38. The Tricarboxylic Acid Cycle: Biochemical Pathways and Mechanisms
Every living organism requires a constant, reliable source of energy to survive, grow, and adapt. At the very heart of this microscopic power grid lies a biochemical marvel that transforms the food organisms consume into cellular currency. This slide deck explores the fundamental mechanics of cellular respiration. The core purpose of these presentations is to demystify complex biochemical structures and metabolic pathways, providing college and medical students with a comprehensive, easily digestible breakdown of how cells sustain human life through precise molecular reactions.
Slide 1: Introduction to the Tricarboxylic Acid Cycle
The Tricarboxylic Acid Cycle stands as one of the most critical topics in biochemistry and cellular metabolism. Often visualized as a simple spinning wheel of chemical reactions, it is actually a highly orchestrated sequence of biochemical events. This presentation delves into the deep mechanisms, structural biology, and metabolic regulation of this essential pathway. Understanding the Tricarboxylic Acid Cycle is fundamental for anyone pursuing advanced biological sciences or medicine. The introductory slide sets a serious, academic tone, featuring a detailed molecular structure that underscores the biochemical complexity awaiting exploration.
At its core, the Tricarboxylic Acid Cycle represents the great metabolic clearinghouse of the cell. It takes the fragmented remnants of carbohydrates, fats, and proteins and systematically extracts their hidden energy. When students examine the cover image of the slide deck, the complex ribbon structure of an enzyme is visible behind the chemical structures. This imagery perfectly encapsulates the intersection of structural biology and enzymatic function. The enzymes driving the Tricarboxylic Acid Cycle are highly specialized machines, carefully folded to execute incredibly specific chemical modifications.
As this educational journey begins, one must appreciate that metabolic regulation is not just about producing energy. It is also about maintaining cellular balance in a dynamic environment. The overarching theme of this deck is how structural biology enables the precise metabolic regulation required for survival. By mastering the fundamental steps outlined in the upcoming slides, a deeper appreciation for cellular bioenergetics will naturally develop. The foundation is now set to explore how molecular structures dictate the energetic yield that ultimately powers complex biological organisms.
To truly grasp the gravity of this metabolic pathway, one must look beyond rote memorization. The biochemical mechanisms discussed in the subsequent slides will reveal an elegant logic for how carbon is oxidized and energy is conserved. This slide deck aims to bridge the gap between static textbook diagrams and the dynamic reality of cellular metabolism. Through this detailed exploration, the true beauty and complexity of cellular respiration will become undeniably clear to the developing medical or scientific mind.
Slide 2: The Amphibolic Hub of the Tricarboxylic Acid Cycle
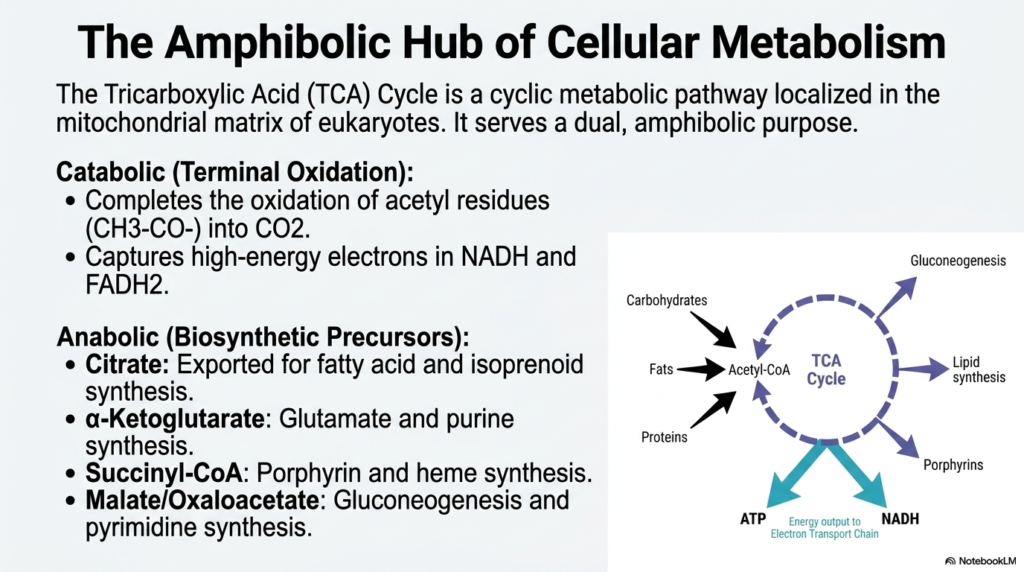
Located deep within the mitochondrial matrix of eukaryotic cells, the Tricarboxylic Acid Cycle operates as a dual-purpose metabolic engine. It is famously known as an amphibolic hub, meaning it serves both catabolic and anabolic functions simultaneously. On the catabolic side, the cycle carries out the terminal oxidation of acetyl residues, reducing them to carbon dioxide. In doing so, the Tricarboxylic Acid Cycle captures high-energy electrons in carrier molecules such as NADH and FADH2. This electron capture is absolutely essential, as these high-energy particles are destined for the electron transport chain, which powers the generation of massive amounts of energy.
Beyond breaking molecules down, the Tricarboxylic Acid Cycle is equally crucial for building them up. The anabolic role of this pathway provides the structural precursors needed for widespread cellular biosynthesis. For instance, citrate can be exported from the mitochondrion to initiate the synthesis of fatty acids and isoprenoids. Similarly, alpha-ketoglutarate serves as a fundamental building block for glutamate and purine synthesis. These intermediates are constantly siphoned off to create the structural and functional molecules required for cellular growth, repair, and replication.
The cycle also provides succinyl-CoA, which is a strictly necessary precursor for the synthesis of porphyrins and heme. Heme is vital for oxygen transport in red blood cells, highlighting the systemic importance of these biochemical pathways. Furthermore, malate and oxaloacetate can be diverted toward gluconeogenesis and pyrimidine synthesis. This massive interconnectivity demonstrates why the Tricarboxylic Acid Cycle is the true metabolic crossroads of the cell. Carbohydrates, fats, and proteins all converge at acetyl-CoA to enter this spinning wheel of biochemical transformation.
Understanding this amphibolic nature is critical for medical students, as metabolic diseases often disrupt this delicate balance. If too many intermediates are pulled for biosynthesis, the catabolic energy-generating function can be compromised. Therefore, the cell must constantly monitor and adjust the flow of molecules into and out of the pathway. The elegance of cellular metabolism lies in this continuous, dynamic balancing act, ensuring that neither energy production nor macromolecule biosynthesis is neglected during physiological stress.
Slide 3: Aldol Condensation Commits Carbon to the Tricarboxylic Acid Cycle

The official starting point of the Tricarboxylic Acid Cycle involves a fascinating biochemical reaction catalyzed by the enzyme citrate synthase. This step marks the exact moment that a two-carbon acetyl group is committed to complete oxidation. The mechanism is a classic aldol condensation, where the methyl group of acetyl-CoA reacts with the carbonyl carbon of oxaloacetate. This joining of a two-carbon molecule with a four-carbon molecule produces the six-carbon compound known as citrate. It is this very molecule that gives the Tricarboxylic Acid Cycle one of its alternative names.
From a thermodynamic perspective, this initial reaction is incredibly powerful. The condensation process involves hydrolysis of a high-energy thioester bond, thereby releasing Coenzyme A (CoA-SH). This hydrolysis is highly exergonic, meaning it releases a substantial amount of energy. Because the energy release is so massive, this specific step of the Tricarboxylic Acid Cycle is practically irreversible under cellular conditions. The massive drop in free energy effectively acts as a biochemical engine, driving the entire metabolic cycle toward equilibrium.
The structural biology of citrate synthase is perfectly adapted to facilitate this specific reaction. The enzyme must carefully exclude water from the active site until the citryl-CoA intermediate is formed, preventing wasteful hydrolysis of the precious thioester bond. Once the condensation is complete, the active site shifts to cleave the bond, releasing the final citrate molecule. This level of precise enzymatic control ensures that the energy stored in the acetyl-CoA linkage is efficiently used to drive the pathway forward rather than being lost as heat.
For students studying metabolic regulation, this committed step is a focal point of cellular control. Because it is highly exergonic and irreversible, it is a logical site to regulate the overall rate of the pathway. By understanding the thermodynamic driver of this aldol condensation, one gains insight into the physical laws governing cellular life. The forceful push generated by citrate synthase ensures that, as long as fuel is available, the cellular metabolic engine will continue to turn.
Slide 4: Stereospecific Isomerization in the Tricarboxylic Acid Cycle
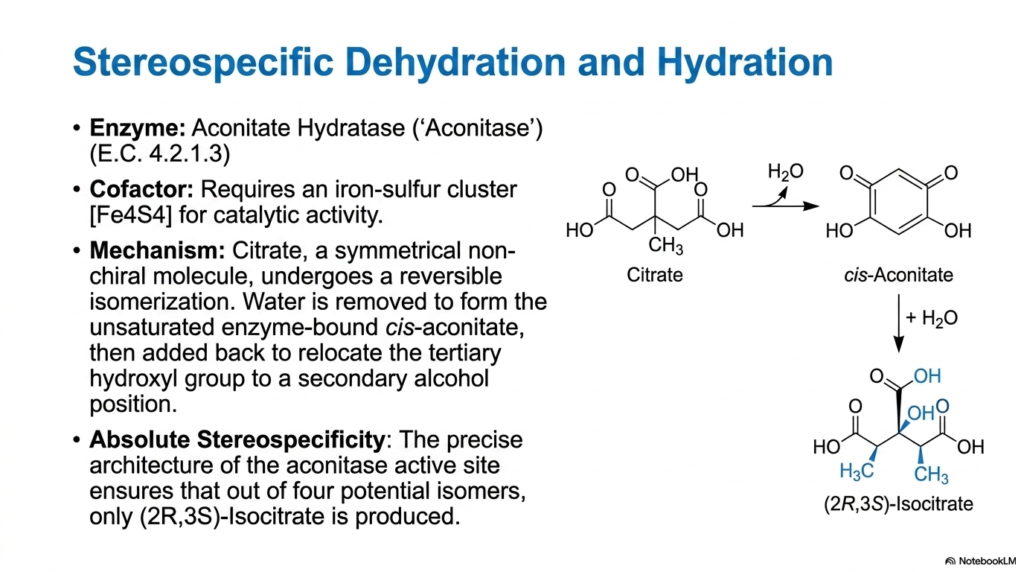
Following the synthesis of citrate, the Tricarboxylic Acid Cycle must rearrange this molecule to prepare it for subsequent energy-extracting steps. This task falls to the enzyme aconitase, officially known as aconitate hydratase. Citrate is a symmetrical, non-chiral molecule, making it difficult to oxidize optimally. Aconitase solves this problem by performing a highly coordinated, two-step reversible isomerization. First, water is removed via dehydration to form a transient, enzyme-bound intermediate called cis-aconitate. Then, water is added back via hydration, relocating the tertiary hydroxyl group to a secondary alcohol position.
The repositioning of the hydroxyl group transforms the molecule into isocitrate. This isomerization is necessary because a secondary alcohol is much more readily oxidized in subsequent stages of the Tricarboxylic Acid Cycle. Aconitase requires a highly specialized iron-sulfur cluster to achieve catalytic activity. This metallic cluster is not involved in electron transfer, its usual biological role, but rather acts directly in binding and precisely positioning the substrate. It holds the citrate molecule in a strict orientation, dictating the precise geometry of the dehydration and rehydration events.
The most remarkable feature of this reaction within the Tricarboxylic Acid Cycle is its absolute stereospecificity. Although the starting molecule, citrate, possesses no chiral centers, the resulting product is intensely specific. Out of four mathematically possible stereoisomers that could be created during the hydration of cis-aconitate, the unique architecture of the aconitase active site ensures that only (2R,3S)-isocitrate is ever produced in the mitochondrion. This biological precision ensures the molecule fits perfectly into the active site of the next enzyme in the pathway.
Aconitase provides a beautiful example of how three-dimensional enzyme structure dictates metabolic function. If the molecule were allowed to freely rotate or if water were added randomly, the pathway would grind to an immediate halt. Absolute stereospecificity prevents the accumulation of useless isomers that the cell cannot metabolize. By carefully escorting the substrate through this transformation, aconitase readies the cellular fuel for the intense oxidative steps that follow, setting the stage for major energy harvesting.
Slide 5: The First Oxidative Decarboxylation of the Tricarboxylic Acid Cycle

The repositioning of the hydroxyl group in the previous step perfectly prepares the molecule for the first major energy harvest of the Tricarboxylic Acid Cycle. Catalyzed by isocitrate dehydrogenase, this step involves oxidative decarboxylation. First, the secondary hydroxyl group of isocitrate is oxidized to form a transient intermediate, oxalosuccinate. This intermediate is highly unstable and spontaneously decarboxylates, losing a carboxyl group as carbon dioxide. The resulting product is the five-carbon molecule known as 2-oxoglutarate, also called alpha-ketoglutarate.
This specific reaction holds immense metabolic significance within the overall Tricarboxylic Acid Cycle. It represents the very first time carbon is lost from the biochemical skeleton as carbon dioxide gas. More importantly, this step features the first significant capture of high-energy electrons. As the substrate is oxidized, a molecule of NAD+ is reduced to form NADH. This newly synthesized NADH acts as a biological battery, carrying its valuable electrons straight to the electron transport chain, where it will eventually drive the generation of roughly 2.5 ATP molecules.
Because this reaction involves the permanent loss of carbon dioxide, the first oxidative decarboxylation is strongly exergonic and biochemically irreversible. Consequently, isocitrate dehydrogenase functions as the primary rate-limiting enzyme for the entire Tricarboxylic Acid Cycle. The speed at which this enzyme processes substrate effectively sets the pace for the entire metabolic pathway. If cellular energy levels are high, this enzyme slows down, conserving fuel. If energy is in drastic demand, the enzyme accelerates, pushing more carbon through the oxidative machinery.
For students analyzing cellular respiration, this enzyme highlights the intersection of chemistry and physiological demand. The transition from a six-carbon molecule to a five-carbon molecule represents a pivotal narrowing of the metabolic funnel. The structural visualization of the electrons moving to NAD+ provides a clear picture of how chemical bonds are translated into mobile energy carriers. Grasping the irreversible nature of this step is essential for understanding how cells prevent futile cycling and maintain forward momentum in energy metabolism.
Slide 6: The Second Oxidative Decarboxylation in the Tricarboxylic Acid Cycle
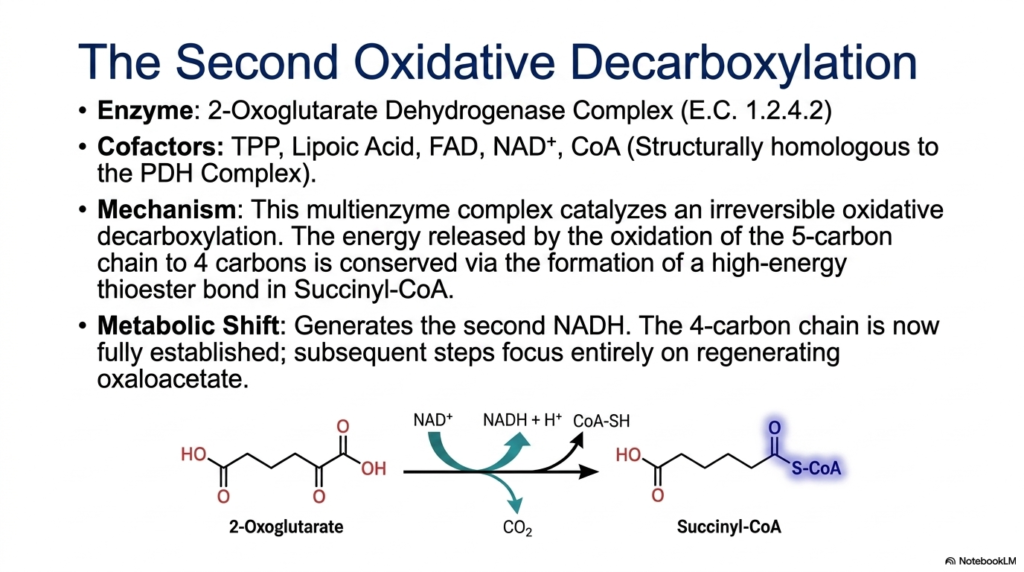
Following the creation of the five-carbon 2-oxoglutarate, the Tricarboxylic Acid Cycle executes its second oxidative decarboxylation. This massive biochemical task is performed by the 2-oxoglutarate dehydrogenase complex. This is not a single enzyme, but rather a massive multienzyme complex composed of three distinct catalytic components. It is structurally and functionally homologous to the famous Pyruvate Dehydrogenase complex. To function properly, this molecular machine requires five separate cofactors: Thiamine Pyrophosphate (TPP), Lipoic Acid, FAD, NAD+, and Coenzyme A.
The mechanism catalyzed by this complex is robust and highly exergonic. The five-carbon chain undergoes an irreversible oxidative decarboxylation, releasing the second and final carbon dioxide molecule of the Tricarboxylic Acid Cycle. As the carbon is oxidized, the massive release of energy is successfully conserved. Instead of being lost as heat, this energy is trapped by forming a new, high-energy thioester bond between the remaining four-carbon skeleton and Coenzyme A. This reaction produces the potent intermediate succinyl-CoA.
Simultaneously, this critical step generates the second NADH molecule. By capturing these high-energy electrons, the cell continues to build its reserves of reducing equivalents for the respiratory chain. The formation of succinyl-CoA marks a major metabolic shift within the Tricarboxylic Acid Cycle. Because two carbons entered the pathway as acetyl-CoA, and two carbons have now departed as carbon dioxide, the net carbon balance is effectively zero. The original four-carbon length of the carrier molecule has been fully reestablished.
From this point forward in the cycle, the biochemical focus shifts completely away from carbon loss. The subsequent steps will be entirely dedicated to extracting the remaining energy and rearranging the four-carbon chain to regenerate the starting molecule, oxaloacetate. For students, mastering the complex cofactor requirements of the 2-oxoglutarate dehydrogenase complex is vital, as vitamin deficiencies significantly affect these components. Without adequate thiamine or niacin, this massive multienzyme machine fails, halting cellular energy production catastrophically.
Slide 7: Direct Energy Harvesting in the Tricarboxylic Acid Cycle
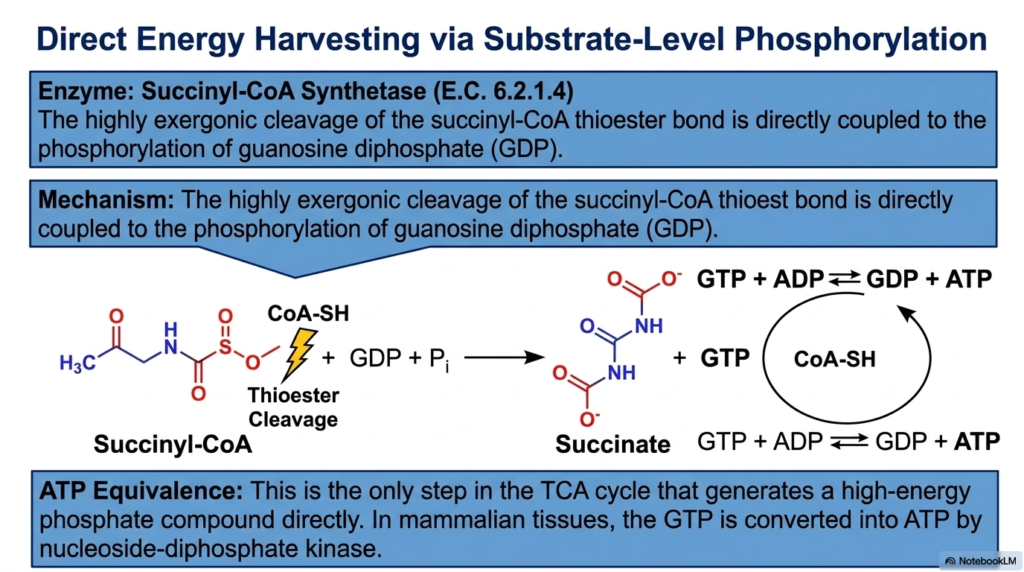
Up to this point in the Tricarboxylic Acid Cycle, energy extraction has relied entirely on capturing electrons through NADH. However, the seventh step introduces a distinct mechanism known as substrate-level phosphorylation. This fascinating reaction is catalyzed by the enzyme succinyl-CoA synthetase. The substrate, succinyl-CoA, contains a high-energy thioester bond that was carefully forged during the previous complex multienzyme step. Succinyl-CoA synthetase is specifically designed to harness the incredible energy locked within that specific sulfur-carbon linkage.
The mechanism begins with the highly exergonic cleavage of the succinyl-CoA thioester bond. As the Coenzyme A molecule is released, the massive drop in free energy is not wasted. Instead, the enzyme directly couples the release of this energy to the phosphorylation of guanosine diphosphate (GDP). An inorganic phosphate is forcefully added to GDP, resulting in the creation of guanosine triphosphate (GTP). The four-carbon skeleton is simultaneously converted to succinate, propelling the intermediate further along the Tricarboxylic Acid Cycle.
The generation of GTP is a landmark event within the Tricarboxylic Acid Cycle. This step is the only point in the pathway where a high-energy phosphate group is generated directly by enzymatic action, bypassing the electron transport chain entirely. The energy contained within GTP is completely equivalent to that of ATP. In most mammalian tissues, this newly synthesized GTP is quickly and seamlessly converted into ATP by a secondary enzyme known as nucleoside-diphosphate kinase.
This direct energy harvest acts as a critical fail-safe for the cell. Even if the downstream electron transport chain is temporarily backed up or oxygen-limited, this specific step guarantees a small but vital baseline production of high-energy phosphates. For biochemistry students, distinguishing between substrate-level phosphorylation and oxidative phosphorylation is a fundamental learning objective. Succinyl-CoA synthetase perfectly illustrates how cells link the thermodynamic breakdown of carbon skeletons directly to the chemical synthesis of usable energy currency.
Slide 8: The Respiratory Chain Link of the Tricarboxylic Acid Cycle
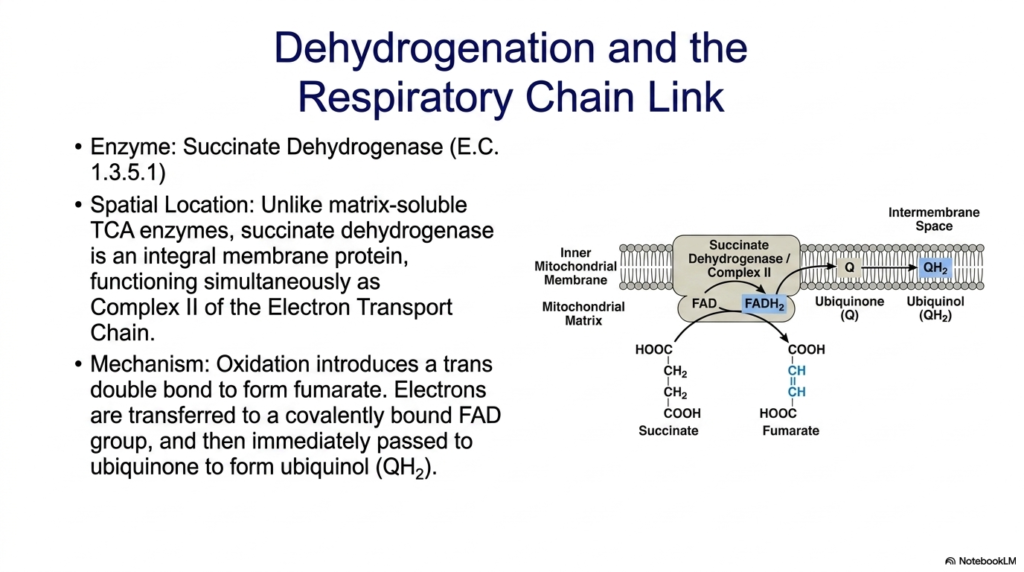
The conversion of succinate to fumarate introduces one of the most unique enzymes in the entire Tricarboxylic Acid Cycle. Succinate dehydrogenase stands completely apart from its biochemical peers. While all other enzymes of this pathway float freely within the soluble environment of the mitochondrial matrix, succinate dehydrogenase is an integral membrane protein. It is deeply embedded in the inner mitochondrial membrane, physically anchoring a portion of the metabolic cycle to the organelle’s structural boundaries.
This unique spatial location exists because succinate dehydrogenase possesses a dual identity. Not only is it an essential step in the Tricarboxylic Acid Cycle, but it simultaneously functions as Complex II of the Electron Transport Chain. The mechanism involves the targeted oxidation of succinate, which removes two hydrogen atoms and introduces a trans double bond, forming fumarate. Because the energy drop in this specific oxidation is relatively small, the electrons cannot be accepted by NAD+.
Instead, the extracted electrons are transferred to a covalently bound FAD prosthetic group embedded deep within the enzyme, reducing it to FADH2. Because succinate dehydrogenase physically bridges the Tricarboxylic Acid Cycle and the respiratory chain, these newly captured electrons do not have to travel far. The FADH2 immediately passes its electrons directly to ubiquinone, a lipid-soluble molecule residing in the membrane, converting it into ubiquinol. This seamless handoff efficiently funnels energy straight into the membrane-bound proton pumps.
For medical students, understanding the enzyme’s physical architecture is paramount to grasping cellular respiration. By acting as a literal bridge between soluble carbon metabolism and membrane-bound electron transport, Complex II eliminates the need for intermediate diffusion. This slide perfectly illustrates the elegant efficiency of cellular design. The enzyme’s embedded nature ensures that the energy extracted from the remaining carbon skeleton is immediately integrated into the machinery that ultimately synthesizes bulk ATP for the organism.
Slide 9: Hydration and Regeneration in the Tricarboxylic Acid Cycle
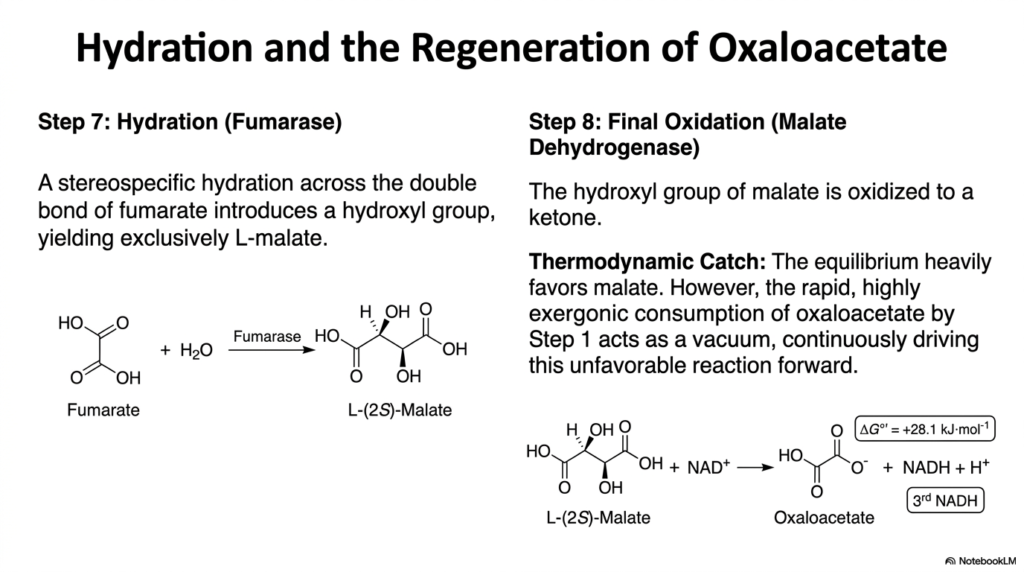
As the cycle nears completion, the four-carbon intermediate must undergo extensive modification to regenerate the original starting material. Following the creation of fumarate, the Tricarboxylic Acid Cycle employs the enzyme fumarase. This enzyme catalyzes a stereospecific hydration reaction directly across the newly formed trans double bond of fumarate. By introducing a water molecule, fumarase successfully adds a hydroxyl group to the carbon skeleton. The extreme precision of the enzyme’s active site ensures the reaction yields the L-isomer of malate exclusively.
The production of L-malate sets the stage for the final biochemical hurdle of the Tricarboxylic Acid Cycle. The eighth and final step is carried out by the enzyme malate dehydrogenase. This enzyme performs a final oxidation, converting the newly attached secondary hydroxyl group of malate into a ketone. This critical oxidation strips the final high-energy electrons from the carbon skeleton, generating the third and final NADH molecule while regenerating the starting molecule, oxaloacetate.
However, this final reaction possesses a massive thermodynamic catch. The oxidation of malate to oxaloacetate is highly endergonic under standard conditions, meaning the chemical equilibrium heavily favors the reverse reaction toward malate. In isolation, this reaction would essentially run backward, halting the Tricarboxylic Acid Cycle. Nature overcomes this massive energy barrier through the principle of metabolic coupling. The rapid, highly exergonic consumption of oxaloacetate by citrate synthase in step one acts as a biochemical vacuum.
By immediately destroying the oxaloacetate product, the first step continuously pulls the unfavorable malate dehydrogenase reaction forward. This elegant interplay demonstrates Le Chatelier’s principle operating flawlessly within a living cell. The continuous regeneration of oxaloacetate ensures the pathway remains a true, uninterrupted cycle rather than a linear dead end. By understanding this thermodynamic relationship, students can deeply appreciate how endergonic biological reactions are successfully driven by the strategic placement of highly exergonic reactions later down the line.
Slide 10: Stoichiometric Yield of the Tricarboxylic Acid Cycle
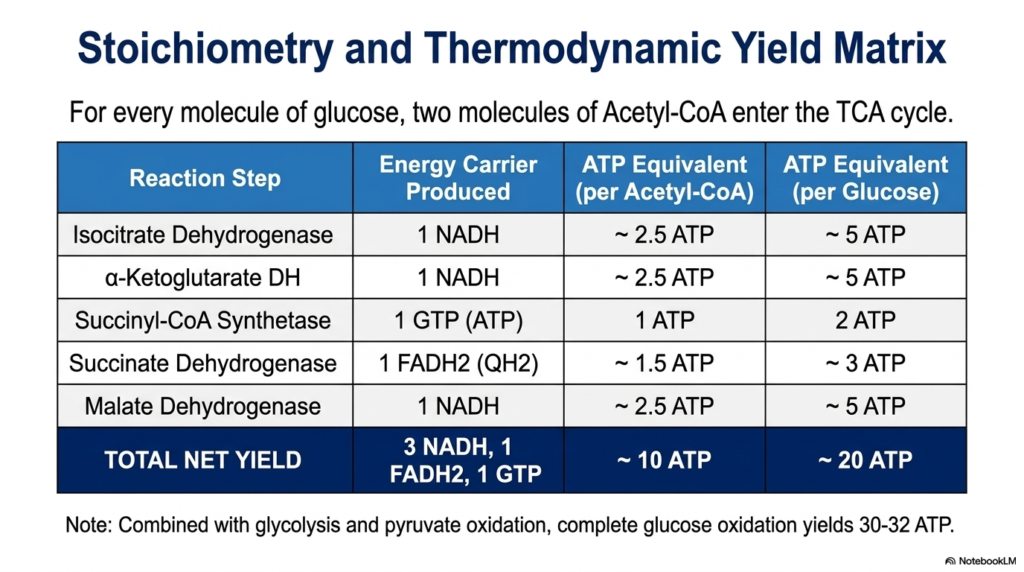
To truly appreciate the cell’s metabolic power, one must analyze the overall thermodynamic yield. This slide breaks down the strict stoichiometry of the Tricarboxylic Acid Cycle. For every single molecule of glucose that enters cellular respiration, two distinct molecules of acetyl-CoA are generated during prior metabolic stages. Therefore, to process one glucose molecule entirely, the metabolic cycle must complete two full revolutions. Calculating the specific energy carriers produced during these revolutions reveals the incredible efficiency of biochemical energy extraction.
During a single turn of the Tricarboxylic Acid Cycle, the sequential enzyme reactions produce a highly specific output. The oxidative decarboxylations by isocitrate dehydrogenase and alpha-ketoglutarate dehydrogenase each yield one NADH. The final oxidation by malate dehydrogenase generates a third NADH. The membrane-bound succinate dehydrogenase creates one FADH2. Finally, the substrate-level phosphorylation by succinyl-CoA synthetase creates one GTP, which is energetically identical to ATP. This yields a net of three NADH, one FADH2, and one GTP per acetyl-CoA.
Translating these molecular carriers into usable energy requires understanding their downstream value. When processed by the electron transport chain, each NADH generates approximately 2.5 ATP, while each FADH2 contributes roughly 1.5 ATP. When the direct GTP is included, a single turn of the Tricarboxylic Acid Cycle yields an equivalent of 10 ATP molecules. Because one glucose molecule drives two complete turns, the cycle ultimately generates 20 ATP equivalents per glucose, representing the vast majority of biological energy production.
By viewing this stoichiometric matrix, students can quantitatively trace how the potential energy hidden in chemical bonds is methodically harvested. When these 20 ATP equivalents are combined with the energy captured during prior glycolysis and pyruvate oxidation, the complete oxidation of glucose yields a staggering 30 to 32 ATP. This massive thermodynamic yield explains why aerobic organisms heavily rely on mitochondrial metabolism. The careful math of energy conversion proves that the cycle is the undisputed powerhouse mechanism of aerobic life.
Slide 11: Allosteric Control of the Tricarboxylic Acid Cycle
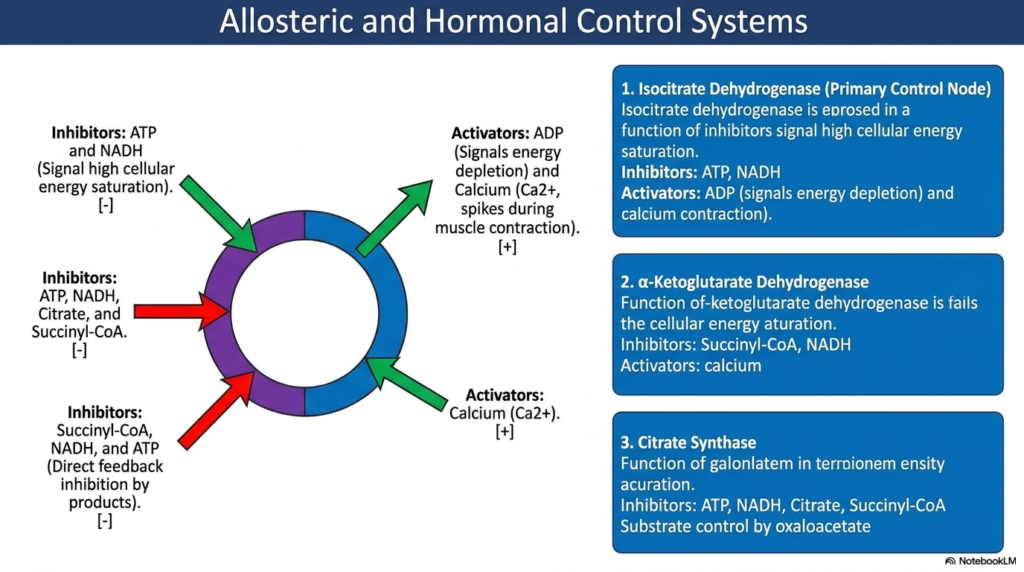
A biochemical pathway as powerful as the Tricarboxylic Acid Cycle must be tightly regulated to prevent cellular disaster. If the cycle ran continuously without restriction, it would quickly burn through all available cellular fuel, wasting precious resources. Therefore, the cell employs complex allosteric and hormonal control systems to monitor energy demands. The regulatory mechanisms primarily target the highly exergonic, irreversible steps of the pathway. By throttling these critical checkpoints, the cell can instantly accelerate or hit the brakes on energy production.
The primary control node of the Tricarboxylic Acid Cycle is isocitrate dehydrogenase. This enzyme acts as the main metabolic speedometer. It is strongly inhibited by high ATP and NADH concentrations, which physically signal a state of high cellular energy saturation. Conversely, when cellular energy is rapidly depleted, rising ADP levels act as strong allosteric activators, forcing the enzyme to accelerate. In muscle tissue, sudden spikes in calcium ions during muscle contraction also strongly activate this node to meet immediate kinetic demands.
The second crucial checkpoint within the Tricarboxylic Acid Cycle involves the alpha-ketoglutarate dehydrogenase complex. Much like the previous step, this enzyme complex fails to advance rapidly when cellular energy is already saturated. It experiences massive direct feedback inhibition from its own immediate products, shutting down when levels of succinyl-CoA and NADH begin to heavily accumulate. As with isocitrate dehydrogenase, this complex is strongly stimulated by the sudden influx of calcium ions, ensuring that energy production ramps up during strenuous physical activity.
Finally, citrate synthase is strongly allosterically regulated. This initial committing enzyme is inhibited by ATP, NADH, citrate, and succinyl-CoA. The overarching theme of this regulatory system is undeniable efficiency. High-energy states cause widespread inhibition, thereby diverting precursor molecules toward anabolic storage. Low-energy states relieve this inhibition, driving the catastrophic breakdown of carbon skeletons. For the medical student, mastering these allosteric triggers provides the foundation for understanding metabolic diseases, exercise physiology, and the dynamic adaptability of human cells.
Slide 12: Maintaining the Pool: Anaplerosis in the Tricarboxylic Acid Cycle
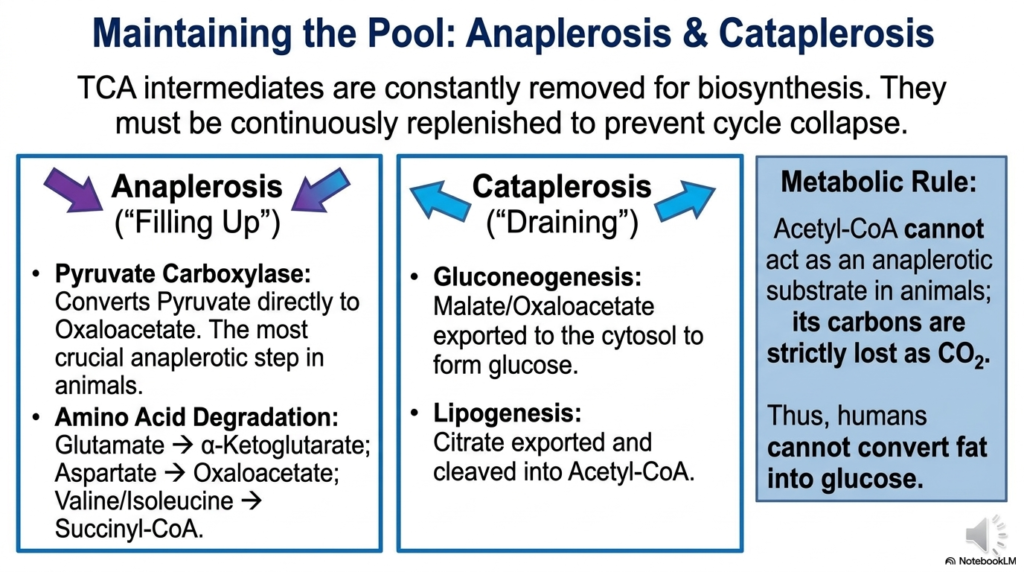
The amphibolic nature of the Tricarboxylic Acid Cycle presents a massive logistical challenge for the living cell. Because metabolic intermediates are constantly being pulled away to build amino acids, fatty acids, and heme groups, the overall concentration of these molecules threatens to drop dangerously low. This draining process is known as cataplerosis. If too many intermediates are siphoned off without replacement, the entire metabolic cycle would physically collapse, halting energy production entirely and leading to rapid cellular death.
To prevent this fatal collapse, the cell utilizes a series of continuous replenishing reactions known as anaplerosis, which essentially means “filling up.” The most critical anaplerotic step in animal cells is catalyzed by the enzyme pyruvate carboxylase. This enzyme bypasses the standard pathway for acetyl-CoA formation, directly converting pyruvate into oxaloacetate. By pumping fresh oxaloacetate directly into the mitochondrion, the cell guarantees the Tricarboxylic Acid Cycle has enough starting material to bind incoming acetyl groups and keep the massive thermodynamic engine turning smoothly.
Amino acid degradation provides another major avenue for anaplerosis. For example, glutamate can be converted directly into alpha-ketoglutarate, while aspartate can be converted into oxaloacetate. Valine and isoleucine can be funneled in as succinyl-CoA. This massive influx of carbon skeletons from protein breakdown ensures the Tricarboxylic Acid Cycle remains fully stocked with intermediates, even during periods of extreme starvation or heavy metabolic demand. The delicate balance between draining and filling dictates the overall health and speed of the pathway.
A fundamental metabolic rule arises from this biochemical architecture. Acetyl-CoA cannot serve as an anaplerotic substrate in animals because its two carbons are strictly lost as carbon dioxide during the cycle. Consequently, animals lack the necessary biochemical enzymes to achieve net synthesis of glucose from fatty acids. The strict division between anaplerotic and cataplerotic flow perfectly illustrates the highly regulated boundaries of mammalian metabolism. Understanding these rules is crucial for interpreting complex physiological states such as diabetic ketoacidosis or starvation.
Slide 13: Clinical Correlations of the Tricarboxylic Acid Cycle
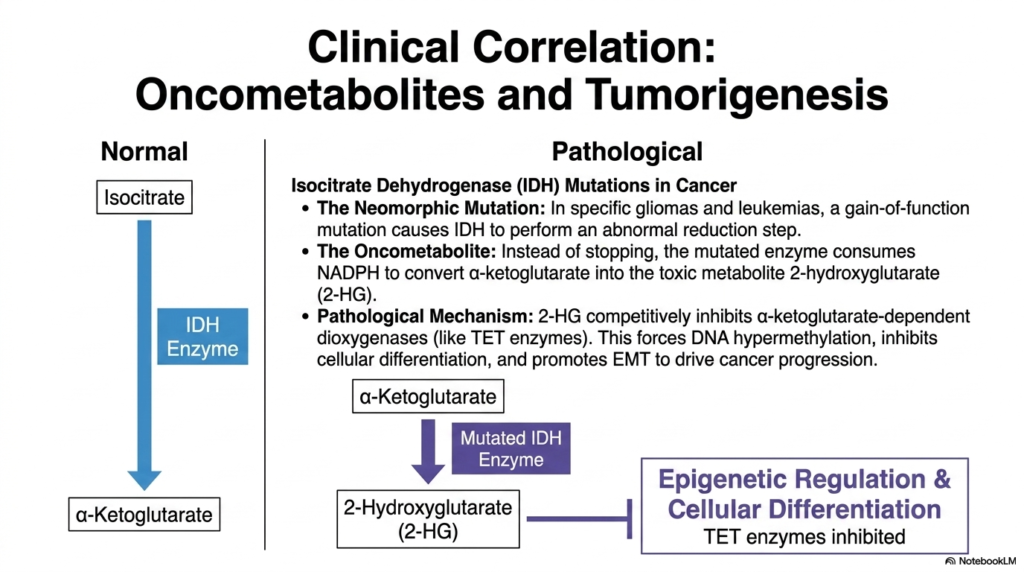
The profound importance of the Tricarboxylic Acid Cycle extends far beyond standard physiology; it plays a highly critical role in modern oncology and disease pathology. When the genes encoding these core metabolic enzymes undergo specific mutations, the resulting biochemical dysfunction can directly drive tumorigenesis. A perfect example of this devastating clinical correlation is found in mutations affecting isocitrate dehydrogenase (IDH). While the enzyme normally functions to produce alpha-ketoglutarate, mutated versions of this enzyme frequently appear in specific human gliomas and aggressive leukemias.
In these cancers, the mutation does not merely break the enzyme; it creates a neomorphic, or gain-of-function, mutation. Instead of stopping at the normal endpoint, the mutated enzyme consumes precious cellular NADPH to carry out an abnormal, catastrophic reduction step. This pathological reaction forcefully converts alpha-ketoglutarate into a highly toxic, rogue molecule known as 2-hydroxyglutarate (2-HG). Because 2-HG is generated entirely by a malfunctioning enzyme of the Tricarboxylic Acid Cycle, scientists classify this dangerous molecule as a primary oncometabolite.
The accumulation of this oncometabolite causes widespread metabolic chaos. 2-HG is structurally similar to alpha-ketoglutarate, allowing it to act as a powerful competitive inhibitor. It specifically blocks alpha-ketoglutarate-dependent dioxygenases, including the critical TET enzymes responsible for proper epigenetic regulation. By shutting down these epigenetic regulators, the mutated Tricarboxylic Acid Cycle enzyme forces massive DNA hypermethylation. This chemical lockdown completely inhibits normal cellular differentiation and traps the cell in a highly proliferative, immature state, thereby directly promoting dangerous cancer progression.
This fascinating clinical correlation bridges the gap between basic biochemistry and advanced pathology. It demonstrates how a single error in a core metabolic enzyme can drastically reshape the epigenetic landscape of an entire cell. For medical students, realizing that intermediate metabolites can directly control gene expression shifts the paradigm of oncology. The oncometabolite concept proves that metabolic pathways are not just passive energy generators, but active regulators of cellular identity, cellular differentiation, and potentially lethal pathological transformations.
Please read our Content Disclaimer Statement.
Check out our social media channels: