55. Amino Acid Biosynthesis: Pathways and Regulation
Have you ever wondered how cells build the fundamental building blocks of life from scratch? Proteins perform almost every critical function in the human body, but they all begin as simple molecules. This slide deck breaks down the complex biochemical structures and processes involved in the creation of these vital molecules. Designed for college and medical students, the presentation will explore the intricate metabolic pathways, nitrogen assimilation strategies, and the elegant regulatory mechanisms that govern these processes.
Slide 1: Introduction to Amino Acid Biosynthesis
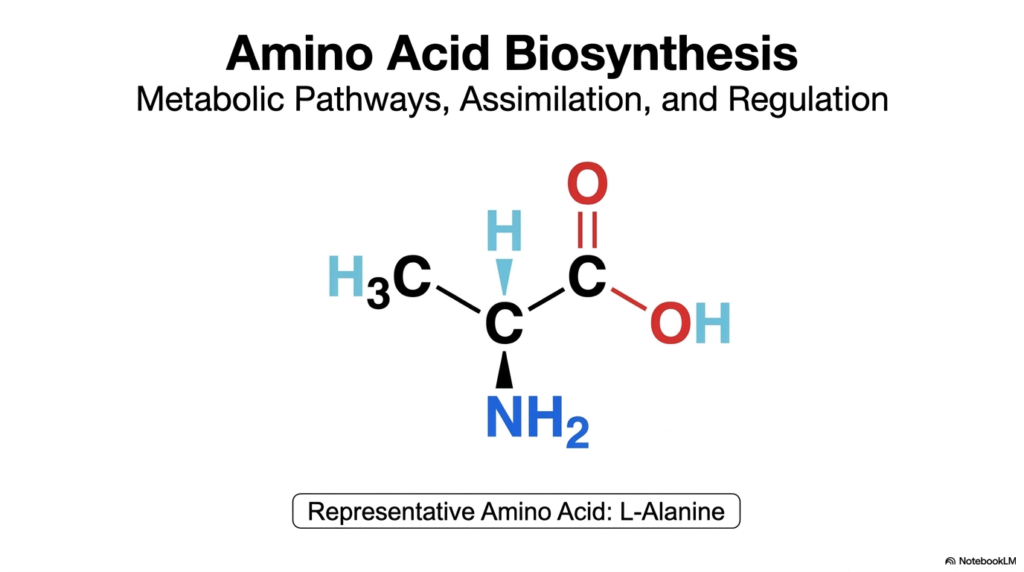
The journey into the molecular machinery of cells begins with a foundational understanding of how organisms construct their most essential building blocks. The study of amino acid biosynthesis reveals the remarkable metabolic pathways, nitrogen assimilation strategies, and regulatory frameworks that sustain life. This introductory slide visually centers on L-Alanine, a representative molecule that perfectly illustrates the classic structure shared by these biological monomers. Understanding amino acid biosynthesis is crucial for medical and college biochemistry students, as it connects basic organic chemistry to large-scale physiological metabolism. It provides the biochemical context needed to appreciate how organisms harness energy to build complexity.
The chemical structure of L-Alanine highlighted here features the central alpha-carbon attached to an amino group, a carboxyl group, and a simple methyl side chain. In the context of amino acid biosynthesis, the focus often shifts to how that crucial nitrogen atom is acquired and incorporated into the carbon skeleton. The cell must employ highly coordinated enzymatic steps to form these bonds without wasting cellular energy. This visual serves as a reminder of the ultimate metabolic goal: constructing specific, stereochemically precise molecules that can be strung together into functional proteins. Every bond represents a carefully regulated metabolic investment by the cell.
By starting with this simple model, the stage is set for exploring the broader biochemical principles of amino acid biosynthesis. The overarching themes of this deck include the diverse origins of the necessary carbon skeletons, the biological imperative of fixing atmospheric nitrogen, and the meticulous ways cells prevent the wasteful overproduction of these molecules. Mastery of these core concepts provides a solid foundation for understanding human metabolic diseases, dietary nutritional requirements, and the fascinating evolutionary history of cellular metabolism. Let this precise chemical structure of L-Alanine anchor the complex biochemical processes, enzymes, and regulatory mechanisms detailed in the following sections.
Slide 2: Regulating Amino Acid Biosynthesis via Feedback Inhibition
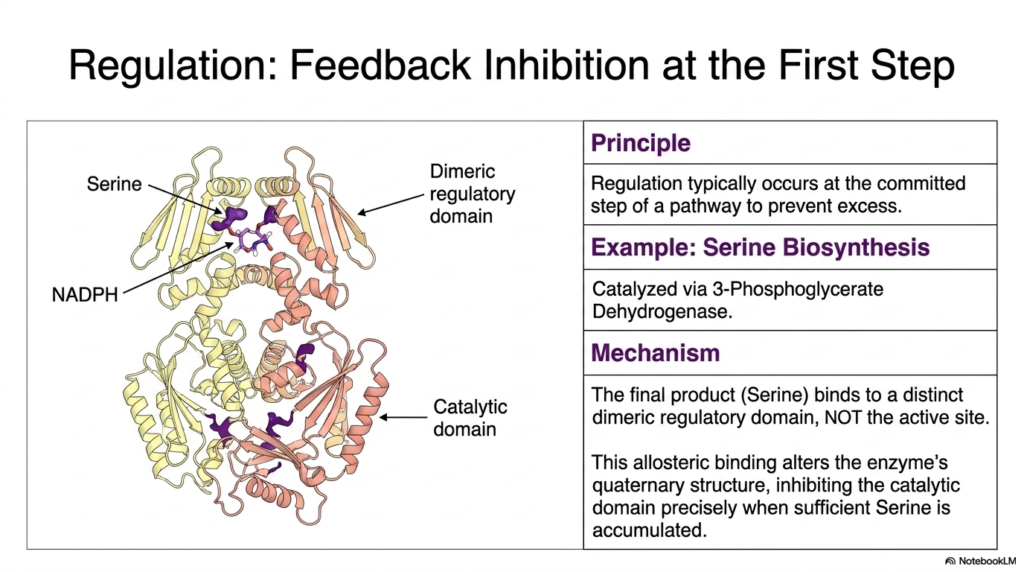
One of the most elegant principles in cellular metabolism is the ability to halt production when resources are sufficient. In the realm of amino acid biosynthesis, regulation typically occurs at the committed step of a metabolic pathway to prevent the wasteful accumulation of excess products. This slide illustrates the classic mechanism of feedback inhibition using serine production as a prime example. For medical students, grasping this concept is vital, as dysregulation in these pathways often underlies metabolic disorders. The enzyme 3-Phosphoglycerate Dehydrogenase catalyzes the first critical step in this pathway, making it a prime target for molecular control.
The visual provides a detailed view of the quaternary structure of the regulatory enzyme, highlighting its distinct dimeric regulatory and catalytic domains. During amino acid biosynthesis, the final product—in this case, serine—does not bind to the active site where the chemical reaction occurs. Instead, serine acts as an allosteric inhibitor by binding specifically to the regulatory domain. This allosteric binding event is a cornerstone of enzyme regulation. When the cellular concentration of serine reaches a sufficient threshold, these molecules attach to the regulatory sites, signaling that the cell has met its metabolic demands for this particular building block.
This binding triggers a crucial conformational change across the enzyme’s structure. The shift in the quaternary architecture effectively inhibits the catalytic domain, immediately pausing the progression of amino acid biosynthesis. This precise mechanism ensures that the cell conserves energy and vital carbon precursors, such as 3-phosphoglycerate, redirecting them to other active metabolic pathways. By utilizing feedback inhibition at the very first committed step, the cell avoids expending resources on intermediate molecules that are no longer needed. This beautiful biochemical logic perfectly balances supply and demand, showcasing the highly evolved efficiency of cellular metabolic networks.
Slide 3: Nitrogen Fixation in Amino Acid Biosynthesis

Before any organism can begin the complex process of amino acid biosynthesis, it must first secure a usable source of nitrogen. Atmospheric nitrogen exists abundantly as a diatomic gas, but its triple bond renders it highly inert and biologically inaccessible to most life forms. To enter the natural nitrogen cycle and become incorporated into biological molecules, this gas must be reduced to ammonia. This slide contrasts the immense energetic hurdles of industrial nitrogen fixation with the remarkable efficiency of biological systems. Understanding this foundational step is essential for appreciating the broader scope of amino acid biosynthesis in the biosphere.
The industrial Haber-Bosch process highlights the extreme difficulty of breaking the nitrogen triple bond. As the slide details, this artificial method requires temperatures of 500 degrees Celsius and staggering pressures of 300 atmospheres to force the chemical reduction. In stark contrast, biological nitrogen fixation achieves the exact same chemical feat at a standard 25 degrees Celsius and a single atmosphere of pressure. This biological miracle is primarily driven by anaerobic microorganisms and symbionts, such as Rhizobium bacteria found in the root nodules of legume plants, which account for about sixty percent of all naturally fixed nitrogen on Earth.
Instead of relying on extreme heat and pressure, microorganisms power this reduction using biochemical energy. The biological process is driven by ATP hydrolysis and the transfer of high-energy electrons from reduced ferredoxin. This enzymatic mastery allows the biosphere to generate the essential ammonia required to support all downstream amino acid biosynthesis. For medical and biochemistry students, this comparison beautifully illustrates the power of biological catalysts. The ability of specialized enzymes to overcome such a massive thermodynamic barrier at ambient conditions remains one of the most awe-inspiring achievements of natural evolution and cellular metabolism.
Slide 4: The Nitrogenase Complex and Amino Acid Biosynthesis
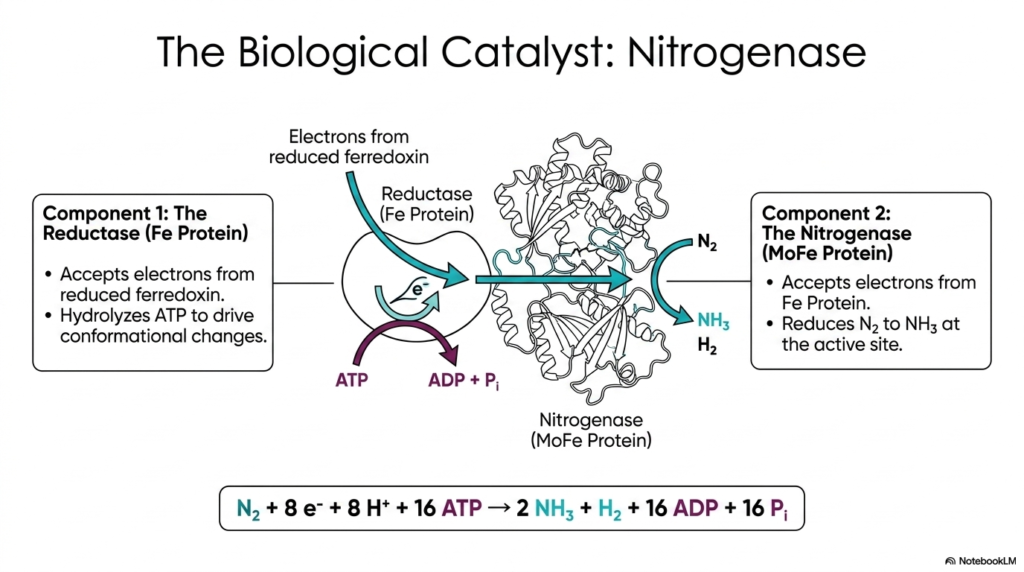
The astonishing biological nitrogen fixation discussed previously is entirely orchestrated by a sophisticated enzyme complex known as Nitrogenase. This slide breaks down the structural and functional components of this critical biological catalyst, which provides the raw ammonia necessary for all subsequent amino acid biosynthesis. The Nitrogenase complex consists of two primary components working in tandem: the Reductase, or Iron (Fe) Protein, and the Nitrogenase itself, known as the Molybdenum-Iron (MoFe) Protein. For students diving into advanced biochemistry, understanding this two-part macromolecular machine is key to grasping how organisms channel energy to perform highly unfavorable chemical reductions.
The process begins with the Fe Protein, which acts as the dedicated electron donor. It accepts high-energy electrons from reduced ferredoxin and utilizes the energy from ATP hydrolysis to drive essential conformational changes. This structural shift allows the Fe Protein to securely dock with the MoFe Protein and transfer its electron payload. This continuous cycle of ATP hydrolysis and electron transfer is a massive energetic investment for the cell, highlighting the absolute necessity of securing bioavailable nitrogen for amino acid biosynthesis. The MoFe protein then serves as the ultimate catalytic site where nitrogen gas is reduced.
At the active site of the MoFe Protein, the accumulated electrons are finally used to directly reduce atmospheric nitrogen to ammonia and hydrogen gas. The summary equation at the bottom of the slide perfectly encapsulates the high metabolic cost of this reaction. Fixing a single molecule of nitrogen requires eight electrons, eight protons, and a staggering sixteen molecules of ATP. This monumental expenditure of biological energy underscores how critical ammonia is for sustaining life. Without the continuous operation of the Nitrogenase complex, the global amino acid biosynthetic pipeline would collapse instantly, halting protein production worldwide.
Slide 5: The Fe Protein’s Role in Amino Acid Biosynthesis
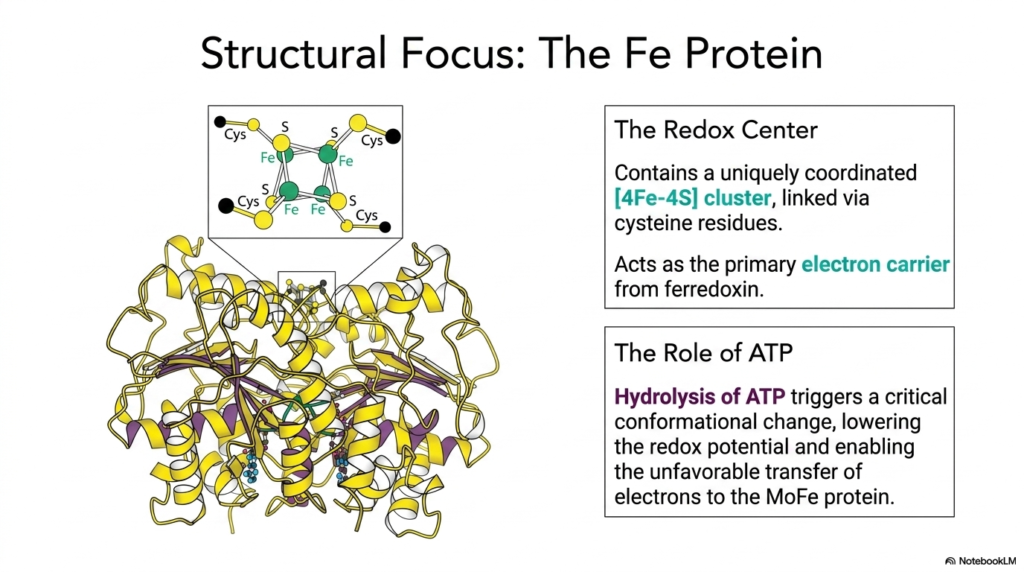
To fully appreciate the mechanics of nitrogen fixation, which sets the stage for amino acid biosynthesis, one must examine the molecular architecture of the Fe Protein. This slide provides a structural focus on the Reductase component, highlighting the vital redox center that makes the entire electron transfer process possible. The Fe Protein contains a uniquely coordinated iron-sulfur cluster, specifically a [4Fe-4S] configuration, which is covalently linked to the protein backbone via four distinct cysteine residues. This specialized metal cluster acts as the primary electron carrier, receiving high-energy electrons from upstream metabolic sources like ferredoxin.
Understanding this structure is essential because the [4Fe-4S] cluster must temporarily house the electron before passing it to the larger catalytic unit. However, transferring an electron to the MoFe Protein is thermodynamically unfavorable. This is where the crucial role of cellular energy comes into play, linking the organism’s energy stores directly to the foundational steps of amino acid biosynthesis. The Fe Protein is not merely a passive wire for electrons; it is an active, ATP-driven molecular machine that must physically maneuver to complete its designated biochemical task.
The binding and subsequent hydrolysis of ATP trigger a massive and critical conformational change across the Fe Protein’s three-dimensional structure. This physical shift forcefully lowers the redox potential of the bound electron, essentially priming it for transfer. This ATP-driven mechanical change is what enables the otherwise unfavorable jump of the electron into the MoFe protein. By coupling ATP hydrolysis to electron transfer, the cell meticulously controls the flow of reducing power. This elegant structural mechanism ensures a steady supply of ammonia, feeding directly into the pathways of amino acid biosynthesis that build life’s essential proteins.
Slide 6: The MoFe Protein in Amino Acid Biosynthesis
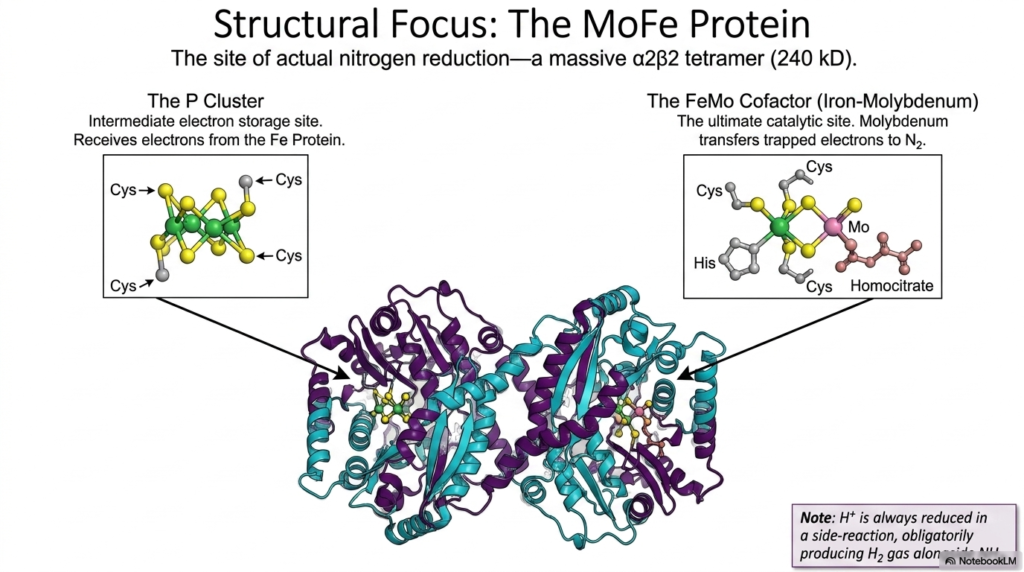
Following the electron’s journey brings us to the MoFe Protein, the massive structural complex where nitrogen is actually reduced. This massive tetrameric enzyme is the heavy lifter that directly supplies the ammonia required for all downstream amino acid biosynthesis. The slide highlights two critical metal centers hidden deep within the protein’s architecture: the P Cluster and the FeMo Cofactor. When the Fe Protein successfully transfers an electron, it does not go straight to the nitrogen gas. Instead, it is first received by the P Cluster, a highly specialized intermediate electron storage site.
The P Cluster, composed of linked iron and sulfur atoms, briefly holds the electrons before funneling them toward the ultimate catalytic destination. This temporary staging area ensures a controlled and steady delivery of reducing power. The electrons eventually arrive at the FeMo Cofactor, an incredibly complex metal cluster featuring iron, sulfur, molybdenum, and homocitrate. It is at this precise biochemical location that atmospheric nitrogen binds and is methodically reduced. The continuous supply of ammonia generated here is the ultimate prerequisite for amino acid biosynthesis across the entire biological spectrum.
The structural intricacy of the FeMo Cofactor is a testament to the evolutionary challenge of breaking the nitrogen triple bond. The molybdenum atom is thought to play a pivotal role in transferring the trapped electrons directly to the nitrogen substrate. Interestingly, the slide notes that the reduction of protons into hydrogen gas is an obligate side reaction of this complex mechanism. This fascinating biochemical quirk means that even the most efficient biological nitrogen fixation expends energy producing hydrogen gas alongside the ammonia desperately needed for amino acid biosynthesis.
Slide 7: Glutamate’s Role in Amino Acid Biosynthesis
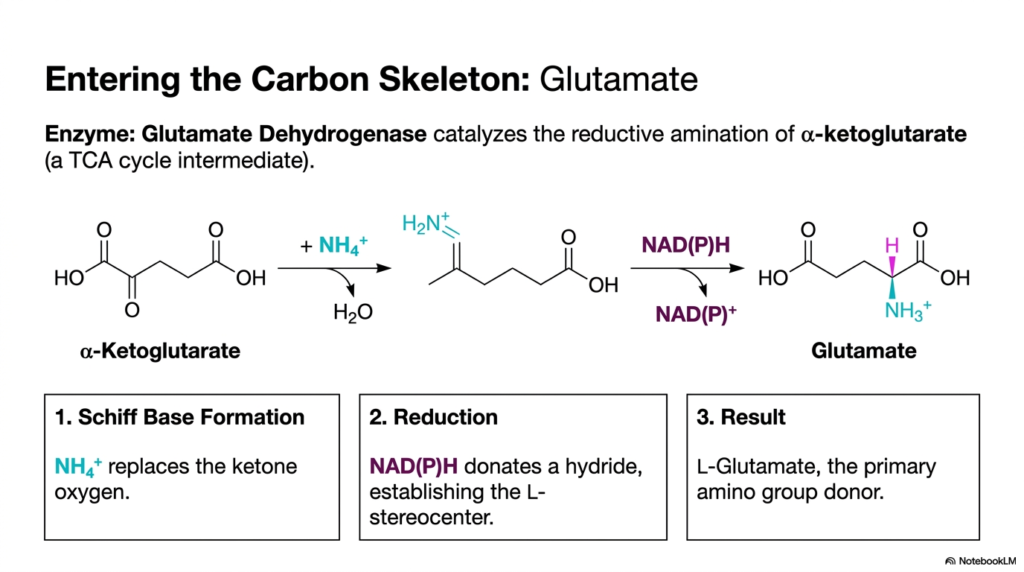
Once inorganic nitrogen is successfully reduced to ammonia, the cell faces a new challenge: safely incorporating this toxic molecule into a stable, organic carbon skeleton. This slide outlines the crucial entry point for nitrogen assimilation in amino acid biosynthesis. The primary gateway is the reductive amination of alpha-ketoglutarate, a ubiquitous intermediate directly pulled from the TCA cycle. The enzyme responsible for this fundamental biochemical transformation is Glutamate Dehydrogenase. This reaction is of paramount importance to medical students, as glutamate serves as the universal nitrogen donor for nearly all other cellular nitrogen requirements.
The chemical mechanism proceeds through a highly specific sequence of events. First, the incoming ammonia replaces the ketone oxygen on the alpha-ketoglutarate molecule, forming an unstable intermediate known as a Schiff base. This step effectively tethers the free nitrogen to the carbon skeleton, neutralizing the toxicity of free cellular ammonia. Following this attachment, a vital reduction step occurs. The cell expends reducing power as NAD(P)H, which donates a hydride ion to the intermediate. This reduction is a hallmark of anabolic pathways such as amino acid biosynthesis, which require significant metabolic investment.
The donation of the hydride ion does more than just complete the chemical bond; it meticulously establishes the required L-stereocenter of the newly formed molecule. The final result of this elegant enzymatic process is L-Glutamate. By transforming a simple TCA cycle intermediate into glutamate, the cell establishes the primary amino group donor required to build the rest of the proteinogenic building blocks. This single, highly regulated reaction bridges the gap between carbohydrate metabolism and amino acid biosynthesis, proving that all cellular metabolic networks are deeply and seamlessly interconnected.
Slide 8: Glutamine as a Sink in Amino Acid Biosynthesis
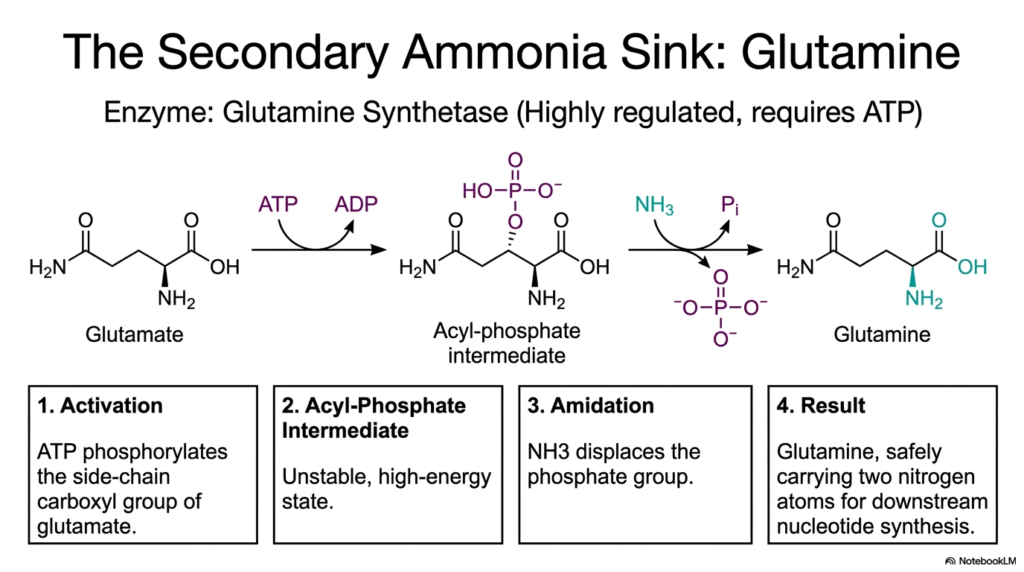
While glutamate serves as the primary nitrogen donor, cells require a secondary mechanism to safely store and transport additional nitrogen atoms for complex metabolic needs. This slide introduces Glutamine Synthetase, a highly regulated enzyme responsible for creating the body’s secondary ammonia sink. In the broader context of amino acid biosynthesis, glutamine is absolutely essential because it safely carries two nitrogen atoms, making it a powerful precursor for downstream nucleotide and protein synthesis. The conversion of glutamate to glutamine is an energy-dependent process that requires strict cellular oversight to manage nitrogen resources effectively.
The biochemical mechanism of Glutamine Synthetase operates in two distinct, beautifully coordinated steps. The first phase is activation, in which the enzyme uses ATP to phosphorylate the side-chain carboxyl group of the existing glutamate. This phosphorylation creates a highly unstable, high-energy acyl-phosphate intermediate. By investing a molecule of ATP, the cell essentially primes the glutamate structure, making it highly reactive for the next chemical step. This energetic priming is a recurring motif across the various pathways of amino acid biosynthesis, ensuring that unfavorable reactions proceed smoothly.
Following the formation of this unstable intermediate, the amidation phase occurs. A second ammonia molecule acts as a nucleophile, forcefully displacing the newly attached phosphate group. This displacement completes the formation of the molecular structure, yielding L-Glutamine. By attaching this second amino group, the cell creates a potent, non-toxic nitrogen carrier that can safely circulate through the bloodstream or cellular cytoplasm. The careful regulation of this specific enzyme ensures that the intricate networks of amino acid biosynthesis have a constant, reliable supply of nitrogen for synthesizing purines, pyrimidines, and other complex biological macromolecules.
Slide 9: The Metabolic Blueprint of Amino Acid Biosynthesis
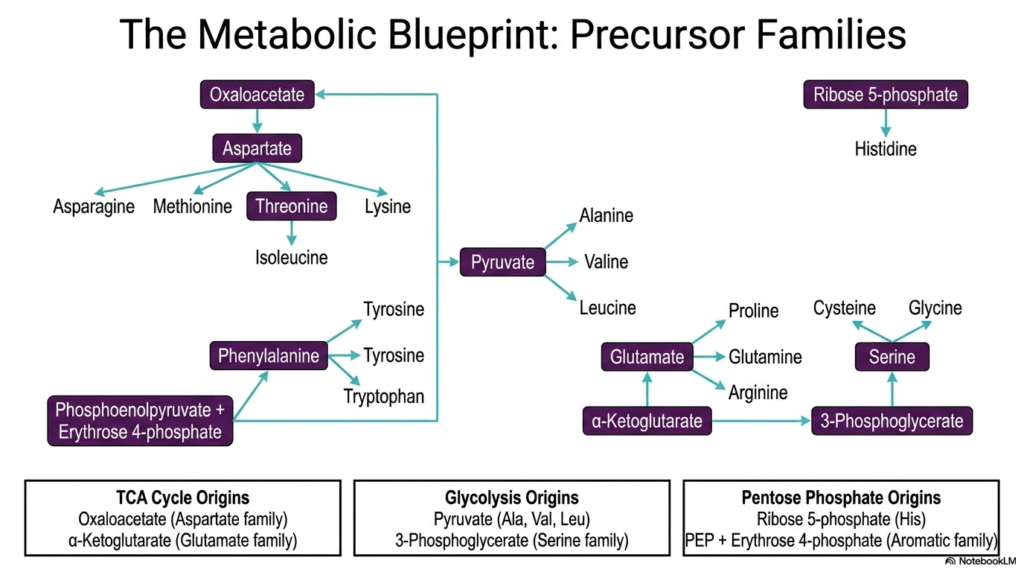
To master the immense complexity of cellular metabolism, students must visualize how diverse molecular pathways interconnect to form a cohesive biological system. This slide presents the comprehensive metabolic blueprint of amino acid biosynthesis, organizing the synthesis of all twenty standard building blocks into distinct precursor families. Rather than memorizing twenty isolated pathways, biochemistry students can group these molecules based on their metabolic origins. The carbon skeletons required for these intricate synthetic processes are strategically siphoned from three central hubs of cellular metabolism: the TCA cycle, Glycolysis, and the Pentose Phosphate pathway.
The visual flow chart elegantly demonstrates how simple central metabolites branch out to form complex molecular structures. For instance, alpha-ketoglutarate from the TCA cycle gives rise to the glutamate family, which includes glutamine, proline, and arginine. Similarly, oxaloacetate gives rise to the aspartate family, serving as the precursor for methionine, threonine, and lysine. Meanwhile, intermediates from glycolysis, such as pyruvate and 3-phosphoglycerate, act as the starting materials for the alanine and serine families, respectively. This organization highlights a beautiful principle of amino acid biosynthesis: nature utilizes pre-existing metabolic infrastructure to efficiently generate new biomolecules.
The inclusion of the Pentose Phosphate pathway adds another layer of complexity, providing the ribose 5-phosphate required for histidine production, as well as erythrose 4-phosphate, which joins with phosphoenolpyruvate to build the complex aromatic rings of phenylalanine, tyrosine, and tryptophan. This blueprint showcases the remarkable efficiency of cellular design. By deriving all proteinogenic precursors from just a handful of central metabolites, the cell tightly integrates energy production with cellular growth. This holistic view of amino acid biosynthesis is absolutely critical for understanding how metabolic fluxes shift during states of starvation, disease, or rapid cellular proliferation.
Slide 10: Transamination and Amino Acid Biosynthesis

A defining characteristic of biological proteins is their strict reliance on molecules with a specific, uniform spatial configuration. This slide explores how the L-stereochemistry of almost all proteinogenic building blocks is rigorously enforced during amino acid biosynthesis. The cell achieves this incredible structural consistency through a single, versatile class of chemical reactions known as transamination. Catalyzed by enzymes called aminotransferases, these reactions are the primary means of shuttling nitrogen around the cell. Understanding this mechanism is vital for medical students, as blood tests often measure specific aminotransferases to assess liver health and metabolic function.
The core principle of transamination involves the direct transfer of an amino group from a donor molecule to an alpha-keto acid acceptor. The slide beautifully illustrates this concept using the classic exchange between oxaloacetate and glutamate. During this reversible reaction, glutamate donates its amino group to oxaloacetate. Through this precise exchange, the original alpha-keto acid (oxaloacetate) is instantly transformed into a new building block (aspartate). Simultaneously, the original donor molecule (glutamate) reverts back into its alpha-keto acid form (alpha-ketoglutarate). This chemical swapping is a cornerstone of amino acid biosynthesis, allowing cells to rapidly balance their molecular inventories.
What makes transamination truly remarkable is its ability to stereospecifically place the new amino group, guaranteeing that only the L-isomer is produced. The aminotransferase enzymes perfectly orient the incoming substrates, preventing the formation of biologically useless D-isomers. By utilizing glutamate as a universal nitrogen donor, the cell can seamlessly synthesize a wide variety of structures through simple, single-step transamination reactions. This elegant enzymatic strategy drastically simplifies the immense complexity of amino acid biosynthesis, ensuring that the cell always has a readily available pool of correctly folded building blocks to sustain continuous protein translation.
Slide 11: Evolution and Amino Acid Biosynthesis
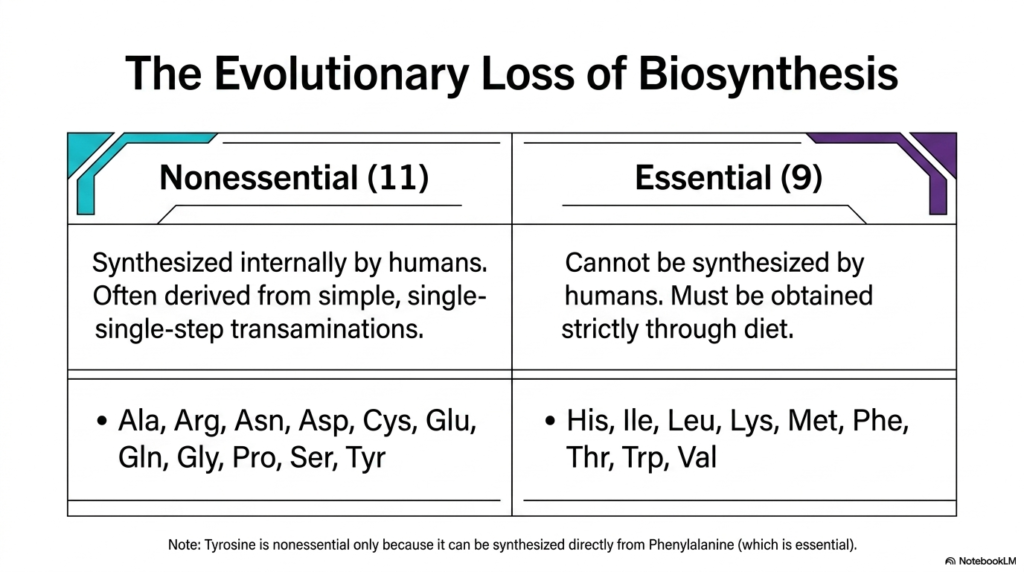
The story of cellular metabolism is deeply intertwined with the evolutionary history of life on Earth. This slide introduces a fundamental biological dichotomy directly related to amino acid biosynthesis: the classification of these molecules into essential and nonessential categories. Over millions of years, human ancestors and other mammalian lineages experienced significant genetic changes that dramatically altered their metabolic capabilities. Understanding which molecular pathways are retained and which are lost is crucial to the fields of nutrition, medicine, and evolutionary biology, as it directly dictates human dietary requirements and physiological health.
As the table illustrates, nonessential molecules are those that humans can synthesize internally from basic metabolic precursors. There are eleven nonessential building blocks, including alanine, aspartate, glutamate, and serine. The pathways for these specific molecules were preserved throughout evolutionary history, largely because they often involve simple, single-step transamination reactions that require minimal cellular energy to maintain. The capacity for internal amino acid biosynthesis of these particular structures ensures that mammalian cells can rapidly adapt to fluctuating metabolic demands without relying entirely on external nutritional sources for their most basic proteinogenic needs.
Conversely, the nine essential building blocks, such as histidine, leucine, and tryptophan, cannot be synthesized by the human body and must be acquired strictly through diet. The complex, multi-step enzymatic pathways required to build these molecules were systematically lost during evolution. By abandoning these energetically expensive anabolic processes, our mammalian ancestors likely gained a selective advantage, conserving massive amounts of molecular resources. This evolutionary trade-off highlights a fascinating aspect of amino acid biosynthesis: sometimes, the most efficient metabolic strategy for a complex organism is to completely outsource the biochemical labor to the plants and microbes in its diet.
Slide 12: The Metabolic Cost of Amino Acid Biosynthesis
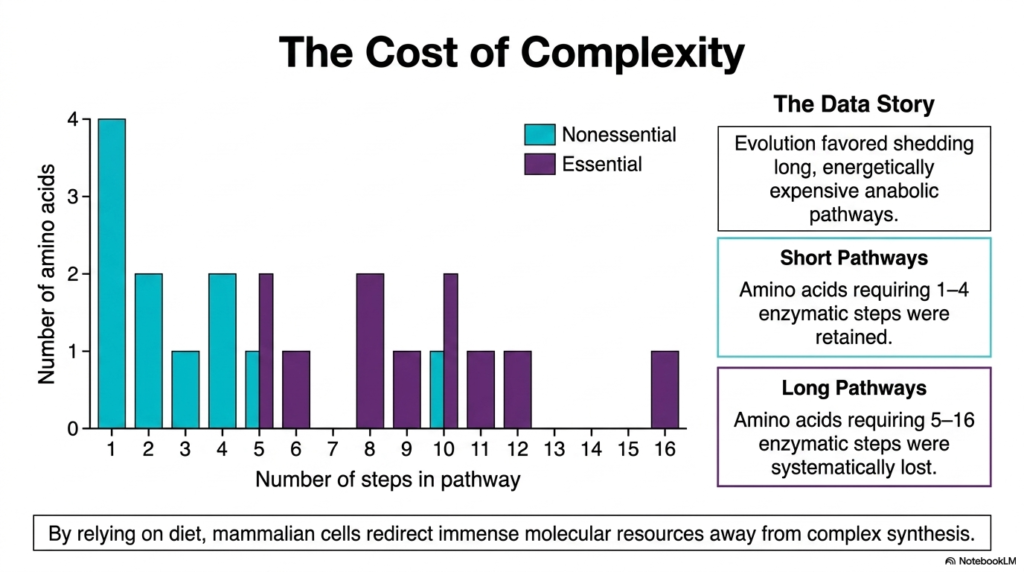
The evolutionary loss of specific metabolic capabilities was not a random genetic occurrence, but rather a calculated consequence of biochemical thermodynamics. This slide visualizes the profound “Cost of Complexity” associated with amino acid biosynthesis. By graphing the number of enzymatic steps required to synthesize each molecule against its essential or nonessential status, a stunning biological data story emerges. This visual perfectly illustrates why human metabolism functions the way it does today. For students of biochemistry, this chart connects molecular energetics directly to evolutionary selection, proving that cellular efficiency dictates survival over millions of years.
The graph clearly demonstrates that evolutionary forces heavily favored the shedding of long, energetically expensive anabolic pathways. Notice how all the nonessential molecules, indicated by the teal bars, cluster on the far left side of the chart. These retained pathways are remarkably short, requiring only one to four enzymatic steps to complete. The human body preserved the genetic instructions for these short pathways because the energetic cost of this specific amino acid biosynthesis remains incredibly low. Cells can easily generate these simple structures on demand without draining their precious reserves of ATP and reducing equivalents.
In sharp contrast, the purple bars representing the essential building blocks stretch far out to the right, with some pathways requiring up to sixteen distinct enzymatic steps. These long pathways were systematically eliminated from the mammalian genome. By relying on dietary intake of these complex molecules, mammalian cells successfully redirected immense molecular resources away from burdensome biosynthetic tasks. This shift allowed organisms to invest that conserved energy into developing complex nervous systems and specialized cellular functions, fundamentally shaping human biology by minimizing the heavy metabolic toll of advanced amino acid biosynthesis.
Slide 13: Managing Branched Pathways in Amino Acid Biosynthesis
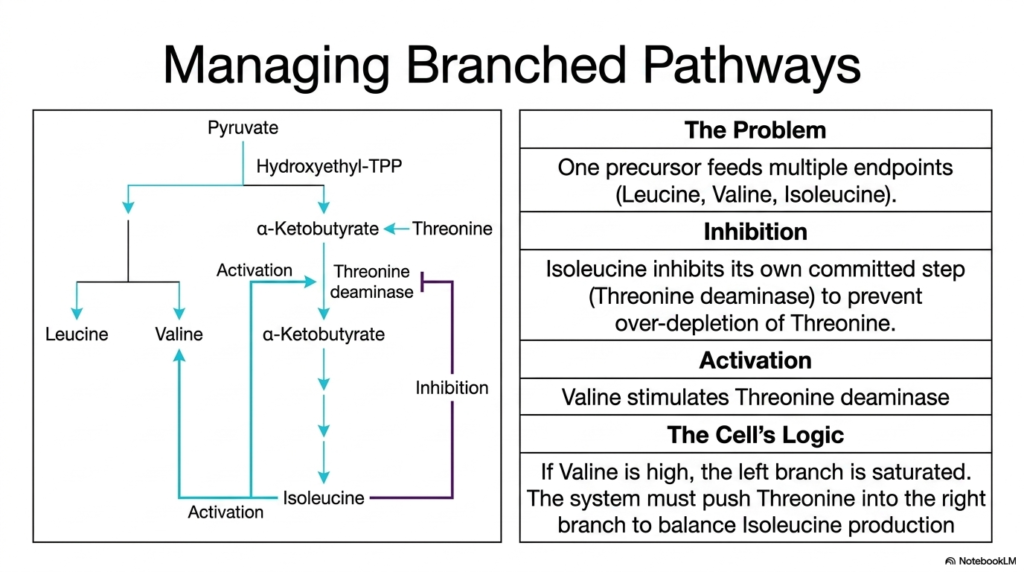
The metabolic networks within a cell are rarely simple, linear assembly lines; they are complex, highly interconnected webs that often share common precursors. This slide addresses the intricate regulatory challenge of managing branched metabolic pathways, a common feature in amino acid biosynthesis. The central problem highlighted here is how a cell manages a single precursor that feeds into multiple different endpoints—in this specific case, the shared pathways for generating leucine, valine, and isoleucine. Biochemistry students must understand these branched dynamics to appreciate how cells maintain molecular homeostasis without overproducing or depleting specific resources.
To balance the production of these related molecules, the cell employs a sophisticated push-and-pull regulatory system. The diagram shows that isoleucine actively inhibits its own committed step by binding to the enzyme threonine deaminase. This negative feedback loop ensures that once sufficient isoleucine is produced, the cell stops cannibalizing its valuable threonine reserves. However, the regulation of this specific amino acid biosynthesis pathway involves more than just simple inhibition. Valine, a product from the opposite side of the branched pathway, acts as an allosteric activator for that exact same enzyme, threonine deaminase.
This dual regulatory mechanism reveals the beautiful logic of cellular metabolism. If valine concentrations are unusually high, it indicates that the left branch of the pathway is completely saturated. To restore molecular balance, the high levels of valine stimulate threonine deaminase, actively pushing the precursor pool down the right branch to increase isoleucine production. This elegant system of cross-regulation ensures that the cell maintains perfectly balanced ratios of all three structures. Such sophisticated molecular communication is a hallmark of amino acid biosynthesis, preventing toxic imbalances and ensuring the continuous, efficient assembly of cellular proteins.
Slide 14: Enzyme Multiplicity in Amino Acid Biosynthesis
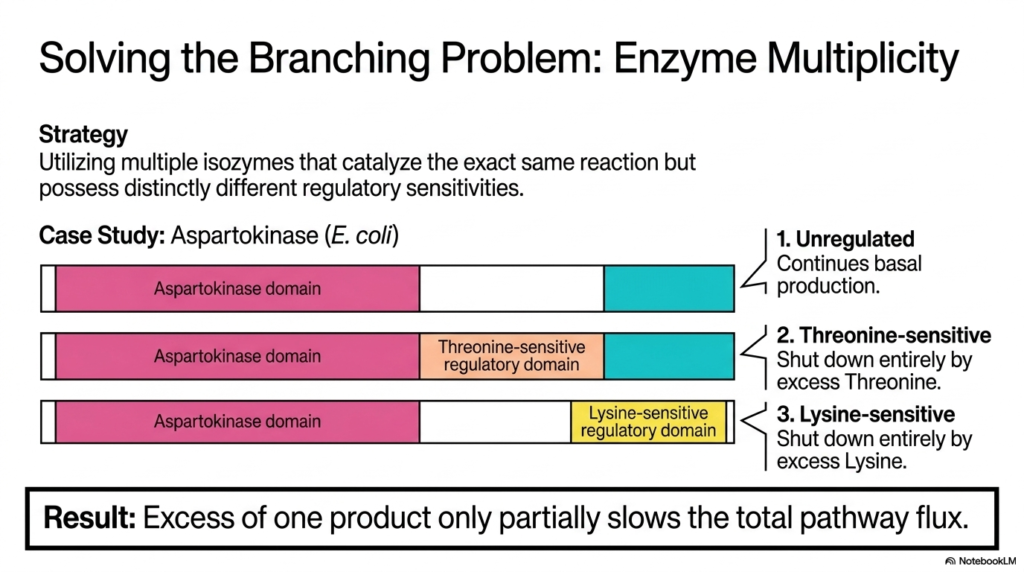
When a single metabolic branch point generates several distinct essential molecules, simple feedback inhibition is no longer sufficient. This slide introduces “Enzyme Multiplicity,” an advanced regulatory strategy for solving complex branching problems in amino acid biosynthesis. The primary strategy involves the cell using multiple isozymes—distinct proteins that catalyze the same chemical reaction but exhibit entirely different regulatory sensitivities. This concept is beautifully illustrated by examining the behavior of the enzyme Aspartokinase in E. coli, providing medical and biology students with a masterclass in molecular control and pathway optimization.
The visual breaks down the three distinct Aspartokinase isozymes operating simultaneously within the bacterial cell. The first isozyme is completely unregulated, ensuring a steady, basal level of continuous production regardless of cellular conditions. The second isozyme features a specific regulatory domain that is highly sensitive to threonine; it shuts down entirely only when threonine is in excess. The third isozyme contains a distinct domain sensitive strictly to lysine. This brilliant arrangement is absolutely crucial for the proper management of highly branched pathways in amino acid biosynthesis, allowing the cell to finely tune its production rates.
The ultimate result of this multiple-isozyme strategy is remarkable metabolic flexibility. Because each isozyme responds to a different downstream product, an excess of only one specific molecule—say, threonine—will only partially slow the total pathway flux. The lysine-sensitive and unregulated isozymes will continue to function normally, ensuring that the other necessary molecular branches do not suffer from starvation. By deploying distinct isozymes at major metabolic checkpoints, the networks of amino acid biosynthesis can gracefully handle complex cellular demands, preventing a localized product excess from accidentally shutting down the entire biochemical assembly line.
Slide 15: Cumulative Feedback in Amino Acid Biosynthesis
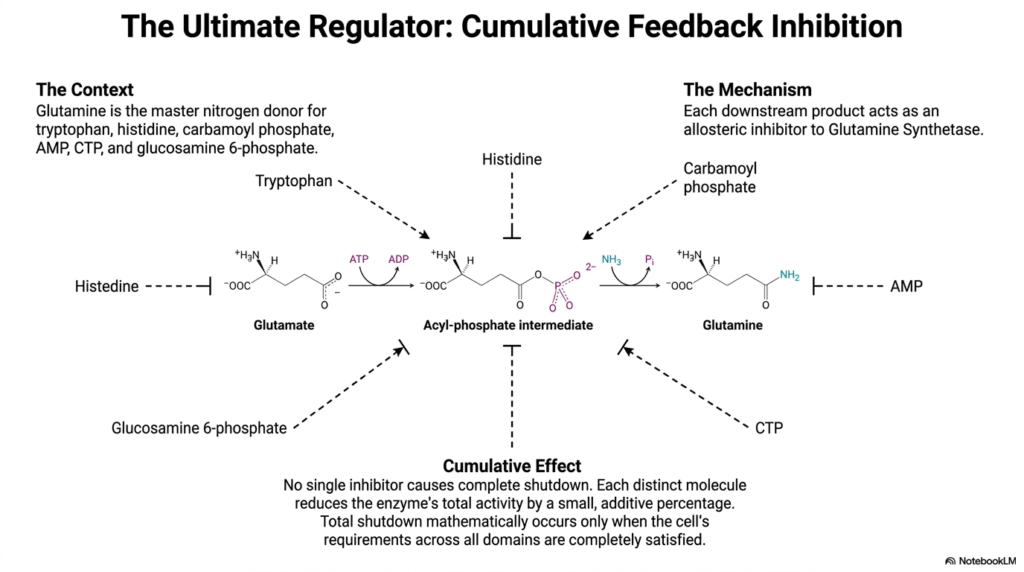
The incredible complexity of cellular metabolism demands a master regulatory switch to oversee the widespread distribution of critical resources. This final slide explores the concept of cumulative feedback inhibition, focusing on Glutamine Synthetase as the ultimate biochemical regulator. Because glutamine acts as the master nitrogen donor for a massive variety of downstream products—including tryptophan, histidine, AMP, and CTP—its production must be monitored with unparalleled precision. In the grand scheme of amino acid biosynthesis, Glutamine Synthetase represents one of the most highly regulated and mathematically elegant control points in the entire biological world.
The diagram illustrates the core mechanism of this advanced regulatory strategy. Rather than relying on a single off-switch, Glutamine Synthetase is subject to allosteric inhibition by multiple distinct downstream products. Molecules such as histidine, tryptophan, and AMP can each bind to the enzyme independently. However, binding of a single inhibitor does not cause a complete shutdown of the system. Instead, each individual downstream molecule reduces the enzyme’s total catalytic activity by a small, additive percentage. This nuanced mechanism ensures that the central hub of amino acid biosynthesis remains remarkably responsive to dynamic cellular conditions.
This cumulative effect provides the cell with an incredibly sophisticated degree of metabolic control. A total shutdown of Glutamine Synthetase mathematically occurs only when the cell’s requirements across all metabolic domains are simultaneously and completely satisfied. If only purines are in excess, the enzyme simply slows down slightly, continuing to provide necessary nitrogen for protein synthesis. This final example perfectly encapsulates the overarching theme of this presentation. The elaborate networks of amino acid biosynthesis are not just chemical reactions; they are highly evolved, mathematically precise biological computers that flawlessly sustain the complex chemistry of life.
Please read our Content Disclaimer Statement.
Check out our social media channels: