37. Glycolysis: Biochemical Pathway and Reactions
Ever wonder how a sprinter bursts off the starting line or how the brain powers through a late-night study session? It all comes down to how cellular machinery extracts energy from food. This comprehensive guide walks through the molecular mechanics of cellular respiration, exploring the distinct reactions, enzymes, and clinical relevance of this primary energy-generating pathway. By dissecting these detailed visual guides, students will master the intricate biochemical steps that sustain human life at the microscopic level.
Slide 1: The Foundations of Glycolysis and Molecular Correlates
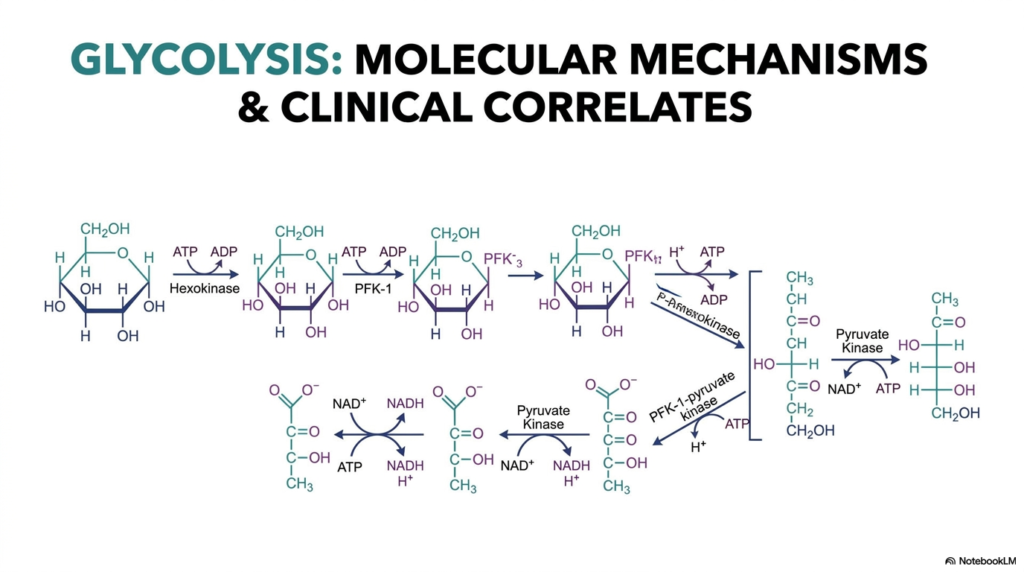
The study of Glycolysis serves as the universal foundation for understanding cellular bioenergetics across almost all living organisms. This initial slide provides a sweeping visual overview of the metabolic pathway, charting the biochemical journey of a single glucose molecule as it undergoes a series of enzymatic transformations. By looking at the complete molecular landscape, students can appreciate the elegant complexity of this process before diving into the deeply specific individual steps. The pathway is not merely a random sequence of events; it is a highly conserved mechanism designed to extract usable energy efficiently.
At its core, Glycolysis is a ten-step enzymatic pathway that occurs entirely in the cell cytosol. The structures shown on this opening slide illustrate the progressive structural breakdown of a six-carbon hexose sugar into two distinct three-carbon molecules of pyruvate. Notice how the molecular intermediates become actively phosphorylated, precisely isomerized, and ultimately cleaved by cellular machinery. These molecular mechanisms are carefully orchestrated by specific enzymes that drastically lower the activation energy of each individual reaction.
Beyond basic metabolism, the clinical correlates of Glycolysis are profoundly important for future physicians and medical researchers. When this metabolic pathway functions normally, it provides a steady, reliable stream of cellular ATP and essential molecular intermediates. However, when these precise molecular mechanisms are violently disrupted by genetic mutations, invasive toxins, or malignant cancerous transformations, the resulting physiological consequences are incredibly severe.
To truly master the nuances of Glycolysis, one must strictly recognize that this pathway fundamentally operates independently of molecular oxygen, distinguishing it from subsequent mitochondrial processes. This ancient, essential evolutionary adaptation allows cellular machinery to survive in deeply hypoxic environments, ensuring continuous cellular function when aerobic respiration is heavily compromised. As we progress through this clinical molecular atlas, keep this overarching structural map firmly in mind, as it forms the ultimate blueprint for cellular survival.
Slide 2: The Net Equation and Phases of Glycolysis
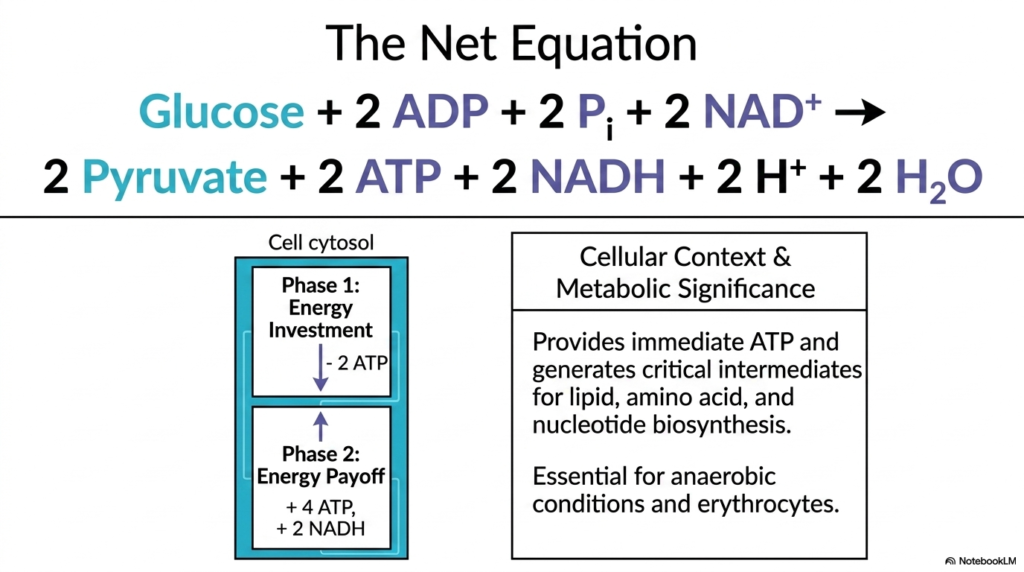
The net equation of Glycolysis reveals the true biochemical accounting of cellular respiration’s first major metabolic phase. The displayed equation perfectly summarizes the transformation of a single glucose molecule into two pyruvate molecules, with a net yield of two ATP and two NADH molecules. Understanding this energetic balance sheet is paramount for medical students, as it clearly demonstrates how much immediate energy is extracted from a carbohydrate without oxygen.
Structurally, Glycolysis is elegantly and distinctly divided into two opposing biochemical phases: the energy investment phase and the energy payoff phase. During the first half of the pathway, the cell must actually spend two valuable ATP molecules to energetically prime the glucose molecule, creating unstable, high-energy intermediates. This initial biochemical investment is entirely necessary to destabilize the hexose ring and structurally prepare it for a clean cleavage. Following this, the payoff phase rapidly generates four ATP molecules, resulting in a net positive yield.
The overarching metabolic significance of Glycolysis extends far beyond just simple, immediate ATP generation. The molecular intermediates seamlessly produced along this catabolic pathway serve as critical, flexible building blocks for other vital cellular processes, including lipid, amino acid, and nucleotide biosynthesis. In tissues that lack mitochondria, such as mature circulating erythrocytes, this cytosolic pathway is the sole source of metabolic energy.
Furthermore, under highly anaerobic conditions, such as intense, sudden muscular exertion, Glycolysis becomes the predominant energy source for the human body. When systemic oxygen supply violently fails to meet cellular metabolic demand, the tissue relies entirely on this pathway to rapidly and continuously generate ATP. This metabolic flexibility highlights the evolutionary brilliance of the process, strictly ensuring that tissues can seamlessly adapt to fluctuating oxygen levels.
Slide 3: Trapping Glucose Inside the Cell During Glycolysis
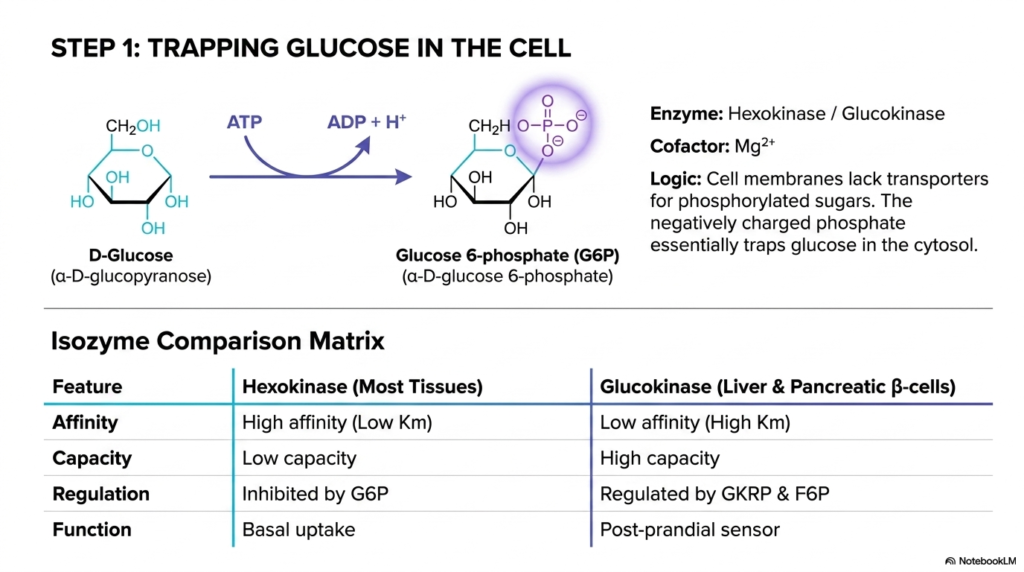
The first crucial biochemical step of Glycolysis involves securely trapping the energy-rich glucose molecule within the cellular environment. Upon rapidly entering the cytosol, glucose is immediately phosphorylated into glucose 6-phosphate (G6P) by the enzyme hexokinase. This highly favorable reaction requires the direct investment of one ATP molecule and relies heavily on a magnesium cofactor to chemically stabilize the intense negative charges of the phosphate groups. By attaching this bulky, negatively charged group, the cell ensures the molecule cannot escape.
This initial trapping mechanism is a masterpiece of biochemical logic that is deeply essential to Glycolysis. Cell membranes naturally and strictly lack any specialized transport proteins capable of successfully moving phosphorylated sugars across the dense lipid bilayer. Therefore, once glucose is converted to G6P, it is irreversibly committed to remain in the cytosol, thereby lowering the intracellular concentration of free glucose. This maintained concentration gradient allows the cell to continuously draw in more glucose.
Students must strongly recognize the critical tissue-specific differences in the enzymatic isozymes that initiate Glycolysis, specifically hexokinase versus glucokinase. Hexokinase is widely found in most peripheral tissues and has a remarkably high affinity for glucose, enabling it to function at maximum enzymatic capacity even when systemic blood sugar levels are extremely low. It is primarily responsible for strict basal glucose uptake and is strongly inhibited by its own chemical product.
In sharp contrast, glucokinase operates predominantly within the liver and pancreatic beta-cells, functioning as a vital, highly sensitive post-prandial glucose sensor. Remarkably, it has a much lower enzymatic affinity for glucose, activating only when blood sugar is markedly high, such as immediately after a heavy meal. Glucokinase is uniquely regulated by specific regulatory proteins and strictly lacks product inhibition, allowing the liver to continuously clear massive amounts of dietary glucose from the portal vein.
Slide 4: Isomerization and the Committed Step of Glycolysis
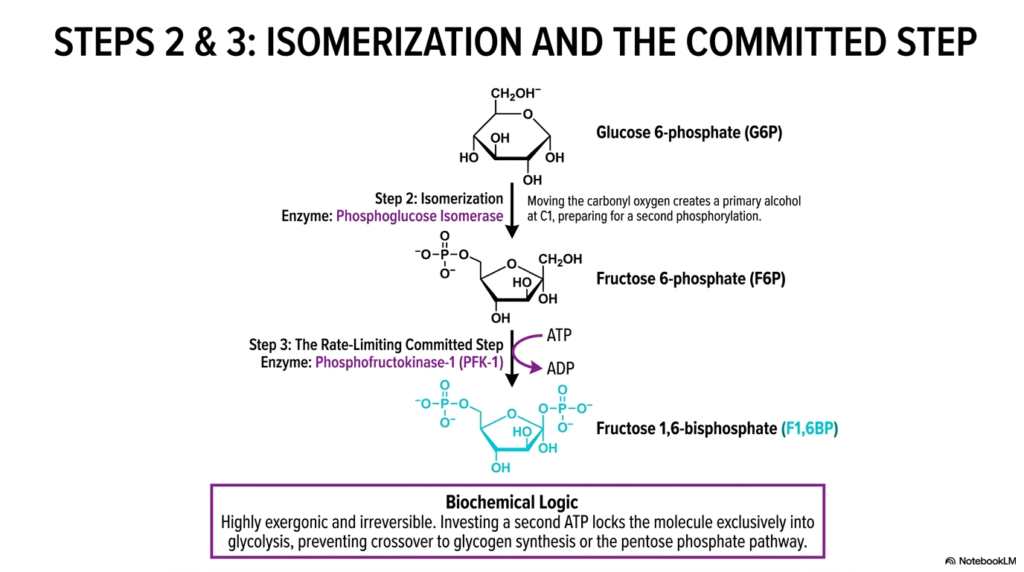
As the metabolic pathway of Glycolysis progresses, the second sequential step involves the delicate structural isomerization of glucose 6-phosphate into fructose 6-phosphate. Flawlessly catalyzed by the enzyme phosphoglucose isomerase, this internal molecular rearrangement precisely shifts the carbonyl oxygen, effectively converting an aldose sugar directly into a ketose sugar. This seemingly subtle biochemical transformation is physically profound, as it successfully exposes a primary alcohol at the first carbon position, perfectly priming the molecule for the next reaction.
Following this isomerization, the third step represents the absolute most crucial regulatory checkpoint in all of Glycolysis. The enzyme phosphofructokinase-1 (PFK-1) directly uses a second ATP molecule to phosphorylate fructose 6-phosphate, immediately generating fructose 1,6-bisphosphate. This specific kinase reaction is highly exergonic and physiologically irreversible, explicitly meaning that once this energetic threshold is crossed, the cellular machinery is entirely committed to pushing the molecule all the way down the pathway.
The biochemical logic that drives this committed step in Glycolysis is incredibly elegant and undeniably vital to overall cellular efficiency. Prior to this highly irreversible phosphorylation, the intermediate glucose 6-phosphate could easily and safely have been diverted into alternative metabolic routes, such as glycogen synthesis for energy storage or the pentose phosphate pathway for nucleic acid production. By heavily investing a second ATP molecule right here, the cell strictly locks the substrate into a catabolic fate.
Understanding this strict biochemical commitment ultimately prevents the deadly, wasteful crossover of precious metabolic resources. For medical students, thoroughly grasping the sheer irreversibility of this specific reaction is the ultimate key to understanding how cells safely prioritize immediate energy generation over long-term storage. By tightly regulating PFK-1, the cell acts as a flawless metabolic traffic cop, authorizing the rapid continuation of the pathway only when intracellular energy demands are critically high.
Slide 5: Lysis and Triose Equilibration in Glycolysis
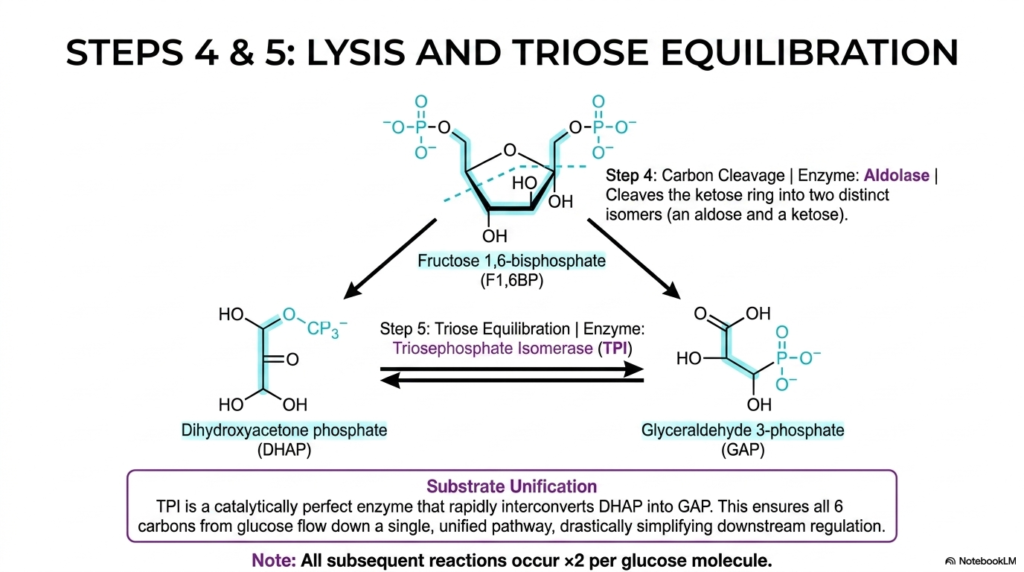
The fourth sequential step of Glycolysis introduces a literal splitting of the molecular framework, known biochemically as lysis. The heavily phosphorylated six-carbon intermediate, fructose 1,6-bisphosphate, is violently cleaved right down the middle by the precise enzyme aldolase. This distinct structural-molecular division produces two distinct three-carbon isomers: an aldose, glyceraldehyde 3-phosphate (GAP), and a ketose, dihydroxyacetone phosphate (DHAP). This cleavage distinctly marks the transition into the energetic processing phase.
However, the downstream catalytic enzymes of Glycolysis are highly specific and can metabolize only one of these two structural isomers. While GAP can directly and seamlessly continue down the energetic pathway, the newly formed DHAP is temporarily biochemically useless in its current structural conformation. To successfully resolve this biochemical dilemma, the pathway relies intensely on the extraordinary catalytic efficiency of a highly specialized enzyme known as triosephosphate isomerase (TPI).
Triosephosphate isomerase is widely celebrated in modern biochemistry as a truly “catalytically perfect” enzyme, flawlessly operating at the absolute maximum speed allowed by the strict laws of physical diffusion. It rapidly and seamlessly interconverts the useless DHAP directly into the highly usable GAP. This rapid molecular equilibration is a deeply critical component of Glycolysis because it strictly enforces a brilliant, unifying metabolic concept securely known as substrate unification.
Thanks to this rapid substrate unification, all six original carbons from the starting glucose molecule now flow continuously down a single, unified biochemical pathway. Students must remember that, because of this specific isomerization event, every subsequent reaction in Glycolysis from this point forward occurs exactly twice per initial glucose molecule. This doubling effect is the mathematical secret to how the cellular machinery ultimately achieves a net positive yield of ATP.
Slide 6: Oxidation, NADH Generation, and the Bottleneck of Glycolysis
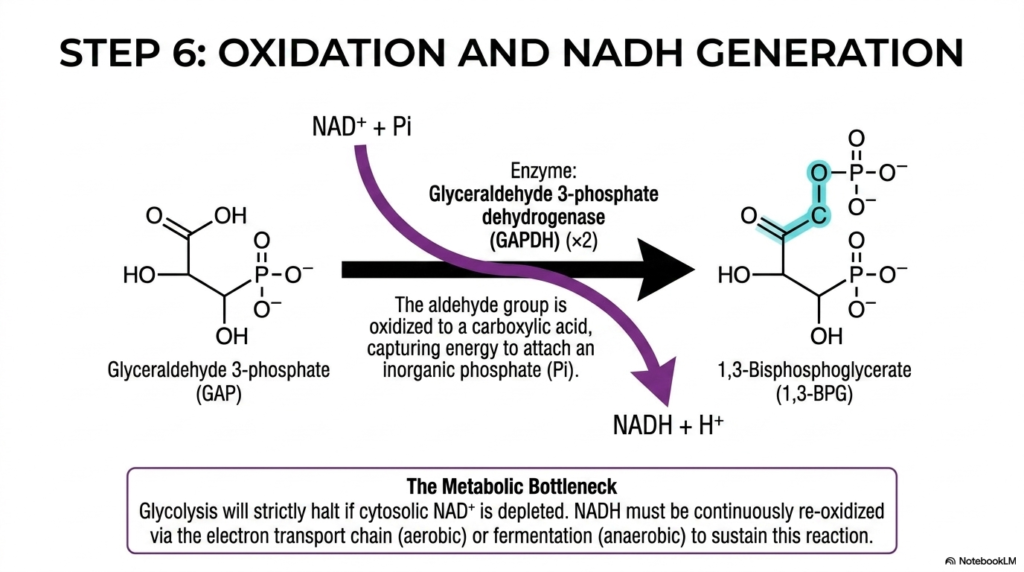
Step six powerfully marks the exact beginning of the crucial energy payoff phase within Glycolysis, properly introducing the first oxidation-reduction reaction of the entire pathway. The specific enzyme glyceraldehyde 3-phosphate dehydrogenase (GAPDH) efficiently catalyzes the oxidation of the aldehyde group on GAPDH directly to a carboxylic acid. This favorable chemical oxidation firmly releases a significant amount of molecular energy, which the enzyme cleverly captures to physically attach free, inorganic phosphate to the carbon skeleton.
This specific reaction in Glycolysis is biochemically monumental because it is the first time the pathway actively harvests energy rather than spending it. Concurrently, the free electrons violently lost during the rapid oxidation of GAP are transferred directly to the waiting coenzyme NAD+, thereby securely reducing it to NADH along with a free hydrogen ion. The newly formed 1,3-bisphosphoglycerate now beautifully contains a highly unstable acyl-phosphate bond, essentially storing the captured energy.
However, this vital oxidative step introduces a significant cellular vulnerability, clinically known as the metabolic bottleneck of Glycolysis. The cytosolic supply of the absolutely essential electron acceptor, NAD+, is highly finite and strictly limited by the cell. If the cellular machinery rapidly consumes all available NAD+ by heavily reducing it to NADH, the entire metabolic cascade will strictly and violently halt at this exact step, totally depriving the cell of further energy.
To sustain continuous energy production, the cell must rapidly re-oxidize the generated NADH back to NAD+. In environments with abundant oxygen, this necessary re-oxidation is gracefully handled by the deeply complex mitochondrial electron transport chain. Conversely, under severe anaerobic conditions, the cell must rely entirely on emergency fermentation. Medical students must deeply understand this delicate electron balance, as it fundamentally dictates whether a cell survives a hypoxic crisis.
Slide 7: The Break-Even Point of Glycolysis
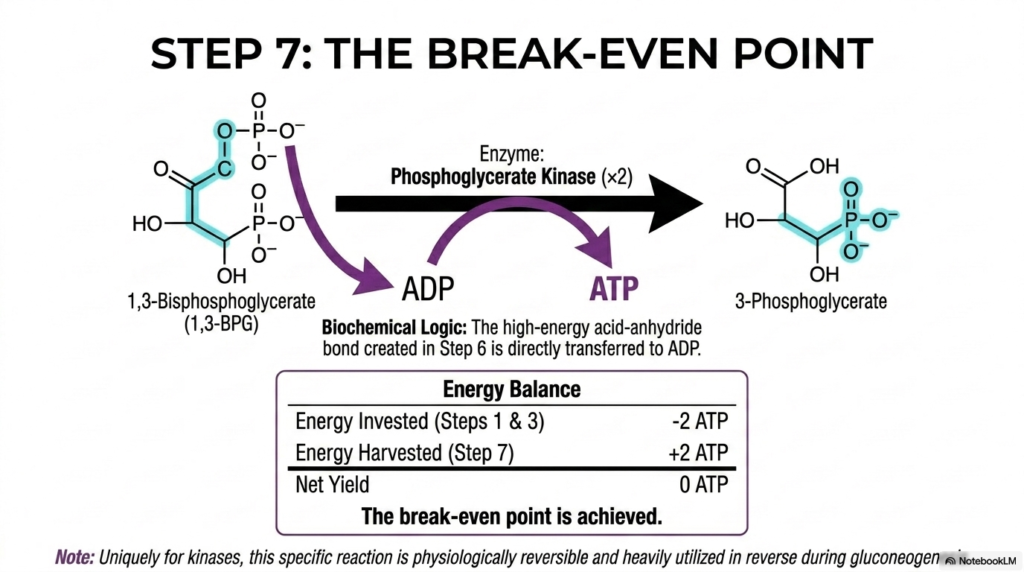
Step seven is a monumental and celebrated milestone in the biochemical journey of Glycolysis, affectionately referred to as the metabolic break-even point. Here, the vital enzyme phosphoglycerate kinase flawlessly catalyzes the strict transfer of the high-energy phosphate group from 1,3-bisphosphoglycerate directly onto an awaiting ADP molecule. This direct, enzyme-mediated transfer beautifully generates a fresh, usable molecule of ATP and seamlessly yields the product 3-phosphoglycerate. Because this phase occurs twice, it produces 2 ATP molecules.
This precise enzymatic mechanism is a totally perfect example of substrate-level phosphorylation, a deeply core physiological concept in Glycolysis. The highly unstable, incredibly energy-rich acid-anhydride bond that was ingeniously formed in the previous biochemical step is now violently cleaved to drive the direct synthesis of ATP. When we carefully tally the strict cellular energy balance sheet at this precise moment, the two ATPs invested earlier have finally been recouped, yielding a net of exactly zero.
An incredibly intriguing biochemical quirk of this highly specific enzyme within Glycolysis is its distinct thermodynamic reversibility. Unlike most kinases in cellular metabolic pathways that drive biochemical reactions in a single, strictly irreversible direction, phosphoglycerate kinase operates very close to true physiological equilibrium. This reversible nature is practically vital, as this exact same enzyme is heavily utilized in reverse during the anabolic process of hepatic gluconeogenesis.
Understanding the strict thermodynamics of the metabolic break-even point is absolutely essential for strongly grasping the overall energetic efficiency of cellular respiration. From here on out, any and all additional energetic units cleanly extracted from the remaining carbon skeleton represent pure, unadulterated cellular profit. This step firmly shifts the biochemical focus of Glycolysis away from energy debt recovery and towards maximizing the final, explosive metabolic output.
Slide 8: Molecular Rearrangement and Dehydration in Glycolysis
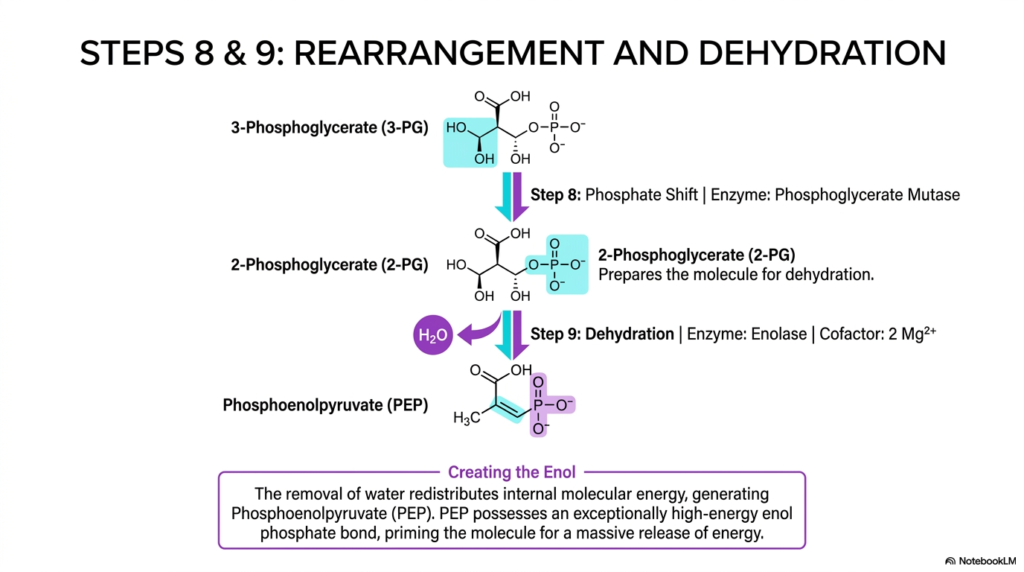
As the pathway of Glycolysis rapidly nears its dramatic conclusion, the remaining carbon skeleton must be structurally rearranged to flawlessly prime it for a massive, terminal energetic release. Step eight specifically utilizes the delicate enzyme phosphoglycerate mutase to execute a precise intramolecular phosphate shift. The phosphate group is physically securely relocated from the third carbon of 3-phosphoglycerate directly to the second carbon. This subtle molecular repositioning structurally prepares the molecule for upcoming dehydration.
Following the shift, step nine forcefully introduces the crucial enzyme enolase, which biochemically drives a powerful dehydration reaction that cleanly removes a single water molecule directly from 2-phosphoglycerate. This violent removal of water fundamentally and totally redistributes the internal molecular energy completely across the physical carbon framework. The stunning structural consequence of this specific dehydration in Glycolysis is the formation of phosphoenolpyruvate (PEP), which possesses an exceptionally high-energy enol phosphate bond.
The sheer biochemical logic behind deliberately creating this specific enol phosphate bond is the true, undeniable secret weapon of Glycolysis. Before the powerful dehydration occurred, the phosphate group was securely attached via a relatively low-energy, stable ester linkage. By violently stripping away the water molecule, the cellular machinery effectively forces the carbon skeleton into a tightly bound, unstable enol configuration that desperately and aggressively seeks to rearrange itself into a far more stable structural state.
For medical students, a deep understanding of the precise biochemical generation of PEP is undeniably critical to understanding how cells maximize their metabolic efficiency. The specific enolase enzyme beautifully acts much like aggressively pulling back the tight string of a biochemical bow, loading the molecular substrate with immense, trapped potential energy. The molecule within Glycolysis is now thoroughly primed, deeply unstable, and perfectly ready to explosively transfer its energetic phosphate group to ADP.
Slide 9: The Final Energy Payoff in Glycolysis
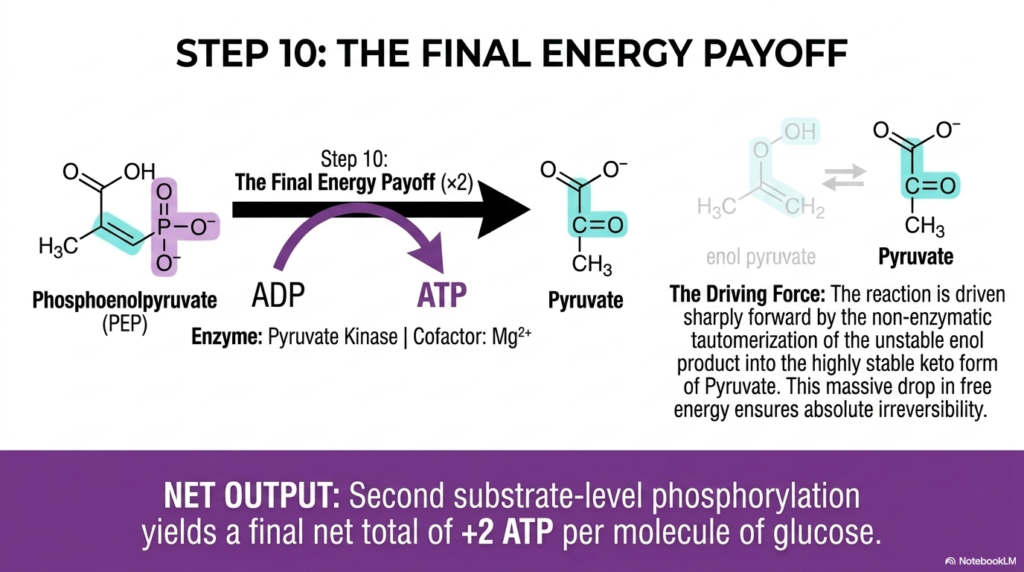
Step ten beautifully represents the triumphant, explosive finale of Glycolysis, delivering the ultimate energy payoff for the entire cell. The critical enzyme pyruvate kinase flawlessly catalyzes the strict, final transfer of the highly energetic phosphate from phosphoenolpyruvate (PEP) directly to a waiting ADP molecule. This reaction beautifully generates fresh ATP alongside the terminal three-carbon product, pyruvate. This step occurs twice per glucose and successfully brings the net ATP output to 2.
The absolute, strict irreversibility of this final crucial step in Glycolysis is violently driven by a massive, truly spontaneous drop in Gibbs free energy. Once the final phosphate group is violently and swiftly removed, the resulting enol pyruvate is extremely and terribly unstable. It tautomerizes rapidly and non-enzymatically into the highly stable keto form of pyruvate. This spontaneous chemical tautomerization acts as a powerful thermodynamic vacuum, sharply driving the reaction firmly forward.
This final, vital generation of ATP successfully represents the second explicit instance of true substrate-level phosphorylation within the entire pathway of Glycolysis. This direct metabolic cellular profit is immediately and fully available to the cell for muscle contraction, ionic active transport, and countless other severe physiological demands. By ensuring that this terminal metabolic step is irreversibly committed, the cellular machinery firmly and flawlessly guarantees that the pathway operates strictly unilaterally.
Deeply understanding this final explosive energy payoff is totally paramount for medical students properly studying cellular bioenergetics. The strict net total of two ATPs generated by Glycolysis might seem entirely modest when compared to total mitochondrial oxidative output, but this precise biochemical reaction is incredibly fast. This sheer explosive speed enables living tissues like skeletal muscle to easily withstand intense, incredibly sudden exertion.
Slide 10: The Thermodynamic Energy Profile of Glycolysis
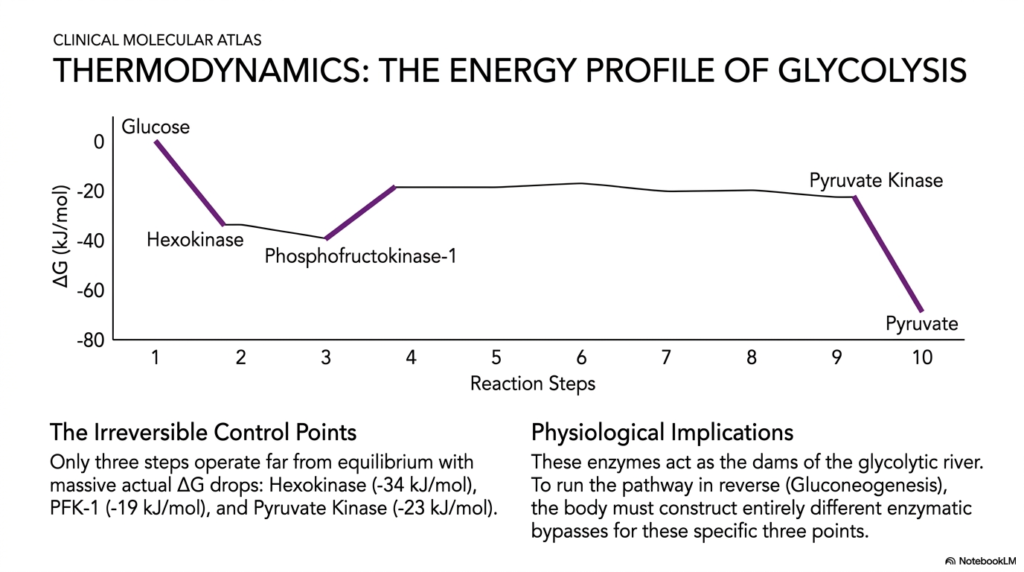
To fully and deeply comprehend the overarching biochemical directionality of Glycolysis, students must carefully analyze the thermodynamic energy profile of the entire pathway. This highly visual clinical atlas accurately charts the distinct changes in Gibbs free energy as raw glucose progressively transforms into pyruvate. The generated graph explicitly reveals that there are exactly three totally irreversible control points that strongly feature massive, sheer negative drops in actual free energy.
These three highly specific, regulated enzymes flawlessly function as the strict metabolic dams of the biochemical river, strictly guiding Glycolysis. Because the total free energy plummets so incredibly drastically at these three distinct molecular locations, the specific catalytic reactions are strictly physiologically irreversible under any normal, healthy cellular conditions. They violently mandate that the entire pathway flow strictly and uncompromisingly in a single, highly catabolic direction without any stagnation.
The deep physiological implications of this specific thermodynamic profile fiercely extend far beyond simple Glycolysis. When a heavily fasting patient’s liver desperately needs to aggressively run this metabolic pathway in complete reverse to violently synthesize new glucose for the starving brain—a complex process properly called gluconeogenesis—it faces a monumental challenge. The human body absolutely cannot push the biochemical river directly back up these three severely steep, physically sheer thermodynamic cliffs.
To properly overcome this, the liver must painstakingly construct distinct enzymatic bypasses to successfully navigate around these three irreversible cellular roadblocks. For medical students, thoroughly memorizing this thermodynamic energy profile is a foundational key to mastering how the human body seamlessly switches between glucose consumption and active glucose production. The strict thermodynamics beautifully dictate this complex regulatory architecture governing Glycolysis and bodily metabolism.
Slide 11: Allosteric Regulation as the Master Controller of Glycolysis
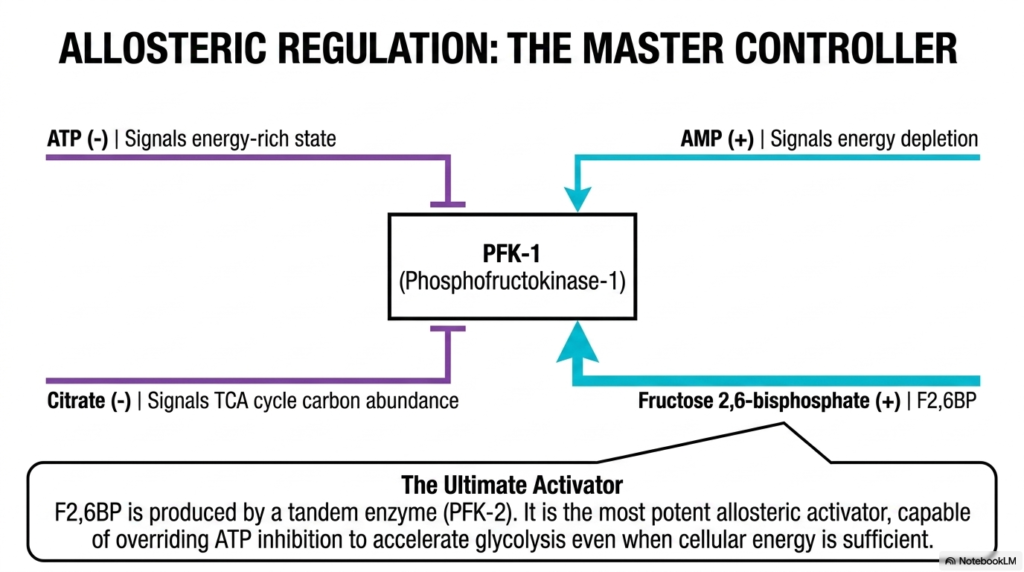
Because Glycolysis is a central hub for rapid energy production, its rate must be tightly and precisely regulated by the cellular environment. The absolute master metabolic controller of the entire pathway is the strict allosteric regulation of the crucial enzyme phosphofructokinase-1 (PFK-1). This specific enzyme carefully integrates multiple competing metabolic signals to precisely match the rapid rate of glucose breakdown to the cell’s immediate, pressing energetic demands.
The highly potent allosteric inhibitors of PFK-1 within Glycolysis brilliantly and rapidly signal a distinct physiological state of sheer energetic abundance. When intracellular ATP levels are excessively high, ATP binds to a distinct regulatory site on the enzyme, thereby aggressively halting its catalytic activity. Similarly, extremely high cytosolic citrate levels act as a powerful negative feedback signal, clearly indicating that the downstream mitochondria are already overflowing with carbon.
Conversely, powerful positive allosteric activators brilliantly signal deep, highly dangerous cellular energy depletion, violently forcing Glycolysis directly into high gear. A sudden buildup of cellular AMP properly indicates that vital ATP stores are severely depleted, thereby strongly binding to PFK-1 to stimulate its activity. However, the most potent metabolic activator of the entire pathway is a highly specialized regulatory molecule, specifically fructose 2,6-bisphosphate, synthesized by a distinct tandem enzyme.
This highly specific regulatory molecule, fructose 2,6-bisphosphate, is incredibly capable of profoundly and aggressively accelerating the pathway, easily overriding even heavily high levels of ATP inhibition to strictly ensure continuous glucose processing. For medical students, a deep understanding of this complex allosteric web governing Glycolysis is essential. It flawlessly reveals how totally individual cells autonomously gauge their own energetic health and seamlessly make microscopic decisions to either safely hoard or violently break down energetic resources.
Slide 12: Hormonal Integration of Glycolysis in the Liver
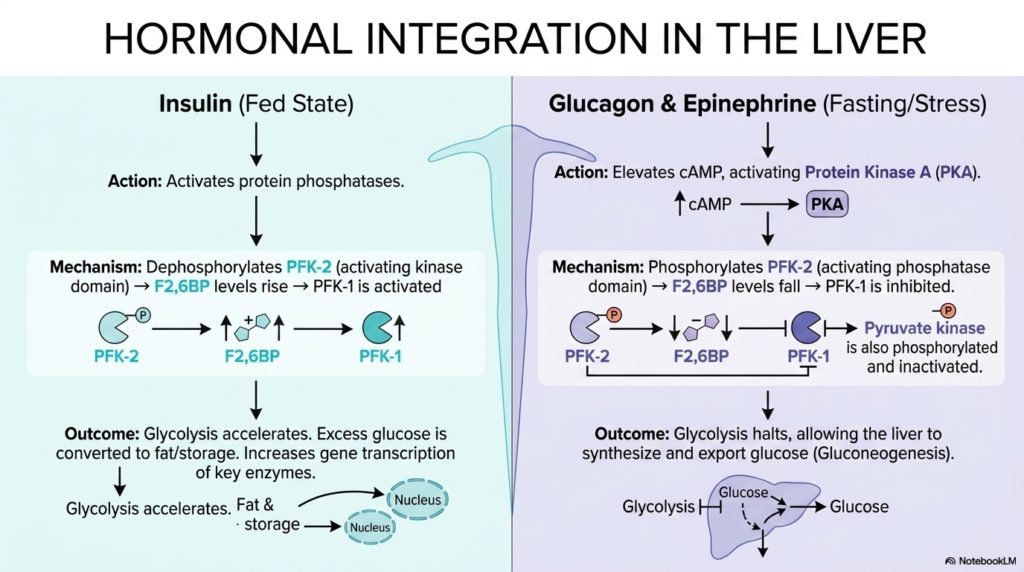
While individual cells successfully and autonomously regulate their own metabolic energy needs, the human liver inherently requires extensive systemic hormonal integration to carefully coordinate glycolysis throughout the entire body. During the highly abundant fed state, the pancreas rapidly releases the hormone insulin. Insulin directly binds to active hepatic receptors and precisely activates intracellular protein phosphatases, which strip bulky phosphate groups from their specific target metabolic enzymes.
Specifically, insulin’s direct cellular action beautifully dephosphorylates the complex tandem enzyme PFK-2, strongly activating its potent kinase domain. This perfectly targeted activation directly triggers a massive, explosive surge in the rapid production of fructose 2,6-bisphosphate. As the ultimate allosteric activator, this molecule powerfully and fiercely stimulates PFK-1, rapidly causing hepatic Glycolysis to accelerate. This rapid biochemical breakdown enables the liver to smoothly convert large amounts of glucose into lipid stores.
Conversely, during severe physiological states of extreme fasting, starvation, or intense stress, systemic hormones like glucagon and epinephrine strictly and violently halt liver Glycolysis entirely. These powerful hormones bind to hepatic receptors, markedly elevating cyclic AMP levels, which in turn aggressively activate Protein Kinase A. This specific kinase rapidly sweeps through the entire liver cell, forcefully phosphorylating target enzymes and strictly destroying the necessary allosteric activators, plunging the pathway into immediate metabolic arrest.
This deeply strict hormonal biochemical switch is an absolute, stunning masterpiece of complex human physiology. By completely shutting down Glycolysis strictly in the liver during a massive fast, the hormone glucagon safely ensures that the vital liver does not selfishly consume the precious glucose it is desperately trying to synthesize for the brain. Medical students must thoroughly master this highly reciprocal hormonal regulation, as it deeply forms the crucial pharmacological basis for understanding systemic diabetes.
Slide 13: The Diverse Fates of Pyruvate After Glycolysis
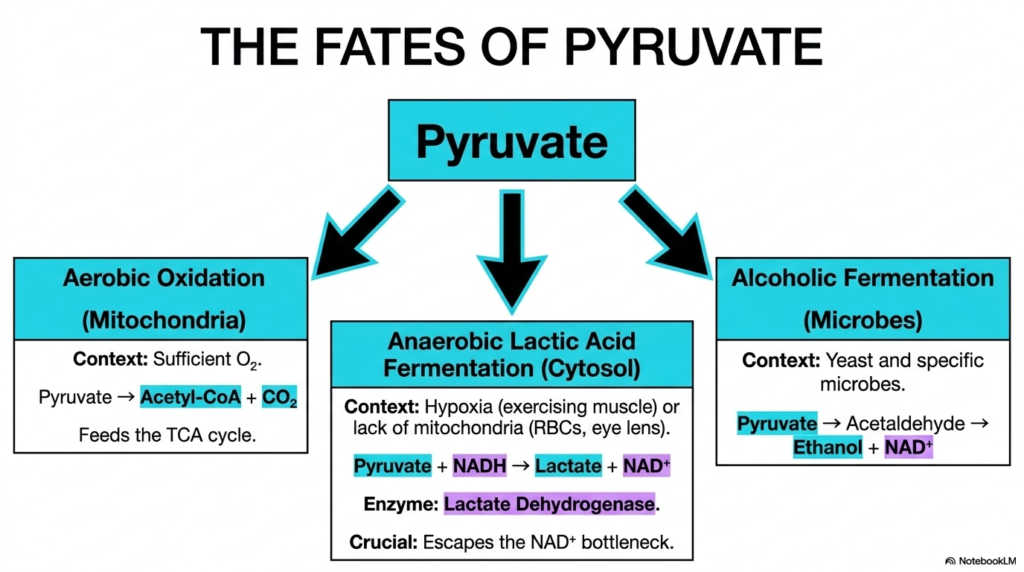
The dramatic biochemical conclusion of Glycolysis does not mark the absolute end of the overall metabolic journey, but rather a vital, critical crossroads that decisively dictates the ultimate fate of the newly formed pyruvate. The highly specific biochemical pathway the generated pyruvate ultimately takes depends entirely on the organism, the tissue type, and the immediate availability of vital molecular oxygen in the surrounding cellular environment.
In the presence of sufficient oxygen and functional mitochondria, pyruvate undergoes strictly aerobic oxidation. It is rapidly transported deeply into the internal mitochondrial matrix, where it is flawlessly converted into Acetyl-CoA. This highly specific biochemical fate following Glycolysis is profoundly efficient, as the produced Acetyl-CoA directly and seamlessly feeds into the energetic TCA cycle. For vital organs like the brain, this continuation is absolutely required for baseline survival.
However, during severe, highly dangerous cellular hypoxia, such as in heavily exercising skeletal muscle, the tissue absolutely cannot rely on the absence of oxygen. To seamlessly survive, the tissue rapidly switches strictly to highly anaerobic lactic acid fermentation. The enzyme lactate dehydrogenase forcefully reduces pyruvate to lactate, simultaneously rapidly oxidizing NADH back to NAD+. This crucial emergency mechanism safely escapes the dangerous metabolic bottleneck that is native to Glycolysis.
Finally, in highly specific, distinct microbes such as yeast, the terminal pyruvate undergoes alcoholic fermentation. Pyruvate is first completely converted into the toxic acetaldehyde and then gracefully reduced to stable ethanol, again perfectly regenerating vital NAD+ in the process. While human physiological biochemistry does not entirely utilize this exact specific route, medical students must properly study it to fully understand the absolute universal necessity of flawlessly maintaining the cytosolic redox balance during Glycolysis.
Slide 14: Glycolysis Pathologies Involving Defects and Toxins
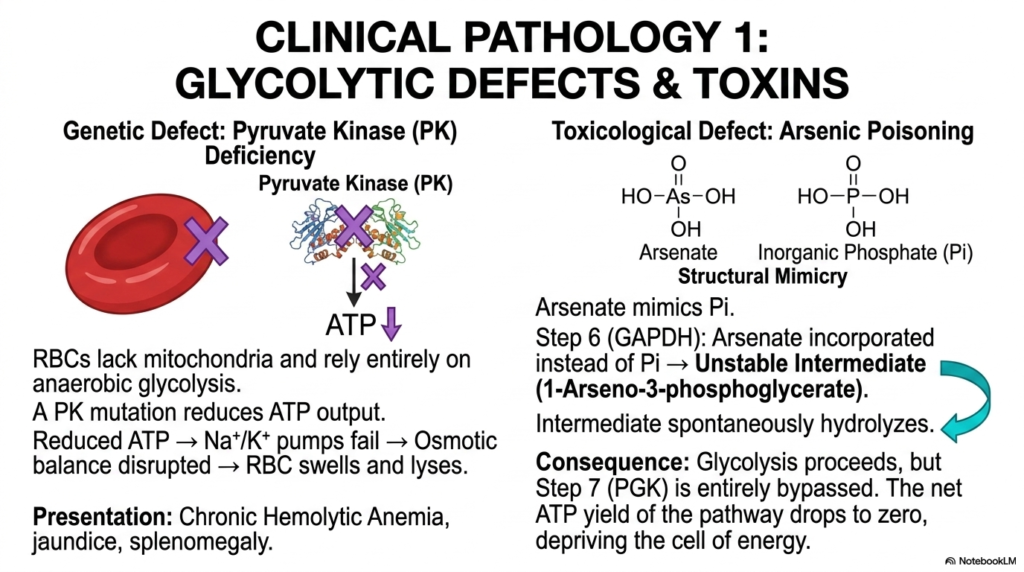
Fiercely transitioning from normal cellular physiology deeply into strict clinical pathology cleanly reveals the entirely catastrophic physiological consequences when the finely tuned molecular machinery of Glycolysis is violently disrupted. One of the most prominent examples of a strict genetic disease is severe Pyruvate Kinase deficiency. Mature red blood cells completely lack internal mitochondria and must rely entirely on the purely anaerobic breakdown of glucose to rapidly generate the vital ATP strictly required to perfectly maintain ionic gradients.
When a highly unfortunate patient inherits a severe Pyruvate Kinase mutation, the terminal energy-yielding step of Glycolysis is severely compromised, drastically reducing total cellular ATP output. Completely deprived of this energy, the vital sodium-potassium pumps on the delicate erythrocyte membrane fail violently. The severely disrupted osmotic balance causes massive cellular swelling, leading the fragile red blood cell to rupture suddenly and violently.
Severe toxicological external interference absolutely also poses a totally systemic, deadly physiological threat to Glycolysis, entirely perfectly demonstrated by severe inorganic arsenic poisoning. Highly toxic arsenate molecules exhibit striking, deadly structural mimicry of normal inorganic phosphate. During step six, the enzyme GAPDH is violently and completely tragically tricked into actively incorporating the poisonous arsenate entirely instead of natural phosphate, strictly creating a highly unstable, highly deadly arseno-intermediate.
The grim consequence of this toxicological molecular mimicry is that, while the physical carbon skeleton blindly and violently continues down the pathway of Glycolysis, the absolutely critical break-even energy generation is entirely and fatally skipped. The net ATP yield of the entire metabolic pathway rapidly plummets to exactly zero. Medical students absolutely must safely recognize how this totally sinister biochemical uncoupling fundamentally deeply forces the vulnerable cell entirely into severe exhaustion.
Slide 15: The Warburg Effect and Cancer Glycolysis
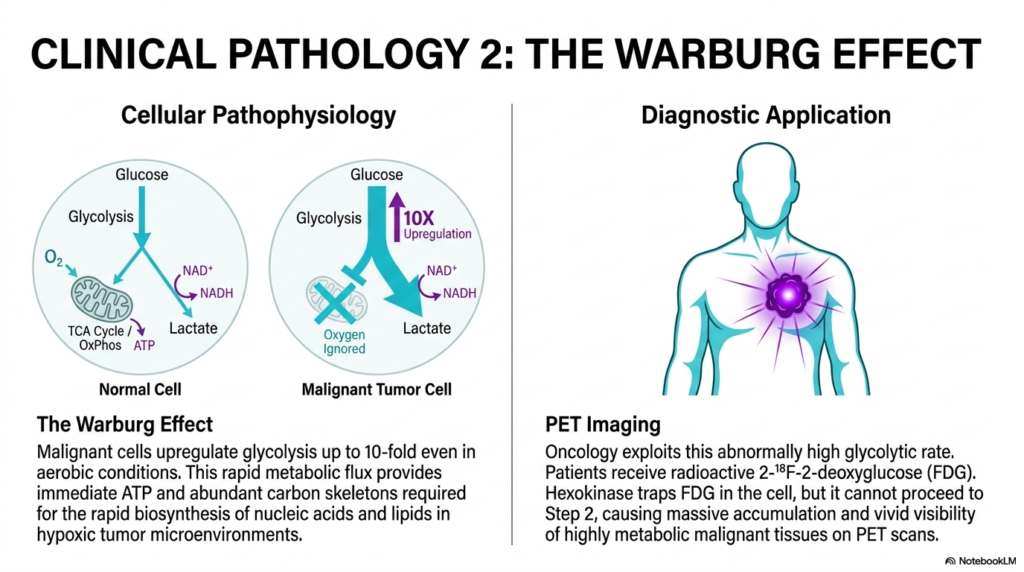
The totally final biochemical slide of this comprehensive clinical atlas strictly explores the deadly Warburg Effect, a highly fascinating cellular pathophysiology that perfectly highlights how aggressive malignant tumors completely violently hijack normal metabolism. In perfectly healthy tissues, Glycolysis typically naturally serves as a minor precursor biochemical pathway. However, deadly cancer cells completely fundamentally alter this metabolic logic, fiercely upregulating the entire exact pathway up to ten-fold, even in highly aerobic environments.
This bizarre, seemingly entirely inefficient total upregulation of Glycolysis by absolutely aggressive malignant tumors is deeply and biologically firmly driven by sheer intense evolutionary survival pressure. While relying heavily on completely anaerobic fermentation undeniably yields far less ATP per molecule of glucose, it acts significantly faster. More importantly, this massive metabolic flux provides the tumor with an incredibly rich, utterly abundant supply of vital cytosolic carbon skeletons for deadly biosynthesis.
Modern clinical oncology brilliantly and totally exploits this highly hyperactive, massive cellular dependence on Glycolysis for highly powerful diagnostic imaging applications, most notably in specialized Positron Emission Tomography scanning. Oncology patients are carefully and strictly injected with a highly specialized, specially formulated, highly radioactive glucose analog, also known as 18F-fluorodeoxyglucose. When this analog rapidly enters the bloodstream, the highly ravenous tumors rapidly utterly violently absorb it.
Once violently trapped inside the highly malignant cancer cell, the initial crucial enzyme deeply governing Glycolysis aggressively and strongly phosphorylates the entirely radioactive analog, firmly trapping it within the enclosed cytosol. However, because of its strictly altered structural molecular structure, the analog cannot safely proceed to the distinct second step. It massively accumulates entirely inside the tumor, powerfully and brilliantly creating a vibrant, visibly glowing clinical hotspot.
Please read our Content Disclaimer Statement.
Check out our social media channels: