35. Energetic Coupling: How ATP Powers Cellular Metabolism
Imagine trying to run a smartphone without a battery. The device simply cannot function. Similarly, biological cells cannot perform the complex, demanding tasks required to sustain life without a constant, reliable supply of usable energy. This comprehensive slide deck explores the fundamental principles of cellular bioenergetics. It carefully breaks down exactly how living cells capture, store, and utilize energy to drive essential biological processes forward. By deeply understanding these mechanisms, medical students will firmly grasp the core metabolic pathways powering life.
Slide 1: Cellular Bioenergetics and Energetic Coupling Fundamentals
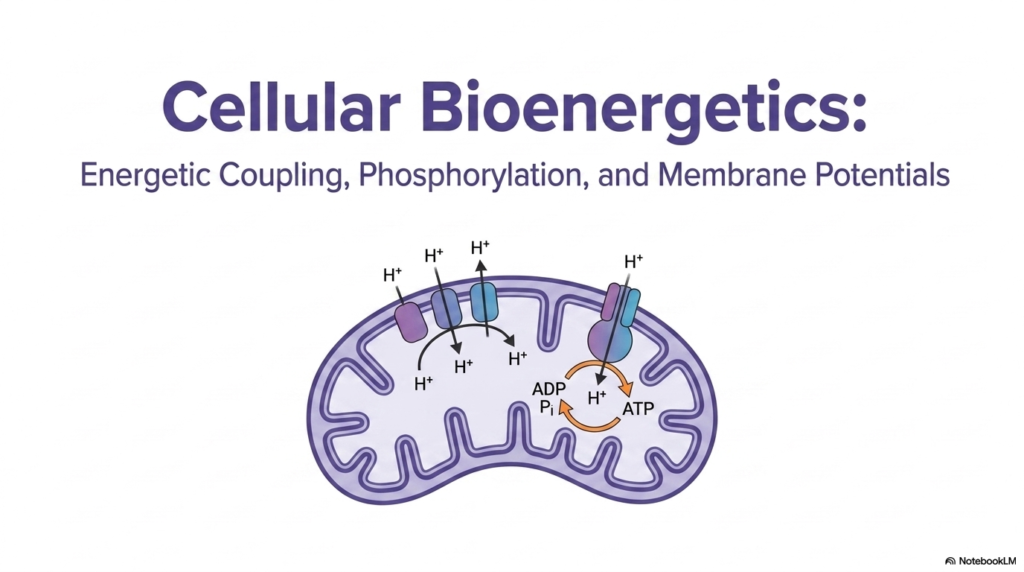
The first slide introduces the grand landscape of cellular bioenergetics, establishing the foundational mechanisms that keep biological cells alive. At the center of this intricate system is the mitochondrion, universally known as the powerhouse of the cell. This distinct organelle is where cellular respiration takes place, transforming raw nutrients into a highly usable biological energy currency. A fundamentally critical concept introduced here is energetic coupling, which serves as the indispensable bridge between energy-releasing and energy-consuming processes. Without this vital link, cellular life would instantly grind to a halt, as biochemical reactions would simply dissipate useless heat instead of performing valuable cellular work.
In the provided illustration, one can clearly observe protons being actively pumped entirely across the inner mitochondrial membrane. This highly precise, active movement creates a massively significant electrochemical gradient, storing vast amounts of potential energy much like heavy water held behind a massive concrete dam. The cell expertly harnesses this stored potential energy to reliably synthesize adenosine triphosphate, universally known as ATP. The remarkable process of energetic coupling strictly ensures that the raw energy released from the dissipating proton gradient is directly utilized to aggressively attach a loose phosphate group to ADP, forming the highly energetic ATP molecule.
ATP consistently functions as the universal energy currency of biological systems, perfectly primed to be spent on demanding cellular activities. The visual illustration elegantly shows how ATP synthase, a massive and complex molecular machine, skillfully facilitates this crucial biological transformation. As numerous protons naturally flow backward into the dense mitochondrial matrix, their rapid movement physically rotates specific structural components of the enzyme. This raw mechanical energy is seamlessly transformed directly into usable chemical energy. The true essence of energetic coupling is perfectly demonstrated here: the spontaneous flow of massive proton waves directly drives the completely non-spontaneous synthesis of ATP molecules.
Understanding this fundamental cellular overview is absolutely crucial for advanced biochemistry and medical students. The overarching thermodynamic principles are not just abstract textbook ideas; they represent the highly tangible realities closely governing every single metabolic pathway operating within the complex human body. By thoroughly mastering exactly how simple phosphorylation and complex membrane potentials intimately interact to sustain ongoing life, the dedicated learner successfully gains a profound appreciation for the immense efficiency of biological design. This comprehensive overview perfectly sets the necessary stage for a much deeper dive into the specific, rapid molecular transactions detailed in the upcoming slides.
Slide 2: The Bifurcation of Biological Energy and Energetic Coupling
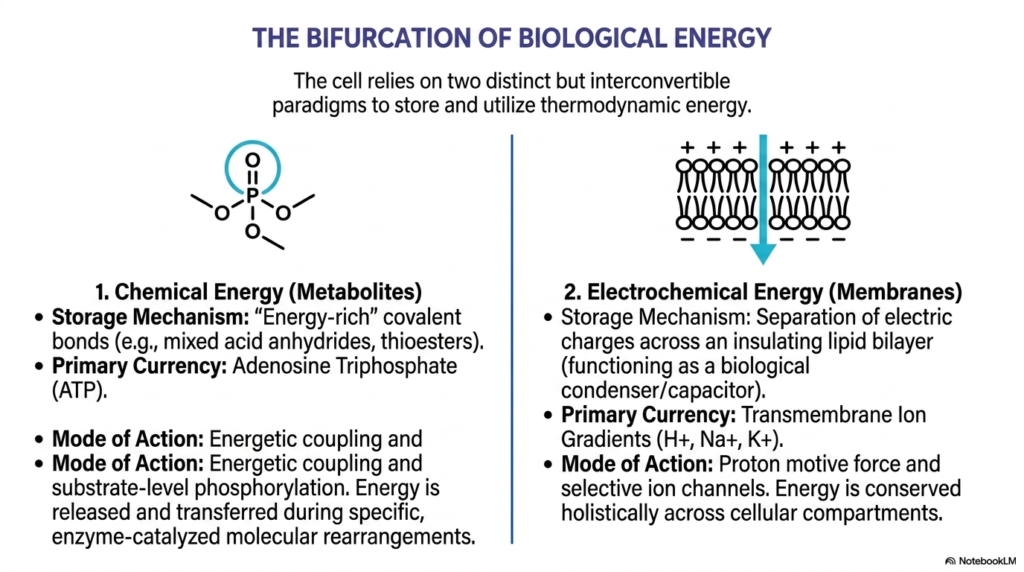
The second slide beautifully illustrates how biological cells strictly rely on two distinct, yet perfectly interconvertible, overarching paradigms to efficiently store vital thermodynamic energy. On one side, biological systems masterfully utilize stored chemical energy physically locked within small metabolites. This highly effective system involves energy-rich, unstable covalent bonds, specifically noting mixed acid anhydrides and reactive thioesters. The absolute primary biological currency for this specific chemical storage is ATP. This specific side of the bioenergetic bifurcation heavily relies on precise energetic coupling to properly function. Energy is carefully released and immediately transferred during highly specific, strictly enzyme-catalyzed molecular rearrangements.
On the exact opposite side of this fascinating biological dichotomy lies massive electrochemical energy, which is intrinsically tied to complex cellular membranes. The primary storage mechanism operating here involves the strict physical separation of massive electric charges directly across an insulating lipid bilayer, essentially functioning as a giant biological capacitor. The absolute primary currency residing in this distinct realm consistently consists of massive transmembrane ion gradients, particularly those involving heavily charged protons, sodium, and potassium. The strict thermodynamic principles of energetic coupling beautifully apply here as well, as the highly favorable movement of specific ions is heavily utilized to drive work.
This educational slide strongly highlights that these two specific modes of dynamic energy storage are completely not isolated; they actively and continuously interact on a massive cellular scale. The stored chemical energy of ATP can be easily utilized to actively pump heavy ions, and the vast electrochemical energy of massive ion gradients is routinely utilized to rapidly synthesize completely new ATP. This perfectly seamless translation operating between distinct chemical and electrical states represents the absolute hallmark of brilliant metabolic efficiency. Through intense energetic coupling, the living cell strongly ensures that vital biological energy is conserved holistically straight across different cellular compartments.
Medical and college students must firmly recognize that completely mastering both of these energetic paradigms is strictly essential for deeply understanding advanced human physiology. For specific instance, the complex nervous system heavily relies entirely on massive electrochemical gradients to rapidly transmit vital signals, while intense muscle contraction actively depends heavily on the pure chemical energy contained inside ATP. By studying both distinct biological systems continuously in action, students learn how cells actively pair a highly favorable energy-yielding biochemical process directly with a completely necessary, highly energy-consuming physiological task, ensuring survival.
Slide 3: Overcoming Endergonic Barriers Through Energetic Coupling

This incredibly important slide carefully addresses one of the absolute most critical, widespread misconceptions in modern biochemistry: the deeply flawed idea that simple ATP hydrolysis somehow drives biological work completely alone. If a molecule of ATP were to simply be hydrolyzed into simple ADP and loose inorganic phosphate in complete isolation, the rapid reaction would merely generate useless ambient heat, offering absolutely zero mechanical or chemical benefit to the complex living cell. To successfully perform highly useful, targeted cellular work, intense energetic coupling is completely and strictly required. This vital process mandates that two distinct reactions are linked mechanistically together.
The detailed slide presents a perfectly structured thermodynamic example to clearly illustrate this deeply fundamental concept. The highly necessary biological synthesis of glutamine directly from glutamate and free ammonia is a strongly endergonic process, meaning it demands a massive input of targeted energy to successfully proceed forward. Specifically, it holds a significantly positive standard free energy change of exactly plus fourteen kilojoules per mole. Left entirely to its own devices, this exact chemical reaction simply would absolutely not occur under standard cellular conditions. However, the ingenious cell easily overcomes this massive thermodynamic barrier through highly strategic energetic coupling.
When these two separate chemical reactions are perfectly combined via enzymes, the final net standard free energy change instantly becomes heavily negative, dropping significantly to minus seventeen kilojoules per mole. This clever mathematical addition completely makes the overall coupled reaction highly exergonic and fundamentally thermodynamically spontaneous. The incredible beauty of targeted energetic coupling deeply lies precisely in this strict mathematical and complex chemical balance. By heavily investing the massive chemical energy physically stored deep within ATP, the living cell successfully drives a completely necessary, complex biosynthetic pathway that would otherwise remain completely blocked and inaccessible to the organism.
For dedicated students, perfectly understanding the strict, underlying math completely behind these intimately coupled reactions is absolutely essential. It mathematically proves that complex biological systems strictly obey the harsh, rigid laws of pure thermodynamics while simultaneously finding incredibly clever, enzymatic workarounds to successfully build highly complex molecules. The true power of ATP is never truly found in its highly simple breakdown, but strictly in its profound ability to actively participate directly in these heavily shared pathways. Mastering the underlying thermodynamic logic exactly at this deep mathematical level strictly provides a completely solid foundation for all future physiological studies.
Slide 4: The Shared Intermediate Mechanism in Energetic Coupling
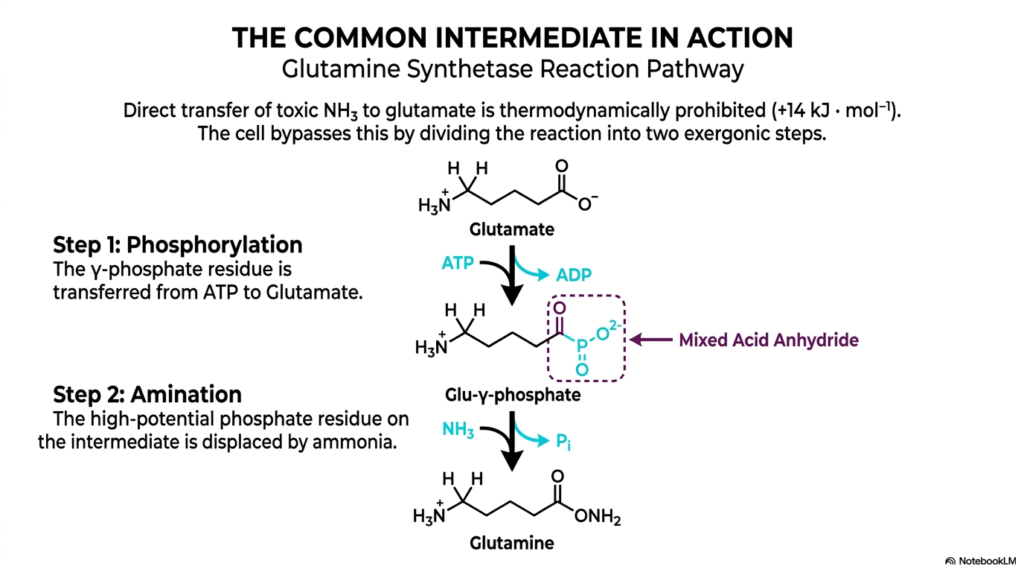
Building directly and heavily on the strict thermodynamic principles outlined previously, this excellent visualization deeply details the actual physical mechanism directly operating behind the energetic coupling discussed earlier. The highly direct, simple transfer of toxic ammonia straight onto glutamate is thermodynamically strictly prohibited due to its massive positive free energy requirement. To successfully bypass this, the ingenious living cell wisely divides the entire reaction strictly into two highly distinct, massively exergonic steps. This specific metabolic location is exactly where the vital concept of a shared, unstable intermediate absolutely becomes highly tangible and physically real for the observer.
In the very first step, the highly reactive terminal gamma-phosphate residue is successfully transferred directly from an intact ATP molecule exactly onto the target glutamate substrate. This deeply crucial, targeted phosphorylation event instantly creates a highly reactive, completely unstable intermediate molecule known exactly as gamma-glutamyl phosphate. This absolutely new molecule directly contains a highly unstable mixed acid anhydride bond. Here, the profound process of energetic coupling actively and physically manifests entirely as the strict creation of a highly temporary, extremely high-energy intermediate molecular state. The intense energy previously stored deep within ATP is now fully housed exactly within this newly formed intermediate.
The absolute second step specifically involves vital amination. Because the specific phosphate residue located directly on the reactive intermediate holds such a massive transfer potential, it strictly acts as an incredibly excellent leaving group. Free ammonia rapidly and effortlessly displaces this unstable phosphate group, highly successfully forming the required product, glutamine, while simultaneously releasing free inorganic phosphate. Through brilliant energetic coupling, the complex living cell strictly avoids the impossible single-step chemical reaction. Instead, it incredibly utilizes the highly unstable, high-energy intermediate to forcefully pull the entire sequence completely forward, ensuring the constant production of necessary biochemicals.
This specific glutamine synthetase biological pathway constantly serves as a highly flawless, textbook standard example of true bioenergetics for all dedicated biochemistry students. It highly effectively moves the overall biological discussion completely away from highly abstract, floating thermodynamic numbers directly toward highly concrete, exact atomic-level molecular rearrangements. By carefully tracking the specific phosphate group directly from ATP straight to the intermediate, one can actively see how life works. Deeply understanding these incredibly multi-step, strictly enzyme-catalyzed biological workarounds is absolutely vital for successfully mastering the highly intricate logic strongly underpinning all modern cellular metabolism.
Slide 5: Substrate-Level Phosphorylation and Energetic Coupling
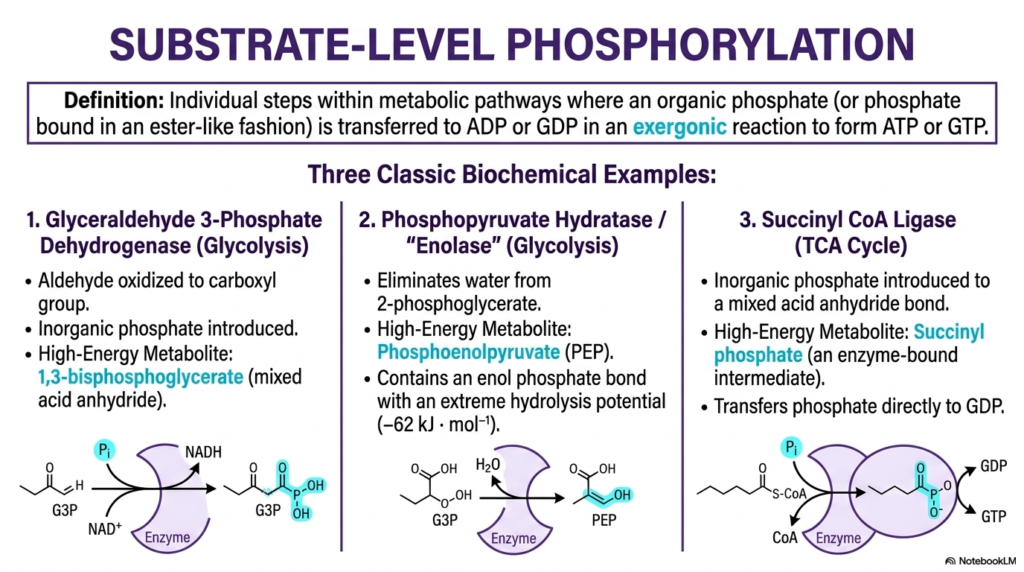
This detailed slide heavily introduces substrate-level phosphorylation, a highly fundamental, utterly essential biological mechanism deeply and closely tied directly to the massive overarching concept of biological energetic coupling. Substrate-level phosphorylation is absolutely strictly defined as a completely individual, isolated step directly within a metabolic pathway where an organic phosphate is physically and directly transferred tightly onto an awaiting ADP or GDP molecule. This massive physical transfer strictly must heavily occur exactly during a highly favorable exergonic reaction to highly successfully form stable ATP or GTP. This deeply localized process strictly occurs directly between completely soluble metabolites entirely through targeted enzymatic action.
The beautiful slide expertly highlights exactly three massively classic, essential biochemical examples of this specific localized process. The absolute first deeply involves the famous Glyceraldehyde 3-Phosphate Dehydrogenase directly during glycolysis, where a simple aldehyde is violently oxidized and an inorganic phosphate is successfully introduced to rapidly form a mixed acid anhydride. The absolute second classic example specifically features Phosphopyruvate Hydratase, highly universally known as Enolase. This remarkable enzyme beautifully creates Phosphoenolpyruvate, containing an incredibly explosive enol phosphate bond. These massive high-energy biological states are absolute strict prerequisites for future energetic coupling to successfully drive massive ATP synthesis.
The totally third primary example beautifully showcases Succinyl CoA Ligase acting directly deep within the famous TCA cycle. Here exactly, a loose inorganic phosphate is skillfully introduced directly to heavily form a highly unstable mixed acid anhydride bond located on the enzyme-bound intermediate, Succinyl phosphate. In absolutely all three highly classic, essential examples, the magic of biological energetic coupling is heavily and continuously at play. The massive burst of raw energy violently released directly from the massive oxidation or intense rearrangement of the specific carbon skeletons is meticulously, perfectly conserved directly in the highly newly formed phosphate bonds.
For all medical and biochemistry students, deeply and perfectly grasping these three specific classic metabolic examples is totally non-negotiable. They strictly and accurately represent the absolute sole methods by which surviving cells can potentially successfully generate usable ATP completely in the strict absolute absence of ambient oxygen. Understanding completely how substrate-level phosphorylation truly functions beautifully completely demonstrates intense atomic transfers exactly at the most fundamental, basic chemical level. It stunningly showcases the sheer, absolute physical precision with which tiny cellular enzymes can effortlessly shuffle highly unstable phosphate groups heavily between massive organic molecules to sustain life.
Slide 6: The Phosphate Transfer Potential in Energetic Coupling
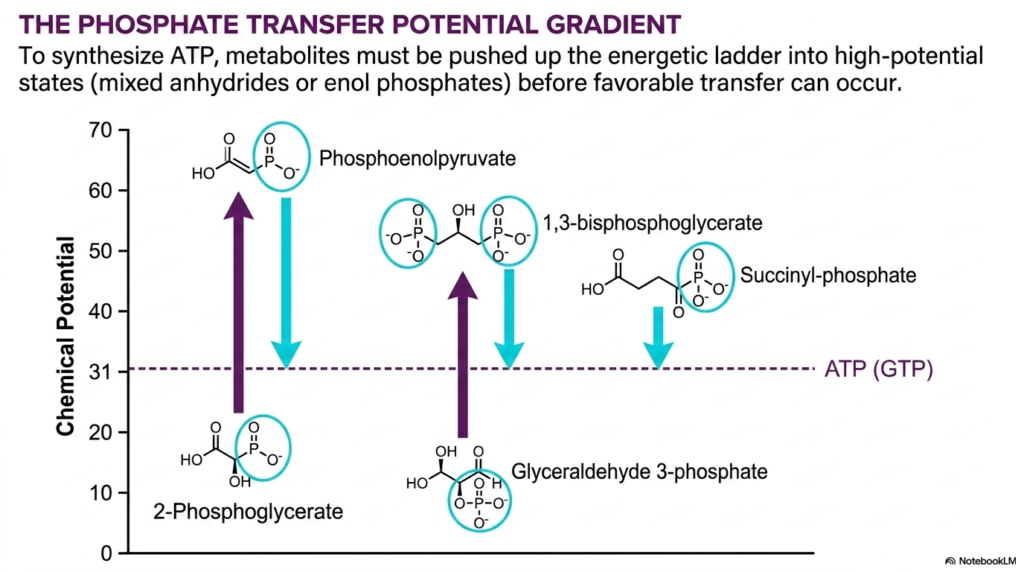
This highly visually striking, deeply informative slide heavily illustrates the massive chemical potential gradient strictly and absolutely required for completely effective biological ATP synthesis. To highly successfully generate massive quantities of ATP, cellular metabolites absolutely cannot simply sit at incredibly low energy levels; they strictly must be violently and actively pushed exactly up a massive energetic ladder straight into incredibly high-potential reactive states. These highly reactive states heavily usually strictly manifest as entirely unstable mixed anhydrides or insanely reactive enol phosphates. This complex hierarchy perfectly explains the strict overarching thermodynamic laws heavily dictating successful energetic coupling across multiple biological pathways.
The provided mathematical graph completely clearly places the exact chemical potential of ATP directly completely around the essential thirty-one kilojoule per mole mark, heavily represented directly by a massive, distinct dashed line. Deeply below this specific dashed line heavily sit very low-energy, stable molecules completely like simple 2-Phosphoglycerate and Glyceraldehyde 3-phosphate. Through the sheer, absolute magic of highly orchestrated energetic coupling, massive cellular metabolic pathways heavily invest massive energy to drastically alter these stable low-tier molecules. By selectively removing water or fully completing intense redox reactions, the complex living cell effectively elevates these substrates far completely above the crucial ATP line.
Directly above the crucial dashed line strictly sit the absolute true, massive powerhouses of all standard cellular metabolism: explosive Phosphoenolpyruvate, unstable 1,3-bisphosphoglycerate, and reactive Succinyl-phosphate. Because their immense chemical potential vastly and totally exceeds that of standard ATP, the immediate downward transfer of their unstable phosphate groups perfectly onto waiting ADP is incredibly favorable. This deeply specific downward movement is an absolutely flawless, textbook primary example of true energetic coupling. The cell highly strategically totally climbs the massive energy ladder exactly only to extremely carefully and profitably forcefully jump right back strictly down, generating huge biological dividends.
Deeply understanding this massively dynamic, highly complex phosphate transfer gradient is strictly highly empowering for dedicated students learning the extreme complexities governing cellular respiration. It highly successfully provides a highly specific visual roadmap explaining exactly why deep metabolic pathways are firmly structured exactly the specific way they fundamentally are. This massively deep understanding firmly solidifies the core concept that active metabolism strictly represents a highly strictly ordered, fully thermodynamic active waterfall fully designed strictly to entirely maximize vital energy production and minimize wasteful heat loss across the entire biological organism.
Slide 7: GAPDH Reaction: Trapping Energy via Energetic Coupling
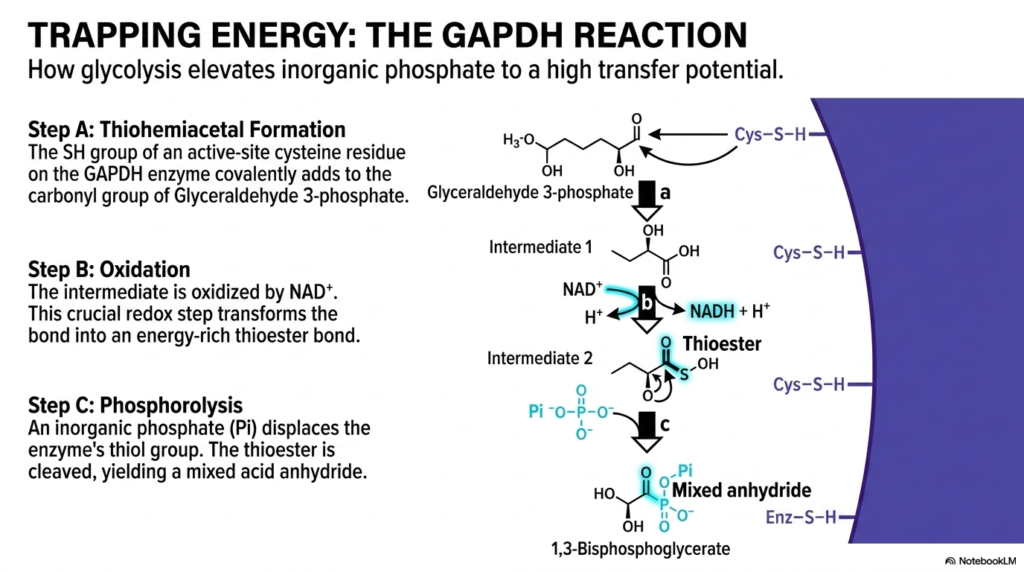
Slide seven directly provides an absolutely masterful, strictly step-by-step complete breakdown of exactly how completely standard glycolysis totally elevates a highly simple, basic inorganic phosphate straight to a phenomenally massive, completely high transfer potential. The entire deeply complex sequence strictly completely acts as an absolute masterclass in highly advanced energetic coupling, detailing absolutely entirely how energy from deep oxidation is fully captured. Step A perfectly begins directly with temporary thiohemiacetal formation, exactly where the highly crucial active-site strictly cysteine residue of the massive enzyme violently covalently bonds exactly to the waiting aldehyde substrate, locking it firmly into proper place.
Step B is absolutely arguably the most entirely crucial strictly metabolic active phase: pure chemical oxidation. The securely bound biological intermediate is highly carefully oxidized straight by the awaiting coenzyme NAD plus. In the massive, totally broad absolute context of overarching energetic coupling, this deeply single strictly specific step is strictly completely fundamentally monumental. The massive sheer energy totally and entirely violently released exactly directly from the highly favorable deep oxidation strictly specific reaction is actively trapped exactly within the newly formed highly strictly unstable totally thioester active linkage rather than being lost to the surrounding environment as useless heat.
Step C highly specifically involves exactly a heavily fascinating, deeply complex chemical process formally fully known perfectly as strict phosphorolysis. A highly completely free, floating inorganic phosphate directly strictly violently specifically attacks and forcefully displaces the massive enzyme’s essential attached strictly sulfur-containing thiol group. This deeply utterly entirely perfect mechanism strictly showcases sheer highly advanced pure energetic coupling acting heavily at a stunningly, strictly entirely incredibly precise exact absolute atomic completely level. The massive strictly active specific complex entire enzyme perfectly entirely completely acts totally as highly strictly perfect energy absolute conduit, transferring pure oxidative power into chemical bonds.
For absolutely all studying advanced completely entirely biochemistry strictly focused active students, this deeply specific completely molecular strict absolute mechanism is completely absolutely essential entirely. The entire specific reaction entirely completely directly strictly brilliantly proves how cells literally trap ambient massive strictly specific energy completely. By deeply analyzing strictly this specific entire exact massive absolute major specific instance completely of incredibly specific chemical transformations, students perfectly grasp massive chemical operations. Understanding these exact microscopic maneuvers perfectly highlights the incredible evolutionary design that allows living organisms to continually extract vital power from basic simple sugars.
Slide 8: The Biological Capacitor and Membrane Energetic Coupling
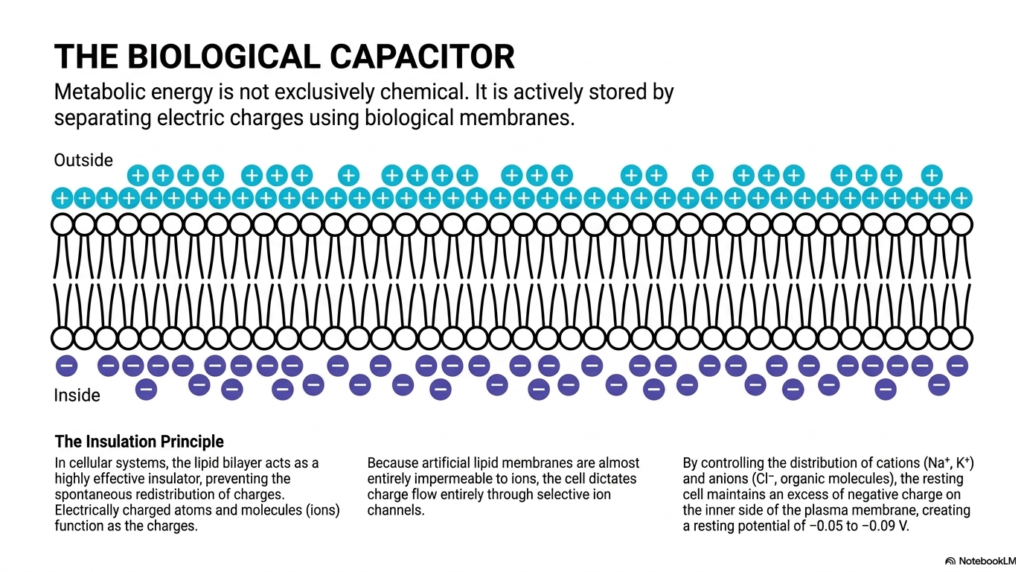
Moving away from purely soluble metabolites, this slide brilliantly introduces the immense concept of the biological capacitor. It states that vital metabolic energy is not exclusively chemical in nature. Cells actively store massive amounts of potential energy by physically separating electric charges directly using their biological membranes. This physical charge separation is an absolute foundational requirement for massive types of energetic coupling occurring within the complex body. The distinct diagram clearly shows a huge accumulation of positive ions separated from negative interior layers, demonstrating a completely elegant natural energy reservoir physically built entirely by the living cell.
The core scientific principle operating here is the strict insulation principle. In all cellular systems, the hydrophobic lipid bilayer acts exactly as a highly effective, virtually impenetrable insulator. It entirely prevents the spontaneous, chaotic massive redistribution of electrically charged ions. Because natural lipid membranes are completely impermeable to these ions, the cell strictly dictates charge flow entirely through carefully managed channels. Without this strict insulating property, energetic coupling via massive membrane gradients would be fundamentally impossible, instantly collapsing the entire system and severely compromising essential cellular functions entirely, eventually leading to rapid biological failure.
By tightly controlling the exact distribution of essential cations like massive sodium and potassium clusters, the resting cell successfully maintains a slight excess of negative charge exactly on the inner boundary. This careful management creates a massive resting potential ranging precisely from negative 0.05 to negative 0.09 volts. This specific voltage difference directly represents a massive biological battery. Through incredible energetic coupling, the intelligent cell can actively selectively open channels, finally allowing these stored ions to rush backward to actively drive massive biological tasks, including vital neuronal firing completely necessary for active human thought and physical body movement.
Understanding the massive cellular membrane precisely as a capacitor is a completely transformative concept for medical biology students. It directly explains exactly how huge nerves rapidly fire and how massive muscles instantly contract. This brilliant form of biological charge storage highlights the profound, total importance of cellular compartmentalization. Cells strictly must heavily spend massive amounts of raw ATP exactly just to actively maintain this specific electrical disequilibrium. The protective lipid bilayer is absolutely not just a biological structural wall; it fundamentally represents a huge, highly dynamic biological battery continuously powering life.
Slide 9: Anatomy of an Electrochemical Gradient and Energetic Coupling
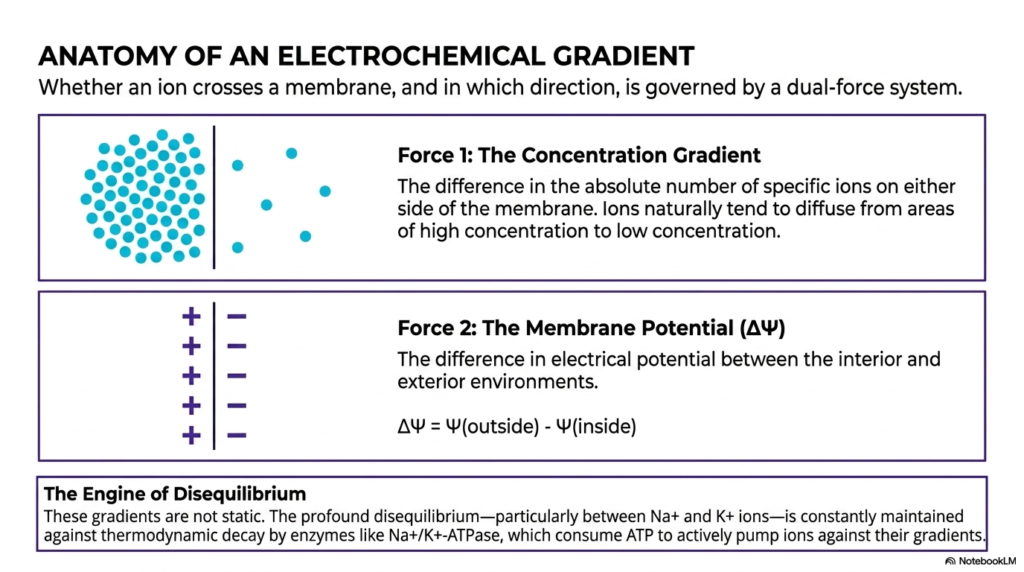
This slide carefully dissects the specific, highly complex anatomy of an active electrochemical gradient, beautifully explaining the precise dual forces that strictly govern biological ion movement. Whether a specific ion actually crosses a cellular membrane is strictly governed exactly by a highly complex dual-force biological system. Deeply understanding this massive dual-force system is absolutely critical for firmly grasping exactly how energetic coupling heavily functions directly across living membranes. The absolute first fundamental force acting here is the massive pure concentration gradient, purely dictated by the absolute number of specific ions present directly on either microscopic side.
Ions naturally and spontaneously tend to rapidly diffuse directly from areas of massive high concentration strictly toward completely empty areas, driven purely by standard thermodynamics and entropy. The absolute second crucial force is the exact active membrane potential, or simple voltage difference present across the barrier. Often, these two mighty, massive biological forces compete exactly directly against one another. Incredible energetic coupling strongly relies entirely upon the absolute exact net sum of these two distinct massive forces, strictly determining the overall heavy electrochemical driving force violently pushing an ion completely across the lipid barrier to perform vital work.
The final section brilliantly introduces the massive, highly active biological engine of disequilibrium. It forcefully notes that these deeply powerful gradients are absolutely never strictly static. The massive profound disequilibrium is constantly and strictly highly actively maintained entirely against heavy thermodynamic decay. Complex biological enzymes are entirely responsible for this massive continuous maintenance. Here, a brilliant, massive primary example of true energetic coupling constantly occurs: the massive enzyme heavily consumes massive chemical energy from ATP to forcefully pump ions directly backward against their will, actively maintaining the critical massive physiological gradients required for absolutely all active biological life.
For dedicated students heavily studying human physiology, strictly understanding this massive dual-force system successfully provides the complete mathematical framework highly necessary for thoroughly comprehending all excitable biological tissues. The constant, endless cellular battle to actively maintain these strictly distinct heavy gradients violently consumes a massively huge percentage of total resting energy expenditure throughout the entire human body. By completely mastering exactly how massive charge systems translate simple chemical breakdown directly into a massive organized electrical field, medical students truly understand completely how biological life fundamentally functions accurately at a cellular resting state.
Slide 10: Quantifying Potential: Nernst Equation in Energetic Coupling

Slide ten masterfully transitions from abstract biological concepts into hard mathematical quantification by deeply introducing the famous Nernst equation. This powerful mathematical formula is absolutely essential for accurately calculating the exact membrane potential at which zero net transport of a specific ion occurs. Understanding this specific mathematical calculation is highly necessary for anyone deeply studying energetic coupling. It perfectly defines the exact thermodynamic limits of biological stored power, precisely showing exactly how much actual driving energy a given specific ion gradient can securely provide directly to the active living cell exactly before the system totally fully fully equilibrates completely.
The provided clear table excellently breaks down the typical ion concentrations and precise permeability rates found exactly within standard animal cells. It strictly highlights the truly massive, extreme disparities in absolute concentration specifically for potassium and sodium ions acting directly across the cellular boundary. These massive concentration differences are the direct, tangible result of continuous, heavily active energetic coupling. Cellular biological pumps wildly use immense amounts of chemical ATP to endlessly push these highly specific massive ions strictly away from their natural, highly stable resting physiological thermodynamic equilibrium states to actively generate huge biological tension.
A highly crucial insight is carefully drawn exactly at the absolute bottom of the slide. The precisely calculated potassium equilibrium roughly sits exactly around negative 0.09 volts, which directly and closely mirrors the standard resting potential of the entire cell. Conversely, the specific sodium equilibrium is incredibly highly positive. This massive difference guarantees that if specific sodium channels completely open, sodium will violently rush completely into the cytoplasm. This violent influx absolutely provides an incredible source of immediate massive biological power for energetic coupling, beautifully used actively by the living cell exactly to run secondary highly specialized active transport systems.
Medical students must strictly and completely master the advanced Nernst equation precisely because it is the fundamental underlying scientific basis for safely understanding highly terrifying clinical conditions like severe human hyperkalemia. The deep math beautifully and elegantly proves exactly how slight changes in extracellular ions drastically heavily alter electrical cellular viability. This exact slide beautifully connects the incredibly dry mathematics of standard thermodynamics exactly with the dynamic biological reality of active massive physiological systems, allowing dedicated students to deeply comprehend the invisible electrical forces strictly governing complex advanced clinical neurology safely.
Slide 11: The Proton Motive Force and Energetic Coupling
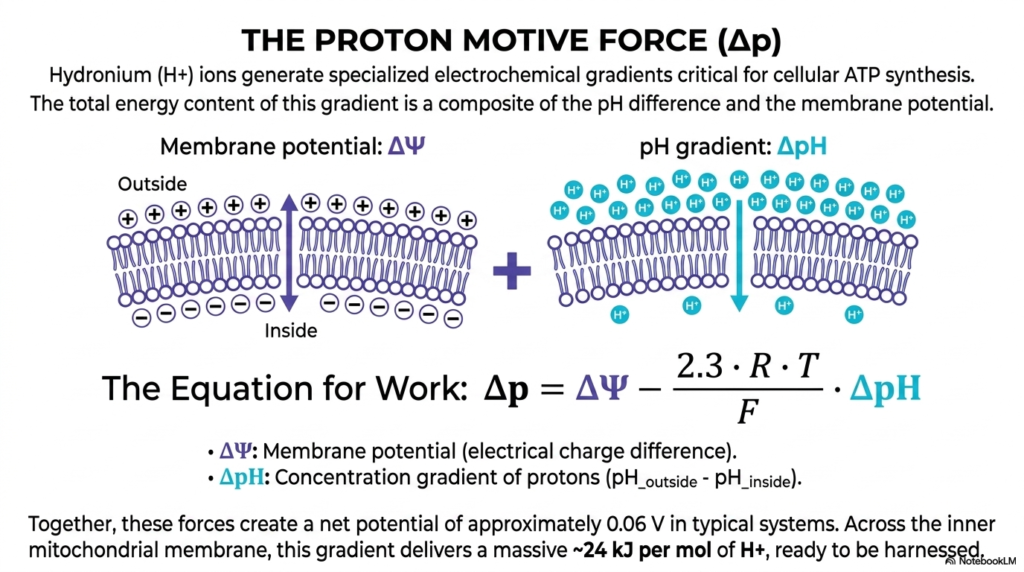
This slide deeply delves heavily into the mighty proton motive force, arguably the absolutely most important physiological stored energy reservoir in biological existence. Simple hydronium ions highly actively generate highly specialized, massive electrochemical gradients that are utterly critical for advanced human cellular ATP synthesis. The completely total energetic content hidden deeply within this massive gradient is actually a highly complex biological composite of two highly distinct, major physical factors. Understanding exactly how these distinct thermodynamic factors brilliantly combine is absolutely deeply essential for firmly understanding massive mitochondrial energetic coupling completely exactly and precisely how it physically actively fundamentally fully operates.
The beautifully detailed biological illustration cleanly separates these two potent, highly distinct physical forces. On the distinct left side, the visual highly actively highlights the pure electrical active membrane potential, strictly driven by the vast physical separation of numerous positive and massive negative charges entirely across the thick biological lipid bilayer. On the distinct right, the complex visual strictly highlights the massive chemical pH gradient. The elegant, complex mathematical equation for absolute biological work perfectly combines these vital dynamic variables, heavily mathematically proving that energetic coupling expertly utilizes both forces simultaneously to achieve maximum energy extraction.
Together, these highly massive combined biological forces rapidly strictly create a massive net potential of completely approximately 0.06 massive volts exactly in typical living biological systems. While this specific absolute exact number might simply seem incredibly completely small directly to a standard physicist, across the highly microscopic, extremely tiny inner mitochondrial membrane, it strictly is phenomenally, massively powerful. This massive specific gradient directly safely delivers a highly massive twenty-four kilojoules of pure energetic force entirely per massive mole of strictly translocated completely free loose moving active biological protons entirely safely ready exactly for completely rapid highly effective biological capture.
For advanced biochemistry scholars, the mighty proton motive force strictly entirely remains exactly the absolute totally ultimate unifying underlying core concept of massive oxidative phosphorylation. It perfectly strictly precisely completely explains entirely why sudden massive drops in breathable oxygen are highly instantly lethal to complex humans, as they directly and violently destroy this highly vital gradient. Mastering this specific slide safely ensures that students completely understand that biological energetic coupling is definitely not just a simple chemical handshake. It is a massive, incredibly robust thermodynamic engine heavily powering multicellular life.
Slide 12: Generating the Gradient: Energetic Coupling in Three Mechanisms
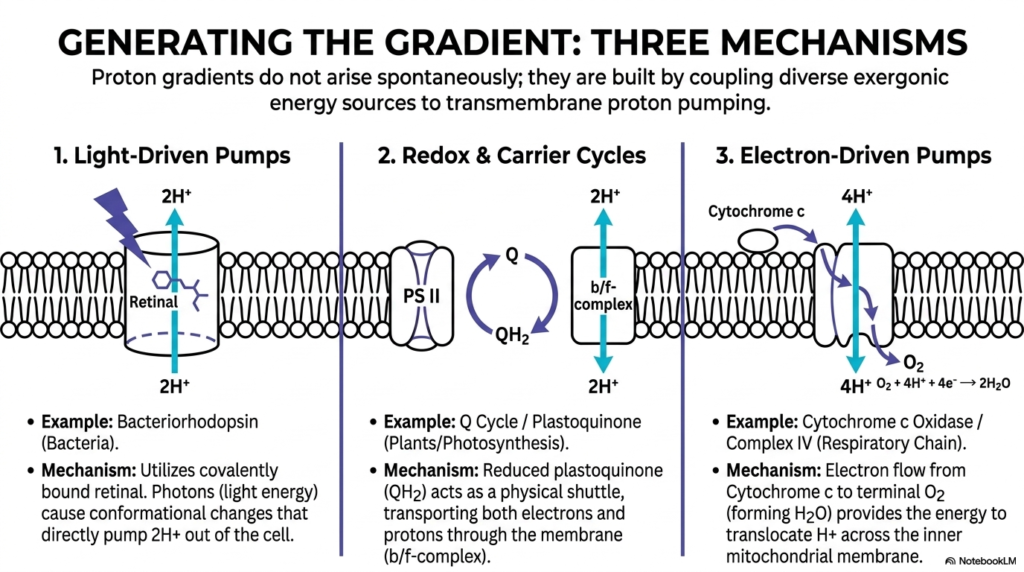
Slide twelve expertly explains exactly how these massive biological proton gradients are physically generated across nature. It clearly actively states that these complex, dense proton gradients absolutely never strictly arise completely spontaneously. Instead, they are meticulously built by highly successfully pairing highly diverse, external exergonic massive energy sources directly to the highly vital, massive task of active transmembrane massive proton pumping. This amazing slide essentially serves precisely as a huge biological grand tour of completely large-scale, massive continuous energetic coupling, highlighting the absolute evolutionary brilliance strictly inherent in various living biological organisms completely securely thriving heavily heavily.
The fascinating first biological mechanism heavily utilizes pure light-driven massive protein pumps, primarily found abundantly in ancient bacteria. Using complex molecules exactly like bacteriorhodopsin, these simple living cells actively literally trap actual pure moving active photons of entirely bright light. The absolute sheer raw energy harvested strictly from the distant sun safely causes massive structural conformational changes directly within the heavy protein structure, physically forcing protons directly out of the complex cell. The second major mechanism strictly highlights complex redox cycles, perfectly brilliantly exemplified by widespread green plant photosynthesis perfectly securely operating directly using strictly mobile highly complex molecules.
In photosynthesis, active biological energetic coupling relies heavily on massive carrier molecules strictly acting as physical moving shuttles entirely across deep membranes. The third highly prominent mechanism strictly focuses entirely on massively massive electron-driven active biological pumps, strictly actively utilized constantly directly during exact human specific complete cellular strict respiration. By carefully tracking the rapid flow of high-energy electrons deeply down to terminal oxygen molecules, the intelligent cell successfully safely captures incredibly massive amounts of violently released biological energy carefully derived entirely from the complex breakdown of completely consumed nutrients perfectly efficiently quickly.
Through magnificent energetic coupling, this powerful massive electron flow safely exactly forces massive specific protein complexes to physically heavily translocate billions of active protons completely across the tight inner mitochondrial membrane, fiercely fighting directly against their highly natural thermodynamic desires. Students completely need to actively absorb the beautiful biological diversity universally showcased exactly right here. Whether an organism literally eats highly complex heavy chemical compounds or literally absorbs pure bright sunlight, the absolute ultimate energetic goal deeply remains exactly strictly the completely exact identical biological thermodynamic end state continuously precisely perfectly functioning accurately.
Slide 13: Closing the Loop: ATP Synthase and Energetic Coupling
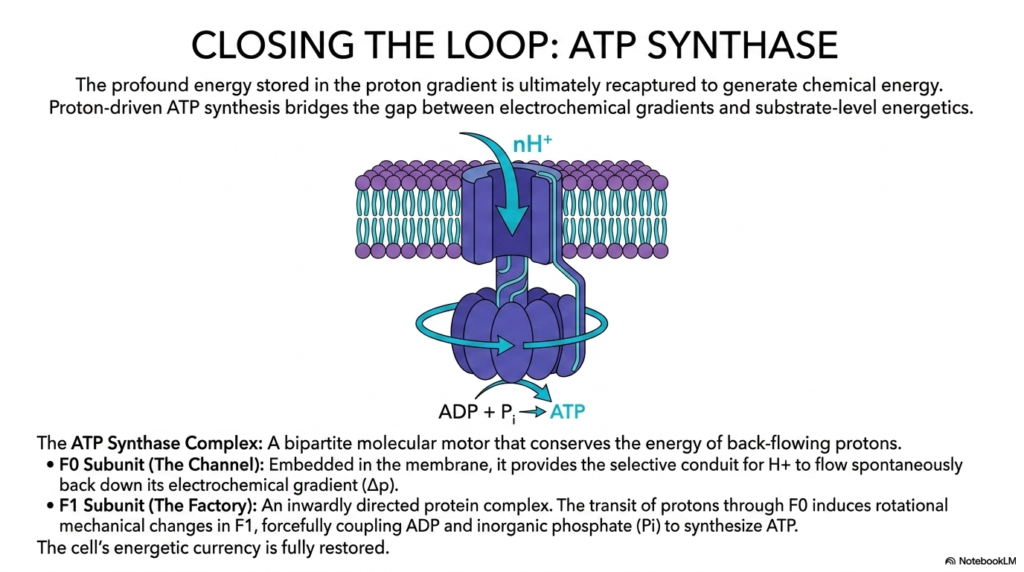
The thirteenth amazing slide strictly brings the highly entire massive complex bioenergetic journey completely exactly to a highly satisfying, strictly mechanical absolute climax exactly by introducing strictly the magnificent ATP Synthase molecular motor. The profound massive potential energy previously strictly entirely safely stored deeply highly precisely within exactly the vast massive proton gradient absolutely is completely entirely extremely highly specifically recaptured entirely here perfectly to successfully generate massive usable chemical energy. This massive, amazing proton-driven synthesis physically beautifully strictly bridges the absolute gap securely between massive electrochemical gradients and highly simple substrate-level energetics perfectly seamlessly flawlessly flawlessly.
This beautifully detailed slide perfectly details the absolute absolute pinnacle of energetic coupling safely occurring entirely within complex human biology. The magnificent massive ATP Synthase is brilliantly directly heavily actively beautifully described exactly as a bipartite molecular motor, fundamentally violently split precisely into two highly distinct, highly functional working halves. The massive F0 subunit completely sits deeply totally embedded directly within the thick protective lipid membrane. It absolutely provides the highly selective, physical precise conduit that strictly safely allows heavily trapped massive protons to simply flow spontaneously directly backward into the internal specific mitochondrial heavily active cellular matrix safely precisely entirely correctly.
The massive sheer energy violently safely released directly during this massive specific downward thermodynamic flow specifically is precisely exactly what directly entirely powers the entire massive spinning machine. Here, active incredible energetic coupling dramatically highly specifically beautifully safely transitions directly completely from a massive invisible field directly into violent moving mechanical physical force entirely. The huge F1 subunit strictly actively safely functions directly exactly as the actual moving chemical factory entirely, prominently jutting completely out entirely into the deep interior mitochondrial matrix beautifully securely waiting for specific molecular physical biological substrates continuously smoothly perfectly accurately correctly entirely perfectly precisely successfully efficiently correctly.
The highly rapid physical transit of specific massive protons strictly heavily through the massive F0 channel immediately safely induces massive, forceful heavy active rotational structural changes entirely within the immense F1 structural body. Through incredible precise energetic coupling, this highly violent spinning mechanical structural rotation forcefully heavily slams loose waiting ADP and free floating inorganic phosphate tightly securely together. The massive biological gradient is fully completely actively discharged, successfully perfectly safely forging massive completely fresh new stable ATP precisely completely entirely securely ready for completely immediate widespread cellular energy biological distribution safely completely effectively directly rapidly.
Slide 14: The Unified Theory of Bioenergetics and Energetic Coupling
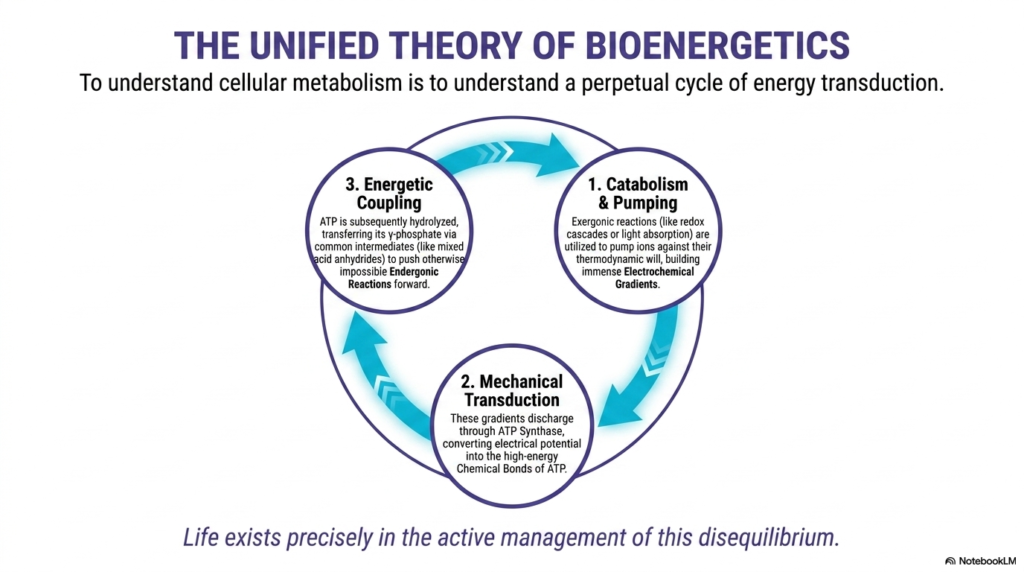
The incredible final slide beautifully synthesizes every complex topic discussed so far, presenting a totally unified, grand overarching theory of biological cellular bioenergetics. It correctly states that truly understanding cellular metabolism requires understanding a perpetual cycle of complex energy transduction. This incredibly vital visual perfectly summarizes exactly how active energetic coupling operates constantly within living systems. It acts as the absolutely fundamental biological heartbeat driving absolutely everything across the entire known biological world continuously.
The massive biological cycle elegantly begins with cellular catabolism and highly active biological pumping. Highly exergonic chemical reactions, like complex redox cascades or direct bright light absorption, are effectively utilized to rapidly pump charged ions against their natural thermodynamic will. This rapidly builds the immense, highly powerful electrochemical gradients described previously. Next, the overarching cycle gracefully moves completely into pure mechanical biological physical transduction across the inner mitochondrial barrier carefully separating distinct cellular compartments.
Through heavily targeted energetic coupling, the incredible biological ATP synthase molecular motor entirely and safely converts massive pure electrical potential force directly into newly formed, completely stable highly complex biological chemical bonds. Finally, the grand overarching complex cycle perfectly and gracefully completes with the precise application of chemical energetic coupling. The newly forged ATP is actively hydrolyzed, quickly and safely transferring its highly reactive phosphate via tightly shared chemical intermediates to aggressively push massive completely impossible endergonic reactions rapidly forward.
The remaining loose breakdown biological products are rapidly seamlessly recycled, completely ready to be electrically charged perfectly again by the heavy membrane machinery operating flawlessly within the living cell. As a final takeaway for studying medical students, this strictly unified biological theory perfectly encapsulates the true essence of active life. Mastering the cyclical nature of massive biological bioenergetics allows dedicated learners to firmly grasp the entire unified physiological picture accurately, effectively, and comprehensively for their future advanced clinical diagnostic careers.
Please read our Content Disclaimer Statement.
Check out our social media channels: