52. Biochemical Mechanisms of Transamination and Deamination
Have you ever wondered how the human body safely processes the protein from a heavy steak dinner? The answer lies in the elegant biochemical pathways of nitrogen metabolism. This introductory slide sets the stage for a deep dive into the molecular world of amino acid processing. The core purpose of this section is to introduce the presentation’s overarching theme: the continuous, vital dance of transamination and deamination in cellular survival.
Slide 1: Introduction to Transamination and Deamination: The Biochemical Core of Nitrogen Metabolism
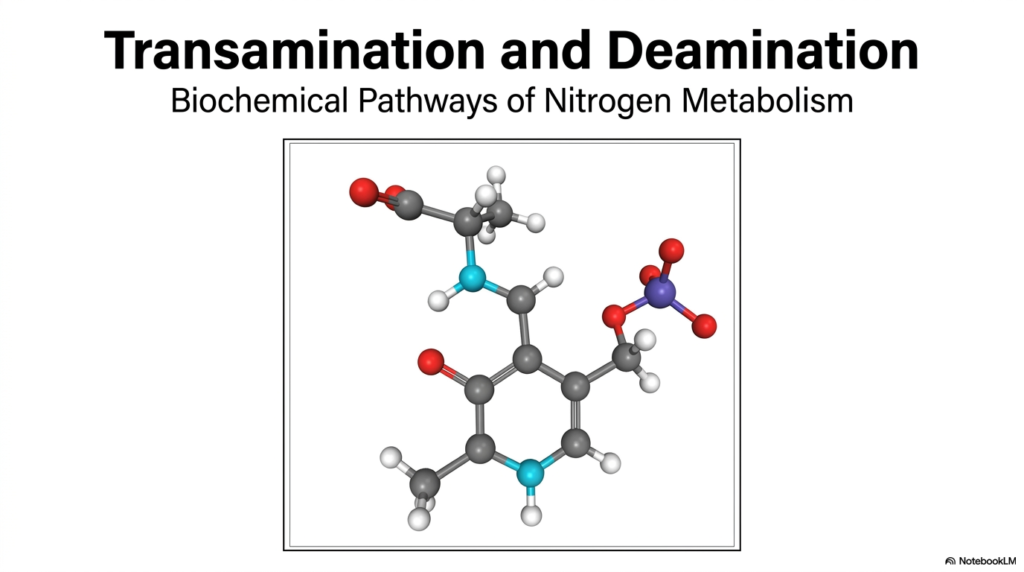
The visual presented highlights a complex molecular structure, representing the delicate chemical interplay required to handle biological nitrogen. In biological systems, managing nitrogen is a constant challenge because free ammonia is highly toxic to tissues, particularly the brain. This is exactly where the dual processes of transamination and deamination become absolute necessities for human health. They act as the body’s microscopic waste-management and recycling system, ensuring that amino groups are moved safely without causing damage.
As students of biochemistry, one must recognize that these pathways do not operate in isolation. The intricate mechanisms of transamination and deamination work synergistically to balance the body’s nitrogen pool. Proteins cannot be stored like fats or carbohydrates; their constituent amino acids must be constantly utilized, repurposed, or degraded. This metabolic necessity makes understanding the underlying chemical structures and enzymatic reactions absolutely critical for future medical professionals and scientists analyzing tissue health.
Throughout this slide deck, the fundamental principles of these reactions will be unpacked. The biochemical journey will begin with the collection of amino groups and end with their safe excretion. By mastering the concepts of transamination and deamination, students will unlock a deeper understanding of human metabolism, organ crosstalk, and the clinical diagnostics used every day in modern medicine to detect cellular damage.
Slide 2: The Biological Problem of Amino Nitrogen: Why Transamination and Deamination are Essential
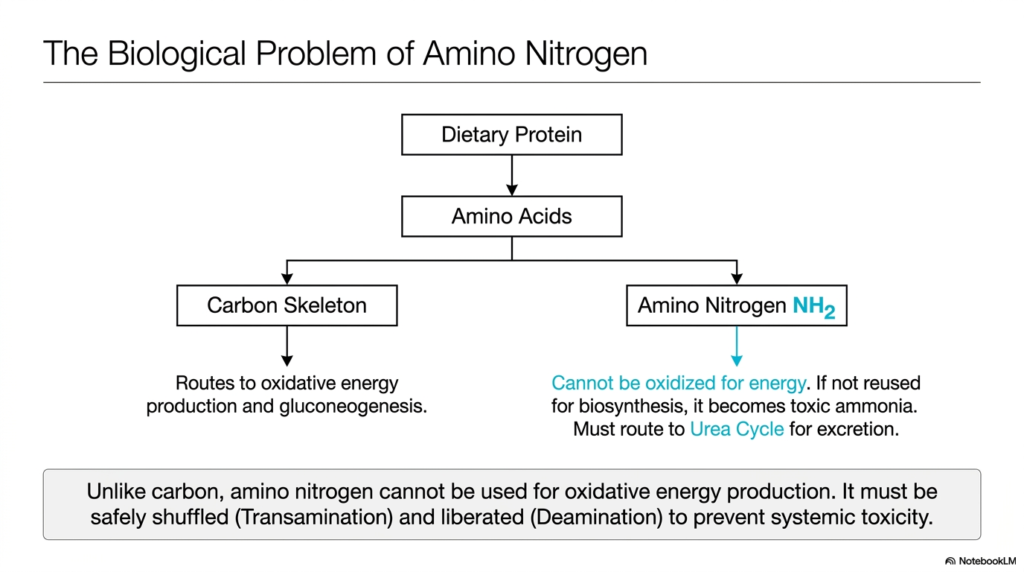
Imagine trying to burn a piece of wet wood for heat; it simply does not work efficiently and produces harmful smoke. Similarly, the human body cannot use dietary protein for energy without first stripping away its nitrogen component. This slide explains the fundamental biological problem of amino nitrogen. Its core purpose is to illustrate why the carbon skeleton of an amino acid can be used for energy, whereas the amino group requires specialized handling via transamination and deamination.
When humans consume dietary protein, it is broken down into constituent amino acids. These molecules are unique because they contain an amino group attached to a carbon skeleton. The carbon skeleton is incredibly versatile. Once separated, it can be routed either directly into oxidative energy production or into gluconeogenesis to maintain blood sugar levels. However, the amino nitrogen cannot be oxidized for energy. It presents a significant metabolic hurdle that cellular machinery must carefully manage to prevent systemic toxicity.
If the amino group is not immediately reused for the biosynthesis of new proteins or other nitrogenous compounds, it poses a severe threat. Left unchecked, it becomes free ammonia, a highly neurotoxic substance that can cause severe brain damage or death. To prevent this, the body relies heavily on the intertwined processes of transamination and deamination. These pathways safely shuttle the dangerous nitrogen away from vulnerable tissues and direct it toward the liver for detoxification.
Unlike carbon, which is exhaled easily as carbon dioxide, this toxic nitrogen waste must be chemically processed. The body first safely shuttles the amino group between molecules via transamination and deamination pathways. Ultimately, the nitrogen is liberated in a controlled environment and fed into the Urea Cycle for safe excretion in the urine. Understanding this overarching problem is the first step in mastering the clinical importance of hepatic function.
Slide 3: Transamination and Deamination Fundamentals: The Core Swap of Amino Groups
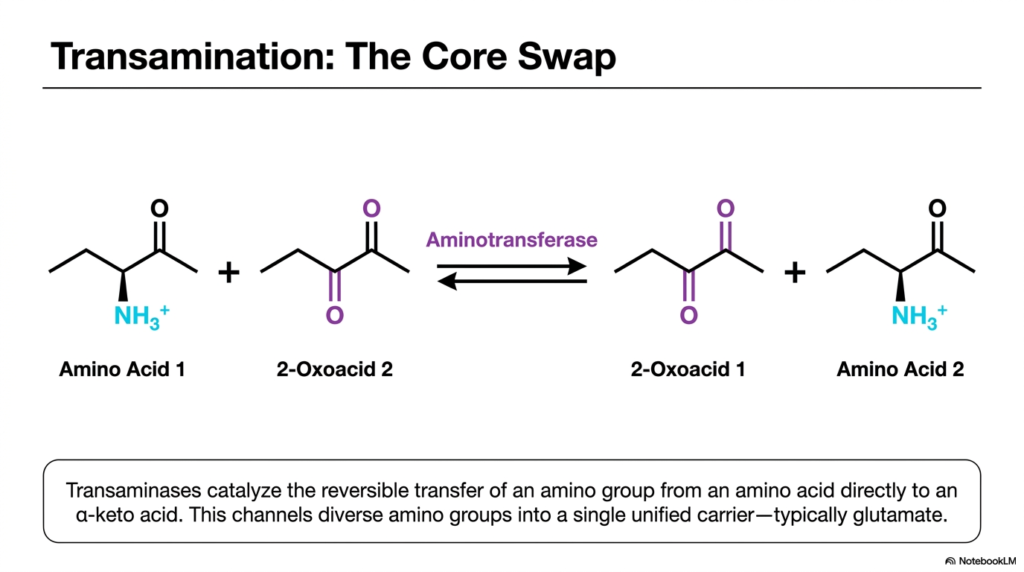
Think of cellular metabolism as a bustling train station where volatile cargo must be constantly transferred between trains without ever touching the ground. This is the essence of amino acid metabolism. This slide focuses on the initial transfer mechanism of this nitrogen cargo. Its core purpose is to break down the reversible transfer of an amino group from an amino acid to an alpha-keto acid, a process intimately linked with the broader cycles of transamination and deamination.
The reaction shown is a molecular swap. An aminotransferase enzyme catalyzes the transfer of an amino group from the first amino acid directly to a second 2-oxoacid. In doing so, the original amino acid becomes a new keto-acid, and the original keto-acid becomes a new amino acid. This elegant exchange is a cornerstone of the transamination and deamination continuum, ensuring that volatile nitrogen is never left floating freely in the cellular environment where it could cause immediate harm.
This transfer is completely reversible, which is a vital feature for cellular flexibility. Depending on the body’s metabolic state, the equilibrium can shift to either build new amino acids for protein synthesis or break them down for energy. This reversible funneling mechanism is what makes transamination and deamination so highly dynamic. By channeling diverse amino groups from various amino acids into a single unified carrier, the cell drastically simplifies its metabolic pathways.
Typically, this unified carrier is glutamate. By transferring various amino groups onto alpha-ketoglutarate to form glutamate, the liver centralizes its nitrogen processing. Glutamate then acts as the primary donor for the subsequent steps in the transamination and deamination cascade. This single-core swap prevents metabolic chaos, allowing the body to efficiently manage the nitrogen derived from the 20 standard amino acids found in the human diet.
Slide 4: Pyridoxal Phosphate (PLP): The Essential Cofactor for Transamination and Deamination
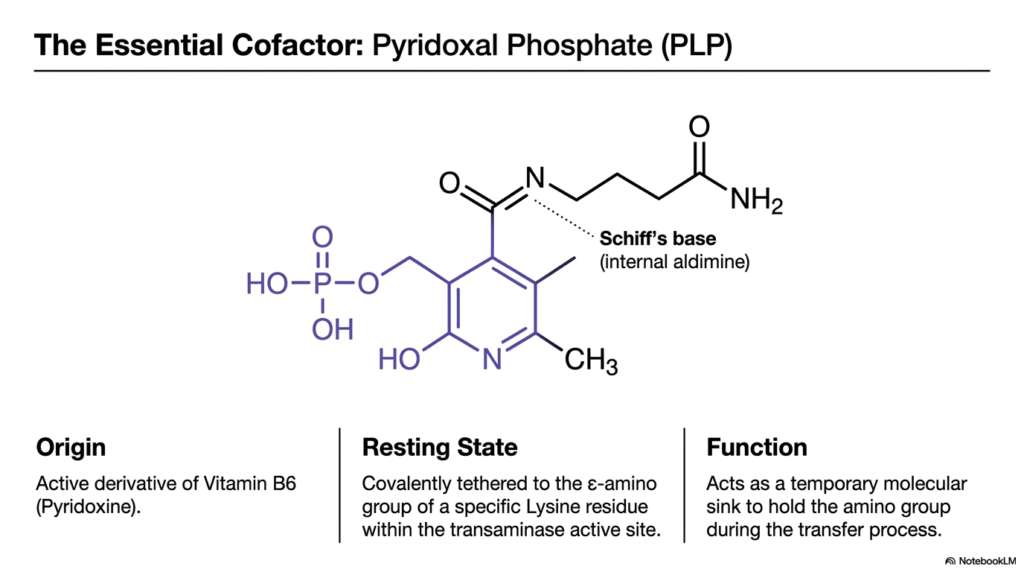
Every master builder requires a reliable tool belt to practice their craft safely and precisely. In the biochemical world of nitrogen metabolism, that essential tool is a remarkable molecule called Pyridoxal Phosphate. This slide examines the vital cofactor required by aminotransferase enzymes. Its core purpose is to illustrate the structure and function of PLP, the active derivative of Vitamin B6, which is absolutely mandatory for the initial steps of transamination and deamination.
Pyridoxal Phosphate, commonly abbreviated as PLP, originates directly from dietary Vitamin B6. Without adequate levels of this vitamin, the entire network of transamination and deamination would grind to a dangerous halt. In its resting state, the PLP molecule is covalently tethered to a specific Lysine residue within the active site of the transaminase enzyme. This connection forms an internal aldimine, priming the enzyme for the arrival of an amino acid substrate.
The primary function of PLP is to act as a temporary molecular sink. When an amino acid enters the enzyme’s active site, the amino group must be held securely while the carbon skeleton is released. PLP safely holds onto this reactive amino group during the transfer process. This temporary storage mechanism is the defining chemical characteristic that allows transamination and deamination pathways to proceed without releasing toxic free ammonia into the surrounding cellular environment.
The structural features of PLP, particularly its electron-withdrawing pyridine ring, make it uniquely suited for this task. It acts as an electron sink, stabilizing the reactive intermediates formed during amino group transfer. Understanding the molecular mechanism of this cofactor is crucial for students, as it clearly demonstrates how transamination and deamination rely on precise structural chemistry to carry out large-scale physiological functions safely.
Slide 5: Ping-Pong Bi Bi Kinetics: The Molecular Shuffle in Transamination and Deamination
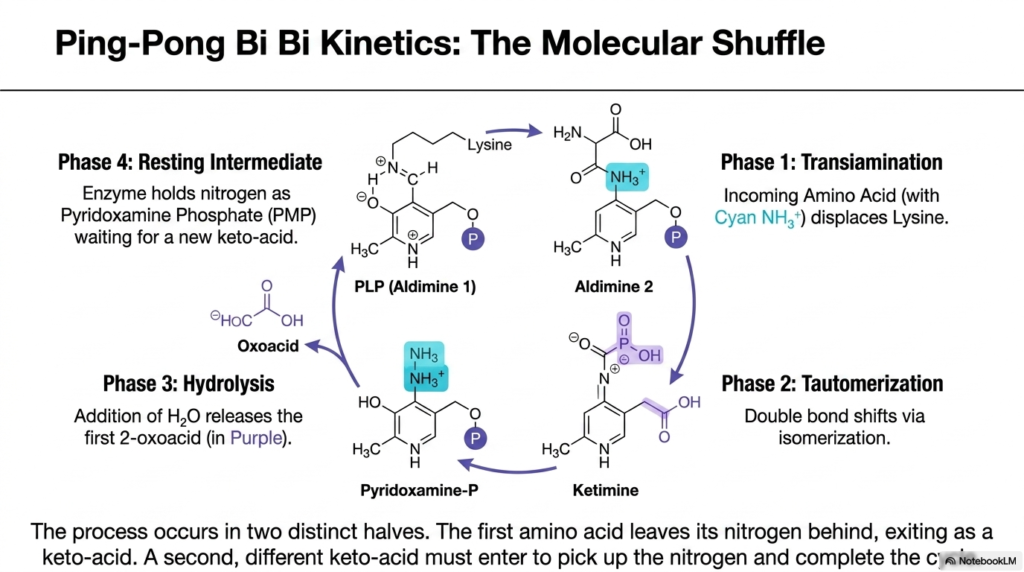
Imagine a precise relay race where the baton must be handed to a stationary official before a new runner can pick it up and continue the race. This perfectly describes the enzymatic mechanism of aminotransferases. This slide delves into the fascinating Ping-Pong Bi Bi kinetics of the transferase enzymes. Its core purpose is to break down the four distinct phases of this molecular shuffle, a process that underpins the entire system of transamination and deamination.
The kinetic mechanism occurs in two distinct halves, perfectly illustrating how transamination and deamination maintain nitrogen safety. In the first phase, the incoming amino acid enters the active site and displaces the lysine residue, binding its amino group to the PLP cofactor to form Aldimine 2. Next, a critical tautomerization occurs, in which a double bond shifts via isomerization to form a ketimine intermediate. This prepares the molecule for the chemical separation of the carbon backbone.
The third phase involves hydrolysis. The addition of a water molecule cleaves the structure, releasing a newly formed 2-oxoacid. However, the nitrogen is left behind, securely attached to the cofactor, which is now called Pyridoxamine Phosphate. This is the resting intermediate stage. The enzyme is patiently holding the nitrogen, waiting for a new, different keto-acid to enter the active site and complete the second half of the transamination and deamination cycle.
To finish the shuffle, a new keto-acid must enter, and the exact reverse of these steps occurs. Pyridoxamine Phosphate transfers the amino group it holds onto the new keto acid, creating a new amino acid and regenerating the original PLP cofactor. This ping-pong mechanism ensures that free ammonia is never produced during this phase of transamination and deamination, showcasing the beautiful, protective complexity of enzyme kinetics in human biochemistry.
Slide 6: Liberating Toxic Ammonia: The Deamination Side of Transamination and Deamination

If transamination is the safe transportation of a dangerous cargo, deamination is the careful unloading of that cargo at a specialized waste facility. This slide marks a critical transition in the metabolic pathway. Its core purpose is to distinguish the safe shuffling of nitrogen from the process that ultimately strips it from the carbon backbone. Here, we explore how the transamination-deamination continuum ultimately liberates toxic ammonia for obligatory excretion.
While transamination safely moves nitrogen between carbon backbones without releasing it, deamination is required to finally detach it as free ammonia. This slide outlines three distinct types of this liberating process. The first is Oxidative Deamination. This reaction, driven by dehydrogenases or oxidases, generates alpha-keto acids and free ammonia. This is the most critical terminal step for urea synthesis, acting as the primary off-ramp for the nitrogen funneled through transamination and deamination networks.
The second mechanism is Hydrolytic Deamination. Unlike the oxidative route, this process directly adds a water molecule to release the ammonia without a simultaneous oxidation reaction. A classic clinical example is the enzyme Glutaminase, which converts Glutamine directly into Glutamate, releasing a free ammonia molecule. Both oxidative and hydrolytic pathways are essential components of the broader transamination and deamination system, ensuring that waste nitrogen from all tissues can be processed.
Finally, the slide introduces Monoamine Oxidation. This involves the oxidation of monoamines and is absolutely critical for the central nervous system. It inactivates powerful neurotransmitters such as serotonin and epinephrine via specialized enzymes. While specific, this is a vital part of the body’s overall transamination and deamination strategy. It highlights that removing amino groups is not just about digesting food; it is fundamental to regulating brain chemistry and neurological function.
Slide 7: Glutamate Dehydrogenase: The Bottleneck of Transamination and Deamination
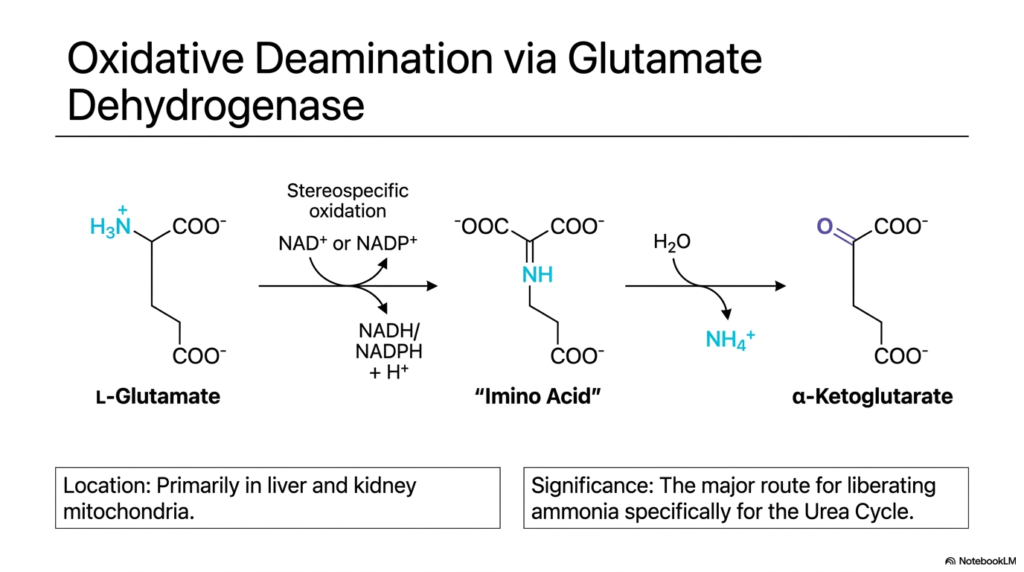
Imagine a massive river system where countless small tributaries eventually converge into a single, roaring waterfall. In the biochemistry of nitrogen, Glutamate Dehydrogenase acts as that powerful waterfall. This slide explores the specific mechanism of oxidative deamination via this crucial enzyme. Its core purpose is to explain how L-Glutamate is processed to release ammonia, acting as the central bottleneck in the overarching pathways of transamination and deamination.
The reaction begins with L-Glutamate, the universal collection point for cellular nitrogen. Through stereospecific oxidation, Glutamate Dehydrogenase converts L-Glutamate into an imino acid intermediate, producing NADH or NADPH and a hydrogen ion. Following this, the addition of a water molecule hydrolyzes the imino group. This precisely orchestrated transamination and deamination sequence finally yields alpha-Ketoglutarate and a free, unbound ammonium ion.
The physiological location of this reaction is incredibly vital. Glutamate Dehydrogenase operates primarily within the mitochondria of the liver and kidneys. By sequestering this reaction inside the mitochondria, the cell protects its delicate cytoplasm from the highly toxic free ammonia that is generated. This compartmentalization is a hallmark of safe transamination and deamination practices, ensuring the dangerous byproduct is immediately adjacent to the enzymes of the Urea Cycle for instant detoxification.
The biochemical significance of this specific reaction cannot be overstated. It represents the major route for liberating ammonia specifically to feed the Urea Cycle. Because almost all amino acids first transfer their nitrogen to form glutamate, this single enzyme controls the flow of nitrogen waste out of the body. Understanding Glutamate Dehydrogenase is essential to understanding how the dual engines of transamination and deamination effectively protect the human body from fatal ammonia toxicity.
Slide 8: Hepatic Integration: The Nitrogen Funnel of Transamination and Deamination
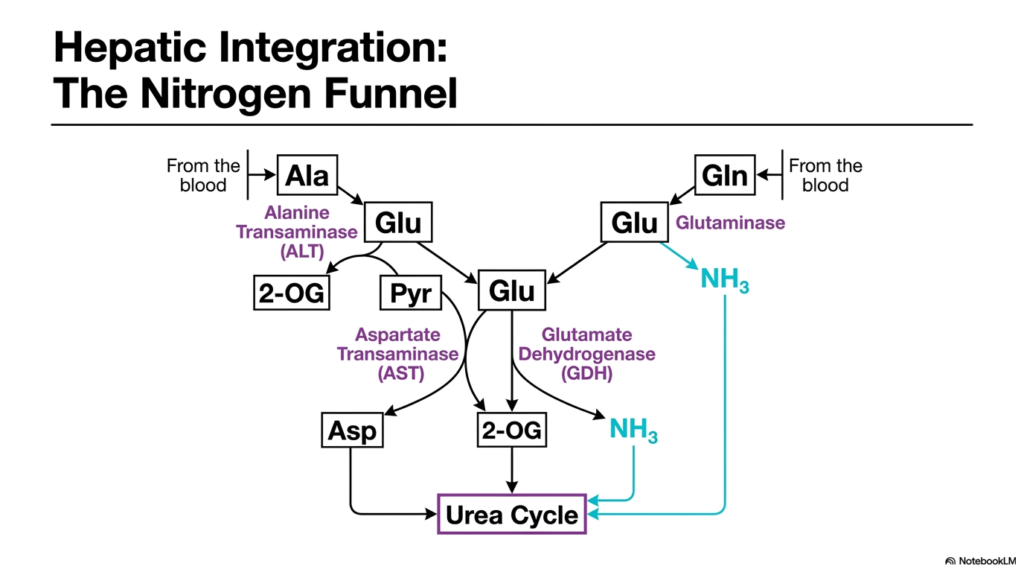
Managing nitrogen waste requires a central processing plant with flawless logistics, and the human liver is perfectly designed for this colossal task. This slide maps out the nitrogen funnel within hepatic tissues. Its core purpose is to demonstrate how multiple metabolic pathways converge on a single organ, highlighting the seamless integration of transamination and deamination to feed the Urea Cycle and prevent systemic ammonia poisoning.
The diagram illustrates nitrogen arriving at the liver from the bloodstream, primarily carried by two non-toxic transport molecules: Alanine and Glutamine. Once inside the liver, the transamination and deamination machinery takes over. Alanine is processed by Alanine Transaminase, which transfers its amino group to Glutamate, forming Pyruvate. This represents the vital initial collection phase of hepatic nitrogen metabolism, gathering waste from peripheral tissues.
Simultaneously, Glutamine from the blood is acted upon by Glutaminase, releasing one molecule of free ammonia and forming additional Glutamate. With the cellular nitrogen concentrated into the Glutamate pool, the system utilizes Aspartate Transaminase and Glutamate Dehydrogenase. Aspartate Transaminase creates Aspartate, while Glutamate Dehydrogenase liberates more free ammonia. This perfectly illustrates how transamination and deamination provide the exact chemical substrates required to drive the Urea Cycle.
This integrated funnel ensures that no matter where the nitrogen originated in the body, it is safely processed. The liver’s ability to utilize both transamination and deamination simultaneously allows it to adapt to varying nutritional loads and metabolic stressors. For medical students, understanding this chart is crucial, as it visually links peripheral tissue protein breakdown to the liver’s ultimate detoxification pathways, underscoring the elegant biochemical design of human metabolism.
Slide 9: The Glucose-Alanine Cycle: Inter-Organ Transamination and Deamination
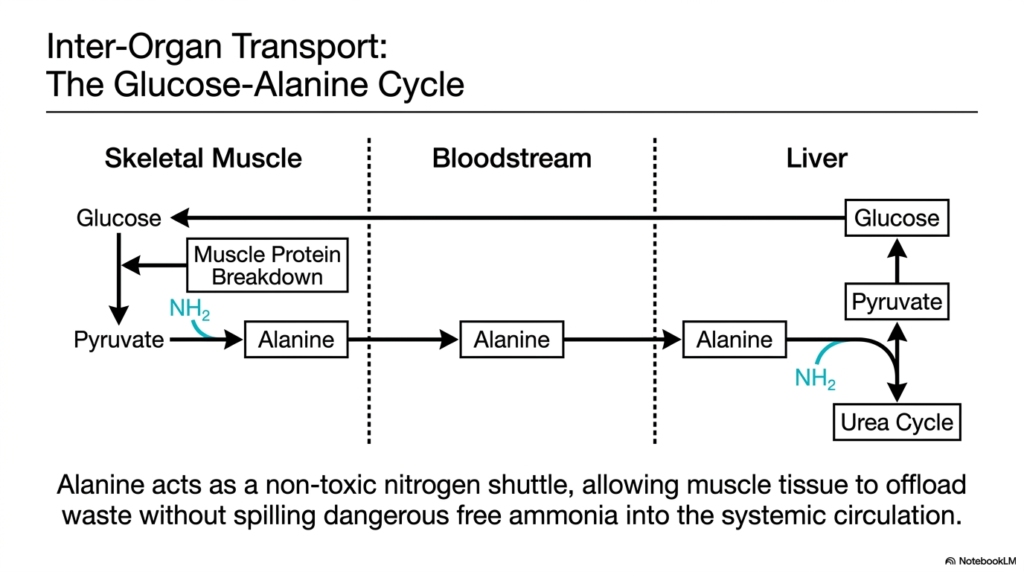
When you lift heavy weights or sprint, your muscles break down protein for energy, generating toxic nitrogen waste right in the middle of an intense workout. How does the muscle survive this sudden biochemical stress? This slide introduces the Glucose-Alanine Cycle. Its core purpose is to explain the brilliant inter-organ transport system that relies on transamination and deamination to shuttle waste from skeletal muscle to the liver without poisoning the blood.
Skeletal muscle lacks the enzymes to perform the Urea Cycle, meaning it cannot detoxify the ammonia generated from muscle protein breakdown locally. Instead, it relies on a clever transport mechanism. Inside the muscle, the waste amino nitrogen is transferred onto pyruvate to form the amino acid Alanine. This is a prime example of how transamination and deamination pathways are strategically utilized across different organ systems to support overall survival.
Alanine acts as a remarkably safe, non-toxic nitrogen shuttle. It seamlessly exits the skeletal muscle and travels through the systemic bloodstream to the liver. Once it arrives in the hepatic tissue, the liver reverses the entire process. Through hepatic transamination and deamination, the amino group is carefully removed from Alanine, regenerating Pyruvate and liberating the nitrogen directly into the Urea Cycle for safe excretion.
The brilliance of this cycle lies in its ability to recycle. The remaining Pyruvate in the liver is not wasted; it is used to synthesize new Glucose via gluconeogenesis. This new Glucose is then pumped back into the bloodstream to feed the working skeletal muscle. By linking energy production to nitrogen disposal, the cycle demonstrates the remarkable physiological efficiency of the body’s expansive transamination and deamination network.
Slide 10: Clinical Enzymology: Serum Transaminases in Transamination and Deamination
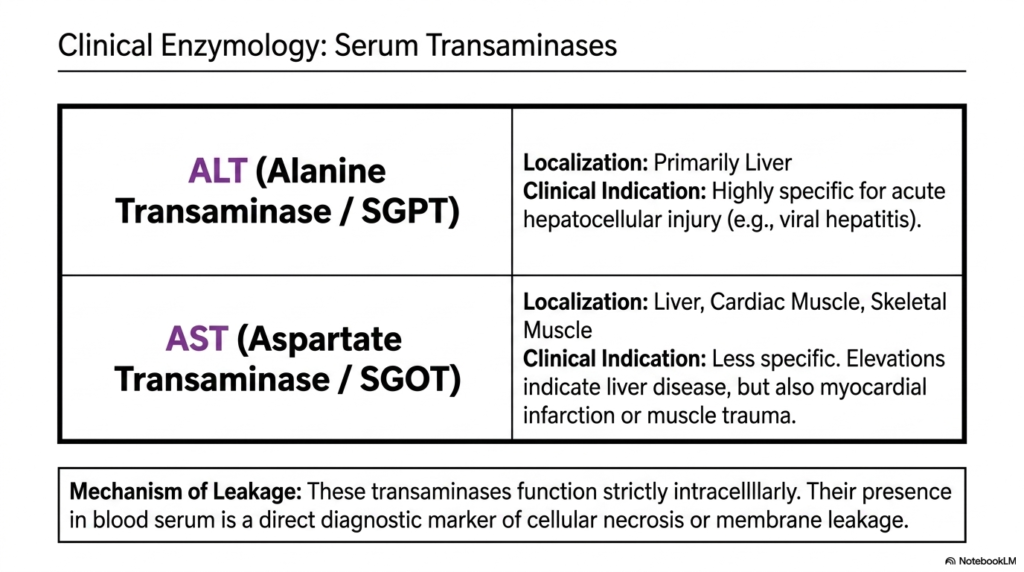
In modern medicine, biochemistry is not just theoretical; it is actively used to diagnose and save lives every single day in the clinic. This slide bridges the gap between basic science and clinical practice by examining serum transaminases. Its core purpose is to explain how the enzymes responsible for transamination and deamination serve as critical diagnostic biomarkers for detecting severe organ damage and cellular necrosis in patients.
The slide highlights two major enzymes: Alanine Transaminase and Aspartate Transaminase. Alanine Transaminase is localized primarily within the liver. Because of this restricted location, an elevation of this enzyme in a patient’s blood is a highly specific clinical indicator for acute hepatocellular injury, such as viral hepatitis. It is a direct measure of the structural integrity of the transamination and deamination machinery in the liver.
Aspartate Transaminase, on the other hand, is found in the liver, cardiac muscle, and skeletal muscle, making it less specific to hepatic tissue. High levels can indicate liver disease, but they can also signal a recent myocardial infarction or severe muscle trauma. Understanding the tissue distribution of these transamination and deamination enzymes allows physicians to pinpoint which tissues are under acute stress or actively dying.
The underlying mechanism of this diagnostic tool relies entirely on membrane leakage. Under normal, healthy conditions, these transaminases function strictly intracellularly, performing their metabolic duties. Their sudden presence in the blood serum means that cell membranes have physically ruptured, spilling their contents into the circulation. Thus, tracking transamination and deamination enzymes is a direct, reliable method for monitoring the progress of cellular necrosis.
Slide 11: Spontaneous DNA Damage: The Dark Side of Transamination and Deamination
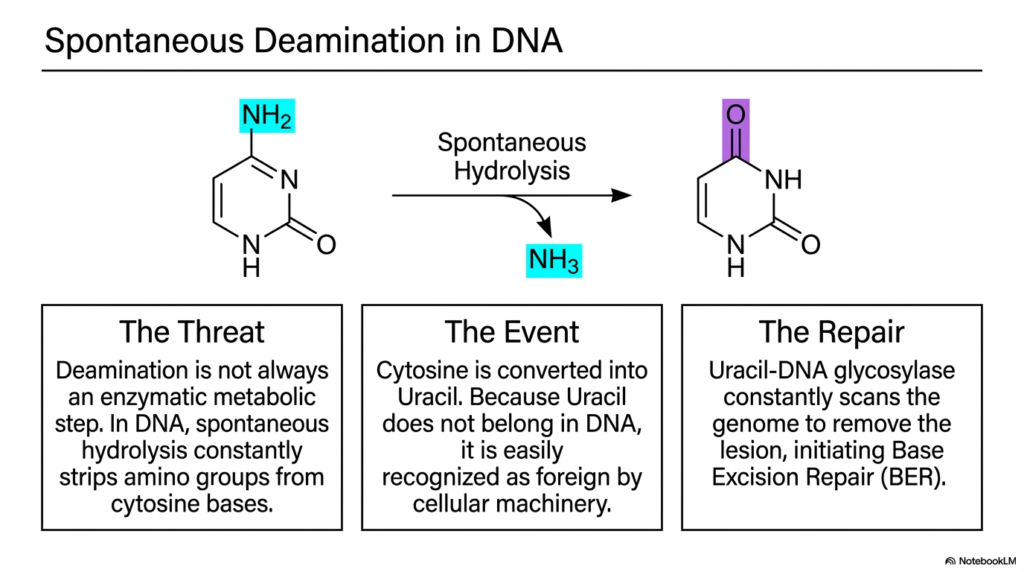
While we usually think of metabolism as a carefully controlled enzymatic process, biology is constantly fighting the chaotic, degrading forces of thermodynamics. Sometimes, vital chemical groups simply fall off vital molecules unprompted. This slide explores the terrifying phenomenon of spontaneous DNA damage. Its core purpose is to illustrate that the chemistry of transamination and deamination isn’t always helpful; sometimes it poses a massive threat to the integrity of our genetic code.
Deamination is not always a beneficial, enzyme-driven metabolic step designed for bodily nitrogen disposal. Within the watery environment of the cell nucleus, spontaneous hydrolysis constantly attacks our DNA. Specifically, it strips amino groups directly from Cytosine bases. This represents the dark side of transamination and deamination chemistry. When this spontaneous hydrolysis occurs, Cytosine is forcibly converted into Uracil, a base that belongs strictly in RNA, not DNA.
Because Uracil absolutely does not belong in the DNA double helix, its sudden appearance constitutes a significant mutational event. However, this specific chemical alteration is actually a biological blessing in disguise. Cellular machinery readily recognizes Uracil as a foreign base in DNA. This recognition is critical because it immediately triggers the body’s powerful repair mechanisms to fix the damage caused by this rogue transamination and deamination event.
The repair process is spearheaded by an enzyme called Uracil-DNA glycosylase. This enzyme constantly patrols the entire human genome, locating and removing these specific lesions. Once the Uracil is removed, the Base Excision Repair pathway is initiated to insert the correct Cytosine back into place. Understanding this constant microscopic battle highlights how the chemical principles of transamination and deamination dictate both metabolic health and genetic stability.
Slide 12: The Mutagenic Threat of 5-Methylcytosine: Evading Transamination and Deamination Repair
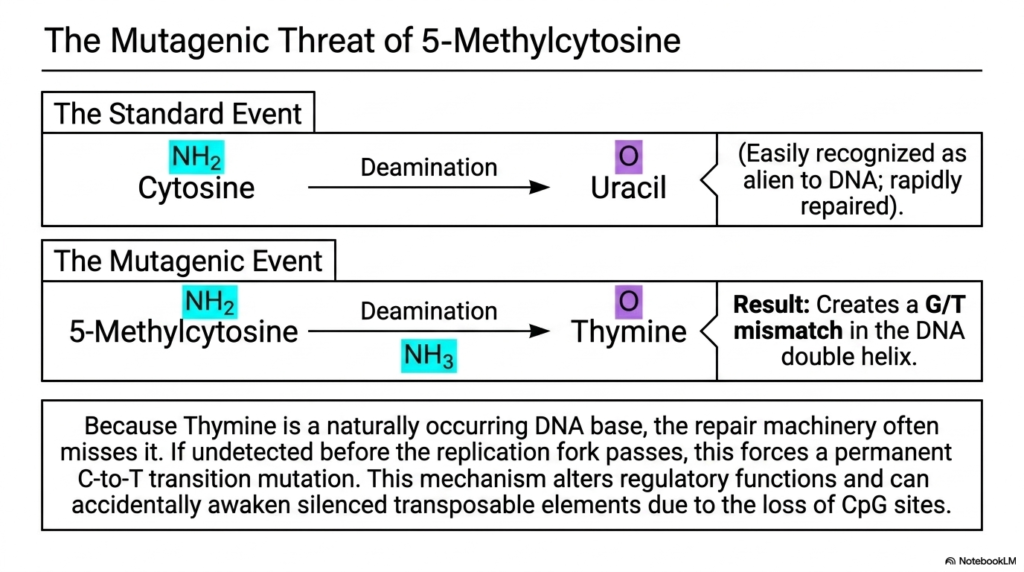
What happens when our cellular DNA security system is tricked by a cleverly disguised molecular intruder? The result is often a permanent, irreversible genetic mutation that can lead to disease. Building on the previous slide’s concepts, this slide delves deeper into a highly dangerous form of DNA damage. Its core purpose is to explain the severe mutagenic threat posed by the deamination of 5-Methylcytosine, an event that frequently outsmarts normal transamination and deamination repair networks.
The slide contrasts a standard mutational event with a highly dangerous one. When a standard Cytosine deaminates to Uracil, it is easily recognized and repaired by the cell. However, human DNA often contains 5-methylcytosine, a modified base involved in epigenetics. When the chemical forces of transamination and deamination act on 5-Methylcytosine, it does not convert to Uracil. Instead, it deaminates directly into normal Thymine, creating a massive problem for repair machinery.
Because Thymine is a naturally occurring, expected DNA base, the repair machinery often completely misses the error because it doesn’t look alien. This creates a dangerous mismatch in the DNA double helix. If this mismatch is undetected before the cellular replication fork passes during cell division, the error becomes permanently locked in. This rogue transamination and deamination event forces a permanent transition mutation into the genome of the daughter cell.
The consequences of this specific mutation are medically profound. This mechanism disrupts vital regulatory functions and can inadvertently awaken silenced transposable elements by removing key structural sites. Many human genetic diseases and aggressive cancers can be traced back to this exact chemical vulnerability. It serves as a stark reminder that the chemical rules governing transamination and deamination apply universally across all biology.
Slide 13: Purine Deamination and Pathology: Transamination and Deamination in Immune Defense
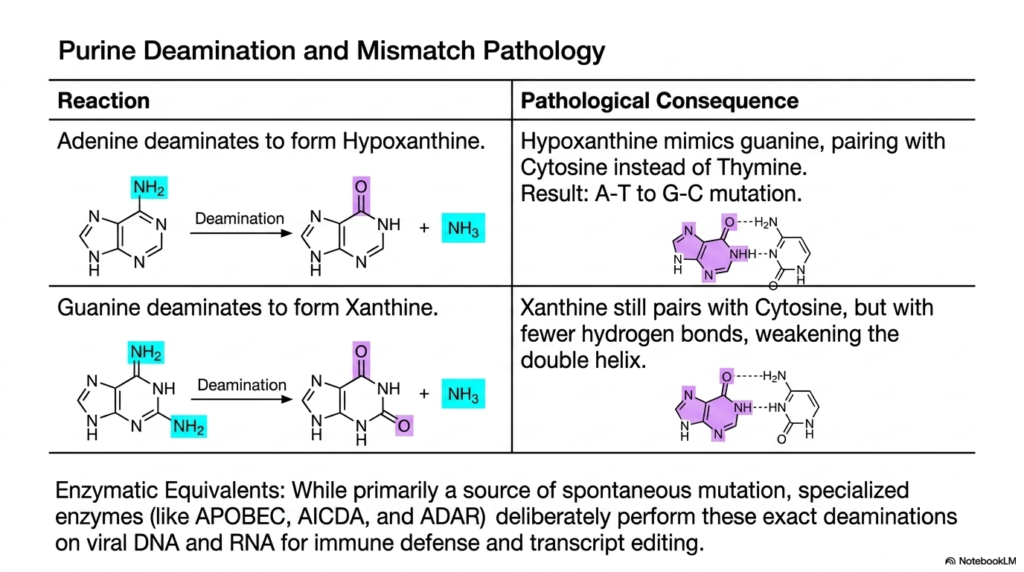
The structural integrity of the DNA double helix relies entirely on precise hydrogen bonding between paired bases. When that molecular bonding is compromised, the genetic code destabilizes rapidly. This slide examines the pathological consequences of purine modification. Its core purpose is to detail how the accidental removal of amino groups from Adenine and Guanine weakens DNA, while also revealing how the immune system weaponizes these transamination and deamination mechanisms for defense.
The slide details the specific degradation reactions for the purine bases. When Adenine undergoes spontaneous deamination, it forms Hypoxanthine. This is highly pathological because Hypoxanthine chemically mimics Guanine, causing it to pair with Cytosine instead of Thymine during DNA replication. This results in a permanent mutation. This demonstrates how uncontrolled transamination and deamination reactions can quietly rewrite the genetic code, leading to cellular dysfunction over a patient’s lifetime.
Similarly, when Guanine deaminates, it forms Xanthine. While Xanthine still pairs with Cytosine, it forms fewer hydrogen bonds. This significantly weakens the structural integrity of the double helix, making it far more prone to physical breakage and replication errors. These spontaneous errors reflect the biological cost of living in an aqueous environment, where the chemical reactions of transamination and deamination can occur spontaneously at any moment.
However, this chemistry is not always accidental or harmful. The slide highlights specialized enzymatic equivalents that act intentionally. These cellular enzymes deliberately perform these exact deaminations on viral DNA and RNA as a powerful form of immune defense and transcript editing. By intentionally weaponizing transamination and deamination, the human body can actively mutate invading viruses to death, showcasing the incredible duality of these biochemical mechanisms in combating disease.
Slide 14: Synthesis and Comparison: Transamination and Deamination Dimensions
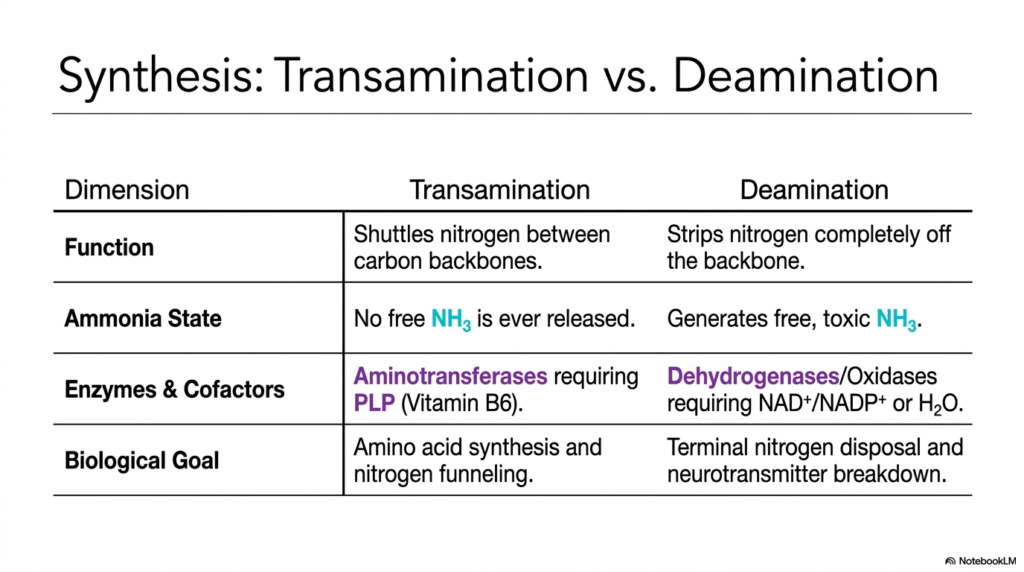
To truly master complex biochemical pathways, one must be able to synthesize information and contrast the subtle differences between intimately interconnected systems. This slide provides the essential high-level academic summary. Its core purpose is to present a definitive comparison between the two major pathways we have explored, using a clear dimension table to clarify the distinct roles of transamination and deamination in maintaining human nitrogen homeostasis.
The table breaks down the processes across four vital dimensions. First, we look at biological function. Transamination’s role is to safely shuttle nitrogen between different carbon backbones, serving as a molecular transport service. Conversely, deamination’s function is to strip the nitrogen completely from the backbone. This clear distinction helps students understand that transamination and deamination are two halves of a whole; one organizes the waste, and the other prepares it for disposal.
The ‘Ammonia State’ dimension is critical for toxicological understanding. During transamination, no free, toxic ammonia is ever released; the nitrogen is always securely bound to a molecule. Deamination, however, is the dangerous terminal step that generates this free, toxic ammonia. This highlights exactly why the transamination and deamination enzymes require vastly different cofactors to perform their specific chemical duties, utilizing Vitamin B6 versus specialized oxidation molecules.
Finally, the ‘Biological Goal’ dimension unites the overarching concepts. Transamination aims to synthesize necessary amino acids and funnel waste nitrogen to the liver for processing. Deamination represents the terminal disposal of nitrogen and the necessary breakdown of neurotransmitters. By reviewing this synthesized data, medical students can solidify their understanding of how transamination and deamination work in tandem to balance anabolic growth with catabolic waste management in humans.
Slide 15: The Grand Integration of Pathways: Mastering Transamination and Deamination
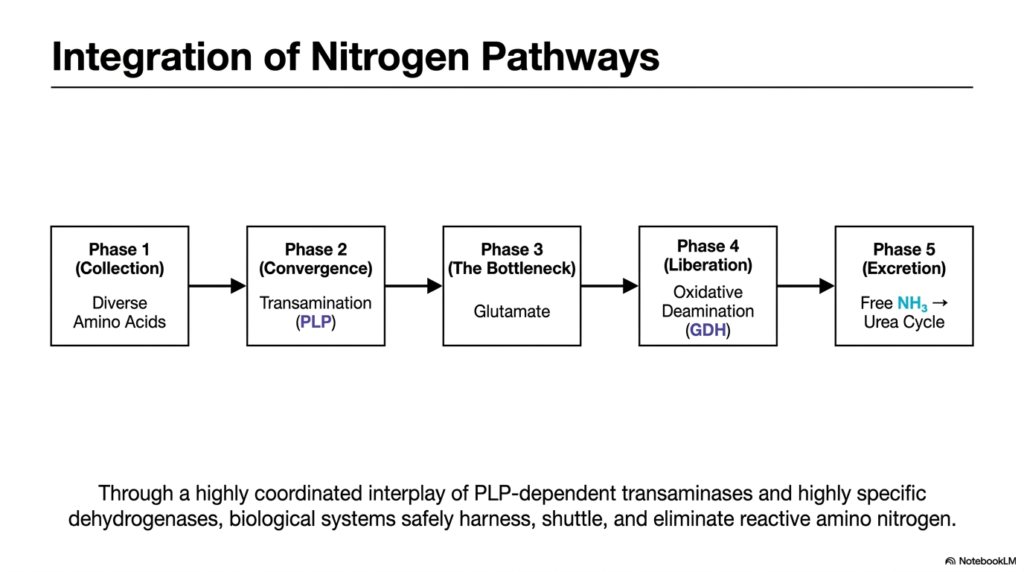
Every complex symphony relies on the precise, timed coordination of its individual instruments to create a masterpiece. The human metabolism operates exactly the same way. This final slide brings the entire biochemical journey together into a single, cohesive flowchart. Its core purpose is to illustrate the grand integration of nitrogen pathways, showing the step-by-step phases by which transamination and deamination sequentially process diverse amino acids into the disposable urea.
The model logically divides the entire process into five phases. Phase 1 is the collection of diverse amino acids from our diet and tissue breakdown. Phase 2 represents convergence, in which transamination channels these varied amino groups into unified carriers. This is the beauty of the transamination and deamination network: it takes chaotic, diverse inputs and forces them into a highly standardized, manageable chemical format for the liver to process safely.
Phase 3 is the bottleneck, centered heavily around the molecule Glutamate, which acts as the temporary storage reservoir for all collected bodily nitrogen. Phase 4 is liberation, in which oxidative deamination finally rips the nitrogen away, forming free ammonia. These middle phases perfectly demonstrate the metabolic hand-off between transamination and deamination, proving they are inextricably linked in the quest to protect the body from fatal neurotoxicity.
Finally, Phase 5 is excretion, where the freshly liberated free ammonia is immediately captured by the Urea Cycle and safely removed from the body as waste. Through this highly coordinated interplay of specific enzymes and cofactors, biological systems safely harness, shuttle, and eliminate reactive amino nitrogen. Mastering this flowchart means fully understanding the indispensable, life-sustaining power of human transamination and deamination pathways.
Please read our Content Disclaimer Statement.
Check out our social media channels: