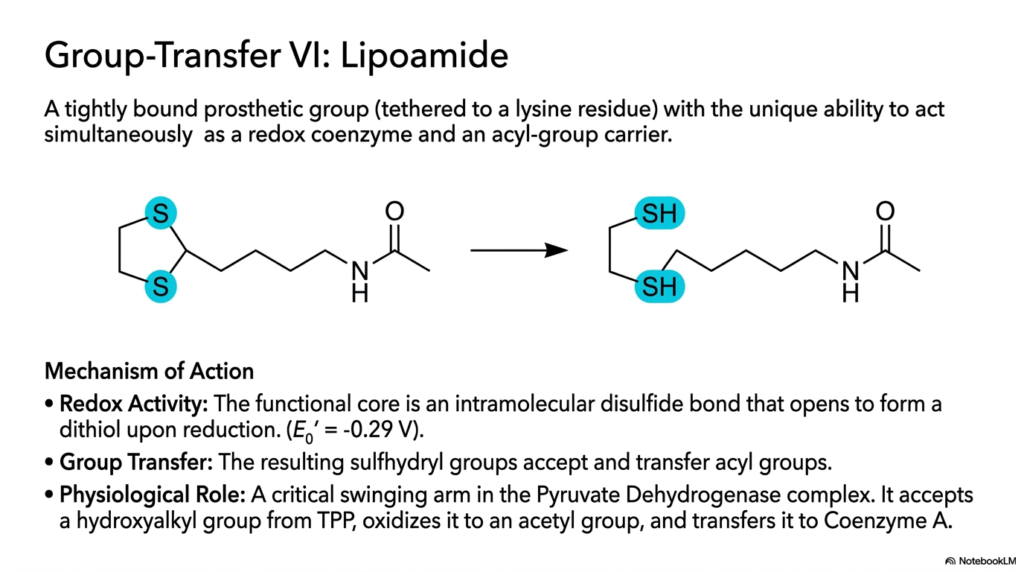
Slide thirteen specifically introduces a truly spectacular structural marvel of highly complex, deep metabolic-physical integration: the unique heavy Lipoamide group. This profoundly unique, massive molecule is specifically an incredibly tightly bound, heavy, permanent prosthetic group, chemically tethered directly and strongly to a highly specific, long, flexible protein lysine residue. What truly, entirely, perfectly, and highly distinguishes these specific, complex, tightly bound coenzymes is their remarkable, highly versatile dual biological action. They are physically capable of efficiently performing entirely distinct, massive chemical roles, sequentially and strictly, within the exact same massive, continuous biological enzymatic catalytic reaction cycle.
The profoundly complex, heavy, precise mechanism of Lipoamide’s chemical action is entirely dependent on its uniquely reactive functional structural core: an incredibly tightly constrained, active intramolecular disulfide bond. When acting strictly inside its primary highly active redox capacity, this critical deep active bond forcefully and rapidly accepts highly specific electrons to cleanly and efficiently form a fully reduced dithiol intermediate. This highly specific biological chemical reduction rapidly structurally primes the entire massive swinging tethered molecular arm for its absolutely crucial, entirely distinct second major metabolic biological job, directly transitioning the entire massive molecule into a highly aggressive group-carrier mode.
Once fully reduced, the highly reactive terminal sulfhydryl groups rapidly and forcefully accept bulky, high-energy acyl groups directly from adjacent tightly associated coenzymes residing entirely within the massive multi-enzyme complex. Specifically, in the giant Pyruvate Dehydrogenase complex, this remarkably long, flexible, swinging lipoamide arm efficiently accepts a large hydroxyalkyl group exclusively from the firmly bound TPP. It oxidizes this specific heavy substrate incredibly rapidly, directly into a highly vital, fiercely reactive acetyl group. Finally, the swinging arm rapidly and completely transfers this critical energetic acetyl group onto waiting mobile Coenzyme A to continue cellular energy production.
Slide 14: The Symphony of Coenzymes in the Metabolic Network
In the vast and highly dynamic landscape of cellular metabolism, individual biochemical reactions absolutely do not occur in complete isolation. Instead, they are deeply interconnected, continuously forming a massive, continuous chemical network. This slide provides a crucial macroscopic view of how different coenzymes precisely synchronize to drive the massive energy-harvesting pathways of the human cell. The true beauty of biochemistry lies strictly in observing how these diverse organic cofactors successfully operate sequentially, rapidly passing chemical intermediates and high-energy electrons down the line like a highly coordinated molecular relay race.
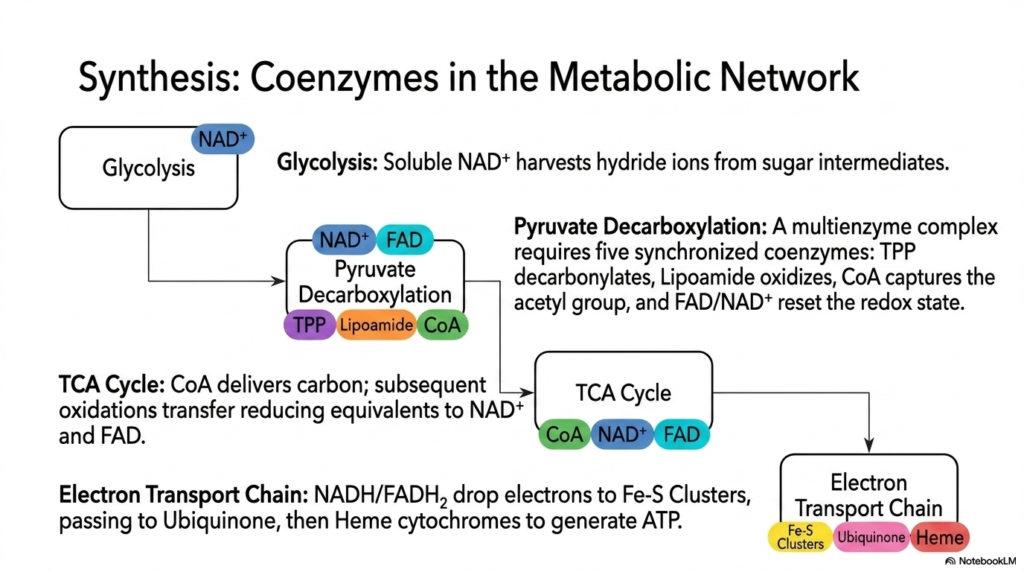
The biochemical journey begins with standard glycolysis in the cellular cytosol, where the simple breakdown of sugar intermediates relies entirely on soluble NAD+. As the basic glucose molecule is physically split and heavily oxidized, these highly mobile coenzymes rapidly harvest crucial hydride ions. The resulting NADH molecules then aggressively ferry these essential reducing equivalents directly toward the mitochondria. This initial biological step perfectly demonstrates how freely diffusing electron carriers link isolated cytoplasmic reactions to the broader, massive energy-generating machinery located deep within the cell.
Next, the metabolic pathway physically transitions strictly into the deep mitochondria for the incredibly complex process of pyruvate decarboxylation. This highly pivotal transition strictly requires a massive multienzyme complex that brilliantly relies on five perfectly synchronized coenzymes to function. TPP rapidly decarbonylates the waiting substrate, while a highly flexible swinging Lipoamide arm quickly oxidizes it. Coenzyme A successfully captures the resulting reactive acetyl group, while tightly bound FAD and highly mobile NAD+ effectively reset the giant enzyme’s internal redox state. This spectacular chemical choreography perfectly highlights their essential, highly collaborative nature.
Following this massive step, the newly generated Acetyl-CoA is delivered directly and rapidly to the famous TCA Cycle (Krebs Cycle), where it provides its vital two-carbon payload. Throughout this continuous circular biochemical pathway, intense oxidations occur systematically, transferring massive amounts of reducing equivalents directly to large waiting pools of NAD+ and FAD. These specific coenzymes act as immense energetic sponges, aggressively soaking up nearly all the available high-energy electrons originally trapped within the carbohydrate structure, completely priming the entire cell for the massive final stage of biological energy conservation.
Finally, all these successfully harvested electrons are forcibly delivered directly to the massive Electron Transport Chain, which is entirely embedded deep within the inner mitochondrial membrane. Here, NADH and FADH2 rapidly drop their electrons off directly to dense inorganic Iron-Sulfur clusters. The highly lipid-soluble Ubiquinone efficiently ferries them rapidly between massive membrane-bound protein complexes, cleanly handing them off to complex Heme cytochromes. This seamless, highly ordered molecular handoff between completely different coenzymes strictly actively drives the massive proton pump absolutely required for ultimate cellular ATP synthesis, beautifully completing the entire metabolic network.
Slide 15: Clinical Pathologies and the Crucial Need for Coenzymes
The incredible biochemical elegance of cellular metabolism, unfortunately, comes with a highly significant evolutionary trade-off, particularly in human biology. Because humans lack the necessary complex biological machinery to synthesize most of these absolutely essential molecules de novo, they must obtain the critical vitamin precursors directly through a healthy, highly balanced diet. This final, highly crucial slide bridges the gap between microscopic molecular biology and macroscopic human pathology, illustrating exactly what inevitably occurs clinically when the body suffers a massive, systemic deficit of these vital coenzymes.
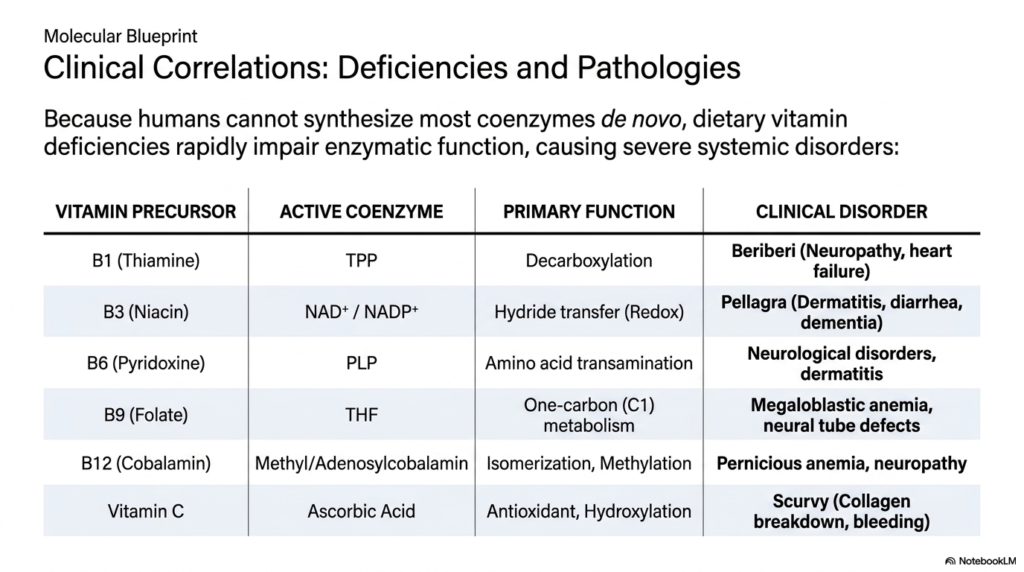
When direct dietary intake of specific essential vitamins is completely absent, the corresponding active coenzymes cannot be synthesized by the body. Consequently, the massive, intricate metabolic networks we previously explored completely grind to a highly destructive, violent halt. For instance, a severe dietary deficiency in the vital Vitamin B1 (Thiamine) rapidly and heavily depletes cellular TPP. Without this highly specific molecule, essential oxidative decarboxylation simply ceases, violently starving highly active bodily tissues of vital cellular energy. This directly manifests clinically as the severe disease Beriberi, characterized heavily by rapid neurological degeneration and massive, fatal heart failure.
Similarly, a severe, prolonged lack of Niacin (Vitamin B3) directly and violently impairs the massive cellular pool of highly mobile NAD+ and NADP+. Because these ubiquitous, essential coenzymes are absolutely required for nearly every major redox reaction in the body, a severe deficiency violently disrupts global human metabolism. The resulting clinical condition, known historically and clinically as Pellagra, is devastatingly characterized by the classic, severe symptoms of massive dermatitis, massive uncontrollable diarrhea, and rapidly progressive clinical dementia, directly showcasing the massive systemic importance of these molecules.
The specific dietary lack of other crucial vitamin precursors causes highly targeted, massive clinical failures. A severe deficiency in Folate (Vitamin B9) strictly starves the body of essential THF coenzymes, rapidly stopping critical single-carbon cellular transfers. Because rapidly dividing cells absolutely require this exact mechanism for successful massive DNA precursor biosynthesis, affected patients rapidly develop severe, highly dangerous megaloblastic anemia. Furthermore, a total systemic lack of Vitamin B12 completely impairs the highly massive Cobalamin structures, violently interrupting vital neurological pathways and causing rapidly destructive pernicious anemia and severe, completely irreversible neuropathy.
Finally, a profound dietary deficiency of Vitamin C completely removes Ascorbic Acid from the human body, directly halting critical non-specific antioxidant cellular protection and specific essential hydroxylation reactions. Without this highly crucial, wildly important water-soluble reducing agent, vital structural collagen heavily fails to cross-link properly. This massive structural failure directly causes rapid tissue breakdown, massive spontaneous bleeding, and the classic severe symptoms of Scurvy. Understanding these highly direct clinical correlations fundamentally proves to students that coenzymes are absolutely not just theoretical academic concepts, but absolutely vital, literal molecules of life.
Please read our Content Disclaimer Statement.
Check out our social media channels: