41. Gluconeogenesis: The Pathway of De Novo Glucose Synthesis
Imagine the human brain running out of fuel during a long fast; the consequences would be catastrophic. Thankfully, human physiology possesses a metabolic failsafe to prevent this exact scenario. This academic slide deck explores the critical biochemical mechanisms that enable cellular structures to generate their own energy from scratch. By analyzing the complex structural pathways, distinct enzyme regulatory steps, and severe clinical implications of endogenous glucose production, this text provides a comprehensive blueprint of incredible metabolic survival.
Slide 1: Gluconeogenesis: The Metabolic Blueprint of Glucose Biosynthesis
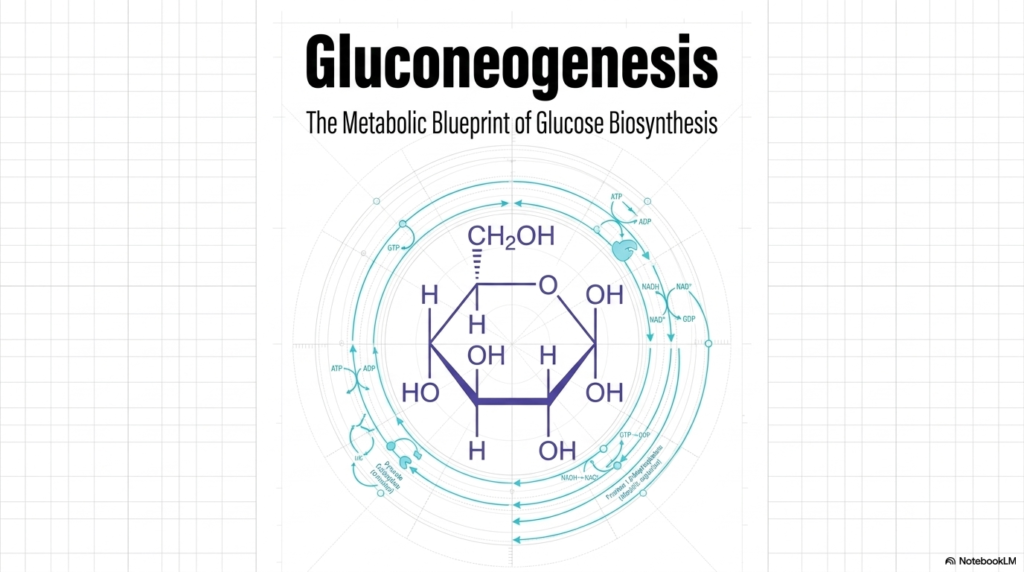
The initial slide introduces the foundational concept of Gluconeogenesis, serving as a comprehensive master blueprint for de novo glucose biosynthesis. At the very center of this metabolic visualization lies the chemical structure of glucose, a critical six-carbon sugar that functions as the primary energy currency for countless cellular operations. Surrounding this central molecule are the intricate, cyclical pathways representing the metabolic steps required to synthesize it. The circular design of the blueprint emphasizes that cellular metabolism is a continuous, interconnected loop rather than a simple linear progression.
When studying Gluconeogenesis, students must recognize that this pathway is not merely a straightforward reversal of glycolysis. It is a highly specialized, independent biological process governed by its own unique thermodynamic rules and distinct regulatory mechanisms. The background blueprint aesthetic highlights the architectural precision required by the cell to build this vital molecule from scratch. Every atom and chemical bond must be meticulously assembled through a series of specific enzymatic reactions, consuming a vast amount of cellular energy in the process.
This introductory visualization prepares medical students to delve into the complex microscopic world of cellular metabolism. By examining the process of Gluconeogenesis from a broad, systemic perspective first, one can better appreciate the complex molecular choreography that follows. The outer rings of the diagram illustrate the continuous flux of nucleotides such as ATP, ADP, and GTP, as well as electron carriers such as NADH. These critical cofactors are the unsung heroes of Gluconeogenesis, providing the necessary thermodynamic driving force.
Ultimately, this slide establishes the central overarching theme of the entire presentation: metabolic resilience and biochemical adaptation. The intricate blueprint of Gluconeogenesis illustrates how human physiology has evolved to safeguard the brain and other obligate tissues against starvation. As the subsequent slides unfold, this structural foundation remains paramount. The detailed enzymatic steps will reveal exactly how the body seamlessly shifts from consuming dietary carbohydrates to relying entirely on this remarkable endogenous synthesis pathway to maintain systemic homeostasis.
Slide 2: The Biological Imperative for Gluconeogenesis and Endogenous Glucose
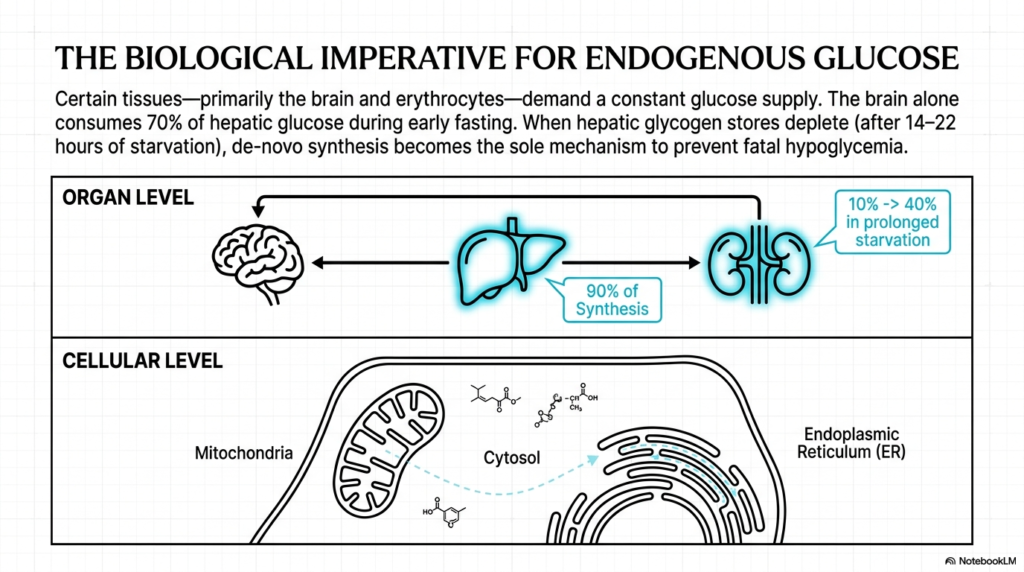
The biological necessity of Gluconeogenesis cannot be overstated when examining human metabolic physiology. Certain tissues within the body—primarily the brain and erythrocytes (red blood cells)—are obligate glucose consumers, meaning they demand a constant, uninterrupted supply of this hexose sugar to function. During the early stages of fasting, the brain alone consumes roughly 70 percent of all hepatic glucose output. As an expert biochemist will note, maintaining this supply is non-negotiable for neurological survival and fundamental cellular operations across these vital organ systems.
A critical transition occurs when the body’s readily accessible carbohydrate reserves begin to be depleted. After approximately 14 to 22 hours of starvation, hepatic glycogen stores are entirely exhausted. At this critical juncture, de novo synthesis via Gluconeogenesis becomes the sole biological mechanism capable of preventing fatal hypoglycemia. The pathway activates dramatically, shifting the body’s entire metabolic machinery from a state of carbohydrate storage and consumption to one of rigorous, energy-intensive endogenous production to sustain the organism.
When examining the architecture at the organ level, the liver emerges as the primary command center. Under normal fasting conditions, hepatocytes are responsible for roughly 90 percent of all glucose synthesis through the Gluconeogenesis pathway. However, the kidneys also play a crucial supplementary role. While they normally handle only 10 percent of synthesis, their contribution can astonishingly increase to 40 percent during prolonged starvation, demonstrating remarkable metabolic plasticity under systemic stress.
Finally, at the cellular level, this pathway is a marvel of spatial organization. The enzymatic reactions of Gluconeogenesis do not occur in a single compartment; rather, they are intricately partitioned across the mitochondria, the cytosol, and the endoplasmic reticulum. This strict compartmentalization prevents futile cycling and enables exquisite regulation, ensuring that intermediates are precisely shuttled between organelles to complete the complex assembly of a brand-new glucose molecule.
Slide 3: Gluconeogenesis and Sourcing the Carbon Backbone
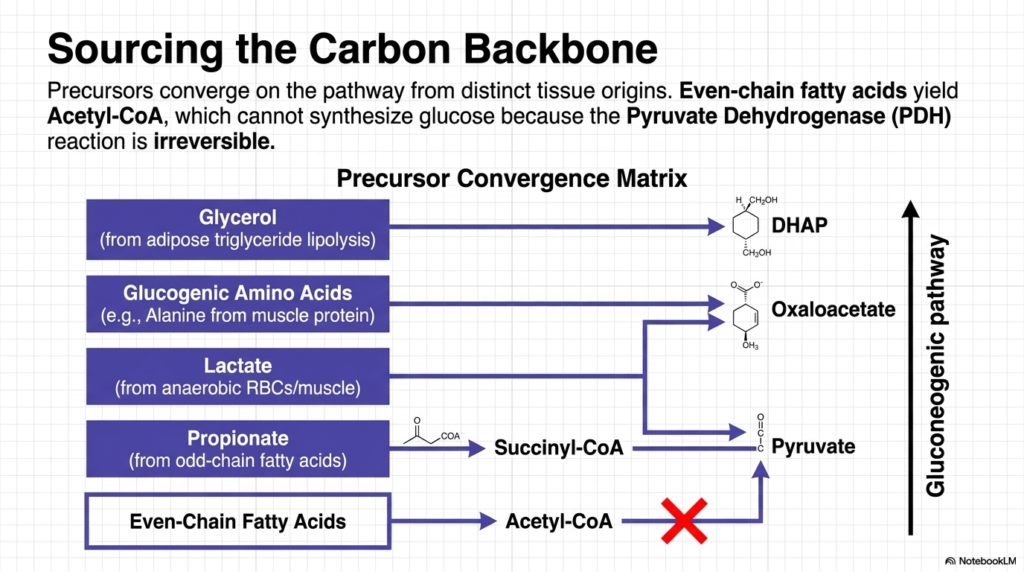
A thorough understanding of Gluconeogenesis requires examining the sourcing of its carbon backbones. The precursors for this pathway originate from distinct tissue sources, highlighting remarkable physiological coordination. The precursor convergence matrix illustrates how glycerol, released from adipose tissue triglyceride lipolysis, directly enters the cascade as Dihydroxyacetone phosphate (DHAP). This demonstrates how fat stores contribute structurally to sugar synthesis. Meanwhile, glucogenic amino acids, primarily alanine, mobilized from skeletal muscle protein breakdown, are funneled into the cycle by forming crucial intermediates such as oxaloacetate and pyruvate.
Another vital substrate for Gluconeogenesis is lactate, which is continuously generated by red blood cells and anaerobically exercising muscle tissue. This lactate travels through the bloodstream and is converted back into pyruvate within the liver. Furthermore, propionate, a byproduct of the oxidation of odd-chain fatty acids, provides another carbon source by entering the cycle as succinyl-CoA. These diverse molecular inputs underscore the pathway’s remarkable versatility, enabling the liver to scavenge a range of metabolic byproducts and convert them into life-sustaining energy.
A common misconception among early students is the role of standard fats in Gluconeogenesis. The diagram explicitly strikes out the pathway from even-chain fatty acids to glucose. When these common lipids undergo beta-oxidation, they yield vast amounts of Acetyl-CoA. However, the Pyruvate Dehydrogenase (PDH) complex catalyzes an irreversible reaction; Acetyl-CoA cannot be converted back into pyruvate. Consequently, the carbon atoms from typical even-chain fats can never serve as a structural backbone for new sugar synthesis.
Therefore, the biological constraints of Gluconeogenesis dictate that while the body can synthesize sugar from proteins and select metabolic waste products, it cannot directly convert the bulk of its fat reserves into glucose. Acetyl-CoA instead acts as a powerful allosteric regulator, providing the thermodynamic energy necessary to drive the pathway, yet it remains structurally distinct from the newly formed hexose molecule. This fundamental limitation explains why muscle wasting occurs during prolonged starvation despite abundant adipose tissue.
Slide 4: Peripheral Recycling to Fuel Gluconeogenesis: The Cori and Cahill Cycles
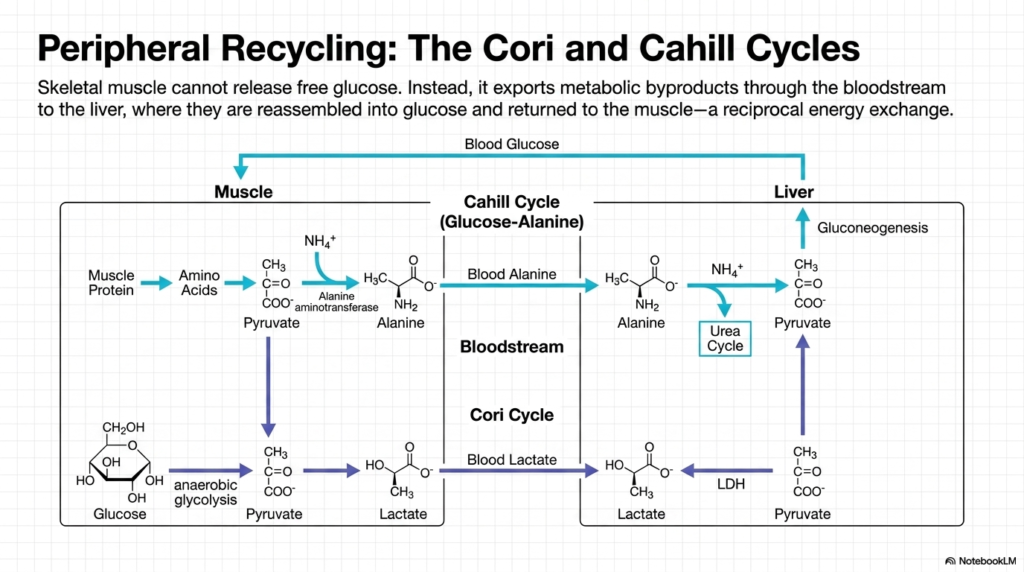
The concept of peripheral recycling is essential when studying the broad systemic impact of Gluconeogenesis. Skeletal muscle poses a unique biochemical challenge: it lacks the specific enzyme required to release free glucose into the circulation. Instead of sharing its energy directly, the muscle exports metabolic byproducts into the bloodstream for delivery to the liver. The liver then reassembles these smaller molecules into complete sugars and returns them to the muscle, establishing a reciprocal energy exchange that sustains the organism during exertion or starvation.
One half of this recycling system is the Cahill Cycle, also known as the glucose-alanine cycle. During states of starvation, muscle protein is aggressively broken down into free amino acids. These amino acids transfer their nitrogen groups to pyruvate, forming alanine. This alanine is then secreted into the bloodstream and travels to the liver. Upon arrival, the liver extracts the carbon skeleton to fuel Gluconeogenesis, while the toxic ammonium byproduct is safely sequestered and disposed of via the specialized Urea Cycle.
The second major recycling mechanism intimately tied to Gluconeogenesis is the Cori Cycle. When skeletal muscle undergoes rigorous anaerobic exercise, it rapidly breaks down its glycogen stores, producing large quantities of lactate as a metabolic end product. This lactate diffuses out of the muscle cells and enters the systemic circulation. Because the muscle cannot further metabolize lactate efficiently under anaerobic conditions, it relies entirely on the liver to clear this acidic byproduct and restore metabolic equilibrium.
Upon reaching the hepatic tissue, lactate is swiftly converted back into pyruvate by the enzyme Lactate Dehydrogenase. This newly formed pyruvate is immediately funneled into the Gluconeogenesis pathway, successfully regenerating a new molecule of glucose. The liver subsequently releases this fresh sugar back into the blood, where it travels directly back to the working skeletal muscle. This elegant inter-organ cooperation ensures a continuous supply of energy while simultaneously preventing the dangerous accumulation of lactic acid in the peripheral tissues.
Slide 5: The Gluconeogenesis Pathway Mirror: Overcoming Thermodynamic Roadblocks
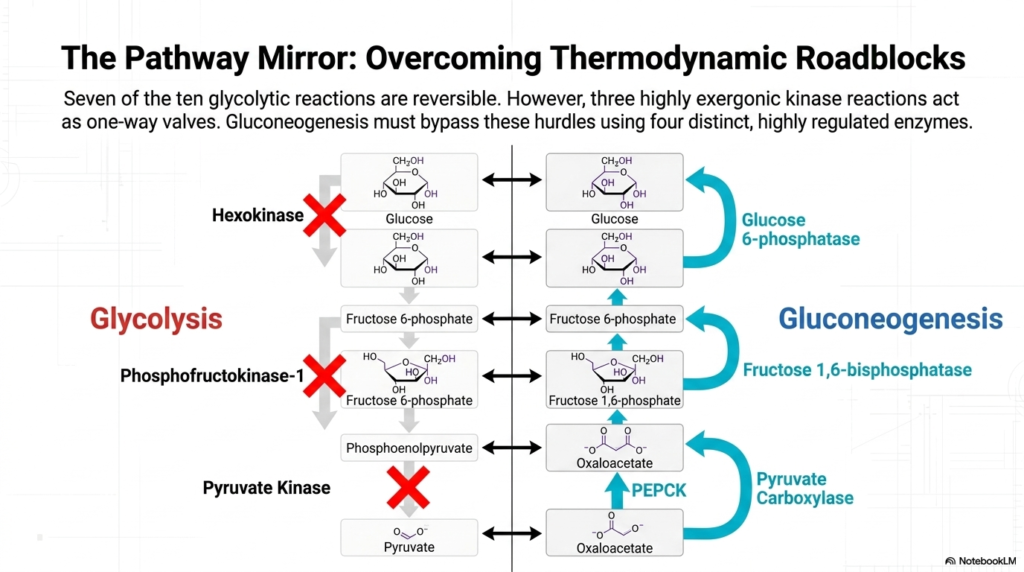
When examining the cell’s metabolic pathways, glycolysis and Gluconeogenesis act as a biochemical mirror, yet they are not identical. The architecture of cellular metabolism reveals that seven out of the ten glycolytic reactions are entirely reversible. These near-equilibrium reactions can readily proceed in either direction depending on the concentrations of substrates and products. However, the cell employs three highly exergonic kinase reactions that act as strictly regulated, irreversible one-way valves, preventing a simple backward flow.
To successfully manufacture glucose, the pathway of Gluconeogenesis must ingeniously bypass these three thermodynamic hurdles. The glycolytic enzymes Hexokinase, Phosphofructokinase-1 (PFK-1), and Pyruvate Kinase catalyze reactions with large negative free-energy changes, making them physically impossible to reverse under normal physiological conditions. To overcome these roadblocks, the body employs a specialized set of four distinct, highly regulated enzymes dedicated exclusively to synthesizing glucose, allowing the cell to navigate around the impenetrable barriers of glycolysis.
The first major obstacle is bypassing Pyruvate Kinase. Gluconeogenesis achieves this by utilizing a two-step maneuver involving the enzymes Pyruvate Carboxylase and Phosphoenolpyruvate Carboxykinase (PEPCK). This elegant detour transforms pyruvate into oxaloacetate, and then subsequently into phosphoenolpyruvate, effectively vaulting over the first major thermodynamic barrier. This specific bypass is the most energy-intensive segment of the entire process, requiring significant ATP and GTP hydrolysis to physically force the molecules up the energetic gradient.
The remaining two hurdles in Gluconeogenesis involve stripping phosphate groups off intermediate molecules. The pathway bypasses PFK-1 by employing Fructose 1,6-bisphosphatase to convert fructose 1,6-bisphosphate back into fructose 6-phosphate. Finally, the barrier imposed by Hexokinase is overcome by Glucose 6-phosphatase, which carries out the final hydrolysis to release free, unphosphorylated glucose. By utilizing these specialized phosphatases, the liver perfectly counteracts the irreversible kinase enzymes of glycolysis, allowing the seamless production of vital energy.
Slide 6: Bypass I of Gluconeogenesis (Phase 1): The Mitochondrial Detour
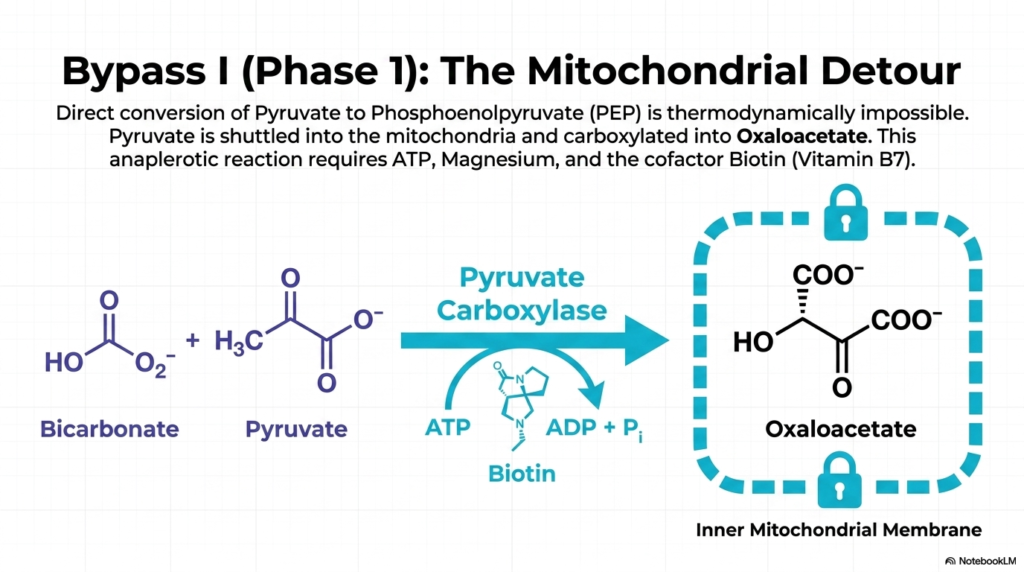
The first crucial phase in overcoming the thermodynamic barriers of Gluconeogenesis is known as Bypass I, specifically focusing on the mitochondrial detour. As previously established, the direct, single-step conversion of pyruvate back into phosphoenolpyruvate (PEP) is a thermodynamic impossibility. To navigate this biochemical roadblock, cellular machinery must first shuttle the cytosolic pyruvate deep into the inner mitochondrial matrix. This spatial compartmentalization is visually represented as a locked vault, indicating that this initial step is tightly guarded and sequestered from the cytosol.
Once securely inside the mitochondria, pyruvate encounters the indispensable enzyme Pyruvate Carboxylase. This enzyme facilitates an anaplerotic reaction, meaning it aggressively replenishes the metabolic intermediates of the mitochondrial cycles. Pyruvate Carboxylase attaches a carbon dioxide molecule—derived from ambient bicarbonate—directly to the three-carbon pyruvate, successfully forging the four-carbon compound known as oxaloacetate. This reaction serves as the foundational building block for the rest of the Gluconeogenesis cascade, setting the structural stage for glucose synthesis.
The biochemistry of this specific step in Gluconeogenesis is highly complex and resource-intensive. The enzymatic action of Pyruvate Carboxylase is strictly dependent on ATP hydrolysis, using the energy from a broken phosphate bond to drive the addition of a new carbon atom. Furthermore, the reaction requires magnesium ions to stabilize the nucleotide and the critical cofactor Biotin, also known as Vitamin B7, which physically carries the activated carbon dioxide molecule to the active site.
Understanding this mitochondrial detour is paramount for medical students, as it highlights the sheer energetic cost of initiating gluconeogenesis. The cellular investment of ATP at this very early stage emphasizes that producing endogenous glucose is a demanding task, reserved only for situations of absolute physiological necessity. The successful formation of oxaloacetate completes the first half of Bypass I, paving the way for the molecule’s eventual return to the cellular cytosol.
Slide 7: Bypass I of Gluconeogenesis (Phase 2): The Cytosolic Escape
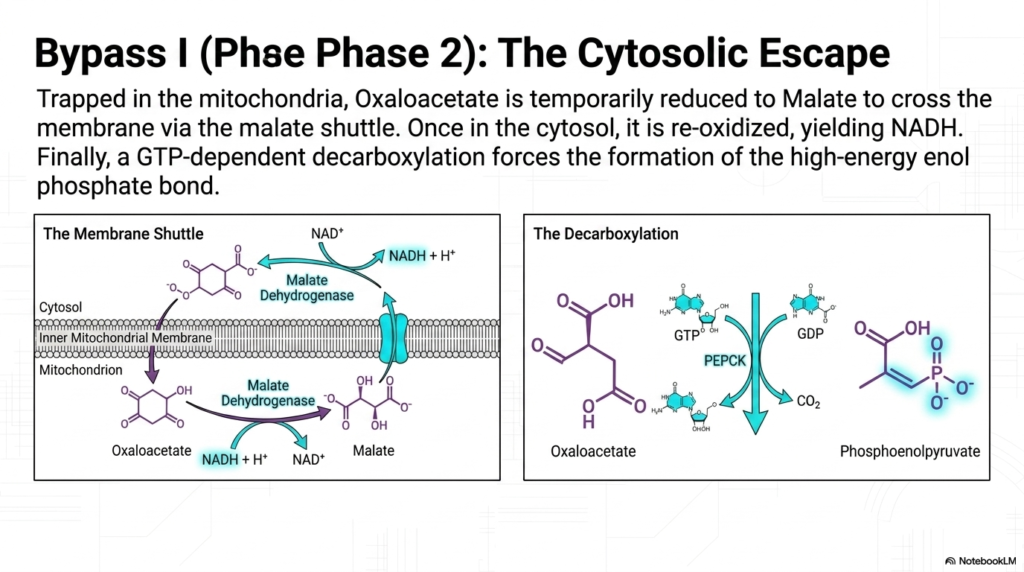
The second phase of Bypass I in Gluconeogenesis presents a unique logistical challenge: the newly synthesized oxaloacetate is completely trapped inside the mitochondria. The inner mitochondrial membrane lacks a dedicated transport protein for oxaloacetate, thereby physically imprisoning the molecule within the organelle’s matrix. To continue the biosynthetic pathway, the cell must execute a clever biochemical workaround known as the cytosolic escape, temporarily altering the structure of the intermediate to smuggle it across the impermeable lipid bilayer.
To achieve this escape, oxaloacetate is rapidly reduced into malate by the mitochondrial isoform of the enzyme Malate Dehydrogenase. Unlike oxaloacetate, malate possesses a specific membrane shuttle that allows it to pass freely into the cytosol. This temporary disguise is absolutely vital for the progression of Gluconeogenesis. Once malate safely arrives in the cytosolic compartment, the cytosolic isoform of Malate Dehydrogenase immediately goes to work, reversing the previous reaction and reoxidizing malate to the functional oxaloacetate.
This shuttle system serves a brilliant dual purpose for the overall machinery of Gluconeogenesis. When malate is re-oxidized in the cytosol, the reaction transfers electrons to NAD+, generating a new molecule of NADH. This is a critical physiological triumph, as the later steps of glucose synthesis heavily depend on a robust supply of cytosolic NADH to provide reducing equivalents. The mitochondria thereby successfully export both the carbon backbone and the necessary reducing power simultaneously.
Once oxaloacetate is successfully relocated, the final step of Bypass I can occur. The enzyme Phosphoenolpyruvate Carboxykinase (PEPCK) acts on the molecule, utilizing energy derived from GTP hydrolysis. PEPCK drives a simultaneous decarboxylation and phosphorylation reaction, stripping away the recently added carbon dioxide while forcefully attaching a high-energy phosphate group. This complex maneuver yields phosphoenolpyruvate, effectively completing the most arduous and energetically demanding step in the Gluconeogenesis pathway.
Slide 8: Bypass II of Gluconeogenesis: The Rate-Limiting Checkpoint
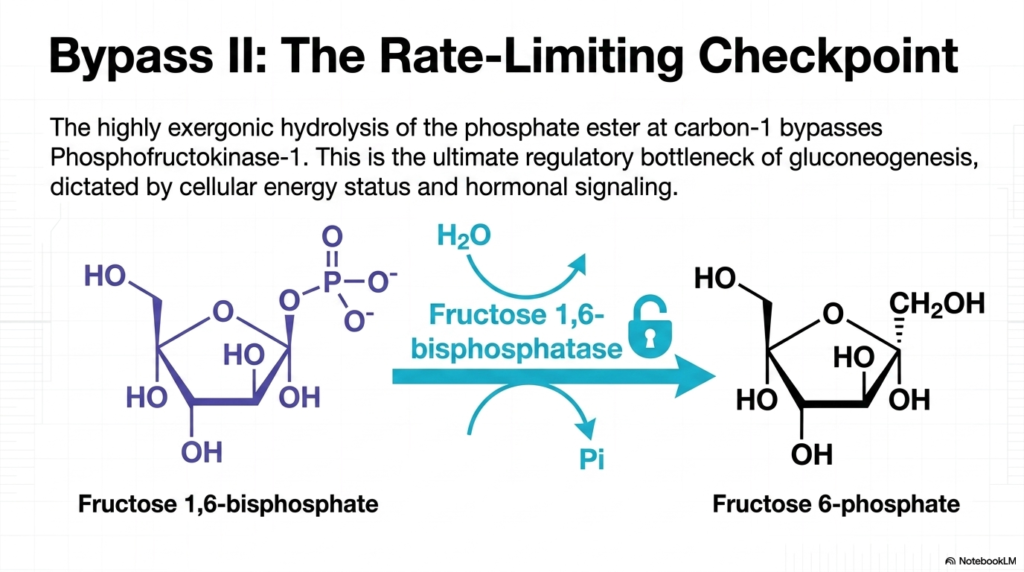
Following the difficult creation of phosphoenolpyruvate, the pathway proceeds through a series of reversible reactions until it encounters its next major roadblock. Bypass II of Gluconeogenesis involves navigating around the highly exergonic glycolytic enzyme Phosphofructokinase-1. The cell accomplishes this by using a specialized enzyme, Fructose 1,6-bisphosphatase. This enzyme catalyzes the hydrolysis of the phosphate ester bond at carbon-1 of Fructose 1,6-bisphosphate, generating Fructose 6-phosphate and releasing inorganic phosphate into the cellular environment.
The biochemical significance of Fructose 1,6-bisphosphatase cannot be overstated; it represents the ultimate rate-limiting checkpoint of the entire Gluconeogenesis pathway. This enzymatic step functions exactly like a master lock or a heavily guarded tollbooth. Because it is irreversible, it dictates the overall pace and flow of endogenous glucose production. If this enzyme is inhibited, the entire assembly line violently halts, preventing the wasteful consumption of cellular energy reserves during inappropriate times.
The strict regulation of this specific Gluconeogenesis checkpoint is primarily governed by the cell’s internal energy status. When energy levels drop dangerously low, signaling molecules rapidly bind to this enzyme, halting its activity to conserve precious ATP. Conversely, when the cell is metabolically rich and has abundant structural precursors, the locks are removed, and Fructose 1,6-bisphosphatase churns out Fructose 6-phosphate at an accelerated rate, pushing the system ever closer to the final product.
Medical professionals must deeply understand this locked gateway, as it serves as the primary target for vast arrays of hormonal signaling. The body uses this specific junction in Gluconeogenesis to translate systemic messages from the pancreas directly into metabolic action. By tightly controlling Fructose 1,6-bisphosphatase, the organism can flawlessly synchronize the liver’s biochemical output with the macroscopic needs of the brain and red blood cells, ensuring metabolic harmony during periods of intense fasting or physiological stress.
Slide 9: Bypass III of Gluconeogenesis: The Endoplasmic Reticulum Release
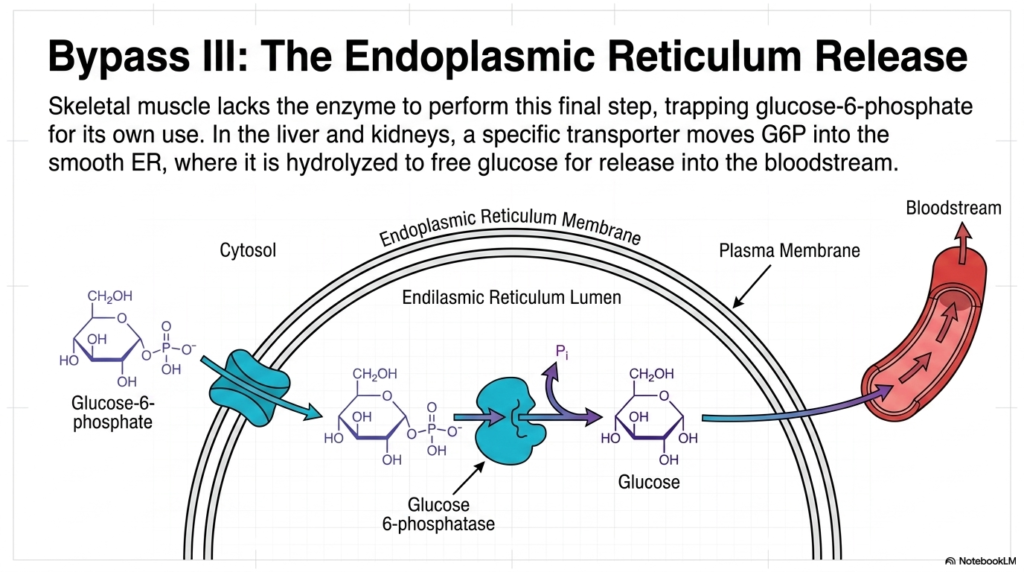
The final, definitive step in the synthesis of endogenous glucose is referred to as Bypass III of Gluconeogenesis. At this late stage in the process, the cell has successfully manufactured Glucose-6-phosphate. However, this phosphorylated molecule carries a strong negative charge, which completely prevents it from crossing the cellular membrane and entering the bloodstream. Interestingly, skeletal muscle tissue lacks the specific enzymatic machinery to remove this phosphate, thereby trapping the newly formed sugar for its own selfish metabolic use.
To nourish the rest of the starving organism, the liver and kidneys use a highly specialized protein structure to complete Gluconeogenesis. A dedicated transport channel actively pulls the cytosolic Glucose-6-phosphate directly into the lumen of the Endoplasmic Reticulum (ER). This strict compartmentalization is a brilliant evolutionary adaptation, ensuring that the final, delicate stage of glucose production is physically separated from the chaotic, enzyme-rich environment of the general cellular cytosol.
Once safely sequestered inside the Endoplasmic Reticulum, the crucial enzyme Glucose 6-phosphatase initiates the final biochemical alteration. It acts as molecular scissors, cleanly hydrolyzing the remaining phosphate bond. This vital reaction strips away the charged inorganic phosphate, successfully yielding a pure, uncharged molecule of free glucose. This specific enzymatic step marks the final step in the rigorous Gluconeogenesis assembly line, transforming a once-useless metabolic intermediate into a universally accepted biological currency.
Now that the glucose is free of its restrictive phosphate group, it can finally escape the cell. The newly formed sugar molecule diffuses out of the Endoplasmic Reticulum, travels across the cellular plasma membrane, and is continuously secreted directly into the bloodstream. By successfully executing this final step of Gluconeogenesis, the liver fulfills its primary biological mandate: rescuing the brain and erythrocytes from impending energy failure and single-handedly maintaining the fragile balance of systemic blood sugar levels.
Slide 10: The Thermodynamic Ledger of Gluconeogenesis: The Cost of Glucose Synthesis
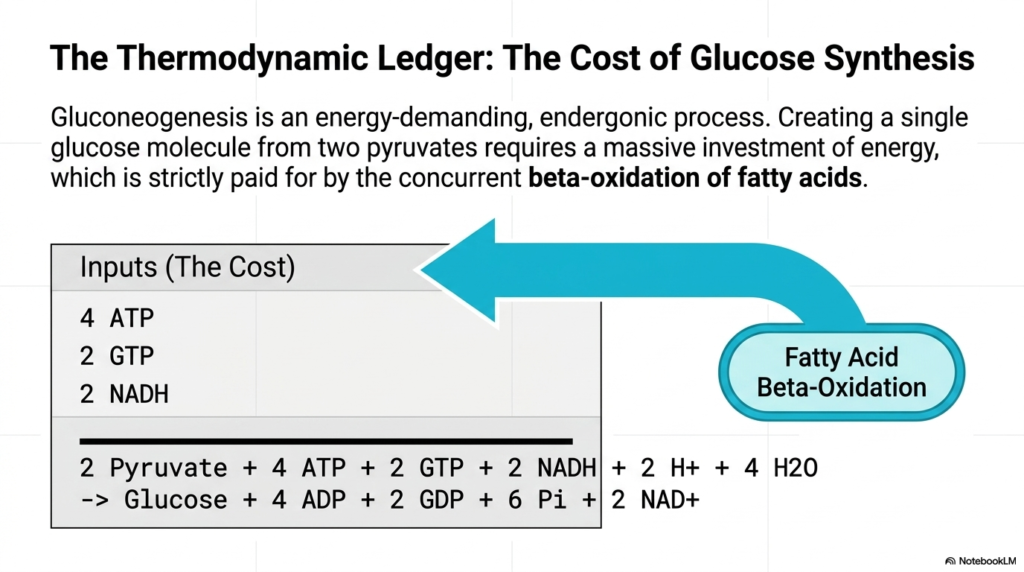
A comprehensive analysis of cellular biochemistry requires a meticulous review of the thermodynamic ledger associated with Gluconeogenesis. This biological pathway is incredibly energy-intensive and highly endergonic, meaning it fundamentally cannot occur without a massive influx of external energy. While breaking down a sugar molecule yields an abundance of free ATP, the cellular effort required to reverse the process and construct a new glucose molecule from scratch exacts a shockingly high metabolic toll on the liver.
The mathematical reality of Gluconeogenesis is clearly illustrated in the provided biochemical ledger. To successfully bind two three-carbon pyruvate molecules into a single six-carbon glucose structure, the cell must sacrifice a significant portion of its stored energy reserves. Specifically, the reaction strictly requires the hydrolysis of four ATP molecules and two GTP molecules, and the consumption of two energy-rich NADH electron carriers. This massive structural investment visually underscores the extreme thermodynamic difficulty of reversing cellular glycolysis.
When evaluating the thermodynamic ledger of Gluconeogenesis, a crucial question emerges from the equation: 2 Pyruvate + 4 ATP + 2 GTP + 2 NADH -> Glucose + 4 ADP + 2 GDP + 6 Pi + 2 NAD+. How does an actively starving cell, which is desperately trying to synthesize energy for the brain, afford this exorbitant molecular price tag? The answer highlights a beautiful synergy within human metabolism: the necessary thermodynamic power is exclusively supplied by an entirely separate biochemical process.
The immense cost of Gluconeogenesis is perfectly offset by the concurrent beta-oxidation of free fatty acids in hepatic tissue. As the liver rapidly burns through lipid reserves, it generates the vital ATP, GTP, and NADH required to power the endogenous glucose assembly line. Therefore, fat burning and sugar synthesis are inextricably linked during starvation. Without the energetic influx provided by lipid oxidation, the endergonic reactions of Gluconeogenesis would grind to a thermodynamic halt.
Slide 11: Local Control of Gluconeogenesis: The Allosteric Seesaw
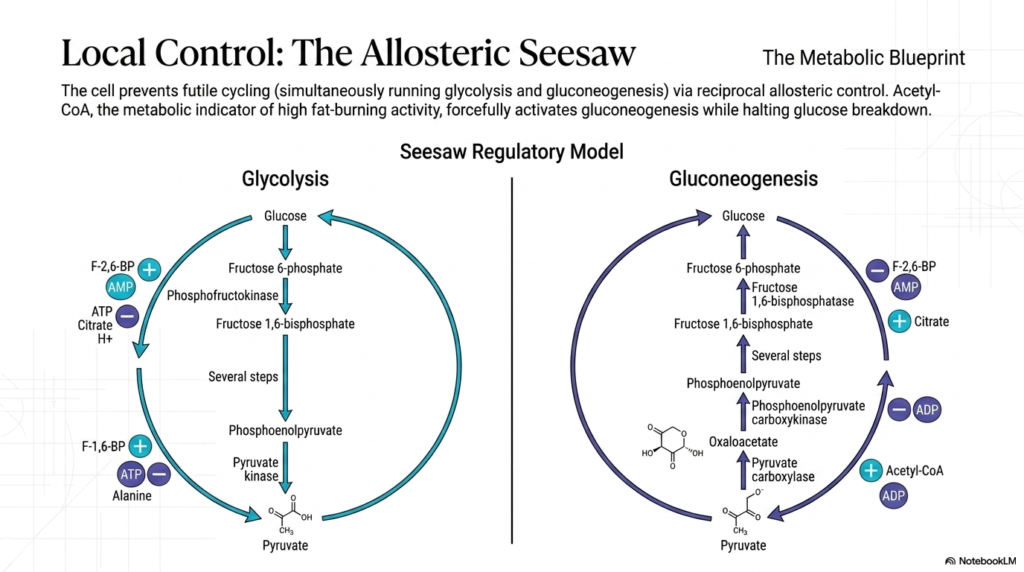
At the microscopic level, the cell must employ strict local control mechanisms to prevent metabolic chaos. A primary danger during starvation is the initiation of futile cycling, a disastrous biological scenario where the cell attempts to run both glycolysis and Gluconeogenesis simultaneously. If this occurred, the cell would endlessly burn high-energy ATP and GTP just to spin its biochemical wheels, producing zero net glucose. To prevent this, metabolism relies on a brilliant system of reciprocal allosteric control that functions like a molecular seesaw.
The concept of the allosteric seesaw guarantees that when one metabolic pathway is forcefully activated, the opposing pathway is simultaneously suppressed. Acetyl-CoA acts as the ultimate local indicator of high fat-burning activity within the mitochondria. When fatty acids are aggressively oxidized, the accumulation of Acetyl-CoA directly binds to and activates Pyruvate Carboxylase, violently pushing the system toward Gluconeogenesis. At the very same moment, it strongly inhibits the pyruvate dehydrogenase complex, effectively shutting down the competing pathways of glucose breakdown.
Conversely, intracellular molecules like AMP act as emergency alarm bells, signaling that the cell’s immediate energy reserves are dangerously depleted. High levels of AMP strongly inhibit the rate-limiting enzyme of Gluconeogenesis, Fructose 1,6-bisphosphatase. Simultaneously, this same AMP molecule acts as a powerful allosteric activator for the glycolytic enzyme Phosphofructokinase-1. This ensures that a starving, low-energy cell will immediately halt the expensive construction of new glucose and instead prioritize the rapid breakdown of available fuels to survive.
Another critical allosteric regulator in this system is Citrate, an intermediate of the citric acid cycle that signals an abundance of thermodynamic energy. Elevated citrate levels positively regulate Gluconeogenesis by activating Fructose 1,6-bisphosphatase while reciprocally inhibiting glycolysis. This intricate web of local molecular interactions ensures that the metabolic assembly line operates only when the liver possesses both the necessary chemical precursors and the abundant energy required to sustain the heavy demands of glucose synthesis.
Slide 12: Hormonal Master Control of Gluconeogenesis: The F-2,6-BP Switch
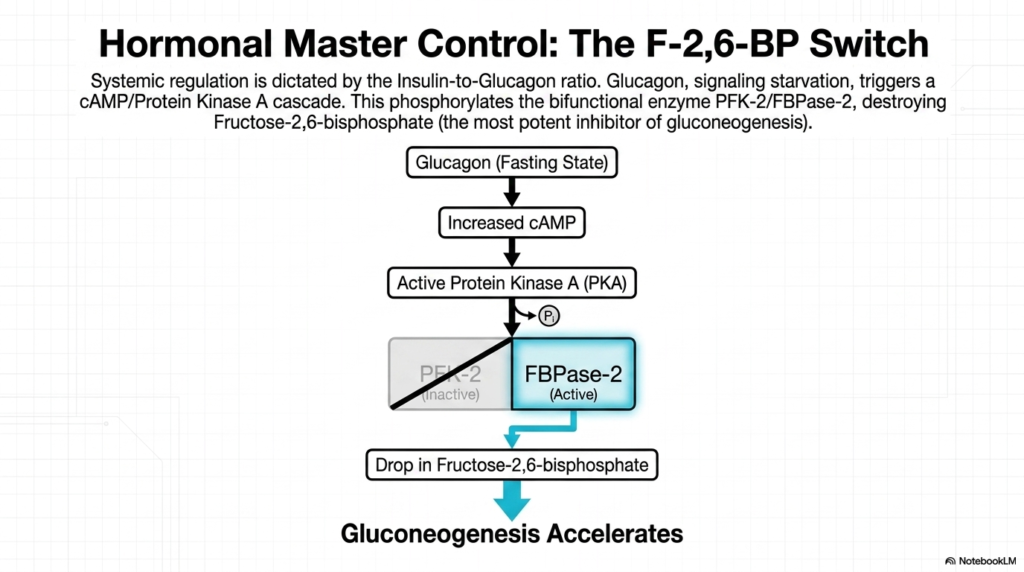
While local allosteric molecules manage cellular logistics, the overarching pace of Gluconeogenesis is dictated by powerful systemic hormonal signals. The biological master control switch for human metabolism is determined by the crucial Insulin-to-Glucagon ratio circulating within the bloodstream. During periods of extended fasting, the pancreas detects falling blood sugar levels and secretes massive quantities of glucagon. This hormone acts as a systemic distress signal, binding directly to specialized receptors on the outer membrane of liver cells to initiate a widespread metabolic response.
The binding of glucagon triggers a powerful intracellular cascade, rapidly increasing cyclic AMP (cAMP) production within the hepatocyte. This surge in cAMP subsequently activates Protein Kinase A (PKA), a master phosphorylating enzyme that fundamentally alters the behavior of internal cellular machinery. PKA aggressively targets a unique bifunctional enzyme, PFK-2/FBPase-2, and covalently attaches a phosphate group to it. This specific modification is the primary mechanism by which fasting directly influences the rate of Gluconeogenesis.
The phosphorylation of this bifunctional protein yields a dramatic result: it completely inactivates the PFK-2 domain while simultaneously hyperactivating the FBPase-2 domain. This activated FBPase-2 acts as molecular scissors, rapidly destroying intracellular reserves of Fructose-2,6-bisphosphate (F-2,6-BP). The rapid destruction of this specific molecule is arguably the single most important regulatory event in fasting metabolism, as F-2,6-BP is the most potent naturally occurring inhibitor of the entire gluconeogenesis pathway.
With the rapid, widespread drop in Fructose-2,6-bisphosphate levels, the heavy biochemical brakes are finally removed from the system. Relieved of this potent inhibition, the key enzymes of Gluconeogenesis drastically accelerate their activity. Through this elegant hormonal cascade, a systemic signal of starvation is flawlessly translated into a massive spike in cellular biosynthesis, ensuring that the liver rapidly produces and exports life-saving glucose to nourish the brain before permanent neurological damage can occur.
Slide 13: Clinical Pathology Impairing Gluconeogenesis: Von Gierke Disease

The profound importance of Gluconeogenesis becomes devastatingly clear when examining what happens when the pathway genetically fails. Clinical pathology frequently intersects with biochemistry, and Von Gierke Disease, also known as Glycogen Storage Disease Type 1, is the premier diagnostic example. This severe metabolic disorder arises from a rare genetic mutation resulting in a complete deficiency of the enzyme Glucose-6-phosphatase. Without this crucial protein, the liver is stripped of its ability to perform the final, necessary step in glucose release.
The diagnostic pathology table clearly outlines the catastrophic chain reaction that occurs when this specific enzyme is missing. Because the liver cannot hydrolyze the phosphate group, the final product of Gluconeogenesis, Glucose-6-phosphate, accumulates heavily within the cell. The hepatocyte becomes utterly incapable of releasing free glucose into the bloodstream, leading to severe, life-threatening fasting hypoglycemia. Without medical intervention, the patient’s brain is rapidly starved of its obligate fuel source mere hours after their last carbohydrate-rich meal.
The absence of this single enzymatic step completely deranges multiple interacting metabolic pathways. The excessive accumulation of Glucose-6-phosphate drives glycogen synthesis to proceed uncontrollably, trapping vast quantities of complex sugars within the liver tissue and resulting in massive, painful hepatomegaly. Furthermore, because the primary exit route is structurally barricaded, the heavily backed-up intermediates of Gluconeogenesis are forcibly shunted away from sugar production and aggressively diverted down alternative biochemical avenues, wreaking systemic havoc on the patient’s internal chemistry.
This severe molecular traffic jam forces accumulated pyruvate to spontaneously convert into lactic acid, causing a dangerous drop in blood pH known as lactic acidosis. Simultaneously, the trapped intracellular carbons are continuously shunted into the lipid and purine biosynthesis cascades. This relentless misdirection of substrates biochemically manifests as extreme hypertriglyceridemia and hyperuricemia. By analyzing Von Gierke Disease, medical professionals can truly grasp how a single broken node in Gluconeogenesis can completely destabilize the entire human metabolic network.
Slide 14: Metabolic Sabotage of Gluconeogenesis: Enzymatic Blocks vs. Redox Imbalance
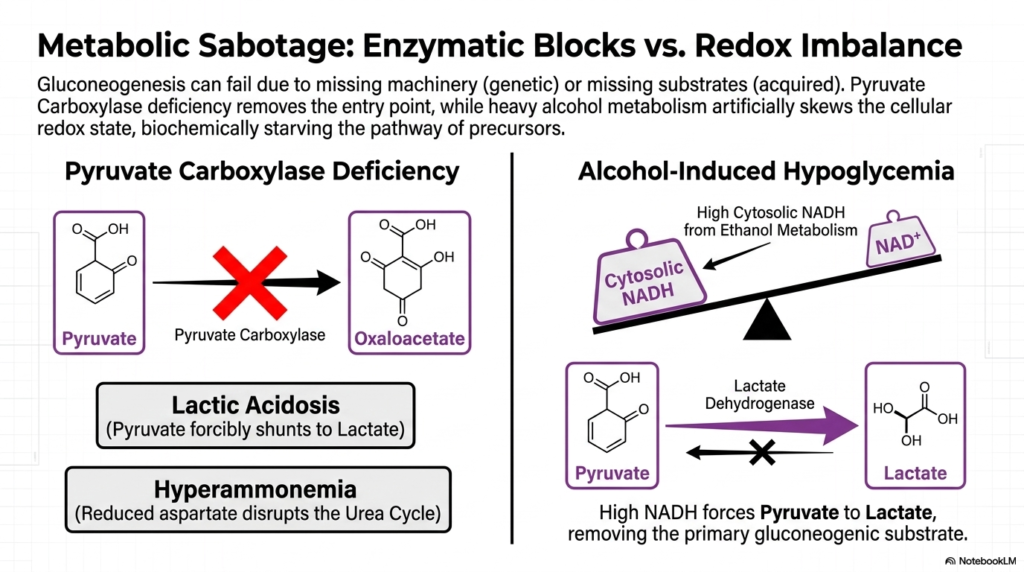
Beyond inherited genetic defects, the delicate machinery of Gluconeogenesis can also be severely compromised by acquired metabolic sabotage. Endogenous glucose production relies on a fragile equilibrium among functional enzymes and balanced chemical conditions. When either of these parameters is disrupted, the biosynthetic assembly line crashes. Two prominent examples of this sabotage are Pyruvate Carboxylase deficiency, representing a hard enzymatic block, and severe alcohol intoxication, which fundamentally deranges the internal cellular redox state to halt production.
A genetic deficiency in Pyruvate Carboxylase physically removes the absolute primary entry point for the entire Gluconeogenesis pathway. Without this critical enzyme, pyruvate cannot enter the mitochondria to form oxaloacetate. Consequently, the accumulated cytosolic pyruvate is forcibly shunted away from glucose synthesis and irreversibly converted into lactate, resulting in severe clinical lactic acidosis. Furthermore, this specific mitochondrial blockade drastically reduces cellular aspartate production, a necessary structural intermediate for maintaining the closely associated urea cycle.
The subsequent disruption of the urea cycle inevitably leads to hyperammonemia, a highly toxic accumulation of ammonia in the bloodstream. This vividly demonstrates how the failure of Gluconeogenesis extends its destructive reach far beyond simple hypoglycemia. Alternatively, alcohol-induced hypoglycemia represents an acquired form of metabolic sabotage. The rapid breakdown of massive amounts of ethanol forces the liver to generate an extremely high concentration of cytosolic NADH, aggressively skewing the delicate cellular redox balance toward a highly reduced state.
This artificially high concentration of NADH violently tips the biochemical scales, heavily forcing the enzyme Lactate Dehydrogenase to convert available pyruvate strictly into lactate. By driving the reaction entirely in the wrong direction, ethanol metabolism physically starves the Gluconeogenesis pathway of its absolute most important precursor. Without sufficient pyruvate to feed the mitochondrial entry point, the liver simply cannot construct new sugar, leaving the intoxicated patient highly vulnerable to severe, medically profound drops in blood glucose levels.
Slide 15: Gluconeogenesis as the Integrated Biosynthetic Network
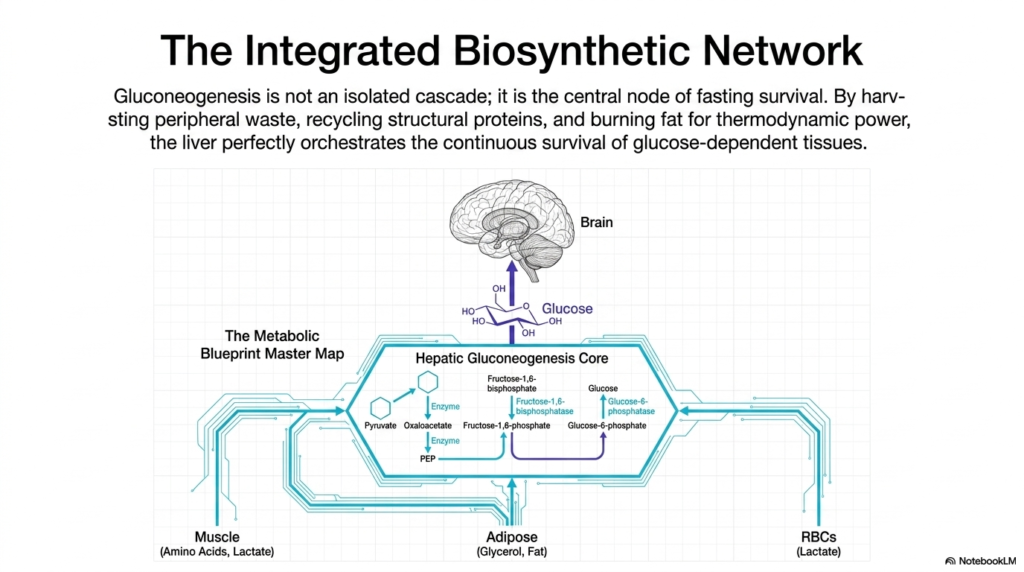
The concluding slide of this presentation synthesizes the immense complexity of metabolism, portraying Gluconeogenesis not merely as a linear sequence of isolated chemical reactions, but as the master integrated biosynthetic network of the human body. During periods of severe nutritional deprivation, the liver serves as a vital biological orchestrator. It seamlessly coordinates the intricate functions of multiple distant organ systems, transforming chaotic metabolic waste products into a unified, life-sustaining stream of pure cellular energy.
The metabolic blueprint master map visually reinforces how the hepatic core harvests necessary resources from the periphery. The liver actively scavenges circulating lactate discarded by red blood cells, ensuring that energetic potential is never wasted. Simultaneously, it oversees the aggressive dismantling of skeletal muscle, importing a steady flow of structural amino acids to serve as fresh carbon backbones. This incredible level of systemic coordination is absolutely crucial to guarantee that the primary machinery of Gluconeogenesis is never starved of raw materials.
To fund this monumental structural effort, the liver must also communicate with widespread adipose tissue. Massive quantities of stored triglycerides are continuously broken down into glycerol—which feeds directly into the pathway—and free fatty acids. The relentless beta-oxidation of these liberated fats inside the liver provides the massive thermodynamic power necessary to drive the highly endergonic reactions of Gluconeogenesis. Therefore, endogenous glucose production is the ultimate culmination of fat, protein, and carbohydrate metabolism working in perfect, desperate synergy.
Ultimately, this beautifully integrated biochemical system exists for one primary purpose: continuous survival. By meticulously recycling peripheral waste, harvesting structural proteins, and aggressively burning fat for thermodynamic power, the liver perfectly orchestrates the unyielding operation of Gluconeogenesis. This biological masterpiece guarantees that the brain, erythrocytes, and other obligate tissues receive a continuous, uninterrupted supply of glucose, allowing human beings to survive and even thrive during prolonged periods of starvation, famine, and extreme physiological stress.
Please read our Content Disclaimer Statement.
Check out our social media channels: