65. Bacterial Cell Structure: Exploring E. coli and Cytoplasmic Functions
Imagine a bustling metropolis operating at maximum efficiency without a single central traffic controller; this is the reality of the cellular interior. The core purpose of this slide deck is to illuminate the foundational principles of cellular life, breaking down complex biochemical environments into digestible concepts. Understanding Bacterial Cell Structure provides an essential framework for understanding how life functions at the molecular scale. Medical students and future biochemists must grasp these foundational dynamics to understand disease mechanisms and pharmacological interventions.
Slide 1: Bacterial Cell Structure: The Cytoplasm – Why Intracellular Dynamics Matter

The first slide introduces the vibrant and densely packed nature of the cellular interior. It visually establishes that Bacterial Cell Structure is far from static or empty. Instead, it is a highly dynamic, organized space defined by its unique composition and rapid biochemical functions. The visual representation highlights various geometric shapes that symbolize the diverse array of macromolecules, metabolites, and ions that fill the cytoplasmic space. This foundational image prepares the observer for a detailed exploration of the microorganism’s physical and chemical reality.
At the heart of Bacterial Cell Structure lies the cytoplasm, a matrix that orchestrates every critical life process. It is the primary arena where vital molecules are synthesized, maintaining the delicate balance of life. The composition of this space dictates its dynamics, influencing how proteins fold, how enzymes locate their substrates, and how genetic information translates into functional machinery. The sheer density of this environment forces a reevaluation of traditional biochemical models, which often study enzymes in dilute solutions rather than their natural, crowded habitat.
Ultimately, mastering the concepts presented in this deck is crucial for anyone entering the medical or biological sciences. The cytoplasm is not merely a background substance; it is the active participant in cellular biochemistry. By studying these dynamics, students unlock the secrets of metabolic regulation, antibiotic targeting, and cellular adaptation. The journey through this deck will reveal that the apparent chaos of the cellular interior is, in fact, a marvel of evolutionary engineering, perfectly tuned for survival and rapid reproduction.
Slide 2: Bacterial Cell Structure: The Standard Model – Why E. coli Defines Molecular Biology
When architects design skyscrapers, they rely on universal physical laws; similarly, biologists rely on a foundational blueprint to understand complex life. The core purpose of this slide deck is to deconstruct this blueprint, providing a rigorous biochemical analysis of the cellular interior. By examining Bacterial Cell Structure through the lens of the model organism Escherichia coli, researchers establish a baseline for all molecular biology. This framework is crucial for students, as it translates abstract biochemical theories into tangible, measurable physical spaces within a living organism.
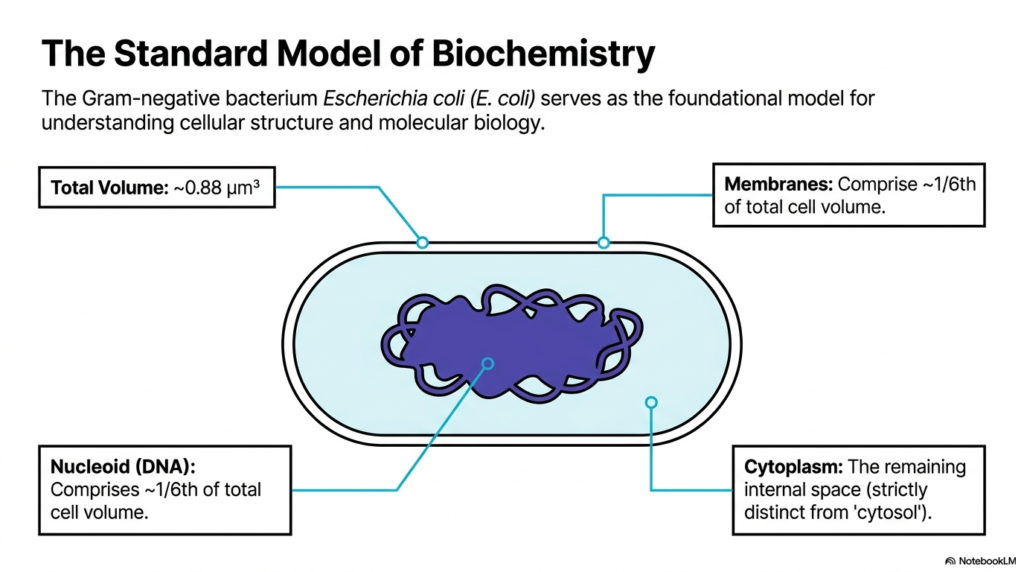
This slide outlines the precise volumetric proportions of the standard E. coli model, which boasts a total volume of approximately 0.88 cubic micrometers. It divides the cell into three primary distinct regions: the membranes, the nucleoid containing the DNA, and the cytoplasm. Interestingly, the membranes and the nucleoid each account for roughly one-sixth of the total cell volume. The cytoplasm occupies the remaining internal space. A crucial distinction is made here in Bacterial Cell Structure: the term cytoplasm denotes the entire crowded matrix, which is strictly distinct from the conceptual liquid-only cytosol.
Understanding these spatial boundaries is vital to grasping how biochemical reactions are compartmentalized and organized in the absence of membrane-bound organelles. In the context of Bacterial Cell Structure, the absence of internal membrane compartments means the nucleoid and cytoplasm must coexist in the same general space, separated primarily by biophysical properties rather than by physical barriers. This close proximity allows for simultaneous transcription and translation, a hallmark of prokaryotic efficiency. The specified volumes determine the concentrations of all proteins and metabolites, directly affecting the thermodynamic favorability of essential survival pathways.
For medical students, recognizing these proportions helps explain why certain antibiotics that target the bacterial membrane or the protein synthesis machinery are so effective. The tight spatial constraints mean any disruption to the volume or macromolecular crowding can be catastrophic for the organism. The E. coli model serves as an ideal educational tool, offering a simplified yet highly accurate representation of life’s minimal spatial requirements. It provides the necessary quantitative context to appreciate the sheer density of biochemical reactions explored in subsequent slides.
Slide 3: Bacterial Cell Structure: Macromolecular Composition – How Cells Balance Water and Dry Mass

Consider a complex recipe where the exact ratio of ingredients determines success or failure; cellular biology operates on a similar principle. The core purpose of this slide deck is to reveal the exact molecular inventory that makes life possible. Understanding Bacterial Cell Structure requires a deep dive into its macromolecular composition. Medical students must appreciate these exact ratios, as altering the delicate balance between water and dry mass disrupts every known pharmacological and physiological cellular process.
This slide breaks down the gross chemical composition of the cell, illustrating that water constitutes a massive seventy percent of the cellular environment. Despite being a single-molecule type, water is critical for Bacterial Cell Structure, serving as the universal solvent for life. The remaining 30% comprises the dry mass, a highly diverse mixture of macromolecules. This stark contrast between a single solvent and a complex matrix defines the physical reality of the cytoplasm.
Within that thirty percent dry mass, proteins dominate, making up seventeen percent of the total volume with approximately three thousand distinct types. RNA follows at eight percent, while DNA, lipids, and sugars each constitute one percent of the mass. A vital aspect of Bacterial Cell Structure is the presence of inorganic ions and small molecules, which together represent the final 1%. These diverse categories highlight the immense biochemical variety required to maintain homeostasis, reproduce, and generate cellular energy.
For future biochemists, memorizing this macromolecular composition is less about rote numbers and more about understanding physical constraints. The limited space means that every protein, transcript, and lipid must be precisely regulated. The ratios presented here prove that the cell is a finely tuned engine. Any deviation in these percentages, whether caused by viral infection or environmental stress, fundamentally compromises the integrity and survival of the organism.
Slide 4: Bacterial Cell Structure: Protein Density – Why Scale Dictates Cellular Function
Imagine attempting to fit an entire automated car factory into a space smaller than a speck of dust. The core purpose of this slide deck is to illustrate exactly how biological systems achieve this incredible feat of miniaturization. When analyzing Bacterial Cell Structure, the concept of scale becomes paramount. For students navigating molecular biology, understanding protein density is the key to unlocking how microscopic organisms exert such profound impacts on human health and global ecosystems.
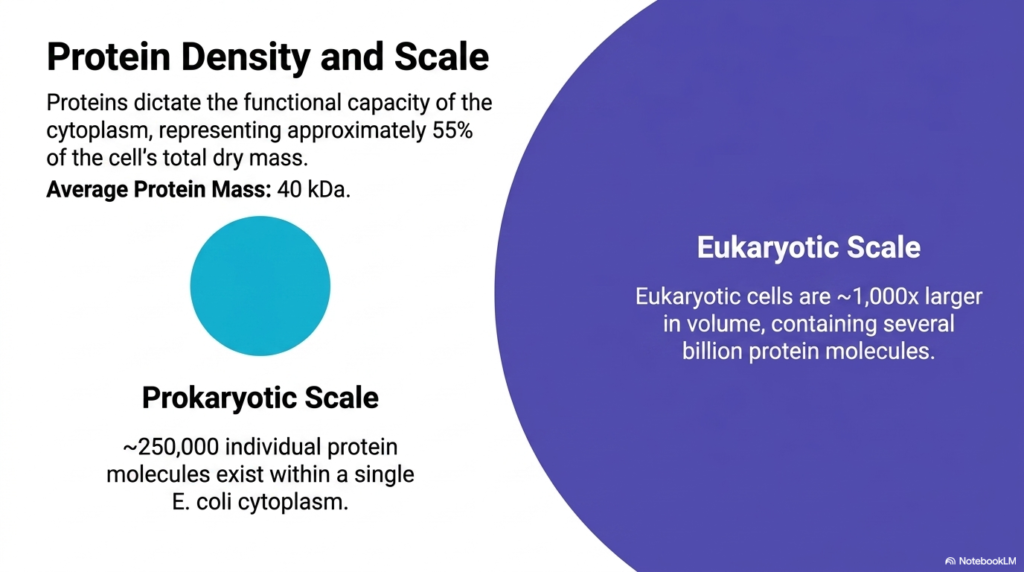
The functional capacity of the cytoplasm is entirely dictated by its protein density, which represents roughly 55% of the cell’s total dry mass. With an average protein mass of forty kilodaltons, these macromolecules are the primary workforce of Bacterial Cell Structure. The slide visualizes this immense density, emphasizing that proteins are not floating freely in an empty void. Instead, they are packed so tightly that they constantly interact, fold, and function in a highly crowded, fiercely competitive microscopic environment.
A dramatic scale comparison is presented to contextualize this density. Within a single prokaryotic organism like E. coli, approximately 250,000 individual protein molecules exist simultaneously. In stark contrast, eukaryotic cells are roughly 1,000 times larger in volume and contain several billion protein molecules. This comparative anatomy of Bacterial Cell Structure demonstrates that while the absolute numbers differ, the fundamental physical principle of extreme macromolecular crowding remains a universal constant across all domains of life.
Understanding this immense scale is critical for grasping drug kinetics and enzyme kinetics within the cell. When a pharmaceutical agent enters this space, it must navigate a dense forest of a quarter-million proteins to find its specific target. The visual representation of protein density provided here shatters the illusion of the cell as a simple bag of water, replacing it with the reality of an incredibly sophisticated, densely woven matrix of active biochemical machinery.
Slide 5: Bacterial Cell Structure: The Micro-Environment – How Molecular Proportions Shape Life
Visualizing the invisible requires a radical shift in perspective, akin to mapping a galaxy by looking at individual grains of sand. The core purpose of this slide deck is to provide that necessary magnification, making the abstract world of biochemistry tangible. To truly comprehend Bacterial Cell Structure, students must shrink their conceptual framework to the nanometer scale. This shift is essential for medical professionals attempting to understand how single molecules can trigger massive systemic changes within a patient.
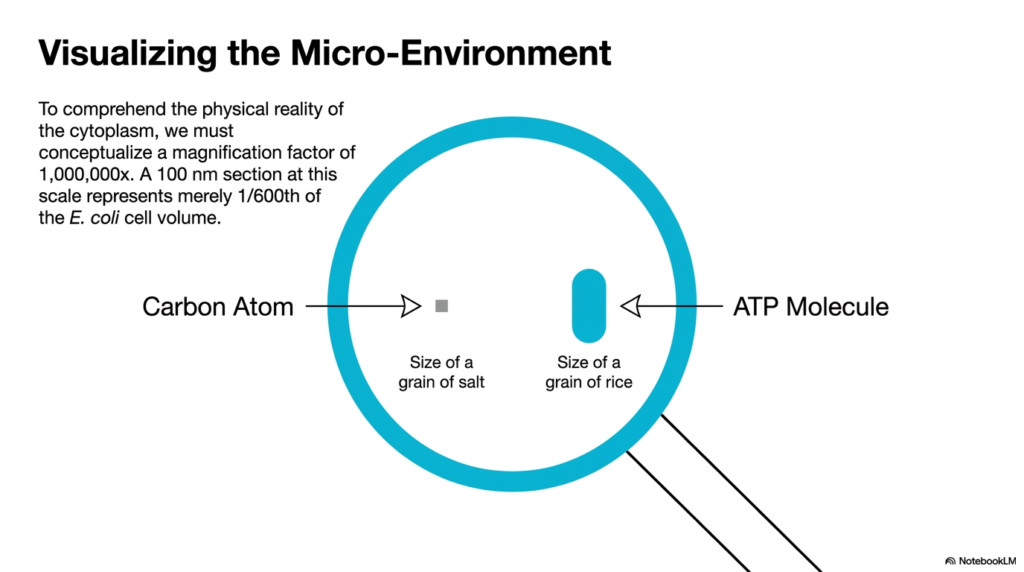
This slide challenges the observer to conceptualize a magnification factor of 1,000,000× to grasp the physical reality of the cytoplasm. At this extreme scale, a 100-nanometer section represents only one-six-hundredth of the total E. coli cell volume. Within this context of Bacterial Cell Structure, the relative sizes of fundamental molecules are mapped out using relatable physical objects. This dramatic scaling bridges the gap between complex biochemical equations and actual physical space within the organism.
By establishing these visual anchors, the slide makes molecular proportions easier to understand. For instance, a single carbon atom is equated to the size of a grain of salt, while an ATP molecule is compared to a grain of rice. This analogy emphasizes a critical rule of Bacterial Cell Structure: small molecules are physically substantial in the microscopic realm. The sheer size of an ATP molecule relative to a carbon atom dictates how energy is physically transported and utilized.
For a biochemistry student, adopting this micro-environmental perspective is highly transformative. It transitions the study of metabolism from a theoretical exercise into a mechanical reality. Knowing the relative physical size of metabolites helps explain steric hindrance, active-site specificity, and molecular transport mechanisms. By visualizing these molecules as physical objects occupying defined space, learners can better predict how enzymes interact with their substrates in the crowded environment of the cell.
Slide 6: Bacterial Cell Structure: The Crowded Paradigm – Why Physical Density Governs Thermodynamics

Navigating a packed subway train during rush hour offers a perfect analogy for the chaotic energy of the cellular interior. The core purpose of this slide deck is to dispel the outdated myth of the cell as a liquid-filled balloon and replace it with accurate biophysical models. Examining Bacterial Cell Structure reveals a tightly compartmentalized, fiercely crowded arena. For biochemistry students, embracing this crowded paradigm is necessary to understand why reactions behave differently in a living organism than in a glass test tube.
The slide explicitly states that the cytoplasm is tightly packed with macromolecules and is definitely not an empty, aqueous swimming pool. This establishes the crowded-cytoplasm paradigm, a foundational concept for understanding modern Bacterial Cell Structure. Organic molecules are separated by only a few individual water molecules, leaving almost no empty space. The visual representation displays a chaotic, interlocking puzzle of proteins and nucleic acids, highlighting the minimal physical separation between distinct biological machines working in tandem.
This immense physical density dramatically alters the rules of chemistry. In the realm of Bacterial Cell Structure, crowding significantly affects thermodynamic properties, protein folding pathways, and molecular diffusion rates. Molecules cannot move freely; they must constantly push past one another. This physical constraint means that the effective concentration of molecules is much higher than their absolute concentration would suggest in a dilute solution. Consequently, biochemical reactions are driven with a thermodynamic efficiency that is impossible to replicate under standard in vitro laboratory conditions.
Recognizing the crowded paradigm forces a massive shift in medical and biological research. Drug design must account for this density, as therapeutic molecules must navigate this incredibly viscous environment to reach their targets. The slide perfectly illustrates that life does not occur in a vacuum, nor in a simple liquid. It thrives in a highly restricted, physically demanding matrix that directly dictates biological function and survival.
Slide 7: Bacterial Cell Structure: Biosynthesis Density – How Translation Machinery Packs the Cytoplasm
Imagine attempting to fit an entire printing press, with all its moving parts, into a space no larger than a thimble. The core purpose of this slide deck is to quantify exactly how much machinery fits within the cell’s microscopic dimensions. Delving into Bacterial Cell Structure reveals a breathtaking concentration of biosynthetic equipment. Medical students must grasp this density to understand how quickly pathogens can multiply, produce toxins, and adapt to host environments through rapid protein synthesis.
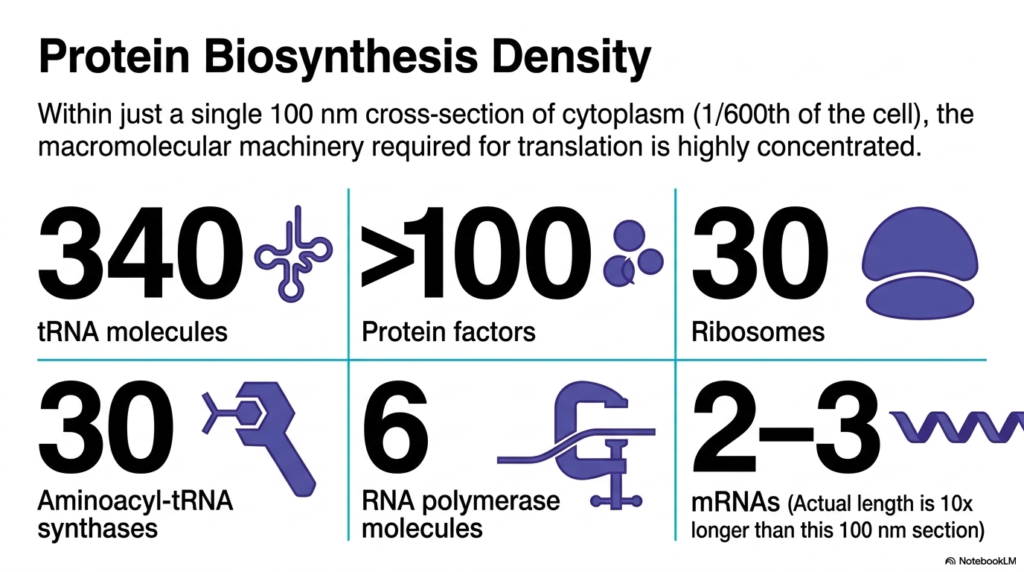
This slide isolates a mere one-hundred-nanometer cross-section of the cytoplasm, representing just one-six-hundredth of the cell, to catalog the machinery of translation. It reveals that Bacterial Cell Structure can accommodate a staggering concentration of macromolecules within this tiny area. The inventory includes three hundred and forty transfer RNA molecules, over one hundred protein factors, and thirty massive ribosomes. This numerical breakdown provides a stark, quantitative look at the sheer physical volume occupied by the protein biosynthesis apparatus.
Furthermore, the slide details the presence of thirty aminoacyl-tRNA synthases, six RNA polymerase molecules, and two to three messenger RNAs. A crucial insight into Bacterial Cell Structure is that the actual length of the mRNA is ten times longer than the section itself, meaning these strands are tightly coiled and interwoven with other components. This extreme packing ensures that, the moment an mRNA is transcribed, ribosomes immediately bind to it, executing rapid, co-transcriptional translation that is essential for bacterial survival.
For students of biochemistry, these numbers are not mere trivia; they represent the kinetic reality of genetic expression. The extreme density of this machinery means that translation is a highly localized, hyper-efficient process. Substrates do not need to travel far to find their required enzymes. This slide brilliantly demonstrates that the cell is a masterclass in spatial economy, cramming an astonishing amount of functional hardware into a fraction of its available volume.
Slide 8: Bacterial Cell Structure: Enzyme Density – Why Metabolic Machinery Requires Proximity
Consider a highly efficient assembly line where workers stand shoulder to shoulder, instantly passing products down the line without taking a single step. The core purpose of this slide deck is to map out this precise metabolic architecture within the cellular matrix. Exploring Bacterial Cell Structure uncovers the brilliant spatial organization of metabolic enzymes. Grasping this concept allows future clinicians to understand metabolic disorders and the precise mechanisms by which specific inhibitors can stall the entire engine of a living cell.
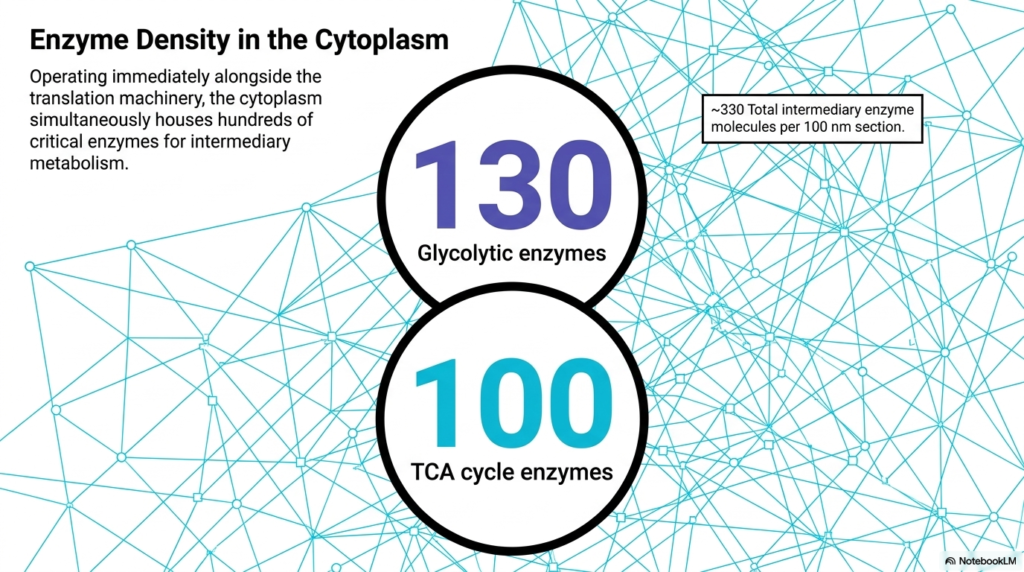
Operating immediately alongside the massive translation machinery, the cytoplasm simultaneously houses hundreds of critical enzymes dedicated to intermediary metabolism. This slide emphasizes that Bacterial Cell Structure is not defined by physical walls but rather by intermingled functional networks. Within the same 100-nanometer section previously discussed, approximately 330 intermediary enzyme molecules are present. This intense crowding ensures that multiple different metabolic pathways are running concurrently in the exact same physical space.
The slide breaks down the total enzyme count, highlighting 130 glycolytic enzymes and 100 tricarboxylic acid cycle enzymes. This specific feature of Bacterial Cell Structure illustrates the necessity of extreme proximity for metabolic efficiency. Because these enzymes are packed so tightly together, the product of one reaction instantly becomes the substrate for the adjacent enzyme. This physical closeness minimizes transit time and prevents highly reactive intermediate molecules from diffusing away or degrading before they can be utilized.
Understanding this enzyme density provides students with a profound appreciation for metabolic channeling. The cytoplasm acts as a solid-state reactor where enzymes are functionally, if not physically, linked. This visual representation clarifies why biochemical pathways operate with such astonishing speed and efficiency. It proves that the physical arrangement of the cell is just as important as the chemical equations themselves, highlighting the perfect marriage between biological form and life-sustaining thermodynamic function.
Slide 9: Bacterial Cell Structure: Micro-Solutes – How Small Molecules Power Massive Reactions
If macromolecules are the massive bricks that construct a biological building, then small molecules and ions are the essential mortar that holds it all together. The core purpose of this slide deck is to highlight every component of the cellular interior, leaving no molecule unaccounted for. When studying Bacterial Cell Structure, one must look beyond the giant proteins to see the dense matrix of micro-solutes. Students must recognize these tiny players, as they directly control osmotic pressure, nerve conduction, and enzyme catalysis.
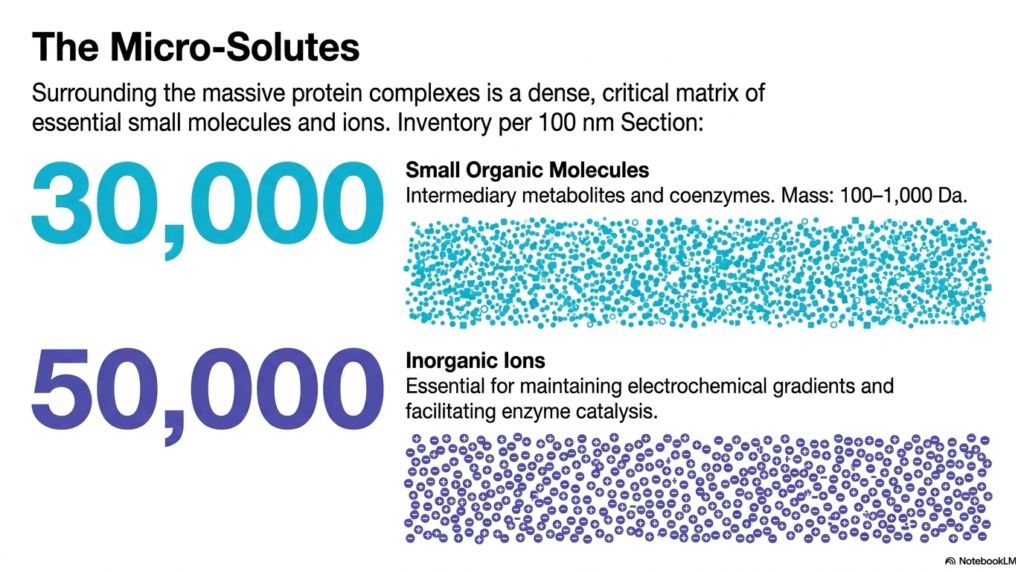
Surrounding the massive protein complexes is a dense, critical matrix of essential small molecules and inorganic ions. The slide inventories this hidden world within Bacterial Cell Structure, revealing 30,000 small organic molecules per 100-nanometer section. These include intermediary metabolites and coenzymes ranging from 100 to 1,000 Daltons in mass. Despite their small individual size, their collective presence creates a highly concentrated chemical soup that fills every conceivable gap between the larger cellular machinery.
Additionally, the slide accounts for fifty thousand inorganic ions packed into this same tiny space. In the context of Bacterial Cell Structure, these ions are not merely passive spectators; they are actively essential for maintaining electrochemical gradients and facilitating enzyme catalysis. The visual depiction shows a massive swarm of tiny dots and charges, illustrating how these micro-solutes coat protein surfaces. They neutralize charges, stabilize folded structures, and act as the immediate chemical currency required for thousands of simultaneous reactions.
For the aspiring biochemist, recognizing the immense quantity of these micro-solutes bridges the gap between gross anatomy and quantum chemistry. The sheer number of these molecules guarantees that enzymes never have to wait for a cofactor or a substrate. This slide successfully shifts the focus to the unsung heroes of the cellular environment, demonstrating that the smallest physical components are absolutely vital to driving the massive energy reactions that sustain biological life.
Slide 10: Bacterial Cell Structure: Molecular Kinematics – Why Brownian Motion Drives Biochemistry
Imagine playing a game of pinball where the ball bounces off thousands of bumpers every second, yet always hits the exact target required to win. The core purpose of this slide deck is to reveal the statistical magic behind these seemingly random cellular movements. Analyzing Bacterial Cell Structure requires an understanding of how molecules physically navigate a crowded space. Medical students must learn these principles of molecular kinematics to predict how fast a drug can diffuse to its cellular target.
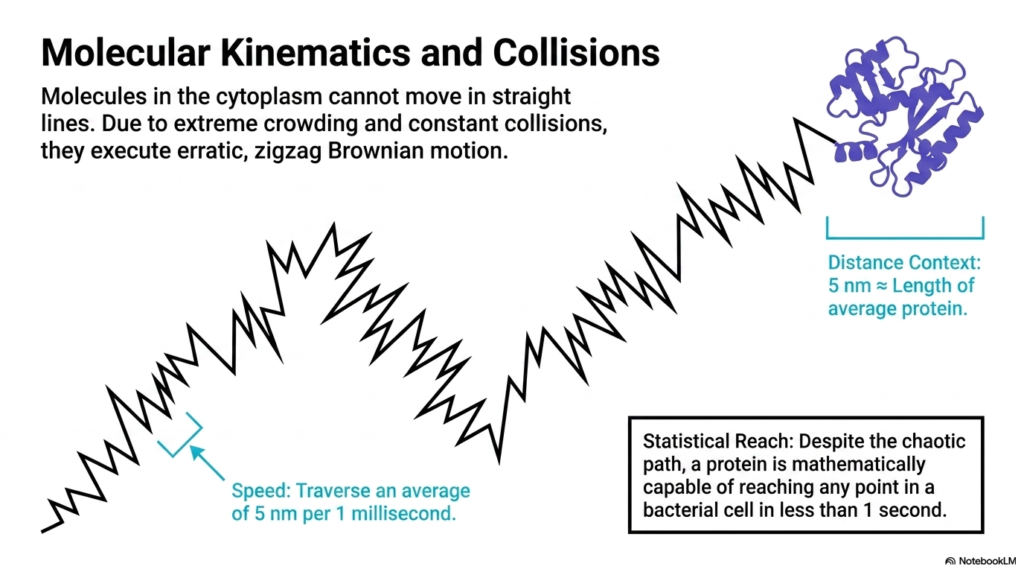
Due to extreme crowding and constant collisions, molecules in the cytoplasm cannot move in straight lines. This slide illustrates a foundational rule of Bacterial Cell Structure: particles execute erratic, zigzag paths known as Brownian motion. The visual depicts a jagged, unpredictable trajectory terminating at a folded protein. This visual perfectly captures the chaotic physical reality of the cellular environment, where thermal energy constantly violently throws molecules against one another, completely preventing any direct, unimpeded transit across the microscopic space.
Despite this erratic path, the slide details the astonishing speed of these molecules, which traverse an average of 5 nanometers per millisecond. The distance context provided shows that five nanometers is roughly the length of an average protein in Bacterial Cell Structure. Furthermore, the statistical reach is clearly defined: despite the chaotic, bouncing path, a single protein is mathematically capable of reaching any point in a bacterial cell in less than one second, driven entirely by this thermal kinetic energy.
Understanding this kinetic reality is crucial for biochemistry students, as it explains why cells do not require internal motorized transport for small-scale operations. Brownian motion, combined with the cell’s microscopic volume, ensures that every molecule will eventually collide with its intended target by sheer statistical probability. This slide brilliantly transforms what appears to be random, inefficient chaos into a highly reliable, incredibly fast mechanism for intracellular communication and the delivery of metabolic substrates.
Slide 11: Bacterial Cell Structure: Central Reaction Space – How the Cytoplasm Connects Cellular Pathways
Consider a massive transit hub connecting dozens of distinct railway lines, allowing millions of passengers to seamlessly transfer to their final destinations. The core purpose of this slide deck is to map out this central cellular junction, illustrating how diverse biochemical pathways interconnect. When viewed globally, Bacterial Cell Structure reveals the cytoplasm as the undisputed central reaction space. Future biochemists must understand this connectivity to see how a failure in one pathway cascades through the entire biological system.
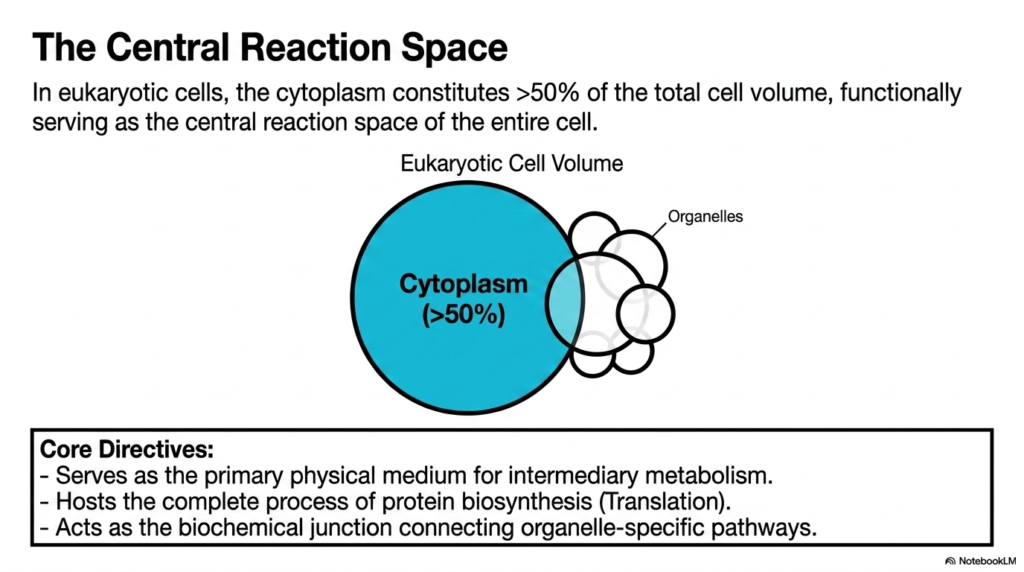
While eukaryotic cells possess numerous internal compartments, their cytoplasm still constitutes over fifty percent of the total cell volume. This slide highlights a universal truth applicable to Bacterial Cell Structure as well: the cytoplasm is the functional, central reaction space of the entire cell. The visual contrasts the massive cytoplasmic volume against smaller surrounding organelles. This comparison emphasizes that the cytoplasm is not just filler but the primary, dominant physical arena where the vast majority of biological life occurs.
The slide outlines the core directives of this critical space. It serves as the primary physical medium for intermediary metabolism and hosts the complete process of protein biosynthesis. More importantly for understanding broader biology beyond Bacterial Cell Structure, it acts as the biochemical junction connecting organelle-specific pathways. Metabolites must cross this vast, crowded space to travel between different specialized compartments. The cytoplasm orchestrates this massive flow of materials, ensuring that the nucleus, mitochondria, and other structures remain chemically unified.
For students, this concept elevates the cytoplasm from a simple structural component to a dynamic, regulatory organelle in its own right. It is the ultimate biological crossroad. By establishing the cytoplasm as the central reaction space, this slide prepares the observer to understand how energy, genetic information, and physical building blocks are seamlessly integrated. It underscores the incredible coordination required to maintain life, driven entirely by the physical properties of this crowded, fast-moving matrix.
Slide 12: Bacterial Cell Structure: Intermediary Metabolism – Why Core Pathways Reside in the Cytoplasm
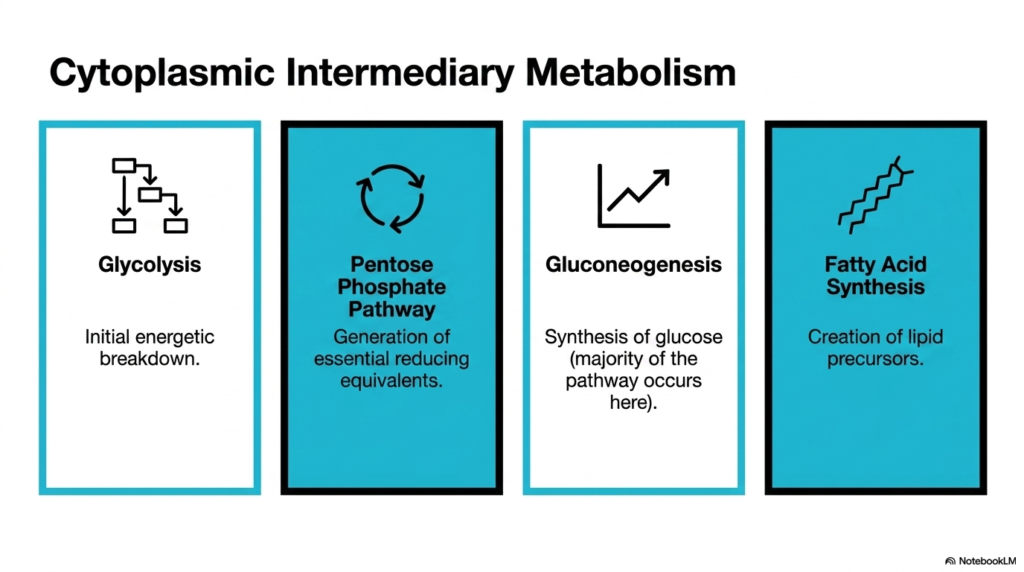
Picture the city’s main power grid, silently breaking down raw resources and routing electricity to millions of individual homes simultaneously. The core purpose of this slide deck is to isolate and explain the foundational metabolic pathways that power cellular life. Exploring Bacterial Cell Structure reveals that the most crucial energy-generating mechanisms reside directly in the cytoplasm. Medical students must master these intermediary networks, as they form the fundamental basis for understanding human nutrition, metabolic disease, and pharmacology.
The slide categorizes four critical pillars of cytoplasmic intermediary metabolism. It begins with glycolysis, the initial energetic breakdown of glucose, which provides immediate cellular fuel. Next, it highlights the pentose phosphate pathway, which generates essential reducing equivalents for anabolism. A key lesson in Bacterial Cell Structure is that these foundational energetic pathways do not require specialized organelles; they operate highly efficiently within the dense, crowded matrix of the main cellular compartment, directly powering immediate biological needs.
Furthermore, the slide details gluconeogenesis, noting that the vast majority of this glucose-synthesizing pathway occurs exactly here in the cytoplasm. Finally, it outlines fatty acid synthesis, the creation of vital lipid precursors required for membrane construction. By grouping these pathways together, Bacterial Cell Structure demonstrates a high level of functional integration. The cell places both the breakdown of sugars and the synthesis of crucial building blocks in the same physical space, enabling rapid, localized regulation in response to immediate energy demands.
For aspiring biochemists, memorizing these pathways is the cornerstone of cellular biology. The visual organization of this slide clearly defines the cytoplasm’s metabolic responsibilities. It proves that the internal matrix is a highly capable chemical factory, fully equipped to handle massive energy transformations. Understanding that these major pathways share the same physical real estate is crucial for grasping how the cell elegantly shifts between burning energy for fuel and storing it for future division.
Slide 13: Bacterial Cell Structure: Spatial Segregation – How Cells Optimize Thermodynamic Efficiency

Imagine trying to operate a massive blast furnace in the exact same room where engineers are attempting to assemble delicate microchips; chaos and failure would ensue. The core purpose of this slide deck is to explain how biology solves this problem through strict metabolic zoning. While Bacterial Cell Structure generally lacks organelles, understanding spatial segregation in eukaryotes highlights universal principles of thermodynamic efficiency. Students must recognize how life strategically separates opposing chemical reactions to prevent futile metabolic cycles and wasted energy.
This slide introduces a critical comparative concept: eukaryotic cells segregate opposing metabolic pathways into distinct compartments. The chart clearly outlines the division of labor between the cytoplasm and the mitochondria. While Bacterial Cell Structure relies on a single continuous space, eukaryotic evolution created boundaries. For instance, the synthesis of fatty acids occurs in the cytoplasm, whereas their degradation via beta-oxidation is strictly sequestered within the mitochondria. This prevents newly synthesized lipids from being broken down immediately, conserving large amounts of energy.
The chart further details how carbohydrates are handled, with glycolysis and most gluconeogenesis occurring in the cytoplasm, while complete oxidation via the TCA cycle occurs in the mitochondria. Energy production is similarly divided, contrasting cytoplasmic substrate-level phosphorylation with mitochondrial oxidative phosphorylation. Interestingly, when comparing this to Bacterial Cell Structure, one observes that prokaryotes must meticulously regulate these opposing pathways kinetically and chemically, rather than physically, since they lack the luxurious physical barriers provided by mitochondrial membranes.
This comparative slide is an essential learning tool for biology students. It perfectly illustrates the concept of thermodynamic optimization through spatial organization. By strictly defining where specific reactions occur, complex cells maintain massive chemical gradients and prevent destructive metabolic loops. It beautifully reinforces the idea that structure directly dictates function, showing that the physical location of an enzyme is just as important as its catalytic mechanism in the grand orchestra of cellular metabolism.
Slide 14: Bacterial Cell Structure: The Reaction Net – Why Metabolic Integration is Crucial
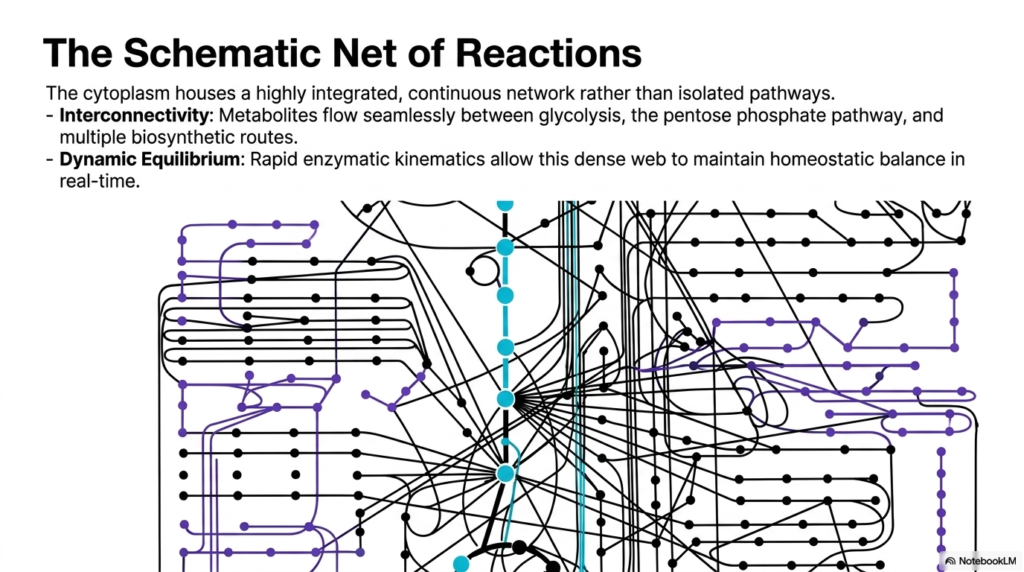
Think of a sprawling, intricately woven spider web where touching a single strand of silk sends immediate vibrations through the entire structure. The core purpose of this slide deck is to visualize this profound interconnectivity within the biochemical landscape. When mapping Bacterial Cell Structure, it becomes evident that metabolic pathways do not exist in isolation. Medical and biology students must adopt this integrated perspective to understand how pharmaceutical interventions can cause widespread, unintended side effects across seemingly unrelated cellular systems.
The slide presents a comprehensive schematic network of reactions, visually demonstrating that the cytoplasm houses a highly integrated, continuous network. It destroys the textbook illusion that glycolysis or the pentose phosphate pathway exists as independent, linear roads. In the reality of Bacterial Cell Structure, these pathways are deeply entangled. The visual features complex webs of black, purple, and blue nodes, demonstrating how intermediate molecules branch off and feed into multiple different biosynthetic routes simultaneously, creating a massively parallel processing system.
Two major points are highlighted: interconnectivity and dynamic equilibrium. Metabolites flow seamlessly between different metabolic and biosynthetic routes in response to cellular demand. Because of the incredible density of Bacterial Cell Structure, rapid enzymatic kinetics allow this dense web to maintain homeostatic balance in real time. If one pathway slows down, intermediates rapidly spill over into alternative interconnected pathways. This incredible dynamic shifting guarantees that the cell can rapidly adapt to fluctuating nutrient availability or sudden environmental stress without missing a beat.
For future clinicians, this slide is a vital lesson in systemic biology. It visually illustrates why biochemistry is deeply complex: altering a single enzyme’s function ripples through the entire cytoplasmic network. The representation of metabolism as a massive, interconnected net forces students to think holistically. It demonstrates that cellular survival is a delicately balanced act of chemical juggling, brilliantly managed by the continuous, rapid flow of thousands of specific molecules through a deeply intertwined metabolic highway.
Slide 15: Bacterial Cell Structure: The Biochemical Synthesis – How Crowding Creates Efficiency
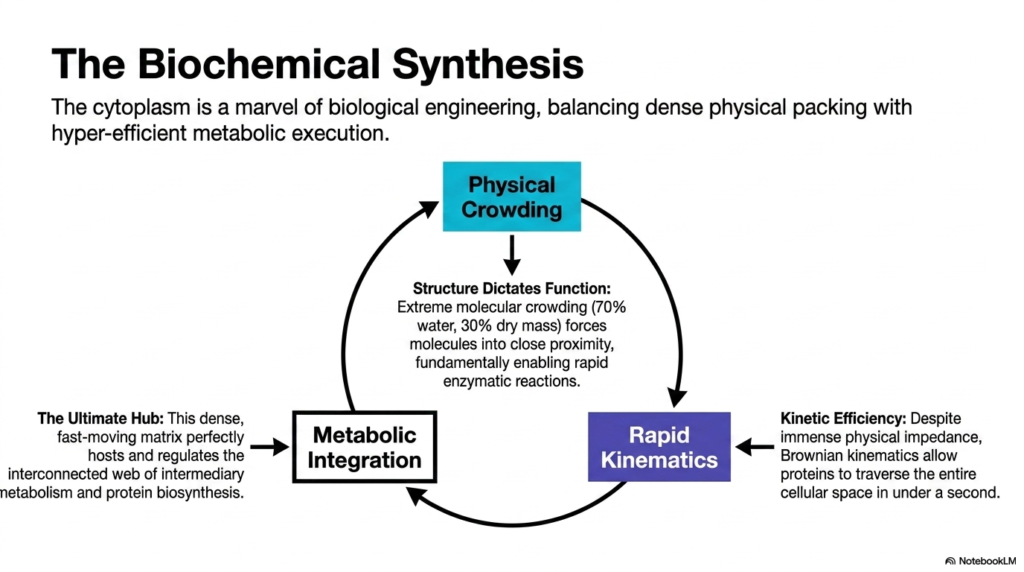
Consider a masterful symphony where every musician is crammed tightly onto a tiny stage, yet they play in such perfect synchrony that the resulting music is flawlessly executed. The core purpose of this slide deck is to synthesize all previous concepts into one unifying biophysical theory of cellular life. The study of Bacterial Cell Structure culminates in understanding how extreme physical limitations actually breed biochemical perfection. Students must grasp this final synthesis to truly appreciate the brilliant engineering of living organisms.
This concluding slide elegantly summarizes that the cytoplasm is a marvel of biological engineering. It visually balances physical crowding with hyper-efficient metabolic execution through a cyclic diagram. The core tenet of Bacterial Cell Structure is reiterated here: structure dictates function. The extreme molecular crowding, with 70% water and 30% dry mass, fundamentally forces molecules into close proximity. This tight packing is not a hindrance but rather the driving force that enables rapid, highly efficient enzymatic reactions.
The cycle progresses from physical crowding to rapid kinematics. Despite immense physical impedance, Brownian kinematics allow proteins to traverse the entire cellular space in under a second. This incredible kinetic efficiency directly leads to the final step: metabolic integration. The final lesson of Bacterial Cell Structure is that this dense, fast-moving matrix perfectly hosts and regulates the interconnected web of intermediary metabolism and protein biosynthesis. The cytoplasm acts as the ultimate hub, seamlessly combining physics and chemistry to create life.
For the biochemistry student, this final slide ties the entire presentation together perfectly. It proves that the chaotic, crowded nature of the cellular interior is a highly selected evolutionary advantage. The tight spaces guarantee that molecules collide, react, and metabolize with astonishing speed. By understanding this biochemical synthesis, learners walk away with a profound appreciation for the microscopic world, realizing that the cellular matrix is the most efficient, dynamic, and integrated chemical factory in the known universe.
Please read our Content Disclaimer Statement.
Check out our social media channels: